
Abstract
Chronic Obstructive Pulmonary Disease (COPD) is a progressive lung disease largely caused by cigarette smoking (CS) and is characterized by lung inflammation and airflow limitation that is not fully reversible. Approximately 50% of people with COPD die of a cardiovascular comorbidity and current pharmacological strategies provide little benefit. Therefore, drugs that target the lung and the cardiovascular system concurrently may be an advantageous therapeutic strategy. The aim of this study was to see whether losartan, an angiotensin-II AT1a receptor antagonist widely used to treat hypertension associated with cardiovascular disease, protects against CS-induced lung inflammation in mice. Male BALB/c mice were exposed to CS for 8 weeks and treated with either losartan (30 mg/kg) or vehicle daily. Mice were euthanized and bronchoalveolar lavage fluid (BALF) inflammation, and whole lung cytokine, chemokine and protease mRNA expression assessed. CS caused significant increases in BALF total cells, macrophages, neutrophils and whole lung IL-6, TNF-α, CXCL-1, IL-17A and MMP12 mRNA expression compared to sham-exposed mice. However, losartan only reduced CS-induced increases in IL-6 mRNA expression. Angiotensin-II receptor expression was reduced in lung tissue from CS-exposed mice. In conclusion, losartan did not inhibit CS-induced BALF cellularity despite reducing whole lung IL-6 mRNA and Ang-II receptor expression.
Similar content being viewed by others


Introduction
Chronic Obstructive Pulmonary Disease (COPD) is a devastating lung disorder most commonly caused by repeated inhalation of cigarette smoke (CS)1. The diseases encompasses emphysema, chronic obstructive bronchiolitis, and mucus-plugging of the airways1,2. There is also increased lung inflammation consisting of macrophages, neutrophils, and lymphocytes3. It is recognised that macrophages and neutrophils are major drivers of the pathology observed in COPD. Cigarette smoke induces the release of inflammatory factors such as tumour necrosis factor (TNF)-α, interleukin (IL)-6, C-C Motif Chemokine Ligand (CCL) 2, and reactive oxygen species (ROS) from alveolar macrophages2,3. Similarly, the number of neutrophils in the bronchoalveolar lavage fluid (BALF) and lung tissue of COPD patients in also increased2,3. These neutrophils secrete multiple proteases in the lungs of patients which then destroy the lung tissue leading to emphysema3,4.
In addition to lung inflammation and pathology, patients often develop other chronic medical conditions (called comorbidities) including cardiovascular disease which further impact the patient’s health and survival5,6,7. It is well recognised that the majority of the burden and healthcare cost in COPD is due to treating the various comorbidities which affect disease morbidity and mortality8. There is evidence that up to 50% of all COPD deaths are because of a cardiovascular comorbidity. In addition, prospective clinical trials have not identified a pharmaceutic that can decrease the risk of death in COPD7,9,10,11,12. There are studies showing that cardiovascular disease accounts for a significant proportion of first and second hospitalizations (42% and 44%, respectively) in COPD13 and that arrhythmias were associated with significant in-hospital mortality of COPD patients14,15.
Current COPD therapy is relatively ineffective16,17 and since a significant number of COPD patients will die of a cardiovascular comorbidity, drugs that target the lung and the cardiovascular system concurrently may be an advantageous therapeutic strategy. Epidemiological studies have suggested that angiotensin converting enzyme (ACE) inhibitors which are used to treat hypertension and cardiac failure (common comorbidities of COPD), may benefit COPD, by reducing mortality and exacerbations18,19,20,21. In addition, COPD patients on angiotensin II receptor blockers (ARBs) had reduced rates of pneumonia and mortality compared to COPD patients on ACE inhibitors22. The beneficial actions of ACE inhibitors in COPD may go beyond them attenuating pulmonary hypertension because angiotensin II (ang II) has pro-inflammatory effects23. In fact, ARBs reduce hyperinflation in subjects with COPD24 and attenuate CS-induced lung injury in mice25. Moreover, ACE inhibitors and ARBs have been shown to slow the progression of percent emphysema, especially among former smokers26. Randomized clinical trials of ARBs are underway to evaluate their effect on the progression of emphysema in COPD (https://clinicaltrials.gov/ct2/show/NCT02696564).
In this study, we examined whether losartan, an angiotensin-II AT1 receptor antagonist widely used to treat hypertension associated with cardiovascular disease, would significantly reduce lung inflammation in mice chronically exposed to CS. Identification of drugs that target the lung and the cardiovascular system concurrently may be advantageous for the treatment of COPD and comorbid cardiovascular disease.
Methodology
Mice
Specific pathogen-free male BALB/c mice (7 weeks of age) were obtained from the Animal Resource Centre Pty. Ltd. (Perth, Australia). Mice were housed in clean micro-isolator cages at 21 °C on a 12-hour day/night cycle with access to as much food or water as desired. After an acclimatisation period of four days, mice were weighed and organised into groups to assure the same average starting weight in each group. CS-exposed mice had access to as much food or water as desired. Mice were individually weighed 5 days per week (Monday to Friday). All experiments were approved by the Animal Ethics Committee of RMIT University (Ethics ID1521) and performed in accordance with the guidelines of animal experimentation established by the National Health and Medical Research Council of Australia.
Losartan treatment
Mice were treated with either losartan (Losartan Potassium, Sigma-Aldrich Co., Australia) or vehicle (water). Losartan Potassium was dissolved in the drinking water and mice had ad libitum access (0.25 g/L to correspond to a dose of 30 mg/kg). Losartan was prepared in fresh water daily for the duration of the protocol.
Cigarette smoke exposure
Mice were placed in an 18-L perspex chamber (The Plastic Man, Huntingdale, Victoria, Australia) in a class II biosafety cabinet (AES Environmental Pty Ltd, Melbourne, Victoria, Australia) and exposed to CS generated from three cigarettes (Winfield Red, ≤16 mg tar, ≤15 mg carbon monoxide, ≤1.2 mg nicotine; Philip Morris, Moorabbin, Australia) spaced evenly over 1 hour, and carried out three times per day (08:00, 11:00, and 14:00 h) for five days a week for 8 weeks as previously published27. Cigarette smoke was generated in 50-ml tidal volumes over 10 s as previously published27. The mean total suspended particulate mass concentration in the chamber containing CS was ~420 mg m−3; this level of CS exposure causes increases in blood carboxyhemoglobin comparable to that observed in human smokers27,28. Sham-exposed mice were handled the same way but were exposed to room air. At the end of the protocol, mice were euthanised with an overdose of anaesthetic (Lethabarb; 60 mg/kg; Virbac Pty. Ltd., Australia) via intraperitoneal injection (ip), and the analyses described below performed.
Bronchoalveolar lavage and tissue collection
Lungs were lavaged in situ with a 400 µl aliquot of PBS, followed by three 300 µl aliquots as previously described27,29. Approximately 1 ml of bronchoalvealor avage fluid (BALF) was recovered per mouse. The total number of viable cells in the BALF was determined, cytospins were prepared using 50 μl of BALF (~5 × 104 cells). Cytospin cells (500 per slide) were fixed in Kwik-Diff Reagent 1 (Thermo-Fisher, Massachusetts, USA), and then stained with Hemacolor® Rapid Solution 2 and 3 (Merck Millipore, Massachusetts, USA), and discerned into macrophages, neutrophils and lymphocytes by standard morphological criteria. The remaining BALF was centrifuged at 660 g for 5 min at 4 °C to collect the supernatant and stored at −80 °C until required. 5 mL of chilled PBS was used to perform a right ventricular perfusion of the heart to clear the lungs of blood. Lungs were then quickly excised, weighed, snap-frozen in liquid nitrogen, and stored at −80 °C until required. The liver, kidneys, retroperitoneal fat, and testicular fat were rapidly dissected, weighed, snap-frozen in liquid nitrogen and stored at −80 °C until required.
RNA extraction, cDNA synthesis and quantitative real time PCR
The whole lung from each mouse was crushed into a powder using a mortar and pestle with liquid nitrogen to avoid thawing. Two small scoops of tissue powder (~15 mg) were then transferred into a 1.7 mL Eppendorf (Germany) tube containing 600 μL RLT Lysis buffer (RNeasy® Mini Kit 250, Qiagen, Germany) supplemented with a 1:100 dilution of β-Mercaptoethanol (1000 × 2-Mercaptoethanol, Life Technologies Co., USA). The tissue was homogenised by passing it through a 21 g needle (Terumo Co., Japan) and 1 mL syringe (Livingstone International Pty. Ltd., Australia) 5–10 times. The Eppendorf tube was then centrifuged at 16,000 g for 3 minutes on a Sigma Laborzentrifugen 1–14 centrifuge (John Morris Scientific Pty. Ltd., Australia) and supernatant was collected. The RNA was then purified with an RNeasy Mini Kit 250 (Qiagen, Germany) and the concentration and purity of the extracted RNA determined with a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific Inc., USA). Single strand cDNA was synthesised by reverse transcription using the High Capacity RNA-to-cDNA Kit (Applied Biosystems, USA) and 1 μg of RNA used for cDNA synthesis.
Quantitative real time polymerase chain reaction (RT-qPCR) was performed using mouse-specific TaqMan Gene Expression Assays (Table 1, Applied Biosystems, Australia), with Taqman Fast Advanced Master Mix (Thermo Fisher Scientific Inc., Australia) on a QuantStudio 7 Flex Real-Time PCR System (Applied Biosystems, Australia). All samples were assayed in triplicate, and the assay was run for 40 cycles. All samples were standardised to GAPDH (housekeeping gene) and fold-change was determined in comparison to the sham vehicle group. Results were analysed via the 2−ΔΔCT method27,29,30,31.
Statistical analyses
Data are expressed as mean ± standard error of the mean (SEM) and n denotes the number of mice used in each group. Statistical differences were decided by two-way ANOVA followed by Sidak’s post-hoc test for multiple comparisons. GraphPad Prism for Windows (Version 8.0.1, GraphPad Sofware, Inc., USA) was used for all analyses and P < 0.05 was used to indicate statistical significance.
Results
Effect of losartan on CS-induced BALF inflammation and lung weight
Cigarette smoke exposure significantly increased the number of BALF total cells, macrophages and neutrophils (P < 0.05) (Fig. 1A–C) but did not affect lymphocyte numbers (P > 0.05) (Fig. 1D). However, losartan had no effect on CS-induced BALF cellularity (P > 0.05). In addition, CS significantly increased the lung weight compared to sham-exposed vehicle mice (P < 0.05) and losartan did not affect this response (P > 0.05) (Fig. 1E).

Effect of cigarette smoke exposure and losartan treatment on inflammatory cell numbers in bronchoalveolar lavage fluid (BALF) and lung weight. Number of total cells (A), macrophages (B), neutrophils (C) and lymphocytes (D) in BALF, and lung weight (E). Mice were exposed to cigarette smoke (CS) or room air (sham) for 8 weeks and treated with losartan (30 mg/kg) or vehicle (saline) administered in the drinking water. Data are expressed as mean ± SEM (n = 9–10 mice per group) and analysed by two-way ANOVA with Sidak’s post hoc test for multiple comparisons. *P < 0.05 compared to sham (vehicle).
Effect of losartan on lung tissue gene expression
To further characterise the inflammatory response to CS and losartan treatment, the mRNA expression of proinflammatory cytokines (IL-6, TNF-α), chemokines (C-X-C Motif Chemokine Ligand [CXCL]-1, C-C Motif Chemokine Ligand 2 [CCL2], IL-17A) and proteases (Matrix Metalloproteinase 12 [MMP12]) was measured in whole lung tissue.
Cigarette smoke exposure caused a significant increase in the mRNA expression of the proinflammatory cytokines, IL-6 and TNF-α, in the lung of CS-exposed mice when compared to sham vehicle mice (P < 0.05) (Fig. 2A,B). Losartan caused a significant decrease in CS-induced increases in IL-6 mRNA expression (P < 0.05) but was without effect on the increased TNF-α mRNA expression (P > 0.05) (Fig. 2A,B).

Effect of cigarette smoke exposure and losartan treatment on inflammatory cytokine and chemokine mRNA expression in whole lung tissue. Gene expression of interleukin-6 (A), tumor necrosis factor-α (B), chemokine ligand 1 (C), interleukin-17A (D), matrix metalloproteinase 12 (E) and chemokine ligand 2 (F) in lung tissue. Fold change was determined in comparison to sham (vehicle), after standardising samples to GAPDH (housekeeping gene). Mice were exposed to cigarette smoke (CS) or room air (sham) for 8 weeks and treated with losartan (30 mg/kg) or vehicle (saline) administered in the drinking water. Data are expressed as mean ± SEM (n = 4–10 mice per group) and analysed by two-way ANOVA with Sidak’s post hoc test for multiple comparisons. *P < 0.05 compared to sham (vehicle), #P < 0.05 compared to smoke (vehicle).
Cigarette smoke exposure caused a significant increase in CXCL-1, IL-17A and MMP12 mRNA expression compared to sham + vehicle mice (P < 0.05) (Fig. 2C–E) but losartan did not affect these increases in mRNA expression (P > 0.05) (Fig. 2C–E). Cigarette smoke did not cause a statistically significant increase in CCL2 mRNA expression although it was a 5-fold increase compared to sham-exposed vehicle mice (P > 0.05) (Fig. 2F). A significant increase in CCL2 expression was observed in CS-exposed losartan-treated mice compared to the sham + vehicle mice (P < 0.05) (Fig. 2F).
The mRNA expression of components of the renin angiotensin system (RAS), including angiotensinogen and the two angiotensin-II receptors, Agtr1a and Atr2, were also measured to determine the RAS-specific effects of losartan. Cigarette smoke exposure and losartan had no effect on mRNA expression of angiotensinogen (P > 0.05) (Fig. 3A). However, CS exposure caused a significant reduction in the expression of both Agtr1a and Agtr2 compared to sham + vehicle mice (P < 0.05) (Fig. 3B,C); however, losartan was without effect on these responses (P > 0.05) (Fig. 3B,C).

Effect of cigarette smoke exposure and losartan treatment on mRNA expression of components of the Renin-Angiotensin System in whole lung tissue. Gene expression of angiotensinogen (A), angiotensin-II AT1a (Agtr1a) receptor (B), and angiotensin-II AT2 (Agtr2) receptor. (C) Relative expression was determined in comparison to sham (vehicle), after standardising samples to GAPDH (housekeeping gene). Mice were exposed to cigarette smoke (CS) or room air (sham) for 8 weeks and treated with losartan (30 mg/kg) or vehicle (saline) administered in the drinking water. Data are expressed as mean ± SEM (n = 9–10 mice per group) and analysed by two-way ANOVA with Sidak’s post hoc test for multiple comparisons. *P < 0.05 compared to sham (vehicle).
Effect of cigarette smoke exposure and losartan treatment on body, kidney, liver and white adipose tissue weights
To understand the effect of both CS and losartan on body weight, mice were monitored for the duration of the protocol. The body weight of CS-exposed mice significantly reduced compared to sham-exposed vehicle mice (P < 0.05), but losartan did not affect this response (P > 0.05) (Fig. 4A).

Effect of cigarette smoke exposure and losartan treatment on body, organ and white adipose tissue weights. Progressive body weight of mice across the experimental period (A) and liver (B), kidney (C), testicular fat (D) and retroperitoneal fat (E) weights at the end of the 8-week cigarette smoke exposure protocol. Mice were exposed to cigarette smoke (CS) or room air (sham) for 8 weeks and treated with losartan (30 mg/kg) or vehicle (saline) administered in the drinking water. Data are expressed as mean ± SEM (n = 9–10 mice per group) and analysed by two-way ANOVA with Sidak’s post hoc test for multiple comparisons. *P < 0.05 compared to sham (vehicle).
Various organs (liver, kidney) and white adipose tissue (testicular and retroperitoneal fat) were weighed to further explore the effect of CS exposure and losartan treatment on body weight. Liver mass was significantly decreased in the CS-exposed losartan-treated group when compared to the sham vehicle group (P < 0.05) (Fig. 4B). The kidneys of CS-exposed mice were significantly heavier than those in the sham-exposed vehicle group (P < 0.05) (Fig. 4C). However, losartan did not affect this change in either the sham or CS groups (P > 0.05) (Fig. 4C). CS-exposed mice had significantly reduced testicular and retroperitoneal fat (P < 0.05), independent of losartan treatment (P > 0.05) (Fig. 4D,E). However, losartan-treated sham mice had significantly increased testicular and retroperitoneal fat compared to sham vehicle mice (P > 0.05) (Fig. 4D,E).
Average food and water consumption
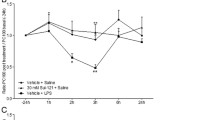
To investigate the effect of CS exposure and losartan on appetite, food and water intake was measured in the CS groups daily throughout the 8 weeks. There were no changes in the food consumed in any treatment group (P > 0.05) (Fig. 5A). Water consumption was measured daily to determine the effects of CS and losartan treatment on water intake. Vehicle-treated mice exposed to CS drank significantly less water when compared to sham-exposed vehicle mice (P < 0.05) (Fig. 5B).

Average food and water consumption during cigarette smoke exposure and losartan treatment. Average food consumed (A), and average water consumed (B) per mouse per day. Mice were exposed to cigarette smoke (CS) or room air (sham) for 8 weeks and treated with losartan (30 mg/kg) or vehicle (saline) administered in the drinking water. Sham groups were pair fed, receiving the same amount of food that the relevant smoke group ate in the previous 24 hours. Food intake of the CS-exposed groups was measured daily at approximately the same time to calculate the amount of food the sham groups would receive. Water intake was measured daily by calculating the difference in weight of water bottles in all cages across 24 hours. Data expressed as mean ± SEM (n = 9–10 mice per group) and analysed by two-way ANOVA with Sidak’s post hoc test for multiple comparisons. *P < 0.05 compared to sham (vehicle).
Discussion
COPD consists of not only a persistent lung inflammation that leads to reduced airflow but also systemic inflammation that leads to comorbidities including cardiovascular disease. Studies have implicated the renin angiotensin system (RAS) in the development of pulmonary and extrapulmonary disorders in COPD thus highlighting the potential therapeutic benefit of RAS blockade in subjects with COPD32. Therefore, we examined whether the angiotensin-II AT1 receptor antagonist losartan would significantly reduce lung inflammation in mice chronically exposed to CS and thus be a promising medication for the treatment of COPD and comorbid cardiovascular disease. We found that losartan did not inhibit CS-induced BALF cellularity despite reducing whole lung IL-6 mRNA and angiotensin II receptor mRNA expression.
Consistent with our previous studies33,34, we found in this study that mice exposed to CS had a significant increase in total inflammatory cells in the BALF, which was largely attributed to increased macrophages and neutrophils. Human studies have also shown that COPD patients have increased macrophages in BALF, sputum, airways and lung parenchyma compared to healthy individuals2. To determine the cause of this inflammation, the mRNA expression of chemokines in the lung tissue were explored. CXCL1, involved in neutrophil recruitment35, was markedly elevated in CS-exposed mice. IL-17A, a neutrophil chemotactic factor, has been shown to be elevated in the small airway epithelium in COPD subjects compared to non-COPD smokers and non-smokers36. In this study, IL-17A mRNA expression was markedly elevated in lung tissue from CS-exposed mice, which is in accord with our previous work demonstrating a pathogenic role of IL-17A in COPD37,38. Wang et al. have shown that CCL2 levels are elevated in AECOPD patients when compared to healthy individuals39. We observed a 5-fold increase in whole lung CCL2 expression in CS-exposed mice, though this was not statistically significant. IL-6 and TNF-α are two pro-inflammatory cytokines that are elevated in lungs of subjects with COPD40. In our chronic CS-exposure model, these cytokines had significantly increased mRNA expression in CS-exposed mice compared to sham vehicle mice. Moreover, the whole lung weighed significantly more in the CS-exposed mice when compared to sham vehicle mice, most likely due to the increased inflammatory response causing oedema.
It is well known that proteases break down lung tissue to cause emphysema and hence play an important role in COPD41. Studies have shown increased concentrations of MMP-1 and MMP-9 in BALF of subjects with emphysema42. In addition, increases in MMP-9 activity in lung parenchyma has also been demonstrated in people with emphysema43. Normal smokers have more MMP-9 in their alveolar macrophages than normal subjects44, and those with COPD have even greater levels45. A pathogenic role of MMP-12 has also been demonstrated in COPD whereby emphysema caused by CS was abolished in MMP-12−/− mice46. Proteolytic enzymes such as MMP-2, MMP-9, MMP-12 are also released from alveolar macrophages as are cathepsin K, L, and S, which all cause destruction of the lung parenchyma2,41. The elevated levels of MMP-12 mRNA in our study was likely due to the accumulation of macrophages.
In addition to lung inflammation, mRNA expression of the various RAS components were investigated in the lung to determine the impact of CS. The two angiotensin-II receptors (Agtr1a and Agtr2) have been shown to be present on epithelial cells in the lung, with localisation of Agtr1a mainly on the parenchyma47. Podowski et al.25 demonstrated no statistically significant change in Agtr1a expression in lung tissue from CS-treated mice. However, CS exposure significantly decreased Agtr1a expression in our model. Similarly, Agtr2 mRNA levels were reduced in the lungs of CS-exposed mice.
A significant quantified decrease in Mac3+ staining in the lung tissue with losartan treatment has been demonstrated in a mouse model of COPD25, suggesting the amount of macrophages recruited to the lung had decreased. In contrast, losartan treatment in our model did not decrease BALF cellularity. CXCL1 increased following CS exposure with both vehicle and losartan treatment and there was no significant difference between the two groups. This is consistent with a study that found BALF levels of CXCL1 were not decreased by treatment with another RAS inhibitor, enalapril, in a model of lipopolysaccharide-induced lung injury48. Treatment with losartan in CS-exposed mice significantly increased CCL2 expression in the lung tissue in comparison to sham vehicle mice suggesting losartan may have been involved in the macrophage infiltration observed in our model. Interestingly, this result contrasts to the cell counts as losartan seemed to induce a slight decrease in the total number of macrophages in the BALF. The CS-induced increase in TNF-α mRNA expression in the lung tissue was unaffected by treatment with losartan; however, losartan induced a significant decrease in IL-6 expression suggesting that it may have had a small effect on lung inflammation. Finally, losartan had no effect on the weight of the lung in CS-exposed mice. As no reduction was observed in BALF cellular inflammation, chemokine, or TNF-α expression, the results of this study suggest losartan has little effect on lung inflammation in this preclinical model of COPD. It is possible that we did not see a decrease in CS-induced BALF inflammation with losartan because the receptor blockade was incomplete. However, this is unlikely given that although losartan did not reduce CS-induced lung inflammation in our study, it did increase peripheral fat accumulation (testicular fat and retroperitoneal fat), reduced CS-induced increases in lung IL-6 mRNA and rescued water intake. These observations give confidence that sufficient drug was being ingested to cause a physiological effect and that the absence of an effect on the inflammatory response was real. The dose of losartan used in our study was also based on the study by Podowski et al.25 who showed that inhibition of TGF-β signalling through angiotensin receptor blockade attenuated cigarette smoke-induced lung injury.
As losartan is an AT1 receptor antagonist, the expression of components of the RAS was measured in the lung tissue to determine the effect of CS and losartan on RAS expression. Angiotensinogen is a precursor of angiotensin-II, the main effector peptide in the system, which signals via its two receptors, AT1 and AT2. There is little known about the effect of CS and losartan on the mRNA expression of these components. Treatment with losartan had no effect on the reduced expression of the two angiotensin-II receptors observed in the CS-exposed mice. In parallel, Podowski et al.25 demonstrated no change in expression of Agtr1a following losartan treatment. The reduced expression of the angiotensin-II receptors in the lung following CS-exposure in our study may explain why losartan did not decrease CS-induced lung inflammation.
COPD is a multi-factorial disease with contributions from inflammation, oxidative stress, hormone signalling, as well as appetite6,49. It is well established that the nicotine component of CS suppresses appetite and causes loss of body weight in both human and animal studies50,51,52. As such, the sham mice were pair-fed in our model to control for changes in appetite and to investigate the contributions of other mechanisms in mediating body and organ weight. Regardless of pair-feeding, losartan did not affect food consumption in our model. The appetite suppression from the CS possibly overrides any differences that may have been observed in the sham groups in the absence of pair-feeding. The dose of losartan chosen for our study is within the range (3–30 mg/kg) used in a similar mouse model of COPD25. In addition, we trialed the 3 and 30 mg/kg doses in a pilot 4 day acute CS-exposure protocol where we found that 30 mg/kg losartan caused a small (but not statistical) decrease in total BALF cellularity in CS-exposed mice and that the CS-exposed mice treated with 30 mg/kg losartan had the largest mean percentage weight gain and water consumption compared to other CS-exposed mice (data not shown).
To explore the effect of CS and losartan treatment on overall body weight, the mice were weighed 5 days per week for the duration of the protocol. A steady increase in weight was observed in all groups; however, the CS-treated mice gained significantly less than the sham groups overall reflecting the results observed in the literature33. According to various studies, the weight loss observed from CS treatment is due to a greater capacity for energy expenditure and not just due to the reduced food consumption53,54. Losartan had no significant effect on bodyweights in either group, consistent with the literature25,55, although losartan-treated sham mice were 2.6% heavier than sham vehicle mice at the end of the 8 week protocol. This effect is most likely explained by the increased white adipose tissue weight (testicular and retroperitoneal fat) in the sham losartan-treated mice. This was somewhat unexpected studies in rats showed that losartan alone had no effect on baseline white adipose tissue weight, suggesting that it could be a species related phenomenon56,57. The CS-treated mice in our model demonstrated a significant decrease in white adipose tissue weight which is consistent with studies showing that Ang II infusion decreases retroperitoneal white adipose (RWAT) fat mass56. However, in that study losartan reversed the Ang II-induced decreases in RWAT fat mass which was not the case in our CS study, highlighting that differences may be a species-dependent observation. In addition, losartan significantly decreased liver weight in CS-exposed mice compared to sham vehicle, and CS-exposure significantly increased kidney weight in vehicle and losartan groups compared to sham vehicle. Body composition in COPD is somewhat complex and involves many factors including genetics, appetite, and exercise tolerance58. As such, patients demonstrate differential responses to CS and have been shown to both increase and decrease body weight58,59. In both human and animal studies, CS causes a preference towards foods high in refined sugar and fat which possibly explains the increased fat weight in some cases60,61. In contrast, BALB/c mice are genetically identical, and all have equal access to normal experimental chow, highlighting a possible limitation in our model.
An enzyme involved in the metabolism of losartan and other drugs, CYP2C9, has demonstrated between-patient differences in the drug pharmacokinetics and pharmacodynamics, which can result in inadequate therapeutic effect or toxicity62,63. Comparisons between Korean and Swedish populations demonstrated that CYP2C9 genotype, ethnicity, and CS are major determinants of enzymatic activity64. Higher enzymatic activity in smokers in each population was observed, with more significant changes in the Korean population64. In addition, CYP2C9 has been demonstrated to be involved in two pathways that trigger the metabolic activation of polycyclic aromatic hydrocarbons found in CS65,66,67. It is thus possible that CYP2C9 enzyme is required for the metabolism of components of CS in our model and is less effective at metabolising losartan. As such, further examination of the levels of losartan metabolites in the blood system is necessary to understand how losartan was metabolised in our model.
In conclusion, 8 weeks of CS exposure in mice caused significant lung inflammation as measured by increased BALF cellularity, increased whole lung mRNA expression for pro-inflammatory cytokines, chemokines and proteases and increases in lung weight. Cigarette smoke exposure also caused significant decreases in whole lung angiotensin II receptor mRNA expression and decreases in body weight, liver and adipose tissue weight. Losartan treatment did not inhibit CS-induced BALF cellularity despite reducing whole lung IL-6 mRNA and Ang-II receptor mRNA expression. In addition, losartan was without affect on CS-induced decreases in body weight, liver and white adipose tissue mass. Thus, it appears unlikely that losartan may be a viable therapeutic to treat CS-induced lung inflammation seen in patients with COPD.
Data availability
All data generated or analysed during this study are included in this published article.
References
Vogelmeier, C. F. et al. Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Lung Disease 2017 Report. GOLD Executive Summary. Am J Respir Crit Care Med 195, 557–582 (2017).
Barnes, P. J., Shapiro, S. D. & Pauwels, R. A. Chronic obstructive pulmonary disease: molecular and cellular mechanisms. Eur Respir J 22, 672–688 (2003).
Vlahos, R. & Bozinovski, S. Recent advances in pre-clinical mouse models of COPD. Clin Sci (Lond) 126, 253–265 (2014).
Di Stefano, A. et al. Upregulation of adhesion molecules in the bronchial mucosa of subjects with chronic obstructive bronchitis. Am J Respir Crit Care Med 149, 803–810 (1994).
Barnes, P. J. Chronic obstructive pulmonary disease: important advances. Lancet Respir Med 1, e7–8 (2013).
Bernardo, I., Bozinovski, S. & Vlahos, R. Targeting oxidant-dependent mechanisms for the treatment of COPD and its comorbidities. Pharmacol Ther 155, 60–79 (2015).
Brassington, K., Selemidis, S., Bozinovski, S. & Vlahos, R. New frontiers in the treatment of comorbid cardiovascular disease in chronic obstructive pulmonary disease. Clin Sci (Lond) 133, 885–904 (2019).
Barnes, P. J. & Celli, B. R. Systemic manifestations and comorbidities of COPD. Eur Respir J 33, 1165–1185 (2009).
Sin, D. D. & Man, S. F. Chronic obstructive pulmonary disease as a risk factor for cardiovascular morbidity and mortality. Proc Am Thorac Soc 2, 8–11 (2005).
Chatila, W. M., Thomashow, B. M., Minai, O. A., Criner, G. J. & Make, B. J. Comorbidities in chronic obstructive pulmonary disease. Proc Am Thorac Soc 5, 549–555 (2008).
Berger, J. S., Sanborn, T. A., Sherman, W. & Brown, D. L. Effect of chronic obstructive pulmonary disease on survival of patients with coronary heart disease having percutaneous coronary intervention. Am J Cardiol 94, 649–651 (2004).
Curkendall, S. M. et al. Cardiovascular disease in patients with chronic obstructive pulmonary disease, Saskatchewan Canada cardiovascular disease in COPD patients. Ann Epidemiol 16, 63–70 (2006).
Anthonisen, N. R. et al. Effects of smoking intervention and the use of an inhaled anticholinergic bronchodilator on the rate of decline of FEV1. The Lung Health Study. Jama 272, 1497–1505 (1994).
Hudson, L. D., Kurt, T. L., Petty, T. L. & Genton, E. Arrhythmias associated with acute respiratory failure in patients with chronic airway obstruction. Chest 63, 661–665 (1973).
Gulsvik, A., Hansteen, V. & Sivertssen, E. Cardiac arrhythmias in patients with serious pulmonary diseases. Scand J Respir Dis 59, 154–159 (1978).
Barnes, P. J. New anti-inflammatory targets for chronic obstructive pulmonary disease. Nat Rev Drug Discov 12, 543–559 (2013).
Barnes, P. J. Development of new drugs for COPD. Curr Med Chem 20, 1531–1540 (2013).
Mancini, G. B. et al. Reduction of morbidity and mortality by statins, angiotensin-converting enzyme inhibitors, and angiotensin receptor blockers in patients with chronic obstructive pulmonary disease. J Am Coll Cardiol 47, 2554–2560 (2006).
Soyseth, V., Brekke, P. H., Smith, P. & Omland, T. Statin use is associated with reduced mortality in COPD. Eur Respir J 29, 279–283 (2007).
Keddissi, J. I. et al. The use of statins and lung function in current and former smokers. Chest 132, 1764–1771 (2007).
Paulin, P. et al. Effect of angiotensin 2 receptor blockers on chronic obstructive lung disease mortality: A retrospective cohort study. Pulm Pharmacol Ther 44, 78–82 (2017).
Lai, C. C. et al. Comparative effects of angiotensin-converting enzyme inhibitors and angiotensin II receptor blockers on the risk of pneumonia and severe exacerbations in patients with COPD. Int J Chron Obstruct Pulmon Dis 13, 867–874 (2018).
Dandona, P., Dhindsa, S., Ghanim, H. & Chaudhuri, A. Angiotensin II and inflammation: the effect of angiotensin-converting enzyme inhibition and angiotensin II receptor blockade. J Hum Hypertens 21, 20–27 (2007).
Andreas, S. et al. Angiotensin II blockers in obstructive pulmonary disease: a randomised controlled trial. Eur Respir J 27, 972–979 (2006).
Podowski, M. et al. Angiotensin receptor blockade attenuates cigarette smoke-induced lung injury and rescues lung architecture in mice. J Clin Invest 122, 229–240 (2012).
Parikh, M. A. et al. Angiotensin-converting inhibitors and angiotensin II receptor blockers and longitudinal change in percent emphysema on computed tomography. The multi-ethnic study of atherosclerosis lung study. Ann Am Thorac Soc 14, 649–658 (2017).
Vlahos, R. et al. Differential protease, innate immunity, and NF-kappaB induction profiles during lung inflammation induced by subchronic cigarette smoke exposure in mice. Am J Physiol Lung Cell Mol Physiol 290, L931–945 (2006).
Wald, N., Idle, M. & Bailey, A. Carboxyhaemoglobin levels and inhaling habits in cigarette smokers. Thorax 33, 201–206 (1978).
Vlahos, R. et al. Neutralizing granulocyte/macrophage colony-stimulating factor inhibits cigarette smoke-induced lung inflammation. Am J Respir Crit Care Med 182, 34–40 (2010).
Austin, V., Ku, J. M., Miller, A. A. & Vlahos, R. Ischaemic stroke in mice induces lung inflammation but not acute lung injury. Sci Rep 9, 3622 (2019).
Yu, X. et al. Matrine reduces cigarette smoke-induced airway neutrophilic inflammation by enhancing neutrophil apoptosis. Clin Sci (Lond) 133, 551–564 (2019).
Vasileiadis, I. E., Goudis, C. A., Giannakopoulou, P. T. & Liu, T. Angiotensin converting enzyme inhibitors and angiotensin receptor blockers: a promising medication for chronic obstructive pulmonary disease? COPD 15, 148–156 (2018).
Hansen, M. J. et al. The lung inflammation and skeletal muscle wasting induced by subchronic cigarette smoke exposure are not altered by a high-fat diet in mice. PloS one 8, e80471 (2013).
Austin, V., Miller, A. & Vlahos, R. Prior cigarette smoke exposure does not affect acute post-stroke outcomes in mice. PLoS One 14, e0214246 (2019).
Sawant, K. V. et al. Chemokine CXCL1 mediated neutrophil recruitment: Role of glycosaminoglycan interactions. Sci Rep 6, 33123 (2016).
Eustace, A. et al. Identification of cells expressing IL-17A and IL-17F in the lungs of patients with COPD. Chest 139, 1089–1100 (2011).
Bozinovski, S. et al. Innate cellular sources of interleukin-17A regulate macrophage accumulation in cigarette- smoke-induced lung inflammation in mice. Clin Sci 129, 785–796 (2015).
Bozinovski, S. & Vlahos, R. Multifaceted Role for IL-17A in the Pathogenesis of Chronic Obstructive Pulmonary Disease. Am J Respir Crit Care Med 191, 1213–1214 (2015).
Wang, Y., Zheng, Y., Zhai, Y. L., Liu, F. Q. & Ding, N. Comparative analysis of MCP-1 and TF in elderly patients with acute exacerbations of COPD and its clinical significance. Eur Rev Med Pharmacol Sci 19, 215–219 (2015).
Barnes, P. J. Cellular and molecular mechanisms of chronic obstructive pulmonary disease. Clin Chest Med 35, 71–86 (2014).
Gualano, R. C., Vlahos, R. & Anderson, G. P. What is the contribution of respiratory viruses and lung proteases to airway remodelling in asthma and chronic obstructive pulmonary disease? Pulm Pharmacol Ther 19, 18–23 (2006).
Finlay, G. A. et al. Matrix metalloproteinase expression and production by alveolar macrophages in emphysema. Am J Respir Crit Care Med 156, 240–247 (1997).
Ohnishi, K., Takagi, M., Kurokawa, Y., Satomi, S. & Konttinen, Y. T. Matrix metalloproteinase-mediated extracellular matrix protein degradation in human pulmonary emphysema. Lab Invest 78, 1077–1087 (1998).
Lim, S. et al. Balance of matrix metalloprotease-9 and tissue inhibitor of metalloprotease-1 from alveolar macrophages in cigarette smokers. Regulation by interleukin-10. Am J Respir Crit Care Med 162, 1355–1360 (2000).
Russell, R. E. et al. Release and activity of matrix metalloproteinase-9 and tissue inhibitor of metalloproteinase-1 by alveolar macrophages from patients with chronic obstructive pulmonary disease. Am J Respir Cell Mol Biol 26, 602–609 (2002).
Hautamaki, R. D., Kobayashi, D. K., Senior, R. M. & Shapiro, S. D. Requirement for macrophage elastase for cigarette smoke-induced emphysema in mice. Science 277, 2002–2004 (1997).
Bullock, G. R. et al. Distribution of type-1 and type-2 angiotensin receptors in the normal human lung and in lungs from patients with chronic obstructive pulmonary disease. Histochem Cell Biol 115, 117–124 (2001).
Arndt, P. G. et al. Systemic inhibition of the angiotensin-converting enzyme limits lipopolysaccharide-induced lung neutrophil recruitment through both bradykinin and angiotensin II-regulated pathways. J Immunol 177, 7233–7241 (2006).
Passey, S. L. et al. Emerging therapies for the treatment of skeletal muscle wasting in chronic obstructive pulmonary disease. Pharmacol Ther 166, 56–70 (2016).
Wager-Srdar, S. A., Levine, A. S., Morley, J. E., Hoidal, J. R. & Niewoehner, D. E. Effects of cigarette smoke and nicotine on feeding and energy. Physiol Behav 32, 389–395 (1984).
Grunberg, N. E., Bowen, D. J. & Winders, S. E. Effects of nicotine on body weight and food consumption in female rats. Psychopharmacology (Berl) 90, 101–105 (1986).
Bellinger, L., Cepeda-Benito, A. & Wellman, P. J. Meal patterns in male rats during and after intermittent nicotine administration. Pharmacol Biochem Behav 74, 495–504 (2003).
Chen, H. et al. Cigarette smoke exposure reprograms the hypothalamic neuropeptide Y axis to promote weight loss. Am J Respir Crit Care Med 173, 1248–1254 (2006).
Chen, H. et al. Long-term cigarette smoke exposure increases uncoupling protein expression but reduces energy intake. Brain Res 1228, 81–88 (2008).
Smith, P. M., Hindmarch, C. C., Murphy, D. & Ferguson, A. V. AT1 receptor blockade alters nutritional and biometric development in obesity-resistant and obesity-prone rats submitted to a high fat diet. Front Psychol 5, 832 (2014).
Cabassi, A. et al. Sympathetic modulation by carvedilol and losartan reduces angiotensin II-mediated lipolysis in subcutaneous and visceral fat. J Clin Endocrinol Metab 90, 2888–2897 (2005).
Wang, T., Lian, G., Cai, X., Lin, Z. & Xie, L. Effect of prehypertensive losartan therapy on AT1R and ATRAP methylation of adipose tissue in the later life of highfatfed spontaneously hypertensive rats. Mol Med Rep 17, 1753–1761 (2018).
Schols, A. M., Broekhuizen, R., Weling-Scheepers, C. A. & Wouters, E. F. Body composition and mortality in chronic obstructive pulmonary disease. Am J Clin Nutr 82, 53–59 (2005).
Canoy, D. et al. Cigarette smoking and fat distribution in 21,828 British men and women: a population-based study. Obes Res 13, 1466–1475 (2005).
Marangon, K. et al. Diet, antioxidant status, and smoking habits in French men. Am J Clin Nutr 67, 231–239 (1998).
Chen, H. et al. Detrimental metabolic effects of combining long-term cigarette smoke exposure and high-fat diet in mice. Am J Physiol Endocrinol Metab 293, E1564–1571 (2007).
Ninomiya, H. et al. Genetic polymorphism of the CYP2C subfamily and excessive serum phenytoin concentration with central nervous system intoxication. Ther Drug Monit 22, 230–232 (2000).
Wadelius, M. Warfarin pharmacogenetics: it matters if you’re black or white. Blood 124, 2171 (2014).
Hatta, F. H. et al. Differences in CYP2C9 Genotype and Enzyme Activity Between Swedes and Koreans of Relevance for Personalized Medicine: Role of Ethnicity, Genotype, Smoking, Age, and Sex. OMICS 19, 346–353 (2015).
Tranah, G. J. et al. Epoxide hydrolase and CYP2C9 polymorphisms, cigarette smoking, and risk of colorectal carcinoma in the Nurses’ Health Study and the Physicians’ Health Study. Mol Carcinog 44, 21–30 (2005).
Brown, B., Avalos, J., Lee, C. & Doolittle, D. The effect of tobacco smoke, nicotine, and cotinine on the mutagenicity of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanol (NNAL). Mutat Res 494, 21–29 (2001).
Smith, C. A. & Harrison, D. J. Association between polymorphism in gene for microsomal epoxide hydrolase and susceptibility to emphysema. Lancet 350, 630–633 (1997).
Acknowledgements
This study was supported by the National Health and Medical Research Council of Australia [Project Grant 1084627].
Author information
Authors and Affiliations
Contributions
R.V. and S.L.P. conceived the study design. M.L.H., S.L.P. and R.V. designed the experiments. M.L.H., H.J.S. and S.L.P. acquired the data. M.L.H., H.J.S., S.L.P. and R.V. analysed and interpreted the data. M.L.H., H.J.S., S.L.P. and R.V. drafted the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hepworth, M.L., Passey, S.L., Seow, H.J. et al. Losartan does not inhibit cigarette smoke-induced lung inflammation in mice. Sci Rep 9, 15053 (2019). https://doi.org/10.1038/s41598-019-51504-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-51504-2
- Springer Nature Limited
This article is cited by
-
Ebselen prevents cigarette smoke-induced cognitive dysfunction in mice by preserving hippocampal synaptophysin expression
Journal of Neuroinflammation (2022)