
Abstract
Here, we describe part of a large-bodied macrophagous plesiosaur jaw from the lower Bajocian (Middle Jurassic) Passwang Formation near Arisdorf in the Basel-Land canton of Switzerland. The specimen preserves the posterior glenoid extremity of the right mandibular ramus comprising the surangular, angular, articular, and probably the prearticular. Notable character states include a transversely expanded surangular that incorporates a prominent medial ridge adjacent to the glenoid and a shallow dorsal fossa, together with a conspicuous trough extending across the lateral surfaces of both the surangular and angular; there is also a basally broad and medially deflected retroarticular process. Collectively, these features suggest affinities with Middle–Late Jurassic pliosaurids, as well as some coeval rhomaleosaurids. Based on such comparisons, we estimate that the Passwang Formation mandible was approximately 1.5 m long when complete, and thus approaches the size range of the largest-known pliosaurids. This discovery supplements the sparse record of diagnostic pliosaurid-like remains from Switzerland and contributes to the currently incomplete knowledge of pre-Callovian Middle Jurassic plesiosaurs globally.
Similar content being viewed by others


Introduction
Although both geographically and stratigraphically ubiquitous, the fossil record of plesiosaurs currently has very incomplete representation from Middle Jurassic strata of pre-Callovian (Aalenian–Bajocian) age (e.g. Buchy 2004; Gasparini 1997; Gasparini and Fernández 2006; Godefroit 1994; Kear 2012; Sachs and Hornung 2016; Vincent et al. 2007, 2013b). To date, only two taxa have been recognised from this interval: the rhomaleosaurid Maresaurus coccai (Gasparini 1997), from the early Bajocian of Argentina, and the probable pliosaurid Simolestes keileni (Godefroit 1994) from the late Bajocian of France. Consequently, the understanding of early Middle Jurassic plesiosaur diversity and distribution is, at present, extremely limited (e.g. Bardet et al. 2014; Kear et al. 2018; Tutin and Butler 2017).
Here, we briefly report the discovery of an incomplete mandible representing a large-skulled pliosaurid-like plesiosaur from the lower Bajocian section of the Passwang Formation in Switzerland. Published accounts of Swiss plesiosaur fossils are, in general, rare (e.g. Geister 1998; Peyer 1940; von Meyer 1856; Wild 1968); thus, although fragmentary, this new specimen provides an important addition to the regional record and also contributes towards filling the gap in knowledge about early Middle Jurassic plesiosaurs globally.
Abbreviations are as follows: CMN Canadian Museum of Nature, Ottawa, Canada; MNHNL Musée national d’Histoire naturelle de Luxembourg; and PIMUZ Paläontologisches Institut und Museum der Universität Zürich, Zurich, Switzerland.
Geological setting
The incomplete plesiosaur mandible described herein (PIMUZ A/III0521) was collected by the fossil enthusiast Dr Hans Holenweg (Muttenz, Switzerland), on the north-western slope of the Eileten (625 m), a prominent hill to the southwest of Arisdorf in the Basel-Land canton of north-western Switzerland (Fig. 1). Shallow marine deposits of Aalenian–Bathonian age occur as localised outcrops at this locality (e.g. Jordan et al. 2008). They overlie the Toarcian–Aalenian Opalinus-Ton on the northern and southern slopes (Wetzel and Allia 2003; Hostettler et al. 2017) and principally comprise the Aalenian–Bajocian Passwang Formation (Burkhalter 1996), which is best exposed over a large area on the western and northern slopes of the Eileten. The Passwang Formation consists of iron oolites, carbonatic sandstones, and claystones and is stratigraphically succeeded by the Hauptrogenstein (Gonzalez and Wetzel 1996), a probable Bajocian–Bathonian unit famous for crinoidal limestones (inter-bedding with minor marls and oolites) that produce well-preserved marine invertebrate fossils.

Locality map of the Eileten and area around Arisdorf in Switzerland. Extent of subsurface and outcrop geological units is based on data from Swisstopo (www.swisstopo.admin.ch)
Systematic palaeontology
Sauropterygia Owen 1860
Plesiosauria de Blainville 1835
?Pliosauridae indet.
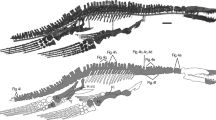
Material: PIMUZ A/III0521, the glenoid extremity of a right mandibular ramus (Fig. 2).

Pliosaurid-like plesiosaur mandible section (PIMUZ A/III0521) recovered from the Passwang Formation near Arisdorf (Switzerland) in (a) lateral, b dorsal, c medial, d ventral, e anterior, and f posterior views. an angular, apr anterior process of angular, art articular, gf mandibular glenoid fossa, lt lateral trough, pf posterior facet of retroarticular process, ps possible suture, rap retroarticular process, sa surangular, sf surangular fossa
Stratigraphic and geographic provenance: Emileia sauzei Zone (early Bajocian) of the upper Passwang Formation exposed on the north-western slope of the Eileten, southeast of Arisdorf, Canton of Basel-Land, north-western Switzerland.
Description and comparisons: PIMUZ A/III0521 includes the surangular, angular, articular, and probably part of the prearticular (Fig. 2). The mandible was broken into several pieces with fracturing and substantial loss of bone having occurred along the damaged surfaces. Subsequent restoration using coloured plaster has obscured sutures, but these appear to have been well-ossified in life. The surangular can be anatomically delimited by its transverse expansion anterior to the glenoid fossa; this incorporates a prominent longitudinal ridge on the medial surface (Fig. 2b). There is a shallow dorsal surangular fossa bounded posteromedially by a rounded ledge (Fig. 2b). Both of these features are characteristics of pliosaurids, such as Marmornectes candrewi Ketchum & Benson, 2011a (Ketchum and Benson 2011a, p. 113, text-fig. 3), Peloneustes philarchus (Seeley 1869) (see Ketchum and Benson 2011b, p. 657, pl. 3, figs. 7–10), Pliosaurus kevani Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum & Forrest, 2013 (Benson et al. 2013, p. 22, fig. 20B, D), and Cryonectes neustriacus Vincent, Bardet & Mattioli (2013) (Vincent et al. 2013a, p. 474, fig. 3). The surangular, as preserved, tapers anteriorly and has a shallowly concave dorsal margin in lateral and medial views (Fig. 2a, c). The dorsolateral edge is damaged, but clearly protruded laterally to form a shelf similar to that illustrated in P. kevani (Benson et al. 2013, p. 22, fig. 20D). A posteroventrally deflected keel on the anterolateral margin of the glenoid fossa (Fig. 2a) may demarcate the surangular–angular contact, as in P. kevani (Benson et al. 2013, p. 22, fig. 20D), Arminisaurus schuberti Sachs and Kear 2018 (Sachs and Kear 2018, p. 490, fig. 2A), and the rhomaleosaurid Rhomaleosaurus thorntoni Andrews 1922 (see Smith and Benson 2014, ppl. 8, fig. 4); this suture is also potentially traceable along the posterolateral process of the glenoid fossa (Fig. 2a).
The lateral surface of the mandible is indented by a prominent trough that extends across both the surangular and angular (Fig. 2a). A similar trough has been reported in pliosaurids, including Stratesaurus taylori Benson, Evans & Druckenmiller, 2012 (Benson et al. 2015, p. 11, fig. 9C) and Hauffiosaurus tomistomimus Benson, Ketchum, Noè & Gómez-Pérez (Benson et al. 2011, p. 552, text-fig. 3), as well as in the rhomaleosaurids Meyerasaurus victor (Fraas 1910) (Smith and Vincent 2010, p. 1054, text-fig. 3C) and Thaumatodracon wiedenrothi Smith & Araújo, 2017 (Smith and Araújo 2017, p. 105, text-fig. 7C). Benson and Druckenmiller (2014) also considered the presence of a lateral mandibular trough to be a synapomorphy for Cretaceous leptocleidians based on its occurrence in Brancasaurus brancai Wegner 1914 (Sachs et al. 2016, p. 16, fig. 4), Nichollssaura borealis (Druckenmiller and Russell 2008) (Druckenmiller and Russell 2008, p. 7, text-fig. 5C), and Plesiopleurodon wellesi Carpenter 1996 (Benson and Druckenmiller 2014, p. 11, fig. 4).
The articulated surangular and angular have a medially constricted ‘hour-glass-like’ cross section in anterior view (Fig. 2e). The ventral profile of the angular is transversely rounded, and the bone gradually widens posteriorly to reach its maximum width beneath the glenoid fossa (Fig. 2d). The lateral and medial sides of the angular are shallowly convex. The tapering anterior process of the articular is traceable on the medial side of the mandible and would have buttressed the prearticular, which appears to be present based on this anatomical association (see Benson et al. 2013, p. 18, fig. 16C, Ketchum and Benson 2011a, p. 114, text-fig. 4G, Ketchum and Benson 2011b, p. 658, text-fig. 12C).
The mandibular glenoid fossa of PIMUZ A/III0521 is 83 mm in maximum width, but only broken remnants of the glenoid processes are preserved (Fig. 2b). Despite this, the glenoid fossa was clearly medially inclined. The adjacent retroarticular process is 64 mm long and integrates both the articular and part of the angular (Fig. 2c). Its longitudinal axis is deflected medially and proportionately approximates the glenoid fossa in length, which is a trait shared with pliosaurids including Liopleurodon ferox Sauvage 1873 (see Andrews 1913, pl. II, fig. 1), some species of Pliosaurus (see Knutsen 2012, p. 266, fig. 6), the Cretaceous brachauchenines Brachauchenius lucasi Williston 1903 (see Albright et al. 2007, p. 34, fig. 3B) and Kronosaurus queenslandicus Longman 1924 (see Holland 2019, p. 7, fig. 5), and rhomaleosaurids, such as Rhomaleosaurus cramptoni (Carte and Bailey 1863) (see Smith and Dyke 2008, p. 976, fig. 1b). In dorsal view, the retroarticular process has a broad base and triangular profile, with convex lateral and medial surfaces. A dorsal flange also extends from the posterolateral edge of the glenoid fossa to the posterior tip of the retroarticular process, which is indented for the insertion of jaw musculature (Fig. 2f).
Affinities and implications for body-size
We tentatively interpret PIMUZ A/III0521 as most likely representing a pliosaurid based on its transversely expanded surangular with a prominent longitudinal ridge on its medial surface anterior to the glenoid, and a shallow dorsal fossa—character states that are apparently unique to this group (Benson and Druckenmiller 2014, Appendix 2, character 127). However, our first-hand comparative inspection of other Middle Jurassic plesiosaur mandibles reveals a similar condition in the rhomaleosaurid Borealonectes russelli Sato and Wu 2008 (CMN 40729, SS pers. obs., Sato and Wu 2008, p. 308, fig. 3A), which possesses both a transversely expanded surangular and a dorsal fossa; the dorsal edge of the surangular is otherwise not preserved in Simolestes keileni (MNHNL BU 159, SS pers. obs., see also Godefroit 1994, pl. 1C). In addition, PIMUZ A/III0521 shares the development of a lateral trough on the surangular and angular, as well as a broad medially deflected retroarticular process with S. keileni (MNHNL BU 159, SS pers. obs) and Maresaurus coccai (Gasparini 1997, p. 137, text-figs. 1A, C), demonstrating close similarities between these stratigraphically (and in the case of S. keileni also geographically) coeval taxa. Nevertheless, the mandibular proportions of PIMUZ A/III0521 indicate a larger individual. For example, the preserved maximum length of PIMUZ A/III0521 is 496 mm, with a corresponding maximum height of 127 mm in the coronoid region. By contrast, the incomplete, but much more intact mandibular ramus of S. keileni measures 1.33 m in maximum preserved length (SS pers. obs.), and is consistent with the extrapolated mandibular dimensions of M. coccai, which we estimate at around 1.1 m in maximum length based on the reconstruction of Gasparini (1997, p. 137, text-fig. 1C). As a result, we posit that the original mandible of PIMUZ A/III0521 could have reached 1.5 m in length and thus approached the sizes of later Jurassic and Cretaceous pliosaurids, whose skull lengths ranged up to over 2 m and had corresponding body lengths of between 9 and 13 m (e.g. Benson et al. 2013; Kear 2003; Knutsen et al. 2012; McHenry 2009).
Conclusions
Our documentation of the incomplete plesiosaur mandible, PIMUZ A/III0521, from the lower Bajocian upper Passwang Formation near Arisdorf in Basel-Land is significant because it provides a new addition to the hitherto sparse record of plesiosaur fossils from Switzerland (e.g. Geister 1998; Peyer 1940; von Meyer 1856; Wild 1968). It is also one of only a handful of pre-Callovian Middle Jurassic plesiosaur occurrences currently reported from Europe, and indeed, worldwide (e.g. Buchy 2004; Gasparini 1997; Gasparini and Fernández 2006; Godefroit 1994; Kear 2012; Sachs and Hornung 2016; Vincent et al. 2007, Vincent et al. 2013b). PIMUZ A/III0521 displays key traits that suggest affinity with Pliosauridae (Benson and Druckenmiller 2014), as well as morphological compatibility with coeval taxa, especially Simolestes keileni (Godefroit 1994), and the rhomaleosaurids Maresaurus coccai (Gasparini 1997) and Borealonectes russelli (Sato and Wu 2008). However, PIMUZ A/III0521 is proportionately larger, with an estimated complete mandible length of around 1.5 m; this not only hints at possible taxonomic distinction, but also implies that pliosaurid-like plesiosaurs had achieved giant body sizes and radiated ecologically as dominant marine apex predators by at least the early Middle Jurassic.
References
Albright, L. B., Gillette, D. D., & Titus, A. L. (2007). Plesiosaurs from the Upper Cretaceous (Cenomanian-Turonian) Tropic Shale of southern Utah, part 1: new records of the pliosaur Brachauchenius lucasi. Journal of Vertebrate Paleontology,27, 31–40.
Andrews, C. W. (1913). A descriptive catalogue of the marine reptiles of the Oxford Clay (Vol. 2). London: British Museum (Natural History).
Andrews, C. W. (1922). Note on the skeleton of a large plesiosaur (Rhomaleosaurus thorntoni sp. n.) from the Upper Lias of Northamptonshire. Annals and Magazine of Natural History,10, 407–415.
Bardet, N., Falconnet, J., Fischer, V., Houssaye, A., Jouve, S., Pereda-Suberbiola, X., et al. (2014). Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Research,26, 869–887.
Benson, R. B. J., & Druckenmiller, P. S. (2014). Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition. Biological Reviews,89, 1–23.
Benson, R. B. J., Evans, M., & Druckenmiller, P. S. (2012). High diversity, low disparity and small body size in plesiosaurs (Reptilia, Sauropterygia) from the Triassic–Jurassic boundary. PLoS ONE,7, e31838.
Benson, R. B. J., Evans, M., Smith, A. S., Sassoon, J., Moore-Faye, S., Ketchum, H. F., et al. (2013). A giant pliosaurid skull from the Late Jurassic of England. PLoS ONE,8, e65989.
Benson, R. B. J., Evans, M., & Taylor, M. A. (2015). The anatomy of Stratesaurus (Reptilia, Plesiosauria) from the lowermost Jurassic of Somerset, United Kingdom. Journal of Vertebrate Paleontology,35, e933739.
Benson, R. B. J., Ketchum, H. F., Noè, L. F., & Gómez-Pérez, M. (2011). New information on Hauffiosaurus (Reptilia, Plesiosauria) based on a new species from the Alum Shale Member (Lower Toarcian: Lower Jurassic) of Yorkshire, UK. Palaeontology,54, 547–571.
Buchy, M.-C. (2004). Plesiosaurs (Reptilia; Sauropterygia) from the Braunjura β (Middle Jurassic; late Aalenian) of southern Germany. Carolinea,62, 51–62.
Burkhalter, R. (1996). Die Passwang-Alloformation (unteres Aalénien bis unteres Bajocien) im zentralen und nördlichen Schweizer Jura. Eclogae Geologicae Helvetiae,89(3), 875–934.
Carpenter, K. (1996). A review of short-necked plesiosaurs from the Cretaceous of the Western Interior, North America. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen,201(2), 259–287.
Carte, A., & Bailey, W. H. (1863). Description of a new species of Plesiosaurus, from the Lias, near Whitby, Yorkshire. Journal of the Royal Dublin Society,4, 160–170.
De Blainville, H. M. D. (1835). Description de quelques espéces de reptiles de la Californie, précédée de l’analyse d’un systéme générale erpetologie et d’amphibiologie. Nouvelles Annales du Muséum d’Historie Naturelle, Paris,4, 233–296.
Druckenmiller, P. S., & Russell, A. P. (2008). Skeletal anatomy of an exceptionally complete specimen of a new genus of plesiosaur from the Early Cretaceous (early Albian) of northeastern Alberta, Canada. Palaeontographica A,283, 1–33.
Fraas, E. (1910). Plesiosaurier aus dem oberen Lias von Holzmaden. Palaeontographica,57, 105–140.
Gasparini, Z. (1997). A new pliosaur from the Bajocian of the Neuquen Basin, Argentina. Palaeontology,40, 135–147.
Gasparini, Z., & Fernández, M. (2006). Middle and Late Jurassic marine reptile faunas of the southeastern Pacific, based on discoveries in Argentina and Chile. Paludicola,5(4), 230–241.
Geister, J. (1998). Lebensspuren von Meersauriern und ihren Beutetieren im mittleren Jura (Callovien) von Liesberg, Schweiz. Facies,39, 105–124.
Godefroit, P. (1994). Simolestes keileni sp. nov., un Pliosaure (Plesiosauria, Reptilia) du Bajocien supérieur de Lorraine (France). Bulletin des Académie et Société Lorraines des Sciences,33(2), 77–95.
Gonzalez, R., & Wetzel, A. (1996). Stratigraphy and paleogeography of the Hauptrogenstein and Klingnau Formations (middle Bajocian to late Bathonian), northern Switzerland. Eclogae Geologicae Helvetiae,89, 695–720.
Holland, T. (2019). The mandible of Kronosaurus queenslandicus Longman, 1924 (Pliosauridae, Brachaucheniinae) from the Lower Cretaceous of northwest Queensland, Australia. Journal of Vertebrate Paleontology,39, e1511569.
Hostettler, B., Reisdorf, A. G., Jaeggi, D., Deplazes, G., Bläsi, H., Morard, A., et al. (2017). Litho- and biostratigraphy of the Opalinus Clay and bounding formations in the Mont Terri rock laboratory (Switzerland). Swiss Journal of Geosciences,110(1), 23–37.
Jordan, P., Wetzel, A., & Reisdorf, A. G. (2008). Jurassic. Swiss Jura Mountains. In T. McCann (Ed.), The geology of Central Europe. Volume 2: Mesozoic and Cenozoic (pp. 880–889). London: Geological Society.
Kear, B. P. (2003). Cretaceous marine reptiles of Australia: a review of taxonomy and distribution. Cretaceous Research,24, 277–303.
Kear, B. P. (2012). A revision of Australia’s Jurassic plesiosaurs. Palaeontology,55, 1125–1138.
Kear, B. P., Fordyce, R. E., Hiller, N., & Siversson, M. (2018). A palaeobiogeographical synthesis of Australasian Mesozoic marine tetrapods. Alcheringa,42, 461–486.
Ketchum, H. F., & Benson, R. B. J. (2011a). A new pliosaurid (Sauropterygia, Plesiosauria) from the Oxford Clay Formation (Middle Jurassic, Callovian) of England: evidence for a gracile, longirostrine grade of Early-Middle Jurassic pliosaurids. Special Papers in Palaeontology,86, 109–129.
Ketchum, H. F., & Benson, R. B. J. (2011b). The cranial anatomy and taxonomy of Peloneustes philarchus (Sauropterygia, Pliosauridae) from the Peterborough Member (Callovian, Middle Jurassic) of the United Kingdom. Palaeontology,54, 639–665.
Knutsen, E. M. (2012). A taxonomic revision of the genus Pliosaurus (Owen, 1841a) Owen, 1841b. Norwegian Journal of Geology,92, 259–276.
Knutsen, E. M., Druckenmiller, P. S., & Hurum, J. H. (2012). A new species of Pliosaurus (Sauropterygia: Plesiosauria) from the Middle Volgian of central Spitsbergen, Norway. Norwegian Journal of Geology,92, 235–258.
Longman, H. A. (1924). A new gigantic marine reptile from the Queensland Cretaceous. Memoirs of the Queensland Museum,8, 26–28.
McHenry, C. R. (2009) Devourer of Gods: The palaeoecology of the cretaceous pliosaur Kronosaurus queenslandicus. Unpublished Ph.D. thesis, University of Newcastle, Australia.
Owen, R. (1860). On the orders of fossil and recent Reptilia, and their distribution in time. Reports of the British Association for the Advancement of Science, London (Vol. 29), pp. 153–166.
Peyer, B. (1940). Saurierwirbel aus dem Schweizer Malm. Eclogae Geologicae Helvetiae,33(2), 285–292.
Sachs, S., & Hornung, J. J. (2016). Marine Tetrapoden aus dem Mittleren Jura (Aalenium und Bajocium) von Bielefeld (Nordwestdeutschland). Berichte Naturwiss. Verein für Bielefeld und Umgegend,53(2015), 52–73.
Sachs, S., Hornung, J. J., & Kear, B. P. (2016). Reappraisal of Europe’s most complete Early Cretaceous plesiosaurian: Brancasaurus brancai Wegner, 1914 from the ‘‘Wealden facies’’ of Germany. PeerJ,4, e2813.
Sachs, S., & Kear, B. P. (2018). A rare new Pliensbachian plesiosaurian from the Amaltheenton Formation of Bielefeld in northwestern Germany. Alcheringa,42, 487–500.
Sato, T., & Wu, X.-C. (2008). A new Jurassic pliosaur from Melville Island, Canadian Arctic Archipelago. Canadian Journal of Earth Sciences,45, 303–320.
Sauvage, H. E. (1873). Notes sur les reptiles fossils. Bulletin of the Geological Society of France,1, 365–380.
Seeley, H. G. (1869). Index to the fossil remains of Aves, Ornithosauria and Reptilia in the Woodwardian Musuem. Cambridge: Deighton, Bell and Co.
Smith, A. S., & Araújo, R. (2017). Thaumatodracon wiedenrothi, a morphometrically and stratigraphically intermediate new rhomaleosaurid plesiosaurian from the Lower Jurassic (Sinemurian) of Lyme Regis. Palaeontographica A,308, 89–125.
Smith, A. S., & Benson, R. B. J. (2014). Osteology of Rhomaleosaurus thorntoni (Sauropterygia: Rhomaleosauridae) from the Lower Jurassic (Toarcian) of Northamptonshire, England. Monograph of the Palaeontographical Society,168(642), 1–40.
Smith, A. S., & Dyke, G. J. (2008). The skull of the giant predatory pliosaur Rhomaleosaurus cramptoni: implications for plesiosaur phylogenetics. Naturwissenschaften,95, 975–980.
Smith, A. S., & Vincent, P. (2010). A new genus of pliosaur (Reptilia: Sauropterygia) from the Lower Jurassic of Holzmaden, Germany. Palaeontology,53, 1049–1063.
Tutin, S. L., & Butler, R. J. (2017). The completeness of the fossil record of plesiosaurs, marine reptiles from the Mesozoic. Acta Palaeontologica Polonica,62, 563–573.
Vincent, P., Bardet, N., & Mattioli, E. (2013a). A new pliosaurid from the Pliensbachian, Early Jurassic of Normandy, Northern France. Acta Palaeontologica Polonica,58, 471–485.
Vincent, P., Bardet, N., & Morel, N. (2007). An elasmosaurid plesiosaur from the Aalenian (Middle Jurassic) of Western France. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen,243(3), 363–370.
Vincent, P., Martin, J. E., Fischer, V., Suan, G., Khalloufi, B., Suchéras-Marx, B., et al. (2013b). Marine vertebrate remains from the Toarcian–Aalenian succession of southern Beaujolais, Rhône, France. Geological Magazine,150(5), 822–834.
von Meyer, H. (1856). Saurier aus der Kreide-Gruppe in Deutschland und der Schweiz. Palaeontographica,6(1), 3–18.
Wegner, T. H. (1914). Brancasaurus brancai n. g. n. sp., ein Elasmosauride aus dem Wealden Westfalens. In: Festschrift für Wilhelm Branca zum 70. Geburtstage 1914 (pp. 235–305). Leipzig: Borntraeger.
Wetzel, A., & Allia, V. (2003). Der Opalinuston in der Nordschweiz: Lithologie und Ablagerungsgeschichte. Eclogae Geologicae Helvetiae,96, 451–469.
Wild, R. (1968). Ein Humerus-Rest eines Plesiosauriers aus dem Oberen Lias von Baden (Kt. Aargau). Eclogae Geologicae Helvetiae,61(2), 581–591.
Williston, S. W. (1903). North American plesiosaurs, Part I. Field Columbian Museum. Geological Series Publication,73(2), 1–77.
Acknowledgements
We are indebted to Dr. Hans Holenweg (Muttenz) for his generous donation of PIMUZ A/III0521 to the Paläontologisches Institut und Museum at the University of Zurich. Our manuscript benefitted from constructive reviews by Rodrigo Otero (Universidad de Chile) and Peggy Vincent (MNHN–CNRS–Sorbonne Université).
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial Handling: D. Marty.
Rights and permissions
About this article

Cite this article
Sachs, S., Klug, C. & Kear, B.P. Rare evidence of a giant pliosaurid-like plesiosaur from the Middle Jurassic (lower Bajocian) of Switzerland. Swiss J Palaeontol 138, 337–342 (2019). https://doi.org/10.1007/s13358-019-00200-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-019-00200-9