
Abstract
In this review, we present the current state of biodiversity knowledge for the class Staurozoa (Cnidaria), including richness estimates, geographical and bathymetric distributions, substrate use, feeding, behavior, life cycle, and conservation. Based on non-parametric, statistical incidence estimators, the global inventory of 50 known and accepted species of stalked jellyfishes might be regarded as close to complete, but we discuss possible bias related to the lower research effort applied in the Southern Hemisphere. Most of the species occur at mid-latitudes, presenting a distributional pattern that disagrees with the classic pattern of diversity (higher richness near the Equator). Specimens are frequently found on algae, but they have also been reported attached to rocks, seagrasses, shells, mud, sand, coral/gorgonian, sea cucumber, and serpulid tube. Most of the species are found in the intertidal and shallow subtidal regions, but species of Lucernaria have been reported at more than 3000 m deep. Amphipods and copepods are the prey items most frequently reported, and stauromedusae have been observed being actively preyed upon by nudibranch mollusks and pycnogonids. Apparently, stalked jellyfishes have a high sensitivity to anthropic impacts in the environment, and promotion of the class, one of the least studied among Cnidaria, is perhaps the best possible conservation strategy.
Similar content being viewed by others

Introduction
Staurozoa is a class of Cnidaria currently represented by 50 species organized in 11 genera, 6 families and 2 suborders (Fig. 1; Table 1) (Miranda et al. 2016a, 2017). As do most of the medusozoans (Petersen 1979; Collins 2002; Collins et al. 2006), stalked jellyfishes have a life cycle with two main generations, known as the stauropolyp and the stauromedusa (Wietrzykowski 1912; Kikinger and Salvini-Plawen 1995; Miranda et al. 2010, 2013). However, the difference between these two stages in Staurozoa is more subtle than in other medusozoan classes (i.e., Cubozoa, Hydrozoa, and Scyphozoa; Marques and Collins 2004; Collins et al. 2006), and completely different from the life cycle of the parasitic Myxozoa (Okamura et al. 2015). In Staurozoa, the metamorphosis is mainly observed in the apical region (i.e., calyx) and the stauromedusa remains attached to the substrate by a peduncle (Fig. 2; Kikinger and Salvini-Plawen 1995; Miranda et al. 2010). The basic structures found in the calyx are the manubrium, the gonads, and eight arms with knobbed tentacular clusters (Fig. 2; Miranda et al. 2013, 2016a). In some species, a rhopalioid (or anchor) lies between each arm (Fig. 2), apparently helping the animals in transience adhere to substrata (Miranda et al. 2013). In general, stauromedusae have a funnel-shaped body (Fig. 2a), although there are many variations, including cruciform species (Fig. 2b, c).

Number of species for staurozoan suborders (Amyostaurida, red shades; and Myostaurida, blue shades), families, and genera

General morphology of stalked jellyfishes. a) Lateral view of Haliclystus borealis (algal substrate shows prominently behind stauromedusa on left half of photograph); b) lateral view of Calvadosia nagatensis (algal substrate shows prominently behind stauromedusa on left half of photograph); c) oral view of Calvadosia cruciformis. Abbreviations: am, arm; an, anchor; cl, calyx; gd, gonad; mn, manubrium; pam, paired arm; pd., pedal disk; pe, peduncle; tc, tentacular cluster
The unique morphology and life cycle of staurozoans have direct implications on their ecology. These animals have a marine benthic habitat and are primarily reported in temperate and boreal waters attached to algae or rocks in intertidal and subtidal pools (Mills and Hirano 2007). Typically, they also have patchy distributions (Miranda et al. 2010, 2012a), possibly related to their asexual reproduction and limited capability of dispersion (Otto 1976, 1978; Miranda et al. 2010). However, there are remarkable exceptions to this general pattern, for example, tropical (Larson 1980; Grohmann et al. 1999) and hydrothermal-associated species (Lutz et al. 1998, 2006; Collins and Daly 2005), which raise important questions about how staurozoans have evolved and diversified. Indeed, information about staurozoan diversity in the literature is still limited, and the class is probably the least known and studied among Cnidaria. Only recently a phylogenetic hypothesis was proposed for Staurozoa, which resulted in a broad taxonomic reassessment (Table 1; Miranda et al. 2016a), demonstrating the need for additional studies focused on this group. We review here for the first time the global diversity of Staurozoa in light of the new phylogenetic proposal (Miranda et al. 2016a). We assess the current state of biodiversity knowledge of the class, including known and estimated richness, geographical and bathymetric distributions, substrate use, feeding, behavior, life cycle, and conservation. This information is crucial for macroecological and macroevolutionary perspectives on the group.
Material and methods
A review of geographic distribution, bathymetric distribution, and substrate use is presented based on information and references listed in the Online Resource 1, Online Resource 2, and Online Resource 3, respectively. Distributional maps were constructed using DIVA-GIS (Hijmans et al. 2001), based on marine realms and ecoregions (Spalding et al. 2007; files available at http://www.marineregions.org/downloads.php). Geographic data were georeferenced from site names described in the literature (Online Resource 1) using Google™ Earth (version 7). We considered that records with equal latitudinal and longitudinal values, to two decimal places, belong to the same locality.
We used the Chao2 statistic (Chao 1987; Colwell and Coddington 1994) in order to estimate the number of undetected species in the assemblage (Colwell and Coddington 1994; Fautin et al. 2013). Chao2 is a non-parametric incidence (presence-absence) estimator that uses counts of singletons (unique) and doubletons (two occurrences), i.e., species that are present in only one and two samples, respectively (rare species), and the total number of observed species in the sample (Basualdo 2011). Chao2 has been shown to perform well in different studies (Melo and Froehlich 2001; Basualdo 2011), and its advantage is that it does not require information on the number of individuals per sample (Unterseher et al. 2008), data rarely mentioned in the literature of Staurozoa. Comparing the observed number of species with an estimate of the total allows one to assess the degree of completeness of inventories (Nakamura and Soberón 2008). EstimateS (version 9; Colwell 2013) was used to calculate the classic and bias-corrected Chao2 estimators, with 95% confidence intervals (see Colwell 2013; see appendix B of EstimateS User’s Guide for more detail). To assess the completeness of the global staurozoan inventory, we calculate the ratio S obs (observed number of species in a sample) to Ŝ Chao2 (estimated number of species), described as the index of completeness (Ĉ; Nakamura and Soberón 2008; Fautin et al. 2013). When Ĉ = 1, the inventory is complete (Nakamura and Soberón 2008; Fautin et al. 2013), and we evaluated this null hypothesis (H 0: Ĉ = 1) using the method described in Nakamura and Soberón (2008), with a significance level of 0.05.
Results & discussion
Ecology and diversity of Staurozoa
Completeness of knowledge and richness estimate
Currently, with 50 valid species (Miranda et al. 2016a, 2017), Staurozoa is one of the least diverse classes among the phylum Cnidaria (Daly et al. 2007). The family Haliclystidae has about half the richness of the class (23 species), and Haliclystus (family Haliclystidae, suborder Myostaurida) and Calvadosia (family Kishinouyeidae, suborder Amyostaurida) are the most diverse genera, with 13 and 11 species, respectively, collectively accounting for almost half of the known staurozoan richness (Fig. 1; Table 1).
We detected five species sampled just once (singletons) and seven doubletons (Tables 2, 3). We compared the observed number of species with richness estimation to assess the degree of completeness of inventories (Ĉ; Table 3). In both cases, using classic and bias-corrected Chao2 estimators, we did not reject the null hypothesis (5% significance level), which suggests that the inventory of stalked jellyfishes, according to these indexes, might be regarded as essentially complete.
However, although this result would indicate a satisfactory current knowledge of staurozoan diversity, a recent staurozoan molecular phylogeny (Miranda et al. 2016a) suggests the existence of at least seven possible new species (including Calvadosia lewisi Miranda et al. 2017, recently described), for which additional material is being collected and/or morphologically analyzed in order to be adequately described (Miranda et al. 2016a, 2017). Another possible indication that the species richness may be an underestimate is the species accumulation curve, which does not seem to be close to saturation (Fig. 3).

Accumulation of the number of described species of Staurozoa through time
In fact, incidence estimators (as Chao2) are expected to compute different species richness values as sampling effort increases (Basualdo 2011). Consequently, these conflicting results can be explained by a bias in sampling effort, which may be related to the restricted number of scientists working on Staurozoa (see example of bias towards collection and taxonomic efforts in Van Soest et al. 2012), especially in the Southern Hemisphere. Six of the seven probable new species pointed out by Miranda et al. (2016a) occur in the Southern Hemisphere (South Africa, Australia, New Zealand, and French Polynesia) and, as sampling in this region increases, the richness of Staurozoa will probably increase (see also Miranda et al. 2017). Indeed, despite broader research programs focusing on the biodiversity of the Southern Ocean (Arntz 1997), the research effort applied to different taxonomic groups has not been uniform and the Northern Hemisphere traditionally has been better sampled than the Southern Hemisphere (López-González and Gili 2000), which is almost certainly the case for Staurozoa (Miranda et al. 2017).
In addition, although most species of Staurozoa occur in shallow waters (see details in the section “Bathymetric range”), there are also species recorded in the deep sea (e.g., Lucernaria janetae in more than 3000 m, Collins and Daly 2005; Lutz et al. 2006). Less than 10% of the deep sea has been mapped, and 60% of the planet is below 1000 m deep (Costello et al. 2012), which might also contribute to a possible underestimation of the diversity of stalked jellyfishes.
Geographic distribution
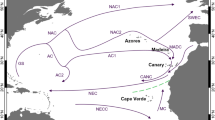
Stalked jellyfishes have been recorded in 415 localities around the world (Fig. 4), 52 (12.5%) of them exclusively for unidentified (or doubtfully identified, i.e., above species level identification) records. These records cover approximately 152° of latitude, from 72.28°S to 79.99°N, and 348° of longitude, from 170.40°W to 177.59°E (Figs. 4, 5 and 6). After assigning published observations to their most probably correct species names, the average number of localities per species (50) is about 10, and 5 species have been recorded from more than 20 localities (Table 2): Haliclystus stejnegeri (21), Depastrum cyathiforme (22), Manania auricula (26), Calvadosia campanulata (33), and Lucernaria quadricornis (66). Twenty-three species (46%) have been recorded in only one band of 10° of latitude, and 5 of them (10%) are known from a single locality in that band (globally unique): Halimocyathus platypus, Manania hexaradiata, Lipkea sturdzii, Lucernaria infundibulum and Lucernaria walteri (Table 2). There are 8 species (16%) exclusive to the polar region, 30 species (60%) exclusive to the temperate region, 8 species (16%) occurring in both temperate and polar regions, and 3 species (6%) exclusive to the tropical region (but one of them, Lucernaria janetae, occurs in the deep sea; Table 2). Thirty-seven species (76%) occur in the Northern Hemisphere, 10 species (18%) in the Southern Hemisphere, and 3 species (6%) in both hemispheres. A higher number of species (6–8) was recorded in the ecoregions “Celtic Seas”, “Gulf of Maine/Bay of Fundy”, “North and East Barents Seas”, “North Sea”, “Northeastern Honshu”, “Sea of Japan/East Sea” and “Yellow Sea” (Fig. 7).

Geographic distribution of Staurozoa. Distribution (filled red circles) plotted over the marine realms proposed by Spalding et al. (2007)

Latitudinal richness of Staurozoa (bands of 10° latitude). Number of species represented by filled black rectangles; hollow rectangles represent genus level identifications, and each rectangle symbolizes a different genus

Global localities where Staurozoa have been recorded. Number of localities (filled black rectangles) where representatives of the class Staurozoa have been recorded, for each band of 10° latitude

Richness of Staurozoa. Number of species of Staurozoa in the marine ecoregions proposed by Spalding et al. (2007). Yellow: 1–2 species; orange: 3–5 species; red: 6–8 species; dark gray (*): genus level identifications (including undescribed species)
Most of the records of stalked jellyfishes are clearly located in the temperate region of the Northern Hemisphere (Figs. 4, 5, 6 and 7). However, there is a possible bias related to collection efforts, as addressed in the section “Completeness of knowledge and richness estimate”. Coastline length can also add a sampling artifact, especially for benthic animals (Fautin et al. 2013). This is likely the case for stauromedusae, with a low number of species at the latitudinal band between 50°–60°S (Figs. 5 and 6), the band with the shortest coastline length.
Even though our understanding of global marine diversity is still limited (Tittensor et al. 2010), the higher diversity of Staurozoa in temperate regions (Fig. 5) disagrees with classic patterns of diversity, in which richness is higher near the Equator and decreases towards the poles (Willig et al. 2003; Fautin et al. 2013). However, there are other exceptions to this pattern, such as the sea anemones (Fautin et al. 2013) and benthic marine algae (Kerswell 2006), groups that have higher species richness at mid-latitudes. The distribution of benthic marine algae is particularly interesting, because algae seem to be the preferential substrate of stauromedusae (see section “Substrate”). Algal genera exhibit a high biodiversity in temperate regions and decreasing richness towards the tropics and polar region (Kerswell 2006), similar to Staurozoa. The biotic interactions between algae and stauromedusae may surpass physical variables, resulting in an apparent close relationship, in which the life cycle of the algae likely affects the life cycle of at least some stalked jellyfishes (Uchida 1929), and these interactions could be a major factor explaining the current distribution of stalked jellyfishes (see section “Substrate” for more information).
The affinity between northern and southern temperate components separated by a distinct equatorial gap, known as antitropicality (Crame 1993), is also remarkable. The evolutionary reason for this pattern is one of the most intriguing questions in biogeography: “how did taxa apparently intolerant to tropical conditions become represented in both Northern and Southern Hemispheres?” (Lindberg 1991; Crame 1993; Burridge 2002: 151). In Staurozoa, the genera Craterolophus, Haliclystus, Lucernaria (excluding the deep-sea records), and Lipkea are antitropical (bipolar and/or bitemperate; Fig. 8). Most of the records of Calvadosia are also antitropical, but the genus is an exception within Staurozoa, with species inhabiting the tropical region (Fig. 8).

Latitudinal distribution of genera of Staurozoa. Deep-sea records in gray
However, apparently there are no bipolar/bitemperate species in the class. Haliclystus auricula, for example, was at one time considered a bipolar species, but molecular and morphological evidence (Miranda et al. 2009, 2010) demonstrate that specimens in the Southern Hemisphere (Argentina: Kramp 1952; Amor 1962; Chile: Zagal 2004a, b, 2008) are actually Haliclystus antarcticus (see Miranda et al. 2010). Haliclystus inabai is also recorded for the temperate Southern and Northern Hemispheres (McInnes 1989; Kramp 1961; Hirano 1986; Zagal et al. 2011; Falconer 2013). However, according to molecular results, the population of the Northern Hemisphere (Japan) is genetically distinct from the Southern Hemisphere (Australia), and the specimens from the Southern Hemisphere could represent a species new to science (Miranda et al. 2016a).
On average, the species of Staurozoa have a latitudinal range two times narrower than their longitudinal range (10° band). Restricted latitudinal ranges might be related to narrow ecological tolerance and/or effective dispersal limitation (Tronholm et al. 2012). Thermal tolerance is one of the most important latitudinal range determinants for marine invertebrates (Tronholm et al. 2012; Jablonski et al. 2013). A potential increase in the temperature of the oceans can negatively affect different life history stages (Byrne 2011; Byrne and Przeslawski 2013), particularly for invertebrates living in the intertidal zone such as many stalked jellyfishes that, in spite of presenting some resilience due to the daily fluctuations in temperature associated with the tidal cycle, are already probably living near their physiological tolerance limits (Byrne and Przeslawski 2013). Additionally, the life cycle of Staurozoa seems to be modulated by seasonal changes in temperature (Miranda et al. 2012a). Therefore, increases in ocean temperature could have serious impacts on staurozoan populations.
Substrate
Stauromedusae are frequently found on algae, for which 8 of 11 genera (72.72%) have been recorded (Fig. 9). The second most common substrate is rock (also recorded as stone and gravel; 54.54%) and seagrass (Zostera; 54.54%), followed by shell (36.36%), mud (18.18%), sand (18.18%) and coral/gorgonian (18.18%), and sea cucumber (9.09%) and serpulid tube (9.09%). The stalked jellyfishes have never been reported attached to artificial substrate (Fig. 9), probably reducing the possibility of human-mediated transport and, consequently, bioinvasion.

Substrate use in Staurozoa. Different substrates recorded for attached stalked jellyfishes (number of substrates for each genus of Staurozoa)
Lucernaria, Calvadosia, and Haliclystus are the genera recorded on the largest number of substrates (six, five and four types, respectively; Fig. 9). These genera are also the most diverse, with 8, 11, and 13 species, respectively (Fig. 1; Table 1).
Generally, stauromedusae are very well camouflaged (color and morphology) to match their surroundings (Fig. 2; Mayer 1910; Corbin 1978; Mills and Hirano 2007). Particularly, camouflage against a background of algae makes these animals very cryptic, although the mechanisms involved in this adaptive (?) process are not well known. Interestingly, it has been observed that when a stalked jellyfish is removed from its algal substrate, it leaves a small whitish disk with the same dimensions of its pedal disk, suggesting that the animal may have absorbed the pigments from the alga (Migot 1922a). There is not enough information to suggest whether stalked jellyfishes are generalists or specialists concerning their algal substrates. However, Haliclystus antarcticus was recorded attached to at least 13 species of algae in the same area in Chile (Zagal 2004a) and Haliclystus “sanjuanensis” was observed on 12 species of algae in the same area in Washington State, USA (CE Mills, pers. obs.).
Several authors have stated that settlement of larvae and metamorphosis are influenced and/or induced by substrate and their characteristics (e.g., associated bacteria) in cnidarians (see review in Leitz 1997; Müller and Leitz 2002; Holst and Jarms 2007) but there is no such information in the staurozoan literature. However, it has been hypothesized that the seasonal appearance of algae in the field has a relationship with the life cycle of stauromedusae. Uchida (1929: 105) reported that along “with the death of Sargassum, the [stauro]medusae gradually disappear”. McInnes (1989) observed that the life cycle of Haliclystus sp. (as Stenoscyphus inabai) “seems to follow the life cycle of the brown seaweed” Cystophora expansa. Berrill (1962: 1249) remarked that Haliclystus auricula “appears to be adapted to living mainly on eel grass”, and with the disappearance of the eel grass in Cape Cod (North American Atlantic Coast) because of pollution and eel grass disease early in the 1930s, the “species has become exceedingly uncommon”. Indeed, studies indicate that shifts in algal abundance may change patterns of larval settlement (Goodbody-Gringley and de Putron 2016), which could be the case in Staurozoa.
In addition, the distribution of Staurozoa is probably influenced by the distribution of their substrates. As for other cnidarians, rafting has an important role on distribution, biodiversity, and evolution of marine organisms (Cornelius 1992a; Thiel and Gutow 2005; Wichmann et al. 2012). Rafting agents, such as macroalgae, can dramatically increase the dispersal range of a variety of marine organisms (Bryan et al. 2012). This long-distance dispersal mechanism may overcome physiological constraints on dispersal ranges for many marine species (Bryan et al. 2012). For example, taxa with “crawl away” juveniles, such as the stalked jellyfishes (benthic, creeping planula; Otto 1976, 1978) probably have significantly lower inherent dispersal ability (Highsmith 1985; Bushing 1994; Wichmann et al. 2012), so rafting could contribute to the breadth of their dispersal range.
Rafting dispersal might have a high impact in the biogeographic structure of the regions in which floating objects (such as macroalgae) are particularly abundant (Wichmann et al. 2012). In this way, the relatively higher abundance of large macroalgae in the temperate region makes rafting a more likely dispersal mode than in the tropics (Highsmith 1985; Bushing 1994; Helmuth et al. 1994; Kerswell 2006). Therefore, animals inhabiting algae in temperate regions, such as stauromedusae, are potential rafters (Highsmith 1985). Different species of the genera Calvadosia, Craterolophus, Haliclystus, Lucernaria, and Manania were collected attached to kelps (e.g., Macrocystis) and/or Sargassum, common rafting agents (Cornelius 1992a; Bushing 1994). Haliclystus kerguelensis, for example, was reported on “detached kelp floating in pool between tide marks” (Kramp 1957).
Additionally, staurozoans were also reported attached to animal substrates, such as mollusks and sea cucumber (Jarms and Tiemann 1996; Salvini-Plawen 2006), so it is also important to highlight the possible contribution of these dispersal agents to the distribution of the benthic stalked jellyfishes. Many benthic invertebrates, including cnidarians, are known to raft on turtles, whales, and fish and, paradoxically, these benthic invertebrates can sometimes traverse greater distances than the potential corresponding to their free swimming stages (Cornelius 1992b).
Evidently, dispersal can occur during different life history stages (Salovius et al. 2005). Although scarce, there are some observations in the literature that different life stages may occupy different substrates from their respective stauromedusa. Polyps of Stylocoronella riedli and Stylocoronella variabilis are interstitial, and were found in coarse sand mixed with shell gravel (Kikinger and Salvini-Plawen 1995), while the stauromedusa stage was recorded just once in the field, attached to a sea cucumber (Salvini-Plawen 2006). The settled planula/frustule of Haliclystus antarcticus (‘microhydrula stage’) was found attached to shells of the bivalve Limopsis hirtella (Jarms and Tiemann 1996; Miranda et al. 2010), whereas the stauromedusa stage is frequently found in algae or rocks (Miranda et al. 2009). Therefore, differential use of substrate along life history amplifies the spectrum of substrate where a researcher can look for these animals in the field, although it may constrain wider distributions when the association with the substrate is obligatory.
In spite of some fragmentary evidence, there is no study on the impact of rafting and/or substrate use on staurozoan distributions. In a recent review on rafting in the marine environment, Thiel and Gutow (2005) revealed a total of 102 hydrozoans, 28 anthozoans, and one scyphozoan for which rafting was confirmed or inferred based on distributional or genetic evidence—no staurozoan was in the list.
Bathymetric range
Specimens of Cnidaria have been recorded in a broad bathymetric range, from intertidal pools to more than 10,200 m deep (Galatheanthemum hadale, Anthozoa; Carlgren 1956). Stauromedusae have been recorded from intertidal pools (82% of the genera) to more than 3000 m deep (Lutz et al. 2006; Fig. 10).

Bathymetric distribution for each genus of Staurozoa. CRATE: Craterolophus; DTRUM: Depastrum; HALIM: Halimocyathus; DEPHA: Depastromorpha; KYOPO: Kyopoda; STYLO: Stylocoronella; HALIC: Haliclystus; LIPKE: Lipkea; CALVA: Calvadosia; MANAN: Manania; LUCER: Lucernaria
The genera Craterolophus, Depastrum, and Halimocyathus have only been recorded in shallow waters, from intertidal to 10 m deep. The genus Depastromorpha was recorded from the intertidal to 14 m deep. The genera Kyopoda and Stylocoronella have never been recorded in intertidal pools, occurring at depths from 8–14 m and 4–22 m, respectively. Haliclystus, Lipkea, Calvadosia, and Manania have been found from the intertidal to intermediate depths, down to 40–60 m (Haliclystus octoradiatus; Kramp 1914), 90 m (Lipkea ruspoliana; Carlgren 1933), 137 m (Calvadosia vanhoeffeni; Smithsonian Online Database, http://collections.nmnh.si.edu/search/iz/), and 110 m (Manania auricula; Kramp 1943), respectively. The genus Lucernaria has the deepest record of the class, being recorded from shallow water to more than 3000 m deep (Fig. 10). Lucernaria janetae, a species inhabiting deep-sea hydrothermal vents (Collins and Daly 2005; Lutz et al. 2006; Voight 2006), is the deepest living stauromedusa ever recorded. Genetic data revealed that specimens of Lucernaria found in different hydrothermal vent sites along the East Pacific Rise are closely related to and maybe conspecific with L. janetae (Lutz et al. 2006), indicating metapopulations with a patchy distribution. In addition, different species of Lucernaria have also been recorded below 200 m deep, such as Lucernaria australis (Vanhöffen 1908), Lucernaria bathyphila (cf. naumov 1961), and Lucernaria sainthilairei (Redikorzev 1925). Lucernaria bathyphila is the only species of Staurozoa reported from a non-vent, abyssal habitat, at a depth of 2800 m, in the Northeast Atlantic Ocean (Naumov 1961; Lutz et al. 2006). It is reasonable to consider that the colonization of deep habitats has some phylogenetic signal in the evolution of Lucernaria, maybe as a synapomorphy of the clade “(L. bathyphila [Arctic and Deep Sea], Lucernaria sp. EPR, L. janetae)” (Miranda et al. 2016a), suggesting a possible shift from an ancestral intertidal habitat to deep waters. However, no special morphological differences in the species living at great depths have been noted (Collins and Daly 2005).
Most reports for Staurozoa refer to the stauromedusa stage. There are few records for other stages, such as planulae and polyps, and their bathymetric ranges (and substrate preferences) in comparison with the respective stauromedusa stages are not known. The settled planula stage of Haliclystus antarcticus was found at about 30 m deep, affixed to a bivalve shell (Jarms and Tiemann 1996; Miranda et al. 2010), whereas the stauromedusa stage is generally recorded in intertidal pools to 8 m deep (Carlgren 1930; Miranda et al. 2010). Polyps of Stylocoronella riedli were recorded from sediments 4 to 22 m deep, whereas the stauromedusa of the same species was found on a holothurian at 8 m deep (Kikinger and Salvini-Plawen 1995; Salvini-Plawen 2006). These examples may indicate that there are different life history stages of Staurozoa living in different habitats and some of these stages probably develop into the respective stauromedusa and migrate under favorable conditions (Kikinger and Salvini-Plawen 1995; Miranda et al. 2012a). Migration, in which the different stages of the life cycle move from the littoral zone to deep waters, and vice-versa, was hypothesized as an explanation for the absence of stauromedusae during some seasons of the year (Miranda et al. 2012a).
Feeding
Zagal (2004b) studied diet by observing gut contents of Haliclystus antarcticus (as H. auricula), collected in Valdivia, Chile, every 2 months over 1 year. The diet consisted of copepods (68.4%), amphipods (15.4%), fly larvae (9.2%), and ostracods (5.9%; Zagal 2004b). Davenport (1998) focused on the trophic relationships of the same species in subantarctic South Georgia and found gut contents with mainly copepods and amphipods, but also remains of an errant polychaete. Besides these two specific studies, staurozoan feeding has been only descriptively reported in several studies: amphipods and copepods are prey items most frequently cited, but insects (fly larvae), polychaete worms, bivalves, isopods, gastropods, ostracods, juvenile decapods (see review in Zagal 2004b), and even sessile ctenophores (Falconer 2013) were also reported. In one instance, a tendency to change prey preference according to size of the stauromedusae was noted: copepods were the most important prey for smaller individuals of H. antarcticus, whereas amphipods were the main item for larger individuals (Zagal 2004b). Further studies are needed to assess whether changing foraging strategies during development is a general characteristic of the group, or may simply reflect the ability of larger individuals to retain larger prey through the entire ingestion process.
Data on feeding behavior is even scarcer. Larson (1980) observed that it would take about 30 min for specimens of Calvadosia corbini to swallow a large prey item. Within 80 min after ingestion, food particles were seen circulating in the gastric cavity and after 5–10 h, the empty carapace of the prey was ejected (Larson 1980). This suggests a somewhat low feeding demand, but, of course, with just a single species investigated, little can be concluded about stauromedusae in general.
Limited information is also available on predation upon stalked jellyfishes. The pycnogonid Phoxichilidium femoratum fed on tentacles of a stauromedusa in aquarium conditions (Prell 1910: 26, Figs. 11, 12), and another pycnogonid of the genus Ammothea often attacks the stauromedusa Manania uchidai (misidentified as M. distincta; Naumov 1961; Uchida and Hanaoka 1933). Stauromedusae have also been seen being preyed upon by nudibranch mollusks (Davenport 1998; Mills and Hirano 2007). Predation pressure was listed as one of the probable reasons that regenerating and injured animals are commonly found in the field (Uchida and Hanaoka 1933; Mills and Hirano 2007; Zagal 2008).
Eight large specimens of H. antarcticus were observed in the stomach of one individual of the fish Notothenia rossii, corresponding to approximately 30% of its stomach content, which also contained about 50% of the volume consisting of macroalgae (Davenport 1998). Typically, macroalgae (the preferential substrate of stalked jellyfishes) is the main stomach content of this fish, which made Davenport (1998) conclude that N. rossii appears to be an opportunistic predator that occasionally takes stauromedusae as bycatch.
Behavior
Stalked jellyfishes have a benthic lifestyle, spending most of their lives attached to substrate (Mills and Hirano 2007). Generally, they use the peduncle to attach to seaweeds, seagrasses, rocks, or other substrates, but they can detach themselves for short periods, floating freely in the water, or moving with the help of the tentacles, anchors, and pad-like structures, sometimes in a somersaulting activity, eventually reattaching with the peduncle (Larson 1980; Mills and Hirano 2007; Miranda et al. 2013). All these structures have characteristic internal organization, with tall cells and gland cells containing granules, probably with supporting and adhesive functions (Miranda et al. 2013, 2016b). However, it has been reported multiple times that Depastrum cyathiforme does not reattach itself once removed from its substrate (Beaumont 1893; Mayer 1910; den Hartog 1976). Migot (1922a) observed that if Haliclystus octoradiatus is accidentally removed from its substrate, it cannot reattach, but remains on the sea bottom and eventually dies. Hyman (1940) reported that detached specimens of “Haliclystus made strenuous efforts to reattach themselves, using the tentacle clusters for adhesion, but never succeeded even when placed on seaweeds” in aquaria. However, personal observations of the same species of Haliclystus (H. “sanjuanensis”) that was observed by Hyman (1940) show that the stauromedusae were able to detach and then reattach to various species of algae in the laboratory, so long as the animals are healthy (CE Mills, pers. obs.). Field observations on Haliclystus antarcticus and Calvadosia corbini indicate that these species can also reattach themselves, moving with the help of anchors or pads and tentacles (LS Miranda pers. obs. of Chilean and Brazilian populations, respectively).
Stalked jellyfishes use mainly the secondary tentacles, with abundant nematocysts (Miranda et al. 2013), to capture their prey (Hyman 1940; Larson 1980). Staurozoa has the least diverse cnidome of the medusozoan classes, with two types of nematocysts in the secondary tentacles, euryteles, and isorhizas (Weill 1934; Larson 1980; Calder 1983; Hirano 1986; Larson and Fautin 1989; Collins and Daly 2005; Fautin 2009; Zagal et al. 2011). Tentacular clusters are probably the most sensitive part of the animal—when touched, they adhere to extraneous objects and usually start a characteristic feeding reaction, bending the arm quickly so that the tentacular cluster with the attached prey encounters the manubrium (Hyman 1940). Larson (1980: 106) described an additional feeding behavior for Calvadosia corbini: when prey touched the tentacles, it adhered to them and the stauromedusa “rapidly and forcefully flipped over”, trapping the prey against the substrate, and the stauromedusa remained with the oral side down, strongly attached to the substrate, until the prey was swallowed. The prey was then engulfed by combined muscular and ciliary activity of the lips, which can stretch to swallow prey as large as its calyx (Larson 1980). The nematocysts and the glandular cells of the gastric filaments and manubrium probably act to subdue prey and to participate in digestion by releasing enzymes (Heeger and Möller 1987; Miranda et al. 2013).
It seems that staurozoans have a great capacity for regeneration, although literature on this topic is scarce. Predation by nudibranchs and pycnogonids (Prell 1910; Uchida and Hanaoka 1933; Mills and Hirano 2007), and the physical action of waves (Amor 1962) can damage body parts, which can later be regenerated. This can explain some abnormal individuals often found in a population (Hornell 1893; Browne 1895; Berrill 1962; Zagal 2008; Miranda et al. 2009). Meyer (1865) studied regeneration of Haliclystus and observed that a calyx can produce a new peduncle, and different parts of the stauromedusa may regenerate, occasionally resulting in an animal with two calyces (Meyer 1865; Mayer 1910). Berrill (1962) also found an individual of Lucernaria quadricornis in nature with two calyces on a single peduncle, apparently sharing the same gastric cavity. Kassianow (1901) cut a Craterolophus convolvulus longitudinally from the oral region to the peduncle and each half regenerated to a new individual. If the section is not so deep, the free edges merge together and restore the former morphology, although a scar may be recognized (Kassianow 1901; Mayer 1910). Additionally, Carlgren (1909), Child (1933), and Hanaoka (1935) found a strong capacity for reconstitution in different staurozoan species (genera Haliclystus, Lucernaria, and Manania) with a remarkable axial physiological gradient.
In general, stauromedusae spawn at night (Wietrzykowski 1910, 1912; Uchida 1929). Females and males of Haliclystus octoradiatus spawn synchronously for generally no longer than 20 min, naturally from 8:00–8:30 pm in July and August (Roscoff, France), but released gametes several hours earlier in the laboratory during the same period (Wietrzykowski 1910, 1912). The animals do not spawn again for 24 h (Wietrzykowski 1912). Species of Haliclystus also spawn in response to light after a period of darkness (Otto 1976; Mills and Hirano 2007). According to Hanaoka (1934), spawning of Manania uchidai (misidentified as M. distincta; Naumov 1961) occurs at 3:00–4:00 am. A natural light-dark cycle with spawning both in the morning and in the evening has also been reported (Otto 1978).
Several species of stalked jellyfishes have been found in high local abundances in nature (e.g., Russell 1904; Corbin 1979; Zagal 2004a), which could be a consequence of physical and/or biological processes (Miranda et al. 2012a). Their eggs have an “extremely sticky” membrane (Otto 1976: 321) and probably remain on the bottom near parent stauromedusae (Mills and Hirano 2007), demonstrating some philopatry. The creeping planulae of Staurozoa are non-ciliated and settle rapidly, which may also contribute to patchy distributions of adults (Otto 1976; Larson 1980). Russell (1904: 63), for example, reported “Depastrum in large numbers under stones”, “very local in its distribution, but generally abundant where it does occur”.
Miranda et al. (2012a) discussed the seasonal abundance in staurozoan populations, correlating these events with the life cycle of the animals, especially with the capability of asexual reproduction. Most staurozoan species are generally more abundant during the summer, but some species have a peak of abundance during winter and autumn (Berrill 1962; Corbin 1979; Zagal 2004a; Mills and Hirano 2007; Miranda et al. 2012a). Different studies confirm that many species of stalked jellyfishes disappear for several months before they reappear under supposedly favorable conditions (Otto 1978; Kikinger and Salvini-Plawen 1995; Mills and Hirano 2007; Miranda et al. 2012a).
Life cycle
Species of the class Staurozoa have a particular life cycle, consisting of a benthic planula larva that settles on the substrate and develops into a stauropolyp, which subsequently undergoes an apical metamorphosis into stauromedusa, without strobilation or budding (Wietrzykowski 1912; Kikinger and Salvini-Plawen 1995; Miranda et al. 2010). As a result, stauromedusae are unique because they bear characteristics found both in polyp and medusa stages of other medusozoans (Collins et al. 2006).
Kowalevsky (1884) was one of the first authors to study the development of a staurozoan, Calvadosia campanulata (as Lucernaria; see Uchida 1929; Hanaoka 1934), from cleavage to the attachment of the larva on a substrate. Bergh (1888) studied the early development of H. octoradiatus and confirmed the results found by Kowalevsky (1884), without many further observations. Wietrzykowski (1910, 1911, 1912) left an important and thorough contribution on the development of staurozoans, detailing the external and internal anatomy of all life cycle stages of H. octoradiatus. Uchida (1929: 150, 151), in reviewing the taxonomy of stalked jellyfishes, added information on the metamorphosis of some species (especially on Haliclystus inabai; Haliclystus tenuis, as H. auricula; and Calvadosia cruciformis), highlighting that “the structure [presence/absence, number, location] of the primary and secondary tentacles and the number of chambers in the stalk […] generally metamorphose with age”. Hanaoka (1934) reported aspects of the early development of Manania uchidai (misidentified as M. distincta; Naumov 1961), from fertilization to the planula stage. Otto (1976, 1978) provided more detailed information on development, locomotion, and settlement of planulae of Haliclystus species. Hirano (1986) listed and described the species of stalked jellyfishes found in Hokkaido (Japan), including notes on their metamorphosis, corroborating Uchida’s (1929) observations about the changes in the number of chambers in the peduncle during development of species of Haliclystus and Manania. Therefore, the generalized idea that “in the development of the stalked medusa, the upper part alone metamorphoses, while the lower part remains unchanged” (Uchida 1973: 137) is incorrect. Kikinger and Salvini-Plawen (1995) succeeded in following the metamorphosis from the polyp into the medusa of Stylocoronella riedli and S. variabilis in the laboratory, adding important data on the development of the group, such as asexual reproduction of the polyps and frustules, and encystment of frustules. More recently, Miranda et al. (2010) unveiled the life cycle of Haliclystus antarcticus using molecular, morphological, and ecological data, associating the miss-assigned hydrozoan species Microhydrula limopsicola as a stage of the life cycle of the staurozoan H. antarcticus. As previous authors (Wietrzykowski 1912; Hanaoka 1934; Otto 1976, 1978), Miranda et al. (2010) discussed the ability of planulae to aggregate and encyst, and included a putative scheme of the life cycle of H. antarcticus (Miranda et al. 2010, Fig. 5), highlighting the ability for asexual reproduction of some stages.
The zygotes (also generally called “eggs”) of stauromedusae are among the smallest in the Metazoa (see Kume and Dan 1968; Strathmann 1987), ranging so far as is known from 30 to 50 μm. The zygotes of Haliclystus octoradiatus are reported to be 30 μm in diameter (Wietrzykowski 1912). Hanaoka (1934) reports that the zygotes of Manania uchidai (misidentified as M. distincta; Naumov 1961) are 50 μm in diameter. Otto (1976) reports zygotes diameters of 35 μm for Haliclystus “sanjuanensis” (as H. stejnegeri) and 40 μm for Haliclystus sp. (as H. salpinx).
Based on all these observations, we can hypothesize the general life cycle of staurozoans to comprise the following series: gametes are released into the gastrovascular cavity and expelled through the mouth (Otto 1976) of male and female stauromedusae (sexes are separate; for details of the structure of the gonads, see Uchida 1929; Eckelbarger and Larson 1993; Miranda et al. 2013); under favorable conditions, they can spawn daily throughout one month (Corbin 1979; Mills and Hirano 2007), generally at night or in response to light stimulus after a period of darkness (Wietrzykowski 1912; Uchida 1929; Hanaoka 1934; Otto 1976, 1978); after fertilization in the water column, an inconspicuous but sticky membrane forms around the zygote, adhering it to the substrate (Hanaoka 1934; Otto 1976); then, the small zygote, granulated with yolk substance (Hanaoka 1934; Otto 1976), undergoes an irregular cleavage and gastrulation through unipolar ingression (Hanaoka 1934; Otto 1976); the gastrula becomes elongated and the planula is formed (Hanaoka 1934); the planula is characterized by 16 endodermal cells (constant number) lined up in a single row, covered by a layer of ectodermal cells, and does not possess cilia, moving by a series of extensions and retractions (Otto 1976, 1978); after creeping for 1–3 days, the planula settles (Otto 1976), recruits, and undergoes metamorphosis into a stauropolyp; the stauropolyp stage develops, forming the primary tentacles (Wietrzykowski 1912; Hirano 1986; Kikinger and Salvini-Plawen 1995; Miranda et al. 2010); gradually, the apical part of the stauropolyp starts to undergo visible modifications (e.g., formation of arms, gastric filaments, gonads, and adradial tentacular clusters; disappearance or modification of the eight primary tentacles) and the stauropolyp metamorphoses into a stauromedusa; however, internally and basally, the stauropolyp may also modify (e.g., change in the number of chambers of the peduncle; Uchida 1929; Hirano 1986); the stauromedusa then grows and reaches its maturity. In general, a life cycle of one generation per year has been reported, with an abundance peak and maturation during the summer, although variations occur depending on the species studied (Berrill 1962; Corbin 1979; Eckelbarger and Larson 1993; Kikinger and Salvini-Plawen 1995; Miranda et al. 2012a). There is no information about the proper stimulus needed for a planula to develop into a stauropolyp (Otto 1976, 1978; Jarms and Tiemann 1996; Miranda et al. 2010). Asexual reproduction has been reported in the settled planula, frustule, and stauropolyp stages (Wietrzykowski 1912; Kikinger and Salvini-Plawen 1995; Jarms and Tiemann 1996; Miranda et al. 2010). Frustules probably constitute resting stages that develop into planulae or small stauropolyps under favorable conditions (Kikinger and Salvini-Plawen 1995; Miranda et al. 2012a). In addition, it has been reported that some planulae can cluster together, secreting a gelatinous substance that encysts all of them, probably constituting an “overwintering stage” (Kowalevsky 1884; Hanaoka 1934; Otto 1978; Miranda et al. 2012a).
Conservation
Currently, there are 882 species of Cnidaria in the IUCN red list of threatened species (www.iucnredlist.org), nearly all belonging to the class Anthozoa (866 species), with the remaining in Hydrozoa (16 species). There is no official data in the IUCN red list regarding endangered species of Staurozoa. However, three staurozoan species (Haliclystus auricula, Calvadosia campanulata, and Calvadosia cruxmelitensis) have been included in the list of UK BAP priority marine species (http://jncc.defra.gov.uk/page-5167).
Although scarce, there is information in the literature concerning the impact of water pollution in populations of stalked jellyfishes. More than 100 years ago, Mayer (1910) reported that pollution in Massachusetts (USA) may have decreased the number of stauromedusae found in this area. According to Mayer (1910), “the contamination of the sea-water in this region has destroyed the Stauromedusae which once abounded there, and which are now exceedingly rare along the entire New England coast”. Berrill (1962) also expressed his concern for the same area and mentioned that “Haliclystus auricula […] appears to be adapted to living mainly on eel grass, and, with the disappearance of the eel grass early in the 1930s, this species has become exceedingly uncommon. Where the eel grass has returned to some extent, it is usually in waters affected by pollution. In Massachusetts Bay, where Clark did most of his collecting, the water has become so polluted that the more delicate coelenterate life has disappeared”. Indeed, one species observed on seagrass and described by Clark (1863) from this region, Halimocyathus platypus, has never been observed since. Therefore, the proposed association between the disappearance of stalked jellyfishes with the disappearance of their substrates (Berrill 1962) seems reasonable. Larson (1976) highlighted that stauromedusae apparently “are very sensitive to environmental conditions”. Zagal (2004a) reported that different factors should be considered in evaluating habitat selection of stalked jellyfishes, including pollution. Miranda et al. (2012b) proposed that specimens of Calvadosia capensis were never found again on the Southern coast of São Paulo (Brazil) because of the “gradual pollution and environmental decay” along the region. In addition, Miranda and Marques (2016) discussed the possible influence of a dam collapse on populations of Calvadosia corbini in Espírito Santo, Brazil. Therefore, although we cannot affirm whether water pollution and other anthropic impacts affects the populations of stauromedusae directly or indirectly (i.e., through their substrate), there are numerous reports in the literature correlating these factors with the decrease of some stauromedusae populations.
In terms of conservation, it may be virtually impossible to directly protect any individual species of Staurozoa. Consequently, conservation of Staurozoa is likely accomplished by protection of the habitat that they require. Perhaps the most effective conservation strategy for stauromedusae is promotion of the group, encouraging increased awareness, recording and monitoring, and studying patterns of behavior and factors that influence their appearance and disappearance.
Staurozoan genera diversity and ecology
Craterolophus
Two valid species: Craterolophus convolvulus (Johnston 1835) and Craterolophus macrocystis von Lendenfeld 1884.
Craterolophus convolvulus was first described based on two specimens from Berwick Bay, Scotland (Johnston 1835), and was later recorded from several different parts of Europe, including England, France, and Germany, as well as on the northeast coast of the USA (Online Resource 4). The other species in the genus, C. macrocystis, was recorded from the east coast of New Zealand, from Port Chalmers (near Dunedin) and Lyttleton (Online Resource 4). It is a rare species (von Lendenfeld 1884) that has not been recorded again since its original description.
Both species were recorded in shallow tide pools (Fig. 10), attached to different species of algae (Fig. 9). Craterolophus convolvulus has two generations per year (i.e., it has two peaks in its seasonal cycle): “a main peak of maximum abundance in spring and a secondary peak in late summer” (Corbin 1979: 387).
Calvadosia
Eleven valid species: Calvadosia campanulata (Lamouroux 1815), Calvadosia nagatensis (Oka 1897), Calvadosia vanhoeffeni (Browne 1910), Calvadosia cruciformis (Okubo 1917), Calvadosia hawaiiensis (Edmondson 1930), Calvadosia tsingtaoensis (Ling 1937), Calvadosia capensis (Carlgren 1938), Calvadosia cruxmelitensis (Corbin 1978), Calvadosia corbini (Larson 1980), Calvadosia tasmaniensis (Zagal et al. 2011), and Calvadosia lewisi Miranda et al. 2017.
Calvadosia has a unique distribution within Staurozoa (Figs 8, Online Resource 5). While most species are found in temperate to polar waters (Figs. 4, 5, 6, 7, and 8), some species of Calvadosia have been reported from tropical waters (Hawaii, USA; Puerto Rico; Veracruz, Mexico; Espírito Santo and Bahia, Brazil; and unidentified/undescribed records in Moorea, French Polynesia—see Miranda et al. 2016a; and India, Panikkar 1944), in addition to several countries in Europe, Japan, China, New Zealand (undescribed species; see Miranda et al. 2016a), Australia, South Africa, and Southern Ocean (Online Resource 5).
Species of Calvadosia have been recorded attached to different substrates: algae, seagrass (Zostera), rocks, shells, and corals (Fig. 9). They have also been recorded over a broad bathymetric range, from intertidal pools to 137 m deep (Fig. 10). Corbin (1979) reported a seasonal cycle of one generation per year for C. campanulata (peak during autumn) and C. cruxmelitensis (peak during winter).
Depastromorpha
Monospecific, Depastromorpha africana Carlgren 1935.
The type locality of D. africana is Cape Town, South Africa (Carlgren 1935), and the species was described based on one specimen (Carlgren 1935; Zagal et al. 2011; see additional material and localities in Miranda et al. 2017). More recently, the species has been recorded in Australia and New Zealand (Online Resource 6; Grohmann et al. 1999; Cairns et al. 2009; Zagal et al. 2011; Miranda et al. 2017); however, further studies may reveal that the Australian material represents a new species (see Miranda et al. 2016a, 2017).
Depastromorpha africana was recorded attached to seagrass and different species of algae (Fig. 9), generally in the intertidal or shallow subtidal zones (Fig. 10; Carlgren 1935; Zagal et al. 2011; Miranda et al. 2017). The species is locally rare in Australia, although ca. 100 individuals were observed in D’Entrecasteaux Channel (Tasmania, Australia; Zagal et al. 2011).
Depastrum
Monospecific, Depastrum cyathiforme (Sars 1846).
Depastrum cyathiforme has been recorded in different parts of England, France, Ireland, Norway (including Spitzbergen), Scotland, and Spain (Canary Islands; Online Resource 7).
Depastrum cyathiforme has generally been reported attached to rocks, but is also found in mud-covered surfaces between stones, or attached to algae (Fig. 9). Specimens were observed in intertidal pools (Fig. 10). Allman (1860: 126) could not discern if these animals have “the power of detaching” themselves after they become attached, and Beaumont (1893: 254) remarked that he never saw a detached specimen reattach if removed from its substrate (see also Mayer 1910: 524); den Hartog (1976: 3) found three specimens in the field and these also did not reattach in the laboratory under running sea water conditions. Russell (1904) reported a large number of specimens, but very local in distribution, so the species is considered to be generally rare (Mayer 1910).
Haliclystus
Haliclystus is the most diverse genus of Staurozoa, with 13 valid species: Haliclystus auricula Clark 1863; Haliclystus octoradiatus Clark 1863; Haliclystus salpinx Clark 1863; Haliclystus inabai (Kishinouye 1893); Haliclystus antarcticus Pfeffer 1889; Haliclystus stejnegeri Kishinouye 1899; Haliclystus kerguelensis Vanhöffen 1908; Haliclystus tenuis Kishinouye 1910; Haliclystus borealis Uchida 1933; Haliclystus sinensis Ling 1937; Haliclystus monstrosus (Naumov 1961); Haliclystus californiensis Kahn et al. 2010; and Haliclystus “sanjuanensis” nomen nudum.
Haliclystus “sanjuanensis” was originally described in Gellermann (1926)’s unpublished M.Sc. thesis, and although the species has never been formally described (nomen nudum), the name has been used repeatedly by, e.g., Hyman (1940), Hirano (1997), Miranda et al. (2009), Kahn et al. (2010); synonymized with H. auricula (Gwilliam 1956; Kramp 1961); or misidentified as H. stejnegeri (Otto 1976, 1978) and H. octoradiatus (Eckelbarger and Larson 1993).
Haliclystus has a widespread and antitropical distribution (Fig. 8), with records from both the west and east coasts of the USA, Arctic, different parts of Europe, Japan, China, Russia, South Korea, Australia, Argentina, Chile, and Antarctica (Online Resource 8). Further studies are necessary to reveal whether Haliclystus inabai from Australia (McInnes 1989; Falconer 2013) represents a new species (see Haliclystus sp. AUS; Miranda et al. 2016a).
Species of Haliclystus have been reported attached to algae, seagrass (Zostera), rocks, and shell (Fig. 9). Migot (1922a, b) and Singla (1976) studied the mechanisms of attachment. Haliclystus is found mainly in shallow waters, from tide pools to 40–60 m deep (Fig. 10). Different stages of the life cycle may occupy different habitats: the stauromedusa of H. antarcticus is generally found in shallow water (0–8 m deep), attached to rocks or algae, whereas its planulae were recorded ca. 30 m deep, attached to shells (Miranda et al. 2010). As far as is known, Haliclystus species have an annual life cycle, with a summer peak (Berrill 1962; Corbin 1979; Zagal 2004a; Miranda et al. 2012a).
Halimocyathus
Monospecific, Halimocyathus platypus Clark 1863. A second species, Halimocyathus lagena (cf. Mayer 1910; Kramp 1914, 1943, 1961), is a synonym of Manania auricula (Clark 1863; Larson and Fautin 1989).
Only a single individual of H. platypus has ever been recorded, at Chelsea Beach, Massachusetts Bay, USA (Online Resource 9; Clark 1863).
Halimocyathus platypus was found attached to Zostera marina (seagrass), at half tide (Figs. 9 and 10; Clark 1863). Mayer (1910) highlighted that the contamination of the seawater at Chelsea Beach (Massachusetts) had devastated the stauromedusae that were once abundant there.
Manania
Seven species: Manania auricula (Fabricius 1780); Manania hexaradiata (Broch 1907); Manania distincta (Kishinouye 1910); Manania atlantica (Berrill 1962); Manania uchidai (Naumov 1961); Manania gwilliami Larson and Fautin 1989; and Manania handi Larson and Fautin 1989.
Recorded from the Northern Hemisphere (Fig. 8), on the east and west coasts of the USA, north and west coasts of Canada, Greenland, Europe, Japan, and Russia (Online Resource 10).
Species of Manania have been recorded in the literature from the intertidal zone to 110 m deep (Fig. 10), attached to algae and seagrass (Zostera; Fig. 9).
Kyopoda
Monospecific, Kyopoda lamberti Larson 1988.
Kyopoda lamberti has been recorded for British Columbia and Southern California (Online Resource 11; Larson 1988), but its range is possibly more or less continuous between these localities, and maybe it has been overlooked because of its small size, cryptic coloration, and high-energy subtidal habitat (Larson 1988).
Specimens have been found attached to cobbles and a coralline algae-covered rock wall at subtidal depths (8–14 m deep; Figs. 9 and 10). The particular morphology of K. lamberti, with stomach and gonads near the substrate, located in the peduncle rather than in the calyx, suggests an adaptation to areas with strong wave surge (Larson 1988).
Lipkea
Three valid species: Lipkea ruspoliana Vogt 1886; Lipkea sturdzii (Antipa 1893), and Lipkea stephensoni Carlgren 1933.
Species of Lipkea have been recorded for Europe and South Africa (Online Resource 12). There are also unidentified Lipkea records from Australia (Zagal et al. 2011), New Zealand (Cairns et al. 2009), and Japan (Miranda et al. 2016a).
Lipkea is one of the three genera of Staurozoa that have never been documented attached to algae, recorded only on rocks, gorgonian, and serpulid tubes (Fig. 9), from the intertidal zone to 90 m deep (Fig. 10). It is the only staurozoan genus that has been observed to appear unexpectedly in aquaria (Monaco, Pisani et al. 2007; Japan, YM Hirano pers. obs.).
Lucernaria
Eight valid species: Lucernaria quadricornis Müller 1776; Lucernaria bathyphila Haeckel 1879; Lucernaria infundibulum Haeckel 1879; Lucernaria haeckeli (Antipa 1892); Lucernaria walteri (Antipa 1892); Lucernaria australis Vanhöffen 1908; Lucernaria sainthilairei (Redikorzev 1925); and Lucernaria janetae Collins and Daly 2005.
The distribution of Lucernaria is bipolar but more concentrated in the Northern Hemisphere (Fig. 8; Arctic, Northern Europe, and East USA) than in the Southern Hemisphere (Antarctica), given the present sampling (Online Resource 13). Additional records are for the deep-sea Lucernaria janetae from the East Pacific Rise (Online Resource 13; Collins and Daly 2005; Lutz et al. 2006).
Species of Lucernaria have been recorded attached to algae, seagrass, rocks, mud/clay (with gravel), shell, and sand (Fig. 9), from the intertidal zone to 3001 m deep, the deepest record for staurozoans (Fig. 10). Lucernaria janetae was recorded near a hydrothermal vent (Collins and Daly 2005; Lutz et al. 2006; Voight 2006).
Stylocoronella
Two valid species: Stylocoronella riedli Salvini-Plawen 1966 and Stylocoronella variabilis Salvini-Plawen 1987.
Species of Stylocoronella have been recorded for Europe (Croatia, England, France, and Spain; Online Resource 14).
Stylocoronella is one of the three genera of Staurozoa never recorded attached to algae (Fig. 9). The stauropolyp was found inhabiting the interstitia of coarse sand and shell-sand, and the stauromedusa was found attached to a sea cucumber (Salvini-Plawen 2006). The species were recorded from 4 to 22 m deep (Fig. 10).
References
Allman GI (1860) On the structure of Carduella cyathiformis. A contribution to our knowledge of Lucernariadae. Trans Microsc Soc London VIII:125–128
Amor A (1962) Sobre Stauromedusae del litoral Patagonico. Notas Mus La Plata Zool 20:89–96
Antipa G (1892) Die Lucernariden der Bremer Expedition nach Ostspitzbergen im Jahre 1889. Zool Jb Abt Syst Geogr Biol Tiere 6:377–397
Antipa G (1893) Eine neue Stauromeduse (Capria n. sturdzii n.) Mitt Zool Stn Neapel 10:618–632
Arntz WE (1997) Investigación antártica en biología marina: situación actual, proyectos internacionales y perspectivas. Bol R Soc Esp Hist Nat Sec Biol 93:13–44
Basualdo CV (2011) Choosing the best non-parametric richness estimator for benthic macroinvertebrates databases. Rev Soc Entomol Argent 70:27–38
Beaumont WI (1893) Note on Lucernarians occurring in the neighbourhood of Port Erin, Isle of Man. Proc Trans Liverpool Biol Soc 7:256–263
Bergh RS (1888) Bemaerkninger om Udviklingen af Lucernaria. Vidensk Meddel Naturhist Foren Kjøbenhavn 1888:214–220
Berrill M (1962) The biology of three New England stauromedusae, with a description of a new species. Can J Zool 40:1262–1249
Broch H (1907) Hydroiden und Medusen. Rep Sec Norw Arct Exp “Fram” 1898–1902 2:1–12
Browne ET (1895) On the variation of Haliclystus octoradiatus. Q J Microsc Sci 38:1–8
Browne ET (1910) Reports of the natural history results of the voyage of the SS Discovery in the Antarctic regions in 1901, under Captain R F Scott RN. Coelentera. V. Medusae. London British Museum (Natural History), London
Bryan SE, Cook AG, Evans JP, Hebden K, Hurrey L, Colls P, Jell JS, Weatherley D, Firn J (2012) Rapid, long-distance dispersal by pumice rafting. PLoS One 7:e40583
Burridge CP (2002) Antitropicality of Pacific fishes: molecular insights. Environ Biol Fish 65:151–164
Bushing W (1994) Biogeographic and ecological implication of kelp rafting as a dispersal vector for marine invertebrates. In: Halvorson WL, Maender GJ (eds) The fourth California Islands symposium: update on the status of resources. Santa Barbara Museum of Natural History, Santa Barbara, pp 103–108
Byrne M (2011) Impact of ocean warming and ocean acidification on marine invertebrate life history stages: vulnerabilities and potential for persistence in a changing ocean. Oceanogr Mar Biol 49:1–42
Byrne M, Przeslawski R (2013) Multistressor impacts of warming and acidification of the ocean on marine invertebrates’ life histories. Integr Comp Biol 53:582–596
Cairns SD, Gershwin L-A, Brook FJ, Pugh P, Dawson EW, Ocaña VO, Vervoort W, Williams G, Watson JE, Opresko DM, Schuchert P, Hine PM, Gordon DP, Campbell HJ, Wright AJ, Sánchez JA, Fautin DG (2009) Phylum Cnidaria: corals, medusae, hydroids, myxozoans. In: Gordon DP (ed) New Zealand inventory of biodiversity, vol Volume 1. Canterbury University Press, Christchurch, pp 59–101
Calder DR (1983) Nematocysts of stages in the life cycle of Stomolophus meleagris, with keys to scyphistomae and ephyrae of some western Atlantic Scyphozoa. Can J Zool 61:1185–1192
Carlgren O (1909) Studien über Regenerations- und Regulationserscheinungen. III. Versuche an Lucernaria. K Sven Vetensk Akad Handl 44:1–44
Carlgren O (1930) Die Lucernariden. Furth Zool Res Swed Antarc Exp 1901–1903 2:1–18
Carlgren O (1933) Zur Kenntnis der Lucernariiden Lipkea, Capria und Brochiella. K Fysiogr Sällsk Handl 44:1–19
Carlgren O (1935) Über eine neue Südafrikanische Lucernariidae, Depastromorpha africana n. gen., n. sp., nebst Bemerkungen über den Bau und die Systematik dieser Tiergruppe. K Sven Vetensk Akad Handl 15:1–24
Carlgren O (1938) Eine neue südafrikanische Lucernariidae, Lucernariopsis capensis. K Fysiogr Sällsk Lund Förh 8:1–6
Carlgren O (1956) Actiniaria from depths exceeding 6000 meters. Galathea Rep 2:9–16
Chao A (1987) Estimating the population size for capture-recapture data with unequal catchability. Biometrics 43:783–791
Child CM (1933) Reconstitution in Haliclystus auricula Clark. Sci Rep Tohoku Imp Univ 8:75–106
Clark HJ (1863) Prodromus of the history, structure, and physiology of the order Lucernariae. J Boston Soc Nat Hist 7:531–567
Collins AG (2002) Phylogeny of Medusozoa and the evolution of cnidarians life cycles. J Evolution Biol 15:418–432
Collins AG, Daly M (2005) A new deepwater species of Stauromedusae, Lucernaria janetae (Cnidaria, Staurozoa, Lucernariidae), and a preliminary investigation of stauromedusan phylogeny based on nuclear and mitochondrial rDNA data. Biol Bull 228:221–230
Collins AG, Schuchert P, Marques AC, Jankowski T, Medina M, Schierwater B (2006) Medusozoan phylogeny and character evolution clarified by large and small subunit rDNA data and an assessment of the utility of phylogenetic mixture models. Syst Biol 55:97–115
Colwell RK (2013) EstimateS, version 9.1.0. Statistical estimation of species richness and shared species from samples (Software and User’s Guide). http://viceroy.eeb.uconn.edu/estimates/index.html. Accessed 15 Jul 2016
Colwell RK, Coddington JA (1994) Estimating terrestrial biodiversity through extrapolation. Philos T Roy Soc B 345:101–118
Corbin PG (1978) A new species of the stauromedusan genus Lucernariopsis (Coelenterata: Scyphomedusae). J Mar Biol Assoc UK 58:285–290
Corbin PG (1979) The seasonal abundance of four species of Stauromedusae (Coelenterata: Scyphomedusae) at Plymouth. J Mar Biol Assoc UK 59:385–391
Cornelius PFS (1992a) Medusa loss in leptolid Hydrozoa (Cnidaria), hydroid rafting, and abbreviated life-cycles among their remote-island faunae: an interim review. Sci Mar 56:245–261
Cornelius PFS (1992b) The Azores hydroid fauna and its origin, with discussion of rafting and medusa suppression. Arquipélago–Life Earth Sci 10:75–99
Costello MJ, Wilson S, Houlding B (2012) Predicting total global species richness using rates of species description and estimates of taxonomic effort. Syst Biol 6:871–883
Crame JA (1993) Bipolar molluscs and their evolutionary implications. J Biogeogr 20:145–161
Daly M, Brugler MR, Cartwright P, Collins AG, Dawson MN, Fautin DG, France SC, McFadden CS, Opresko DM, Rodriguez E, Romano SL, Stake JL (2007) The phylum Cnidaria: a review of phylogenetic patterns and diversity 300 years after Linnaeus. Zootaxa 1668:127–182
Davenport J (1998) Note on the trophic relationships of the stauromedusa Haliclystus antarcticus from subantarctic South Georgia. J Mar Biol Assoc UK 78:663–664
den Hartog JC (1976) A record of the stauromedusa Depastrum cyathiforme (M. Sars, 1846) in France. Trav Stat Biol Roscoff 23:3–4
Eckelbarger KJ, Larson RJ (1993) Ultrastructural study of the ovary of the sessile scyphozoan, Haliclystus octoradiatus (Cnidaria: Stauromedusae). J Morph 218:225–236
Edmondson CH (1930) New Hawaiian medusae. Bernice P. Bishop Mus Occas Pap 9:1–16
Fabricius O (1780) Fauna Groenlandica: systematice sistens animalia Groenlandiae occidentalis hactenus indagata, quoad nomen specificum, triviale, vernaculumque; synonyma auctorum plurium, descriptionem, locum, victum, generationem, mores, usum, capturamque singuli, prout detegendi occasio fuit, maximaque parte secundum proprias observations.Gottlob Rothe, Hafniae et Lipsiae
Falconer A (2013) A stalked jellyfish Stenoscyphus inabai (Kishinouye, 1893) (Stauromedusae), found at the jawbone, Port Phillip Bay, Victoria. Victorian Nat 130:202–207
Fautin DG (2009) Structural diversity, systematics, and evolution of cnidae. Toxicon 54:1054–1064
Fautin DG, Malarky L, Soberón J (2013) Latitudinal diversity of sea anemones (Cnidaria: Actiniaria). Biol Bull 224:89–98
Gellermann MP (1926) Medusae of the San Juan Archipelago. Thesis, University of Washington
Goodbody-Gringley G, de Putron S (2016) Brooding corals: planulation patterns, larval behavior, and recruitment dynamics in the face of environmental change. In: Goffredo S, Dubinsky Z (eds) The Cnidaria, past, present and future. The world of medusa and her sisters. Springer, Switzerland, pp 279–289
Gosse PH (1858) Synopsis of the families, genera, and species of the British actiniae. Ann Mag Nat Hist 3:414–419
Grohmann PA, Magalhães MP, Hirano YM (1999) First record of the order Stauromedusae (Cnidaria, Scyphozoa) from the tropical southwestern Atlantic, with a review of the distribution of Stauromedusae in the Southern Hemisphere. Spec Div 4:381–388
Gwilliam GF (1956) Studies on west coast Stauromedusae. Dissertation, University of California
Haeckel E (1879) Das system der medusen. I, 2: System der Acraspeden. Zweite Hälfte des Systems der Medusen. Gustav Fischer, Jena
Hanaoka K-I (1934) Notes on the early development of a stalked medusa. Proc Imp Acad 10:117–120
Hanaoka K-I (1935) Experiments on the polarity of a stalked medusa Thaumatoscyphus distinctus Kishinouye. Jour Fac Science, Hokkaido Imp Univ, Series VI, Zool 4:159–181
Heeger T, Möller H (1987) Ultrastructural observations on prey capture and digestion in the scyphomedusa Aurelia aurita. Mar Biol 96:391–400
Helmuth B, Veit RR, Holberton R (1994) Long-distance dispersal of a subantarctic brooding bivalve (Gaimardia trapesina) by kelp-rafting. Mar Biol 120:421–426
Highsmith RC (1985) Floating and algal rafting as potential dispersal mechanisms in brooding invertebrates. Mar Ecol Prog Ser 25:169–179
Hijmans RJ, Guarino L, Cruz M, Rojas E (2001) Computer tools for spatial analysis of plant genetic resources data: 1. DIVA-GIS. Plant Genet Resour Newsl 127:15–19
Hirano YM (1986) Species of Stauromedusae from Hokkaido, with notes on their metamorphosis. Jour Fac Science, Hokkaido Imp Univ, Series VI, Zool 24:182–201
Hirano YM (1997) A review of a supposedly circumboreal species of stauromedusa, Haliclystus auricula (Rathke, 1806). In: den Hartog JC (ed) Proceedings of the 6th international conference on coelenterate biology. The Netherlands. Nationaal Naturhistorisch Museum, Noordwijkerhout, Leiden, pp 247–252
Holst S, Jarms G (2007) Substrate choice and settlement preferences of planula larvae of five Scyphozoa (Cnidaria) from German bight, North Sea. Mar Biol 151:863–871
Hornell J (1893) Observations of certain marine animals IV. Abnormalities in Haliclystus (Lucernaria) octoradiatus (preliminary note). Nat Sci 3:33–34
Hyman LH (1940) Observations and experiments on the physiology of medusae. Biol Bull 79:282–296
Jablonski D, Belanger CL, Berke SK, Huang S, Krug AZ, Roy K, Tomasovych A, Valentine JW (2013) Out of the tropics, but how? Fossils, bridge species, and thermal ranges in the dynamics of the marine latitudinal diversity gradient. P Natl Acad Sci USA 110:10487–10494
Jarms G, Tiemann H (1996) On a new hydropolyp without tentacles, Microhydrula limopsicola n. sp., epibiotic on bivalve shells from the Antarctic. Sci Mar 60:109–115
Johnston G (1835) Illustrations in British zoology. Mag Nat Hist J Zool Bot Miner Geol Meteorol 8:59–61
Johnston G (1847) A history of the British Zoophytes. Second Edition, Volume I. John Van Voorst, Paternoster Row, London
Kahn AS, Matsumoto GI, Hirano YM, Collins AG (2010) Haliclystus californiensis, a “new” species of stauromedusa (Cnidaria: Staurozoa) from the northeast Pacific, with a key to the species of Haliclystus. Zootaxa 2518:49–59
Kassianow N (1901) Studien über das Nervensystem der Lucernariden nebst sonstigen histologischen Beobachtungen über diese Gruppe. Z Wiss Zool 69:287–377
Kerswell AP (2006) Global biodiversity patterns of benthic marine algae. Ecology 87:2479–2488
Kikinger R, Salvini-Plawen LV (1995) Development from polyp to stauromedusa in Stylocoronella (Cnidaria: Scyphozoa). J Mar Biol Assoc UK 75:899–912
Kishinouye K (1893) Mushi-kurage, Depastrum inabai n. sp. Zool Mag 5:416–419
Kishinouye K (1899) Contributions to the natural history of the Commander Islands. XIII. A new species of stalked medusae, Haliclystus stejnegeri. Proc U S Nat Mus 22:125–129
Kishinouye K (1910) Some medusae of Japanese water. J Coll Science Imp Univ Tokyo 27:1–35
Kowalevsky AO (1884) Zur Entwicklungsgeschichte der Lucernaria. Zool Anz 7:712–719
Kramp PL (1914) Meduser og Siphonophorer. Catalogue of the Medusae and Siphonophora of Greenland. Meddel Grønland 23:381–456
Kramp PL (1943) The zoology of East Greenland. Medusae, Siphonophora and Ctenophora. Meddel Grønland 121:1–20
Kramp PL (1952) Reports on the Lund University Chile Expedition 1948–49. 2. Medusae collected by the Lund University Chile Expedition 1948–49. Lunds Univ Årsskrift 47:1–19
Kramp PL (1957) Medusae. B.A.N.Z. Antarct Res Exp Rep Ser B 6:151–164
Kramp PL (1961) Synopsis of the medusae of the world. J Mar Biol Assoc UK 40:292–303
Kume M, Dan K (1968) Invertebrate Embryology. Translated from Japanese by JC Dan. PROSVETA & NOLIT Publishing House, Belgrade
Lamouroux JVF (1815) Mémoire sur la Lucernaire campanulée. Mém Mus Hist Nat II:460–473
Larson RJ (1976) Marine flora and fauna of the northeastern United States. Cnidaria: Scyphozoa. NOAA Tech Rep NMFS 397:1–19
Larson RJ (1980) A new stauromedusa, Kishinouyea corbini (Scyphozoa, Stauromedusae) from the tropical western Atlantic. B Mar Sci 30:102–107
Larson RJ (1988) Kyopoda lamberti gen. nov., sp. nov., an atypical stauromedusa (Scyphozoa, Cnidaria) from the eastern Pacific, representing a new family. Can J Zool 66:2301–2303
Larson RJ, Fautin DG (1989) Stauromedusae of the genus Manania (= Thaumatoscyphus) (Cnidaria, Scyphozoa) in the northeast Pacific, including descriptions of new species Manania gwilliami and Manania handi. Can J Zool 67:1543–1549
Leitz T (1997) Induction of settlement and metamorphosis of cnidarian larvae: signals and signal transduction. Invertebr Reprod Dev 31:109–122
Lindberg DR (1991) Marine biotic interchange between the Northern and Southern Hemispheres. Paleobiology 17:308–324
Ling SW (1937) Studies on Chinese Stauromedusae. I. Stauromedusae from Tsingtao. Amoy Mar Biol Bull 3:1–35
López-González PJ, Gili J-M (2000) A new octocoral genus (Cnidaria: Anthozoa) from Antarctic waters. Polar Biol 23:452–458
Lutz RA, Collins AG, Annis ER, Reed AJ, Bennett KF, Halanych KM, Vrijenhoek RC (2006) Stauromedusan populations inhabiting deep-sea hydrothermal vents along the southern East Pacific Rise. Cah Biol Mar 47:409–413
Lutz RA, Desbruyères D, Shank TM, Vrijenhoek RC (1998) A deep-sea hydrothermal vent community dominated by Stauromedusae. Deep-Sea Res II 45:329–334
Marques AC, Collins AG (2004) Cladistic analysis of Medusozoa and cnidarian evolution. Invertebr Biol 123:32–42
Mayer AG (1910) Medusae of the world. Volume III. Scyphomedusae. Carnegie Institution publishing, Washington, publication 109
McInnes DE (1989) A stalked jellyfish (Stauromedusae) found at Black Rock, Port Phillip Bay. A first recording in Australia. Victorian Nat 106:86–92
Melo AS, Froehlich CG (2001) Evaluation of methods for estimating macroinvertebrate species richness using individual stones in tropical streams. Freshw Biol 46:711–721
Meyer A (1865) Über die Reproduktionskraft der Lucernarien. Amtl Ber 40 Verslag Deutsch Naturf Ärzte, Hannover 1865:217
Migot A (1922a) Sur le mode de fixation des Lucernaires à leur support. C R Soc Biol 86:827–829
Migot A (1922b) A propos de la fixation des Lucernaires. C R Soc Biol 87:151–153
Mills CE, Hirano YM (2007) Stauromedusae. In: Denny MW, Gaines SD (eds) Encyclopedia of tidepools and rocky shores. University of California Press, Berkeley, pp 541–543
Miranda LS, Branch GM, Collins AG, Hirano YM, Marques AC, Griffiths CL (2017) Stalked jellyfishes (Cnidaria: Staurozoa) of South Africa, with the description of Calvadosia lewisi sp. nov. Zootaxa 4227:369–389
Miranda LS, Collins AG, Hirano YM, Mills CE, Marques AC (2016b) Comparative internal anatomy of Staurozoa (Cnidaria), with functional and evolutionary inferences. PeerJ 4:e2594
Miranda LS, Collins AG, Marques AC (2010) Molecules clarify a cnidarian life cycle – the “hydrozoan” Microhydrula limopsicola is an early life stage of the staurozoan Haliclystus antarcticus. PLoS One 5:e10182
Miranda LS, Collins AG, Marques AC (2013) Internal anatomy of Haliclystus antarcticus (Cnidaria, Staurozoa) with a discussion on histological features used in staurozoan taxonomy. J Morph 274:1365–1383
Miranda LS, Haddad MA, Mills CE, Marques AC (2012b) Lucernariopsis capensis Carlgren, 1938 (Cnidaria, Staurozoa) in Brazil: first record outside its type locality in South Africa. Zootaxa 3158:60–64
Miranda LS, Hirano YM, Mills CE, Falconer A, Fenwick D, Marques AC, Collins AG (2016a) Systematics of stalked jellyfishes (Cnidaria: Staurozoa). PeerJ 4:e1951
Miranda LS, Marques AC (2016) Hidden impacts of the Samarco mining waste dam collapse to Brazilian marine fauna – an example from the staurozoans (Cnidaria). Biota Neotrop 16:e20160169
Miranda LS, Morandini AC, Marques AC (2009) Taxonomic review of Haliclystus antarcticus Pfeffer, 1889 (Stauromedusae, Staurozoa, Cnidaria), with remarks on the genus Haliclystus Clark, 1863. Polar Biol 32:1507–1519
Miranda LS, Morandini AC, Marques AC (2012a) Do Staurozoa bloom? A review of stauromedusan population biology. Hydrobiologia 690:57–67
Müller OF (1776) Zoologiae Danicae Prodromus, seu animalium Daniae et Norvegiae indigenarum, Characteres, Nomina et Synonyma imprimis popularium. Typis Hallageriis, Havniae
Müller WA, Leitz T (2002) Metamorphosis in the Cnidaria. Can J Zool 80:1755–1771
Nakamura M, Soberón J (2008) Use of approximate inference in an index of completeness of biological inventories. Conserv Biol 23:469–474
Naumov DV (1961) Stsifoidnye meduzy morei S.S.S.R. (scyphozoan medusae in the sea of U.S.S.R.) Akad Nauk SSSR, Opredeliteli po Faune SSSR 75:1–98
Oka A (1897) Sur une nouvelle espèce Japonaise du genre Lucernaria. Ann Zool Jap 1:141–145
Okamura B, Gruhl A, Bartholomew JL (2015) Myxozoan evolution, ecology and development. Springer International Publishing (Switzerland), Cham
Okubo T (1917) Preliminary note on a new genus of Stauromedusae from Hokkaido. Zool Mag 29:317–322
Otto JJ (1976) Early development and planula movement in Haliclystus (Scyphozoa, Stauromedusae). In: Mackie GO (ed) Coelenterate ecology and behavior. Plenum Press, New York, pp 319–329
Otto JJ (1978) The settlement of Haliclystus planulae. In: Chia FS, Rice M (eds) Settlement and metamorphosis of marine invertebrate larvae. Proceedings of the symposium on settlement and metamorphosis of marine invertebrate larvae, American Zoological Society Meeting, Canada. Elsevier, New York, pp 13–22
Panikkar NK (1944) Occurrence of a stauromedusa on the Indian coast. Curr Sci India 13:238–239
Petersen KW (1979) Development of coloniality in Hydrozoa. In: Larwood G, Rosen BR (eds) Biology and systematics of colonial animals. Academic Press, London, pp 105–139
Pfeffer G (1889) Zur Fauna von Süd-Georgien. Jahrb Hamburg Wiss Anst 6:37–55
Pisani V, Otero-Ferrer F, Lotto S, Maurel P, Goy J (2007) Lipkea ruspoliana Vogt, 1887 (Stauromedusae, Scyphozoa, Cnidaria) dans les aquariums du Musée Océanographique de Monaco. Bull Soc Zool Fr 132:183–190
Prell H (1910) Beiträge zur Kenntnis der Lebensweise einiger Pantopoden. Bergens Mus Aarbog 10:1–30
Redikorzev V (1925) Die Lucernose des Weiβen Meeres. Zool Anz 62:155–157
Russell ES (1904) Notes on Depastrum cyathiforme, Gosse. Ann Mag Nat Hist 13:62–65
Salovius S, Nyqvist M, Bonsdorff E (2005) Life in the fast lane: macrobenthos use temporary drifting algal habitats. J Sea Res 53:169–180
Salvini-Plawen LV (1966) Zur Kenntnis der Cnidaria des nordadriatischen Mesopsammon. Veröff Inst Meeresforsch Bremerh 2:165–186
Salvini-Plawen LV (1987) Mesopsammic Cnidaria from Plymouth (with systematic notes). J Mar Biol Assoc UK 67:623–637
Salvini-Plawen LV (2006) First record of a mature stauromedusa Stylocoronella (Cnidaria) in nature. Cah Biol Mar 47:219–222
Sars M (1846) Fauna littoralis Norvegiae oder Beschreibung und Abbildungen neuer oder wenig bekannter Seethiere, nebst Beobachtungen über die Organisation, Lebensweise u. Entwickelung derselben. Druck und Verlag von Johann Dahk, Christiania
Singla CL (1976) Ultrastructure and attachment of the basal disk of Haliclystus. In: Mackie GO (ed) Coelenterate ecology and behavior. Plenum Press, New York, pp 533–540
Spalding MD, Fox HE, Allen GR, Davidson N, Ferdaña ZA, Finlayson M, Halpern BS, Jorge MA, Lombana A, Lourie AS, Martin KD, McManus E, Molnar J, Recchia CA, Robertson J (2007) Marine ecoregions of the world: a bioregionalization of coast and shelf areas. Bioscience 57:573–583
Strathmann MF (1987) Reproduction and development of marine invertebrates of the northern Pacific coast. University of Washington Press, Seattle
Thiel M, Gutow L (2005) The ecology of rafting in the marine environment. II. The rafting organisms and community. Oceanogr Mar Biol 43:279–418
Tittensor DP, Mora C, Jetz W, Lotze HK, Ricard D, Berghe EV, Worm B (2010) Global patterns and predictors of marine biodiversity across taxa. Nature 466:1098–1101
Tronholm A, Leliaert F, Sansón M, Afonso-Carrillo J, Tyberghein L, Verbruggen H, Clerck D (2012) Contrasting geographical distributions as a result of thermal tolerance and long-distance dispersal in two allegedly widespread tropical brown algae. PLoS One 7:e30813
Uchida T (1929) Studies on the Stauromedusae and Cubomedusae, with special reference to their metamorphosis. Jap Jour Zool 2:103–193
Uchida T (1933) Eine neue Becherqualle aus Hokkaido. Proc Imp Acad 9:450–452
Uchida T (1973) The systematic position of the Stauromedusae. Publ Seto Mar Biol Lab 20:133–139
Uchida T, Hanaoka K-I (1933) On the morphology of a stalked medusa, Thaumatoscyphus distinctus Kishinouye. Jour Fac Science, Hokkaido Imp Univ, Series VI, Zool 2:135–153
Unterseher M, Schnittler M, Dormann C, Sickert A (2008) Application of species richness estimators for the assessment of fungal diversity. FEMS Microbiol Lett 282:205–213
Van Soest RWM, Boury-Esnault N, Vacelet J, Dohrmann M, Erpenbeck D, De Voogd NJ, Santodomingo N, Vanhoorne B, Kelly M, Hooper JNA (2012) Global diversity of sponges (Porifera). PLoS One 7:e35105
Vanhöffen E (1908) Die Lucernariden und Scyphomedusen der Deutschen Südpolar-Expedition 1901–1903. Deutsch Südp-Exp 10:25–49
Vogt C (1886) Sur une médusaire sessile, Lipkea ruspoliana. Arch Sci, Genève 16:356–362
Voight JR (2006) Stauromedusae on the East Pacific Rise. Cah Biol Mar 47:347–352
von Lendenfeld R (1884) The Schyphomedusae of the Southern Hemisphere. P Linn Soc N S W 9:155–169
Weill R (1934) Contribution à l’étude des cnidaires et de leurs nématocystes. I, II. Trav. Stn. Zool. Wimereux 10(11):1–701
Wichmann C-S, Hinojosa IA, Thiel M (2012) Floating kelps in Patagonian fjords: an important vehicle for rafting invertebrates and its relevance for biogeography. Mar Biol 159:2035–2049
Wietrzykowski W (1910) Sur le développement des Lucernaridés (note préliminaire). Arch Zool Exp Gen 5:10–27
Wietrzykowski W (1911) Seconde note sur le développement des Lucernaires. Arch Zool Exp Gen 6:49–52
Wietrzykowski W (1912) Recherches sur le développement des Lucernaires. Arch Zool Exp Gen 10:1–95
Willig MR, Kaufman DM, Stevens RD (2003) Latitudinal gradients of biodiversity: pattern, process, scale, and synthesis. Annu Rev Ecol Evol S 34:273–309
Zagal CJ (2004a) Population biology and habitat of the stauromedusa Haliclystus auricula in southern Chile. J Mar Biol Assoc UK 84:331–336
Zagal CJ (2004b) Diet of stauromedusa Haliclystus auricula from southern Chile. J Mar Biol Assoc UK 84:337–340
Zagal CJ (2008) Morphological abnormalities in the stauromedusa Haliclystus auricula (Cnidaria) and their possible causes. J Mar Biol Assoc UK 88:259–262
Zagal CJ, Hirano YM, Mills CE, Edgar GJ, Barrett NS (2011) New records of Staurozoa from Australian coastal waters, with a description of a new species of Lucernariopsis Uchida, 1929 (Cnidaria, Staurozoa, Stauromedusae) and a key to Australian Stauromedusae. Mar Biol Res 7:651–666
AcknowledgEments
We are grateful to David Fenwick for providing information about staurozoan species on the list of UK BAP priority marine species and to Sofia Stepanjants for providing the list of distributions for the Far East representatives of Stauromedusae. We are also grateful to Massimo Avian and Stefano Piraino whose comments helped improve the quality of the manuscript. This study was supported by: Fundação de Amparo à Pesquisa do Estado de São Paulo 2010/52324-6 (ACM), 2011/50242-5 (ACM), 2013/50484-4 (ACM), 2010/07362-7 (LSM), 2015/23695-0 (LSM); Conselho Nacional de Desenvolvimento Científico e Tecnológico 474672/2007-7 (ACM), 563106/2010-7 (ACM), 562143/2010-6 (ACM), 477156/2011-8 (ACM), 305805/2013-4 (ACM), 445444/2014-2 (ACM), 142270/2010-5 (LSM), 165066/2014-8 (LSM); and Comissão de Aperfeiçoamento de Pessoal de Nível Superior PDSE 16499/12-3 (LSM), PNPD (LSM).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Piraino
Electronic supplementary material
Online Resource 1
(PDF 199 kb)
Online Resource 2
(PDF 181 kb)
Online Resource 3
(PDF 184 kb)
Online Resource 4
(PDF 928 kb)
Online Resource 5
(PDF 988 kb)
Online Resource 6
(PDF 954 kb)
Online Resource 7
(PDF 949 kb)
Online Resource 8
(PDF 999 kb)
Online Resource 9
(PDF 945 kb)
Online Resource 10
(PDF 983 kb)
Online Resource 11
(PDF 945 kb)
Online Resource 12
(PDF 953 kb)
Online Resource 13
(PDF 982 kb)
Online Resource 14
(PDF 948 kb)
Rights and permissions
About this article

Cite this article
Miranda, L.S., Mills, C.E., Hirano, Y.M. et al. A review of the global diversity and natural history of stalked jellyfishes (Cnidaria, Staurozoa). Mar Biodiv 48, 1695–1714 (2018). https://doi.org/10.1007/s12526-017-0721-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12526-017-0721-4