
Abstract
Colon and intestinal crypts have been widely chosen to study cell dynamics because of their fairly simple structures. In the colon and intestinal crypts, stem cells (SCs) are located at very bottom of the crypt, fully differentiated cells (FDs) are located in the top of the crypt, and transit-amplifying cells (TAs) are in the middle of the crypt between FDs and SCs. Recently, it has been discovered that there are two types of stem cells in the intestinal crypts: central stem cells (CeSCs) and border stem cells. To investigate dynamics of mutants in colon and intestinal crypts, we develop a four-compartmental stochastic model, which includes two SC compartments, and TAs and FDs compartments. We calculate the probability of the progeny of marked or mutant cells located at each of these compartments taking over the entire crypt or being washed out from the crypt. We found that the progeny of CeSCs will take over the entire crypt with a probability close to one. Interestingly, the progeny of advantageous mutant TAs and FDs will be washed out faster than disadvantageous mutants. Saliently, the model predicts that the time that the progeny of wild-type central stem cells will take over the mouse intestinal crypt is around 60 days, which is in perfect agreement with an experimental observation.









Similar content being viewed by others
References
Bach SP (2000) Stem cells: the intestinal stem cell as a paradigm. Carcinogenesis 21(3):469–476
Baker AM, Cereser B, Melton S, Fletcher AG, Rodriguez-Justo M et al (2014) Quantification of crypt and stem cell evolution in the normal and neoplastic human colon. Cell Rep 8(4):940–947
Bollas A, Shahriyari L (2017) The role of backward cell migration in two-hit mutants’ production in the stem cell niche. PLoS ONE 12(9):e0184651
Bravo R, Axelrod DE (2013) A calibrated agent-based computer model of stochastic cell dynamics in normal human colon crypts useful for in silico experiments. Theor Biol Med Model 10(1):66
Buske P, Galle J, Barker N, Aust G, Clevers H, Loeer M (2011) A comprehensive model of the spatio-temporal stem cell and tissue organisation in the intestinal crypt. PLoS Comput Biol 7(1):e1001045
Cassimerisz L, G. Plopper G, Lingappa VR (2011) Lewin’s cells. Jones and Bartlett Publishers, Sudbury
D’Onofrio A, Tomlinson IPM (2007) A nonlinear mathematical model of cell turnover, differentiation and tumorigenesis in the intestinal crypt. J Theor Biol 244(3):367–74
Durrett R, Foo J, Leder K (2016) Spatial Moran models, II: cancer initiation in spatially structured tissue. J Math Biol 72(5):1369–1400
El Zoghbi M, Cummings LC (2016) New era of colorectal cancer screening. World J Gastrointest Endosc 8(5):252
Fletcher AG, Breward CJW, Chapman SJ (2012) Mathematical modeling of monoclonal conversion in the colonic crypt. J Theor Biol 300:118–133
Johnston MD, Edwards CM, Bodmer WF, Maini PK, Chapman SJ (2007) Mathematical modeling of cell population dynamics in the colonic crypt and in colorectal cancer. Proc Natl Acad Sci USA 104(10):4008–13
Kagawa Y, Horita N, Taniguchi H, Tsuneda S (2014) Modeling of stem cell dynamics in human colonic crypts in silico. J Gastroenterol 49(2):263–269
Kinzler KW, Vogelstein B (1996) Lessons from hereditary colorectal cancer. Cell 87(2):159–170
Komarova NL, Shahriyari L, Wodarz D (2014) Complex role of space in the crossing of fitness valleys by asexual populations. J R Soc Interface 11(95):20140014
Korem Y, Szekely P, Hart Y, Sheftel H, Hausser J et al (2015) Geometry of the gene expression space of individual cells. PLoS Comput Biol 11(7):e1004224
Kozar S, Morrissey E, Nicholson AM, van der Heijden M, Zecchini HI, Kemp R, Tavare S, Vermeulen L, Winton DJ (2013) Continuous clonal labeling reveals small numbers of functional stem cells in intestinal crypts and adenomas. Cell Stem Cell 13(5):626–633
Liu X, Johnson S, Liu S, Kanojia D, Yue W, Singn U et al (2013) Nonlinear growth kinetics of breast cancer stem cells: implications for cancer stem cell targeted therapy. Sci Rep 3:2473
Mahdipour-Shirayeh A, Kaveh K, Kohandel M, Sivaloganathan S (2017) Phenotypic heterogeneity in modeling cancer evolution. PLoS ONE 12(10):e0187000
Mahdipour-Shirayeh A, Darooneh AH, Long AD, Komarova NL, Kohandel M (2017) Genotype by random environmental interactions gives an advantage to non-favored minor alleles. Sci Rep 7:5193
Mirams GR, Fletcher AG, Maini PK, Byrne HM (2012) A theoretical investigation of the effect of proliferation and adhesion on monoclonal conversion in the colonic crypt. J Theor Biol 312:143–156
Nowak MA, Michor F, Iwasa Y (2003) The linear process of somatic evolution. Proc Natl Acad Sci 100(25):14966–14969
Olive KP, Tuveson DA, Ruhe ZC, Yin B, Willis NA, Bronson RT et al (2004) Mutant p53 gain of function in two mouse models of Li–Fraumeni syndrome. Cell 119(6):847–860
Pin C, Watson AJM, Carding SR (2012) Modelling the spatio-temporal cell dynamics reveals novel insights on cell differentiation and proliferation in the small intestinal crypt. PLoS ONE 7(5):e37115
Potten CS, Kellett M, Roberts SA, Rew DA, Wilson GD (1992) Measurement of in vivo proliferation in human colorectal mucosa using bromodeoxyuridine. Gut 33(1):71–78
Potten CS, Kellett M, Rew DA, Roberts SA (1992) Proliferation in human gastrointestinal epithelium using bromodeoxyuridine in vivo: data for different sites, proximity to a tumour, and polyposis coli. Gut 33(4):524–529
Ritsma L, Ellenbroek SIJ, Zomer A, Snippert HJ, de Sauvage FJ et al (2014) Intestinal crypt homeostasis revealed at single-stem-cell level by in vivo live imaging. Nature 507(7492):362–365
Rodriguez-Brenes IA, Komarova NL, Wodarz D (2011) Evolutionary dynamics of feedback escape and the development of stem-cell-driven cancers. Proc Natl Acad Sci 108(47):18983–18988
Sansom OS, Reed KR, Hayes AJ, Ireland H, Brinkmann H et al (2004) Loss of Apc in vivo immediately perturbs Wnt signaling, differentiation, and migration. Genes Dev 18(12):1385–1390
Shahriyari L (2017) Cell dynamics in tumour environment after treatments. J R Soc Interface 14(127):20160977
Shahriyari L, Komarova NL (2013) Symmetric vs. asymmetric stem cell divisions: an adaptation against cancer? PLoS ONE 8(10):e76195
Shahriyari L, Komarova NL (2015) The role of the bi-compartmental stem cell niche in delaying cancer. Phys Biol 12(5):055001
Shahriyari L, Mahdipour-Shirayeh A (2017) Modeling dynamics of mutants in heterogeneous stem cell niche. Phys Biol 14(1):016004
Shahriyari L, Komarova NL, Jilkine A (2016) The role of cell location and spatial gradients in the evolutionary dynamics of colon and intestinal crypts. Biol Direct 11(1):42
Snippert HJ, van der Flier LG, Sato T (2010) Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lgr5 stem cells. Cell 143(1):134–144
Traulsen A, Pacheco JM, Luzzatto L, Dingli D (2010) Somatic mutations and the hierarchy of hematopoiesis. BioEssays 32(11):1003–1008
Vermeulen L, Snippert HJ (2014) Stem cell dynamics in homeostasis and cancer of the intestine. Nat Rev Cancer 14(7):468–480
Vermeulen L, Morrissey E, van der Heijden M, Nicholson AM, Sottoriva A et al (2013) Defining stem cell dynamics in models of intestinal tumor initiation. Science 342(6161):995–998
Werner B, Dingli D, Traulsen A (2013) A deterministic model for the occurrence and dynamics of multiple mutations in hierarchically organized tissues. J R Soc Interface 10(85):20130349–20130349
Wodarz D, Komarova NL (2014) Dynamics of cancer: mathematical foundations of oncology. World Scientific Publishing Company, Singapore
Yatabe Y, Tavare S, Shibata D (2001) Investigating stem cells in human colon by using methylation patterns. Proc Natl Acad Sci 98(19):10839–10844
Zhao R, Michor F (2013) Patterns of proliferative activity in the colonic crypt determine crypt stability and rates of somatic evolution. PLoS Comput Biol 9(6):e1003082
Funding
This research has been supported in part by the Mathematical Biosciences Institute and the National Science Foundation under grant DMS 0931642.
Author information
Authors and Affiliations
Contributions
AMS carried out the analytic calculations, participated in data analysis, participated in the design of the study and writing the manuscript; LS conceived of the study, designed the study, coordinated the study, participated in data analysis, carried out the numerical simulations, obtained parameters’ values, and wrote the manuscript. All authors gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Appendices
Appendix
Materials and Methods
In this section, using the methods described in Shahriyari et al. (2016) and Wodarz and Komarova (2014), a generalized framework will be constructed to investigate cell dynamics in the colonic/intestinal crypt. In this model, we have a four-dimensional multivariable Markov model as the system of random movements over possible states \((e^{*}, b^{*}, d^{*}, f^{*})\) for \(0\le e^{*}\le |S_c|,0\le b^{*}\le |S_b|, 0\le d^{*}\le |D_t|,\) and \(0\le f^{*}\le |D_f|\). The total number of states is \((|S_c|+1)(|S_b|+1)(|D_t|+1)(|D_f|+1)\), where \(|S_c|, \ |S_b|, \ |D_t|, \ |D_f|\) are the sizes of \(S_c, \ S_b, \ D_t, \ D_f\) compartments, respectively.
We denote the probability of moving from the state a to the state b in one time step by \(P_{a \rightarrow b}\), where \(a,b\in \{(e^{*}, b^{*}, d^{*}, f^{*})\}\). For simplicity, indexes a and b only include the parameter(s), which are changing. For example, the probability \(P_{e^{*} \rightarrow e^{*}+1}\) is the probability of moving from the state, which has \(e^{*}\) number of \(S_c\) mutants, to the state that has \(e^{*}+1\) number of \(S_c\) mutants in one time step, while the number of the other mutants (i.e., \(b^{*}, d^{*}, f^{*}\)) is not changed. All possible nonzero transition probabilities are listed below.
1.1 A. Transition Probabilities
-
(1)
\(P_{{f}^{*}\rightarrow {f}^{*}+1} = \left( \frac{f}{f+ f^{*}} \right) ^{2} \left\{ 2\, \lambda _{f} \left( \frac{f}{\mathcal {F}} \right) \left( \frac{r_1\,f^{*}}{\mathcal {F}} \right) \right\} +\)
\( \frac{2 f f^{*}}{(f+ f^{*})^2} \left\{ \lambda _f \left[ \frac{r_1\,f^{*}}{\mathcal{F}} \right] ^2 + (1-\lambda _f) \frac{r_1 d^{*}}{\mathcal{D}} \left[ (1-\lambda _s)\frac{r_1 d^{*}}{\mathcal{D}} + \lambda _s\,(1-\sigma )\frac{r_1 b^{*}}{\mathcal{R}_b} \right] \ \right\} , \)
-
(2)
\(P_{f^{*}\rightarrow f^{*}-1} = \frac{2 f f^{*}}{(f+ f^{*})^2} \,\left\{ \lambda _f \, \left( \frac{f}{\mathcal{F}} \right) ^2 + (1-\lambda _f)\,\frac{d}{\mathcal{D}} \left[ (1-\lambda _s)\, \frac{d}{\mathcal{D}} + \lambda _s\,(1-\sigma )\,\frac{b}{\mathcal{R}_b} + \lambda _s\,\sigma \left( \delta \frac{b}{\mathcal{R}_b} \right. \right. \right. \)
\(\left. \left. \left. + (1-\delta )(1-\gamma )\frac{b}{\mathcal{R}_b} \left( (1-\alpha ) + \alpha \, \frac{b}{b+b^{*}}\, \frac{e}{e+e^{*}} + \alpha \, \frac{b^{*}}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} \right) + (1-\delta )\gamma \,\frac{e}{\mathcal{R}_c}\frac{e}{e+e^{*}}\right) \right] \right\} \)
\( + \left( \frac{f^{*}}{f+ f^{*}} \right) ^2 \left\{ 2\,\lambda _f \left( \frac{f}{\mathcal{F}} \right) \left( \frac{r_1 f^{*}}{\mathcal{F}} \right) \right\} , \)
-
(3)
\(P_{f^{*}\rightarrow f^{*}+2}= \left( \frac{f}{f+ f^{*}} \right) ^2 \,\left\{ \lambda _f \, \left[ \frac{r_1\,f^{*}}{\mathcal{F}} \right] ^2 + (1-\lambda _f)\,\,\frac{r_1 d^{*}}{\mathcal{D}} \left[ (1-\lambda _s)\,\frac{r_1 d^{*}}{\mathcal{D}} + \lambda _s\,(1-\sigma )\,\frac{r_1 b^{*}}{\mathcal{R}_b} \right] \right\} ,\)
-
(4)
\(P_{f^{*}\rightarrow f^{*}-2}=\left( \frac{f^{*}}{f+ f^{*}} \right) ^2 \,\left\{ \lambda _f \, \left( \frac{f}{\mathcal{F}} \right) ^2 + (1-\lambda _f)\,\frac{d}{\mathcal{D}} \left[ (1-\lambda _s)\, \frac{d}{\mathcal{D}} + \lambda _s\,(1-\sigma )\,\frac{b}{\mathcal{R}_b} + \lambda _s\,\sigma \left( \delta \frac{b}{\mathcal{R}_b} \right. \right. \right. \)
\(\left. \left. \left. + (1-\delta )(1-\gamma )\frac{b}{\mathcal{R}_b} \left( (1-\alpha ) + \alpha \, \frac{b}{b+b^{*}}\,\frac{e}{e+e^{*}} + \alpha \, \frac{b^{*}}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} \right) + (1-\delta )\gamma \,\frac{e}{\mathcal{R}_c}\frac{e}{e+e^{*}}\right) \right] \right\} , \)
-
(5)
\(P_{d^{*},f^{*}\rightarrow {d}^{*}-1,f^{*}+2}=\left( \frac{f}{f+ f^{*}} \right) ^{2}\,\left\{ (1-\lambda _{f})\,\frac{r_{1}\, {d}^{*}}{\mathcal {D}} \, \left[ (1-\lambda _s)\, \frac{d}{\mathcal {D}} + \lambda _s\,(1-\sigma )\,\frac{b}{\mathcal {R}_{b}} + \lambda _{s}\,\sigma \,\left( \delta \,\frac{b}{\mathcal {R}_{b}} \right. \right. \right. \)
\(\left. \left. + \,(1-\delta )(1-\gamma )\frac{b}{\mathcal{R}_b} \left( (1-\alpha ) + \alpha \, \frac{b}{b+b^{*}}\,\frac{e}{e+e^{*}} + \alpha \, \frac{b^{*}}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} + (1-\delta )\gamma \,\frac{e}{\mathcal{R}_c}\frac{e}{e+e^{*}} \right) \right] \right\} ,\)
-
(6)
\(P_{d^{*},{f}^{*}\rightarrow {d}^{*}-1,{f}^{*}+1} = \frac{2 f {f}^{*}}{(f+ {f}^{*})^{2}} \,\left\{ (1-\lambda _f)\,\frac{r_{1}\, {d}^{*}}{\mathcal {D}} \left[ (1-\lambda _s)\, \frac{d}{\mathcal {D}} + \lambda _{s}\,(1-\sigma )\,\frac{b}{\mathcal {R}_{b}} + \lambda _{s}\,\sigma \,\left( \delta \,\frac{b}{\mathcal {R}_b} \right. \right. \right. \)
\( \left. \left. +\, (1-\delta )(1-\gamma )\frac{b}{\mathcal{R}_b} \left( (1-\alpha ) + \alpha \, \frac{b}{b+b^{*}}\,\frac{e}{e+e^{*}} + \alpha \, \frac{b^{*}}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} + (1-\delta )\gamma \,\frac{e}{\mathcal{R}_c}\frac{e}{e+e^{*}} \right) \right] \right\} ,\)
-
(7)
\(P_{d^{*}\rightarrow d^{*}+1}= \left( \frac{f}{f + f^{*}} \right) ^2 \, \left\{ (1-\lambda _f)\,\frac{d}{\mathcal{D}}\, \left[ (1-\lambda _s)\,\frac{r_1 d^{*}}{\mathcal{D}} + \lambda _s\,(1-\sigma )\,\frac{r_1 b^{*}}{\mathcal{R}_b} \right] \right\} ,\)
-
(8)
\(P_{d^{*},f^{*}\rightarrow d^{*}+1,f^{*}-1}= \frac{ 2 f f^{*}}{(f+ f^{*})^2} \, \left\{ (1-\lambda _f)\,\frac{d}{\mathcal{D}}\, \left[ (1-\lambda _s)\,\frac{r_1 d^{*}}{\mathcal{D}} + \lambda _s\,(1-\sigma )\,\frac{r_1 b^{*}}{\mathcal{R}_b} \right] \right\} ,\)
-
(9)
\(P_{d^{*},f^{*}\rightarrow d^{*}+1, f^{*}-2}= \left( \frac{f^{*}}{f + f^{*}} \right) ^2 \, \left\{ (1-\lambda _f)\,\frac{d}{\mathcal{D}}\, \left[ (1-\lambda _s)\,\frac{r_1 d^{*}}{\mathcal{D}} + \lambda _s\,(1-\sigma )\,\frac{r_1 b^{*}}{\mathcal{R}_b} \right] \right\} ,\)
-
(10)
\(P_{b^{*},d^{*},f^{*}\rightarrow b^{*}-1,d^{*}+2, f^{*}-2} = \left( \frac{f^{*}}{f + f^{*}} \right) ^2 \, \left\{ (1-\lambda _f)\,\frac{d}{\mathcal{D}}\, \left[ \lambda _s\,\sigma \,\delta \,\frac{r_1 b^{*}}{\mathcal{R}_b} \right] \right\} ,\)
-
(11)
\(P_{b^{*},d^{*},f^{*}\rightarrow b^{*}-1,d^{*}+2, f^{*}-1}=\frac{ 2 f f^{*}}{(f+ f^{*})^2} \,\left\{ (1-\lambda _f)\,\frac{d}{\mathcal{D}}\, \left[ \lambda _s\,\sigma \,\delta \,\frac{r_1 b^{*}}{\mathcal{R}_b} \right] \right\} ,\)
-
(12)
\(P_{d^{*}\rightarrow {d}^{*}-1} = \left( \frac{f^{*}}{f+ {f}^{*}} \right) ^2 \, \left\{ (1-\lambda _{f})\,\frac{r_{1}\, {d}^{*}}{\mathcal {D}} \left[ (1-\lambda _{s})\, \frac{d}{{\mathcal {D}}} + \lambda _{s}\,(1-\sigma )\,\frac{b}{{\mathcal {R}}_{b}} + \lambda _{s}\,\sigma \,\left( \delta \,\frac{b}{{\mathcal {R}}_{b}}\right. \right. \right. \)
\( \left. \left. + \,(1-\delta )(1-\gamma )\frac{b}{\mathcal{R}_b} \left( (1-\alpha ) + \alpha \, \frac{b}{b+b^{*}}\,\frac{e}{e+e^{*}} + \alpha \, \frac{b^{*}}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} + (1-\delta )\gamma \,\frac{e}{\mathcal{R}_c}\frac{e}{e+e^{*}} \right) \right] \right\} ,\)
-
(13)
\(P_{b^{*},d^{*} \rightarrow b^{*}-1,d^{*}+2} = \left( \frac{f}{f+ f^{*}} \right) ^2\, \left\{ (1-\lambda _f)\,\frac{d}{\mathcal{D}}\, \left[ \lambda _s\,\sigma \delta \,\frac{r_1 b^{*}}{\mathcal{R}_b} \right] \right\} ,\)
-
(14)
\(P_{b^{*},d^{*}, \rightarrow b^{*}+1,d^{*}-1} = \left( \frac{f^{*}}{f+ f^{*}} \right) ^2\, \left\{ (1-\lambda _f)\,\frac{r_1 d^{*}}{\mathcal{D}} \left[ \lambda _s\,\sigma \,(1-\delta )\,\left( (1-\gamma )\,\frac{r_1 b^{*}}{\mathcal{R}_b} \left( (1-\alpha ) \right. \right. \right. \right. \)
\(\left. \left. \left. \left. +\alpha \, \frac{b^{*}}{b+b^{*}}\,\frac{e^{*}}{e+e^{*}} +\alpha \, \frac{b}{b+b^{*}}\,\frac{e}{e+e^{*}} \right) + \gamma \,\frac{r_1 e^{*}}{\mathcal{R}_c}\,\frac{e^{*}}{e+e^{*}} \right) \right] \right\} ,\)
-
(15)
\(P_{b^{*},d^{*}\rightarrow b^{*}-1,d^{*}+1} = \left( \frac{f^{*}}{f+ f^{*}} \right) ^2\, \left\{ (1-\lambda _f)\,\frac{r_1 d^{*}}{\mathcal{D}} \left[ \lambda _s\,\sigma \,\delta \,\frac{r_1 b^{*}}{\mathcal{R}_b} \right] \right\} ,\)
-
(16)
\(P_{b^{*}\rightarrow b^{*}+1} = \left( \frac{f}{f+ f^{*}} \right) ^2\, \left\{ (1-\lambda _f)\,\frac{d}{\mathcal{D}}\,\left[ \lambda _s\,\sigma \,(1-\delta )\,\left( (1-\gamma )\,\frac{r_1 b^{*}}{\mathcal{R}_b} \left( (1-\alpha ) +\alpha \, \frac{b^{*}}{b+b^{*}}\,\frac{e^{*}}{e+e^{*}} \right. \right. \right. \right. \)
\(\left. \left. \left. \left. +\alpha \, \frac{b}{b+b^{*}}\,\frac{e}{e+e^{*}} \right) + \gamma \,\frac{r_1 e^{*}}{\mathcal{R}_c}\,\frac{e^{*}}{e+e^{*}} \right) \right] \right\} ,\)
-
(17)
\(P_{b^{*},f^{*}\rightarrow b^{*}+1,f^{*}-1} = \frac{2 f f^{*}}{(f+ f^{*})^2} \, \left\{ (1-\lambda _f)\,\frac{d}{\mathcal{D}}\,\left[ \lambda _s\,\sigma \,(1-\delta )\,\left( (1-\gamma )\,\frac{r_1 b^{*}}{\mathcal{R}_b} \left( (1-\alpha ) \right. \right. \right. \right. \)
\(\left. \left. \left. \left. +\,\alpha \, \frac{b^{*}}{b+b^{*}}\,\frac{e^{*}}{e+e^{*}} +\alpha \, \frac{b}{b+b^{*}}\,\frac{e}{e+e^{*}} \right) + \gamma \,\frac{r_1 e^{*}}{\mathcal{R}_c}\,\frac{e^{*}}{e+e^{*}} \right) \right] \right\} ,\)
-
(18)
\(P_{b^{*},f^{*}\rightarrow b^{*}+1,f^{*}-2} = \left( \frac{f^{*}}{f+ f^{*}} \right) ^2\, \left\{ (1-\lambda _f)\,\frac{d}{\mathcal{D}}\,\left[ \lambda _s\,\sigma \,(1-\delta )\,\left( (1-\gamma )\,\frac{r_1 b^{*}}{\mathcal{R}_b} \left( (1-\alpha ) \right. \right. \right. \right. \)
\(\left. \left. \left. \left. +\,\alpha \, \frac{b^{*}}{b+b^{*}}\,\frac{e^{*}}{e+e^{*}} +\alpha \, \frac{b}{b+b^{*}}\,\frac{e}{e+e^{*}} \right) + \gamma \,\frac{r_1 e^{*}}{\mathcal{R}_c}\,\frac{e^{*}}{e+e^{*}} \right) \right] \right\} ,\)
-
(19)
\(P_{e^{*}\rightarrow e^{*}+1} = \left( \frac{f}{f+ f^{*}} \right) ^2\, \left\{ (1-\lambda _f)\,\frac{d}{\mathcal{D}}\, \left[ \lambda _s\,\sigma \,(1-\delta )\,\left( (1-\gamma )\,\frac{r_1 b^{*}}{\mathcal{R}_b} \, \alpha \, \frac{b^{*}}{b+b^{*}}+ \gamma \,\frac{r_1 e^{*}}{\mathcal{R}_c} \right) \,\frac{e}{e+e^{*}} \right] \right\} ,\)
-
(20)
\(P_{d^{**},f^{**}\rightarrow d^{**}-1,f^{**}+2}= \left( \frac{f}{f+ f^{*}} \right) ^2\,\left\{ (1-\lambda _f)\, \frac{r_2 d^{**}}{\mathcal{D}}\, \left[ (1-\lambda _s)\, \frac{d}{\mathcal{D}} + \lambda _s\,(1-\sigma )\,\frac{b}{\mathcal{R}_b} \right. \right. \)
\( \,\,+ \,\lambda _s\,\sigma \,\left( \delta \,\frac{b}{\mathcal{R}_b} + (1-\delta )(1-\gamma )\frac{b}{\mathcal{R}_b} \left( (1-\alpha ) + \alpha \, \frac{b}{b+b^{*}}\,\frac{e}{e+e^{*}} + \alpha \, \frac{b^{*}}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} \right) \right. \)
\(\left. \left. \left. + \,(1-\delta )\gamma \,\frac{e}{\mathcal{R}_c}\frac{e}{e+e^{*}} \right) \right] \right\} ,\)
-
(21)
\(P_{e^{*}, d^{*}\rightarrow e^{*}+1,d^{*}-1} = \left( \frac{f^{*}}{f+ f^{*}} \right) ^2 \!\! \left\{ (1-\lambda _f) \frac{r_1 d^{*}}{\mathcal{D}} \left[ \lambda _s\sigma (1-\delta ) \left( (1-\gamma ) \frac{r_1 b^{*}}{\mathcal{R}_b} \alpha \frac{b^{*}}{b+b^{*}}+ \gamma \frac{r_1 e^{*}}{\mathcal{R}_c} \right) \,\frac{e}{e+e^{*}} \right] \right\} ,\)
-
(22)
\(P_{e^{*},b^{*}\rightarrow e^{*}-1, b^{*}+1} = \left( \frac{f}{f+ f^{*}} \right) ^2\!\! \left\{ (1-\lambda _f)\, \frac{d}{\mathcal{D}}\! \left[ \lambda _s\,\sigma \,(1-\delta )\,\left( (1-\gamma )\,\frac{b}{\mathcal{R}_b}\,\alpha \, \frac{b}{b+b^{*}} + \gamma \,\frac{e}{\mathcal{R}_c} \! \right) \, \frac{e^{*}}{e + e^{*}} \! \right] \right\} ,\)
-
(23)
\(P_{e^{*},b^{*}, d^{*}\rightarrow e^{*}-1, b^{*}+1,d^{*}-1} = \left( \! \frac{f^{*}}{f+ f^{*}} \! \right) ^2\!\! \left\{ (1-\lambda _f) \frac{r_1 d^{*}}{\mathcal{D}} \! \left[ \lambda _s \sigma (1-\delta ) \! \left( (1-\gamma ) \frac{b}{\mathcal{R}_b} \alpha \frac{b}{b+b^{*}} + \gamma \,\frac{e}{\mathcal{R}_c} \right) \! \frac{e^{*}}{e + e^{*}} \! \right] \!\right\} ,\)
-
(24)
\(P_{b^{*},d^{*},f^{*}\rightarrow b^{*}-1,d^{*}+1,f^{*}+2}= \left( \! \frac{f}{f+ f^{*}} \! \right) ^2\, \left\{ (1-\lambda _f)\,\frac{r_1 d^{*}}{\mathcal{D}} \,\left[ \lambda _s\,\sigma \,\delta \, \frac{r_1 b^{*}}{\mathcal{R}_b} \right] \right\} ,\)
-
(25)
\(P_{b^{*},d^{*},f^{*}\rightarrow b^{*}-1,d^{*}+1,f^{*}+1} = \frac{2 f f^{*}}{(f+ f^{*})^2} \, \left\{ (1-\lambda _f)\,\frac{r_1 d^{*}}{\mathcal{D}} \,\left[ \lambda _s\,\sigma \,\delta \, \frac{r_1 b^{*}}{\mathcal{R}_b} \right] \right\} ,\)
-
(26)
\(P_{b^{*},d^{*}, f^{*}\rightarrow b^{*}+1,d^{*}-1, f^{*}+1} = \frac{2 f f^{*}}{(f+ f^{*})^2} \,\left\{ (1-\lambda _f)\,\frac{r_1 d^{*}}{\mathcal{D}} \left[ \lambda _s\,\sigma \,(1-\delta )\,\left( (1-\gamma )\,\frac{r_1 b^{*}}{\mathcal{R}_b} \left( (1-\alpha ) \right. \right. \right. \right. \)
\(\left. \left. \left. \left. +\,\alpha \, \frac{b^{*}}{b+b^{*}}\,\frac{e^{*}}{e+e^{*}} +\alpha \, \frac{b}{b+b^{*}}\,\frac{e}{e+e^{*}} \right) + \gamma \,\frac{r_1 e^{*}}{\mathcal{R}_c}\,\frac{e^{*}}{e+e^{*}} \right) \right] \right\} ,\)
-
(27)
\(P_{b^{*},d^{*}, f^{*}\rightarrow b^{*}+1,d^{*}-1, f^{*}+2} = \left( \frac{f}{f+ f^{*}} \right) ^2\, \left\{ (1-\lambda _f)\,\frac{r_1 d^{*}}{\mathcal{D}} \left[ \lambda _s\,\sigma \,(1-\delta )\,\left( (1-\gamma )\,\frac{r_1 b^{*}}{\mathcal{R}_b} \left( (1-\alpha ) \right. \right. \right. \right. \)
\(\left. \left. \left. \left. +\,\alpha \, \frac{b^{*}}{b+b^{*}}\,\frac{e^{*}}{e+e^{*}} +\alpha \, \frac{b}{b+b^{*}}\,\frac{e}{e+e^{*}} \right) + \gamma \,\frac{r_1 e^{*}}{\mathcal{R}_c}\,\frac{e^{*}}{e+e^{*}} \right) \right] \right\} ,\)
-
(28)
\(P_{e^{*},b^{*},d^{*}, f^{*}\rightarrow e^{*}-1, b^{*}+1,d^{*}-1,f^{*}+1} \)
\(=\frac{2 f f^{*}}{(f+ f^{*})^2} \, \left\{ (1-\lambda _f)\, \frac{r_1 d^{*}}{\mathcal{D}} \left[ \lambda _s\,\sigma \,(1-\delta )\,\left( (1-\gamma )\,\frac{b}{\mathcal{R}_b}\,\alpha \, \frac{b}{b+b^{*}} + \gamma \,\frac{e}{\mathcal{R}_c} \right) \, \frac{e^{*}}{e + e^{*}} \right] \right\} ,\)
-
(29)
\(P_{e^{*},b^{*}, d^{*}, f^{*}\rightarrow e^{*}-1, b^{*}+1,d^{*}-1,f^{*}+2} \)
\(=\left( \frac{f}{f+ f^{*}} \right) ^2\, \left\{ (1-\lambda _f)\, \frac{r_1 d^{*}}{\mathcal{D}}\left[ \lambda _s\,\sigma \,(1-\delta )\,\left( (1-\gamma )\,\frac{b}{\mathcal{R}_b}\,\alpha \, \frac{b}{b+b^{*}} + \gamma \,\frac{e}{\mathcal{R}_c} \right) \, \frac{e^{*}}{e + e^{*}} \right] \right\} ,\)
-
(30)
\(P_{e^{*},f^{*}\rightarrow e^{*}+1, f^{*}-1} \! = \! \frac{2 f f^{*}}{(f+ f^{*})^2} \! \left\{ (1-\lambda _f) \frac{d}{\mathcal{D}}\! \left[ \lambda _s\,\sigma \,(1-\delta )\! \left( (1-\gamma ) \frac{r_1\,b^{*}}{\mathcal{R}_b} \alpha \frac{b^{*}}{b+b^{*}} + \gamma \frac{r_1\,e^{*}}{\mathcal{R}_c} \right) \frac{e}{e + e^{*}} \right] \right\} ,\)
-
(31)
\(P_{e^{*},f^{*}\rightarrow e^{*}+1, f^{*}-2} \!=\! \left( \! \frac{f^{*} }{f+ f^{*}} \! \right) ^2\! \left\{ (1-\lambda _f)\, \frac{d}{\mathcal{D}}\! \left[ \! \lambda _s \sigma (1-\delta ) \left( (1-\gamma ) \frac{r_1\,b^{*}}{\mathcal{R}_b} \alpha \frac{b^{*}}{b+b^{*}} + \gamma \frac{r_1\,e^{*}}{\mathcal{R}_c} \right) \! \frac{e}{e + e^{*}} \! \right] \right\} ,\)
-
(32)
\(P_{e^{*},d^{*},f^{*}\rightarrow e^{*}+1,d^{*}-1, f^{*}+1} \)
\(=\frac{2 f f^{*}}{(f+ f^{*})^2} \,\left\{ (1-\lambda _f)\, \frac{r_1\,d^{*}}{\mathcal{D}}\,\left[ \lambda _s\,\sigma \,(1-\delta )\, \left( (1-\gamma )\,\frac{r_1\,b^{*}}{\mathcal{R}_b}\,\alpha \, \frac{b^{*}}{b+b^{*}} + \gamma \,\frac{r_1\,e^{*}}{\mathcal{R}_c} \right) \, \frac{e}{e + e^{*}} \right] \right\} ,\)
-
(33)
\(P_{e^{*},d^{*},f^{*}\rightarrow e^{*}+1,d^{*}-1, f^{*}+2} \)
\(=\left( \frac{f }{f+ f^{*}} \right) ^2\, \left\{ (1-\lambda _f)\, \frac{r_1\,d^{*}}{\mathcal{D}}\,\left[ \lambda _s\,\sigma \,(1-\delta )\, \left( (1-\gamma )\,\frac{r_1\,b^{*}}{\mathcal{R}_b}\,\alpha \, \frac{b^{*}}{b+b^{*}} + \gamma \,\frac{r_1\,e^{*}}{\mathcal{R}_c} \right) \, \frac{e}{e + e^{*}} \right] \right\} ,\)
-
(34)
\(P_{e^{*},b^{*},f^{*}\rightarrow e^{*}-1,b^{*}+1,f^{*}-2} \!=\! \left( \! \frac{f^{*}}{f+ f^{*}} \!\! \right) ^2 \!\!\! \left\{ (1-\lambda _f) \frac{d}{\mathcal{D}} \! \!\left[ \! \lambda _s \sigma (1-\delta ) \! \left( (1-\gamma ) \frac{b}{\mathcal{R}_b} \alpha \frac{b}{b+b^{*}} \!+\! \gamma \frac{e}{\mathcal{R}_c} \right) \! \frac{e^{*}}{e + e^{*}} \! \right] \!\right\} ,\)
-
(35)
\(P_{e^{*},b^{*},f^{*}\rightarrow e^{*}-1,b^{*}+1,f^{*}-1} \! =\! \frac{2 f f^{*}}{(f+ f^{*})^2} \!\! \left\{ (1-\lambda _f) \frac{d}{\mathcal{D}}\!\! \left[ \lambda _s \sigma (1-\delta )\!\left( \! (1-\gamma )\frac{b}{\mathcal{R}_b} \alpha \frac{b}{b+b^{*}}+ \gamma \frac{e}{\mathcal{R}_c} \! \right) \! \frac{e^{*}}{e + e^{*}} \! \right] \! \right\} ,\)
-
(36)
\(P_{e^{*},b^{*}\rightarrow e^{*}+1, b^{*}-1} = \left( \frac{f}{f+ f^{*}} \right) ^2\,\left\{ (1-\lambda _f)\, \frac{d}{\mathcal{D}}\, \left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{b}{\mathcal{R}_b}\,\alpha \,\frac{b^{*}}{b+b^{*}}\, \frac{e}{e+e^{*}} \ \right] \right\} ,\)
-
(37)
\(P_{e^{*},b^{*},f^{*}\rightarrow e^{*}+1, b^{*}-1,f^{*}-1} = \frac{2 f f^{*}}{(f+ f^{*})^2} \, \left\{ (1-\lambda _f)\, \frac{d}{\mathcal{D}}\, \left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{b}{\mathcal{R}_b}\,\alpha \,\frac{b^{*}}{b+b^{*}}\, \frac{e}{e+e^{*}} \ \right] \right\} ,\)
-
(38)
\(P_{e^{*},b^{*},f^{*}\rightarrow e^{*}+1, b^{*}-1,f^{*}-2} = \left( \frac{f^{*}}{f+ f^{*}} \right) ^2\,\left\{ (1-\lambda _f)\, \frac{d}{\mathcal{D}}\, \left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{b}{\mathcal{R}_b}\,\alpha \,\frac{b^{*}}{b+b^{*}}\, \frac{e}{e+e^{*}} \ \right] \right\} ,\)
-
(39)
\(P_{e^{*},b^{*},d^{*},f^{*}\rightarrow e^{*}+1, b^{*}-1,d^{*}-1,f^{*}+2} \)
\(= \left( \frac{f}{f+ f^{*}} \right) ^2\,\left\{ (1-\lambda _f)\, \frac{r_1 d^{*}}{\mathcal{D}}\,\left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{b}{\mathcal{R}_b}\,\alpha \,\frac{b^{*}}{b+b^{*}}\, \frac{e}{e+e^{*}} \ \right] \right\} ,\)
-
(40)
\(P_{e^{*},b^{*},d^{*},f^{*}\rightarrow e^{*}+1, b^{*}-1,d^{*}-1,f^{*}+1} \)
\( =\frac{2 f f^{*}}{(f+ f^{*})^2}\, \left\{ (1-\lambda _f)\, \frac{r_1 d^{*}}{\mathcal{D}}\,\left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{b}{\mathcal{R}_b}\,\alpha \,\frac{b^{*}}{b+b^{*}}\, \frac{e}{e+e^{*}} \ \right] \right\} ,\)
-
(41)
\(P_{e^{*},b^{*},d^{*}\rightarrow e^{*}+1, b^{*}-1,d^{*}-1} = \left( \frac{f^{*}}{f+ f^{*}} \right) ^2\! \left\{ (1-\lambda _f)\, \frac{r_1 d^{*}}{\mathcal{D}}\,\left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{b}{\mathcal{R}_b}\,\alpha \,\frac{b^{*}}{b+b^{*}}\, \frac{e}{e+e^{*}} \! \right] \right\} ,\)
-
(42)
\(P_{e^{*},b^{*}\rightarrow e^{*}-1, b^{*}+2} = \left( \frac{f}{f+ f^{*}} \right) ^2\,\left\{ (1-\lambda _f)\, \frac{d }{\mathcal{D}}\, \left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{r_1 b^{*}}{\mathcal{R}_b}\,\alpha \,\frac{b}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} \! \right] \right\} ,\)
-
(43)
\(P_{e^{*},b^{*},f^{*}\rightarrow e^{*}-1, b^{*}+2,f^{*}-1} = \frac{2 f f^{*}}{(f+ f^{*})^2} \, \left\{ (1-\lambda _f)\, \frac{d }{\mathcal{D}}\, \left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{r_1 b^{*}}{\mathcal{R}_b}\,\alpha \,\frac{b}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} \ \right] \right\} ,\)
-
(44)
\(P_{e^{*},b^{*},f^{*}\rightarrow e^{*}-1, b^{*}+2,f^{*}-2} = \! \left( \frac{f^{*}}{f+ f^{*}} \right) ^2\! \left\{ (1-\lambda _f)\, \frac{d }{\mathcal{D}}\, \left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{r_1 b^{*}}{\mathcal{R}_b}\,\alpha \,\frac{b}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} \! \right] \right\} ,\)
-
(45)
\(P_{e^{*},b^{*},d^{*},f^{*}\rightarrow e^{*}-1, b^{*}+2,d^{*}-1,f^{*}+2} \)
\( = \left( \frac{f}{f+ f^{*}} \right) ^2\, \left\{ (1-\lambda _f)\, \frac{r_1 d^{*} }{\mathcal{D}}\,\left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{r_1 b^{*}}{\mathcal{R}_b}\,\alpha \,\frac{b}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} \ \right] \right\} ,\)
-
(46)
\(P_{e^{*},b^{*},d^{*},f^{*}\rightarrow e^{*}-1, b^{*}+2,d^{*}-1,f^{*}+1} \)
\( =\frac{2 f f^{*}}{(f+ f^{*})^2} \, \left\{ (1-\lambda _f)\, \frac{r_1 d^{*} }{\mathcal{D}}\,\left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{r_1 b^{*}}{\mathcal{R}_b}\,\alpha \,\frac{b}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} \ \right] \right\} ,\)
-
(47)
\(P_{e^{*},b^{*},d^{*},f^{*}\rightarrow e^{*}-1, b^{*}+2,d^{*}-1,f^{*}+2} \)
\( = \left( \frac{f^{*}}{f+ f^{*}} \right) ^2\, \left\{ (1-\lambda _f)\, \frac{r_1 d^{*} }{\mathcal{D}}\,\left[ \lambda _s\,\sigma \,(1-\delta )\,(1-\gamma ) \,\frac{r_1 b^{*}}{\mathcal{R}_b}\,\alpha \,\frac{b}{b+b^{*}}\, \frac{e^{*}}{e+e^{*}} \ \right] \right\} ,\)
where \(\mathcal{F}, \mathcal{D}, \mathcal{R}_b,\mathcal{R}_c\) are defined in the following
1.2 B. Fixation Probability
In a one-dimensional Markov model when there is only one compartment X, the fixation probability \(\pi ^{X}_j\), which is the probability that the progeny of j number of mutants will take over the entire compartment X of size \(N>j\), satisfies the following system of the equations.
where \(P_{j \rightarrow m}\) is the probability of transition from the state with j number of mutants to the state with m number of mutants in the population X.
1.3 C. The Fixation Probability of Stem Mutants Inside CeSC Group
To obtain the fixation probability of mutants in CeSCs, it is sufficient to trace mutants only in the stem cell niche, because only BSC mutants can migrate backward to \(S_c\) and non-stem cell mutants cannot change the number of mutants in \(S_c\). Hence, we denote the fixation probability of \(e^{*}\) number of CeSCs and \(b^{*}\) number of BSCs mutants in CeSCs by \(\pi ^\mathrm{Ce}_{(e^{*},b^{*})}\). We consider a bi-variable Markov chain with the initial state \((e^{*},b^{*})=(1,0)\). Additionally, the initial conditions are \(\pi ^\mathrm{Ce}_{(0,0)}=0\) and \(\pi ^\mathrm{Ce}_{(S_c,b^{*})}=1\) for any \(0\le b^{*} \le |S_b|\).
Here, we obtain the fixation probability \(\pi ^\mathrm{Ce}_{(e^{*},b^{*})}\) through a two-variable Markov process of all possible states \((e^{*},b^{*})\) where \((e^{*},b^{*})\ne (0,0)\) and \(0\le e^{*}\le |S_c|-1, 0\le b^{*}\le |S_b|\). The fixation probability \(\pi ^\mathrm{Ce}_{(e^{*},b^{*})}\) satisfies the following system of equations, which is the two-dimensional version of the system of equations (1).
The above system of equations can be rewritten in the following matrix form.
where

The nonzero transition probabilities in \(\mathrm{P}^{Ce}\) are
Here, we supposed that \(\lambda _f<1, \lambda _s\ne 0, \sigma \ne 0\), and \(0\le e^{*}\le |S_c|-1, 0\le b^{*}\le |S_b|\). We calculate the fixation probability \(\pi ^\mathrm{Ce}_{(e^{*},b^{*})}\) of \(e^{*}\) number of CeSC and \(b^{*}\) number of BSC mutants in \(S_c\), by numerically solving the system of Eq. 3.
Note that based on the transition probabilities, when stem cells divide only asymmetrically (i.e., when \(\sigma =0\)), then \(\pi ^\mathrm{Ce}_{(e^{*},0)}=0\). Note, when stem cells divide asymmetrically, no division occurs in the CeSC compartment; thus, the number of CeSC mutants does not change. The same result can be obtained when division is only occurring in \(D_t\) compartment (\(\lambda _s=0\)).
1.3.1 C1. The Fixation Probability of CeSC Mutants in \(S_c\) for \(\alpha =0\)
When \(\alpha =0\), \(\lambda _s > 0\), and \(\gamma >0\), we can simplify the system of Eq. (2) to the following system of equations for all \(0\le b^{*}\le |S_b|\).
From the above system of equations, we get
Therefore, the fixation probability of one mutant central stem cell in \(S_c\), i.e., the probability of the progeny of one CeSC mutant taking over the entire \(S_c\), is
1.4 D. The Fixation Probability of CeSC or BSC Mutants Inside \(S_b\)
Here, we obtain the fixation probability \(\pi ^\mathrm{BC}_{(e^{*}, b^{*})}\), which is the probability that the progeny of \(e^{*}\) number of CeSC and \(b^{*}\) number of BSC mutants will take over \(S_b\). Similar to the previous section, we consider a bi-variable Markov chain. Applying a two-variable Markov process of all possible states \((e^{*},b^{*})\) for \((e^{*},b^{*})\ne (0,0)\) and \(0\le e^{*}\le |S_c|, 0\le b^{*}\le |S_b|-1\), the total number of different possible states is \((|S_c|+1)|S_b| -1\). The fixation probability \(\pi ^\mathrm{BC}_{(e^{*}, b^{*})}\) satisfies the following system of equations
We calculate the fixation probability \(\pi ^\mathrm{BC}_{(e^{*},b^{*})}\) of \(e^{*}\) number of CeSC and \(b^{*}\) number of BSC mutants in \(S_b\), by numerically solving the matrix representation of the above system of equations (9). Note that when \(\lambda _s=0\), \(\lambda _f=1\), or \(\sigma =0\), the number of BSC mutants does not change, therefore \(\pi ^\mathrm{BC}_{(e^{*},b^{*})}=0\) for \(b^{*}<|S_b|\).
1.4.1 D1. The Fixation Probability of BSC Mutants Inside \(S_b\) for \(\alpha =0\)
Assuming \(\lambda _s\ne 0, \lambda _f<1, \sigma \ne 0,\) and \(\alpha =0\), the system (9) can be reduced to the following system for all \(0\le e^{*} \le |S_c|\):
This system of equations reveals that
Therefore, the fixation probability of a single BSC mutant in \(S_b\) is given by
1.5 E. The Fixation Probability of TA Mutants Inside the TA Compartment
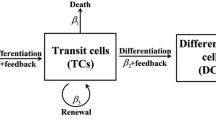
Here, we assume mutants are only located in the TA compartment, and all other cells in the other compartments are wild-type. Since TA mutants can only increase the number of mutants in FD and TA compartments and there is no backward migration from \(D_f\) to \(D_t\), only mutants in \(D_t\) can change the number of mutants in the TA compartment. Thus, the fixation probability \(\pi ^\mathrm{TA}_{d^{*}}\) of \(d^{*}\) number of mutants in \(D_t\) can be obtained using the following system of equations.
where
and
\(P^{TA}_{d^{*}\rightarrow d^{*}+1}\) is the sum of all the transition probabilities, which tend to an increase by one in the number of mutant TA cells, and \(P^{TA}_{d^{*}\rightarrow d^{*}-1}\) is the sum over all the possible transition probabilities leading to a decrease by one in the number of mutant TA cells. More precisely,
The solution to this system is in the following form
where \(\Gamma (t)=\int _0^{\infty } x^{t-1}\,\mathrm{e}^x\, \mathrm{d}x\) is the gamma function. Then, the fixation probability \(\pi _1^\mathrm{TA}\) can be derived from the following relation
Equation (13) implies that if \(\lambda _s=0\), then \( \pi ^\mathrm{TA}_{d^{*}}=\displaystyle {\frac{1}{|D_t|}}\).
1.6 F. The Fixation Probability of Mutant FD Cells in \(D_f\)
Assuming the initial state is \((e^{*}, b^{*}, d^{*},f^{*}) = (0,0, 0, f^{*} )\), only the number of FD mutants can vary. Therefore, the fixation probability of \( f^{*}\) number of FD mutants, \(\pi ^\mathrm{FD}_{f^{*}}\), can be obtained from the following system of equations.
where
and
where
If divisions never occur in the FD compartment, i.e., \(\lambda _f=0\), then the number of FD mutants (\(d^{*}\)) remains constant, implying \(\pi ^\mathrm{FD}_{f^{*}} =0\), for \(1\le f^{*} \le |D_f|-1\).
1.7 G. The Fixation Probability of Mutant TA and FD Cells in the FD Compartment
In this part, we investigate the probability of the progeny of mutant TA cells taking over the entire FD compartment. We assume that initially there are only \(d^{*}\) number of mutant TA cells and \(f^{*}\) number of mutant FD cells, i.e., \((e^{*}, b^{*}, d^{*}, f^{*}) = (0,0, d^{*}, f^{*} )\). Following the methods used in the previous sections, we assume that \(\pi ^\mathrm{FD}_{(d^{*},f^{*})}\) is the fixation probability of starting from \(d^{*}\) number of mutant TA cells and \(f^{*}\) number of mutant FDs (initial state is \((d^{*},f^{*})\)). We consider a bi-variable Markov chain to explore the cell dynamics in the FD and TA compartments. The fixation probability \(\pi ^\mathrm{FD}_{(d^{*}, f^{*})}\) satisfies the following equation.
In order to obtain the fixation probability \(\pi ^\mathrm{FD}_{(d^{*},f^{*})}\), we numerically solve the matrix representation of the system of Eq. (22). Note that if \(\lambda _f=1\), then there is no chance that mutant TA cells divide, thus \(\pi ^\mathrm{FD}_{d^{*},0}=0\) for all \(1\le d^{*} \le |D_t|\).
1.8 H. Numerical Simulation
We use the algorithm given in the methods section to do the numerical simulations, and you can find the python code that we used to generate the results in (https://sites.google.com/site/leilishahriyari/FourCompMoranModel_Shahriyari.py?attredirects=0&d=1). In order to obtain the fixation probability and time to fixation through simulations, we set the maximum updating time T equal to 10,000,000 time steps. We run the algorithm for 100 times, and we calculate the ratio of the number of times that fixation occurs over 100. We repeat this process for 10 times to obtain the mean and the standard deviation of the fixation probabilities. Moreover, to obtain the fixation time, we collect the fixation time for each single run whenever the fixation happens. Then, we calculate the average and standard deviation of these collected fixation times. To represent the simulation time steps in terms of days, we assume that the average cell cycle time is 18 hours for mouse and one day for human (Cassimerisz et al. 2011; Potten et al. 1992; Bach 2000). According to Bach (2000), the average cell cycle time of epithelial cells in the mouse crypts is \(12-13\) hours for rapidly proliferating cells, and 24 hours for stem cells. Potten et al. (1992) suggested that the human colonic cell cycle is 17–48.6 h. This means that if the total number of cells is N, then we present the time step t as \(\frac{t}{N}+1\) day in all simulations done for human crypts in Figs. 4, 5, 6, 7, and 8, and the time step t has been presented as \(0.75\frac{t}{N}+1\) day for all simulations done for mouse crypts in Fig. 3.
Rights and permissions
About this article

Cite this article
Mahdipour-Shirayeh, A., Shahriyari, L. Modeling Cell Dynamics in Colon and Intestinal Crypts: The Significance of Central Stem Cells in Tumorigenesis. Bull Math Biol 80, 2273–2305 (2018). https://doi.org/10.1007/s11538-018-0457-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11538-018-0457-8