
Abstract
Two species of non-native sea lavender, Limonium ramosissimum (Poir.) Maire (Algerian sea lavender) and Limonium duriusculum (Girard) Fourr. (European sea lavender) are prolific invaders of California salt marshes. We examined the efficacy of three non-herbicide treatments [tarping, selective removal (hand pulling), and scraping], and one herbicide (Telar®) treatment, in eliminating L. ramosissimum and L. duriusulum, as well as treatment impacts to native vegetation within three southern California salt marshes. Our experiments demonstrated that the non-herbicide treatments initially suppressed cover of L. ramosissimum and L. duriusulum to near zero but varied over time in their suppression of L. ramosissimum and L. duriusulum, and their effects on native species. Depending on the tarping duration, tarping eradicates L. ramosissimum and L. duriusulum at least a year post treatment with little to no long-term negative impacts to native vegetation. Selective removal had minimal effect on native species but did not consistently suppress L. duriusulum over time. Scraping eradicates L. ramosissimum and L. duriusulum; however, its negative impacts on native species make it an undesirable treatment across large areas. The herbicide treatment was not as effective in suppressing L. ramosissimum, either initially or over time, and it negatively impacted native species. Our findings support the use of tarping as the primary management method to control or eradicate dense infestations of L. ramosissimum and L. duriusulum in salt marshes. We suggest that a combination of tarping and selective removal be used to control L. ramosissimum and L. duriusulum in areas of lower infestation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Non-native invasive plants can form monocultures that inhibit the recruitment and survival of native plants, reduce native biodiversity, alter soil properties and food webs, and affect the overall ecosystem functioning of coastal wetlands (Zedler and Kercher 2004; Levin et al. 2006; Gratton and Denno 2006; Boyer and Burdick 2010; Burdick et al. 2021). Two species of invasive perennial forb, Limonium ramosissimum (Poir.) Maire (Algerian sea lavender) and Limonium duriusculum (Girard) Fourr. (European sea lavender) (Plumbaginaceae) have increased in distribution and abundance in the coastal wetlands of central and southern California over the past 25 years. These Limonium spp. can form monospecific stands that resist the recruitment of, and overgrow, native species (Hubbard and Page 1997; Fig. 1). Both species are horticultural escapees native to the Mediterranean region and proficient invaders of the upper mid-to high-elevation salt marsh and salt marsh-upland transition zone of California salt marshes, occurring at elevations from approximately mean high water (MHW) to above mean higher high water (MHHW) (Hubbard and Page 1997; Archbald and Boyer 2014a).

Infestation of Limonium ramosissimum at Aqua Hedionda Lagoon. Dense vegetative cover of Limonium ramosissimum is in the foreground of the picture and dense flowering of Limonium ramosissimum is pictured between its dense vegetative cover and the stands Juncus acutus L. (spiny rush)
The invasion and spread of non-native Limonium spp. (hereafter, Limonium) in California marshes is likely facilitated by: (1) a wide physiological tolerance of these plants to saline soils; (2) a high reproductive rate with the production of numerous flowers per flower stalk; (3) the potential to establish a large seed bank (> 10,000 seeds m−2) with wide dispersal of buoyant seeds; (4) the potential production of a large number of recruits per year (> 1000 seedlings m−2); and (5) a clonal growth form with a low lying dense canopy and subsurface branching giving rise to new rosettes that may inhibit the germination of native plants (Hubbard and Page 1997; Archbald and Boyer 2014a, b).
Limonium create a plant profile that is much lower and denser than the architecture of the native vegetation they replace such as Salicornia pacifica Standl or the native Limonium californicum (Boiss.) A. Heller. Sensitive species that may directly or indirectly be affected by Limonium include the State and Federally protected Belding’s savannah sparrow (Passerculus sandwichensis beldingi) and light-footed Ridgway’s rail (Rallus longirostris obsoletus; formerly light footed clapper rail), which use native S. pacifica (pickleweed), as breeding and/or foraging habitat, and the Federally protected plant Chloropyron maritimum ssp. maritimum (Nutt. Ex Benth.) A. Heller (salt marsh bird’s-beak) (Archbald and Boyer 2014a, Zembal et al. 2015; U.S. Fish and Wildlife Service 2020 and 2023). L. ramosissimum and L. duriusculum are included in the California Invasive Plant Council (Cal-IPC) list of invasive plants (https://www.cal-ipc.org/plants/profiles) and the subject of several Cal-IPC news articles (Whitcomb 2016; Johnson and Brooks 2018; Stalker 2023).
To date, L. ramosissimum and L. duriusculum have been documented from the San Francisco Bay Area to Southern California. Coastal wetlands invaded by these species include, listed from north to south, sites within multiple counties in and around San Francisco Bay, San Luis Obispo County (Morro Bay), Santa Barbara County (Carpinteria Salt Marsh Reserve), Orange County (Bolsa Chica Wetlands, Huntington Beach Wetlands, Upper Newport Bay), and San Diego County (Aqua Hedionda Lagoon, Batiquitos Lagoon, San Elijo Lagoon, San Diego River Mouth, San Diego Bay, and the Tijuana Estuary).
An effective method to manage and, if possible, eradicate Limonium with minimal negative effects on native species is necessary to avoid further habitat degradation. Here, we report the results of experiments conducted within three southern California tidal wetlands comparing the response of L. ramosissimum and L. duriusculum, and native species, to common techniques that are used for controlling non-native species. These techniques include removing Limonium individuals by hand (selective removal), scraping the marsh surface with garden tools (scraping), securing black plastic tarps for a duration of time (tarping; also known as solarization), and treating individuals with herbicide. Our objective was to evaluate which method was the most effective at removing invasive plants from large areas within the salt marsh with the least negative impact on native species.
Methods
Study sites
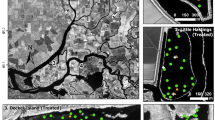
We conducted our experiments in three geographically separated estuaries invaded by Limonium. ramosissimum or L. duriusculum: 1) San Diego River (SDR), 2) Agua Hedionda Lagoon (AHL), and 3) Carpinteria Salt Marsh (CSM) (Fig. 2). The experiment at SDR was conducted on a spit of land on the south side of the San Diego River Flood Control Channel just east of the river mouth (32.755560°, − 117.246924°). Ten native salt marsh plant species are potentially affected by L. duriusculum within this study site (Frankenia salina (Molina) I. M. Johnst., Triglochin maritima L., Limonium. californicum (Boiss.) A. Heller, Distichlis littoralis (Engelm.) H. L. Bell & Columbus, Salicornia pacifica, Jaumea carnosa (Less.) A. Gray, Batis maritima L., Cuscuta californica Costea & M. Wright, Suaeda esteroa Ferren & S. A. Whitmore, and Chloropyron maritimum ssp. maritimum). The experiment at AHL was established at the southeast end of the lagoon (33.139551°, − 117.310964°). Three native plant species are potentially affected by the expansion of L. ramosissimum (S. pacifica, J. carnosa, F. salina and D. littoralis). The experiment at CSM was located at the southwest side of the marsh (34.399078°, − 119.537885°). Five native plant species are potentially affected by L. duriusculum (Distichlis spicata (L.) E. Greene, J. carnosa, D. littoralis, S. pacifica, and C. maritimum ssp. maritimum).

The three salt marshes where experiments were conducted
Experimental design
We used three experimental designs to compare four different, commonly used techniques for controlling non-native plant species. The first design was employed at all three sites and consisted of four treatments distributed in replicate blocks within mid-elevation salt marsh (Table 1). Blocks measured 4 × 4 m and treatment plots measured 1.8 × 1.8 m. The treatment plots were: (1) selective removal where above ground L. ramosissimum or L. duriusculum and associated root mass was selectively removed by hand pulling, but native species were left intact; (2) scraping where garden tools (e.g. hoe, rake, shovel) were used to remove all above ground vegetation and one half inch of topsoil from the plots; (3) tarping where the entire plot was covered with 2 mm thick black plastic and secured to the ground with bricks or staples for a duration of time, after which the tarp, bricks, and staples were removed from the marsh; and (4) an unmanipulated control. Some details of these experiments, such as replication, duration of tarping, duration and frequency of monitoring differed among sites for logistical reasons. Consequently, wetland site is not included as a main effect in the analysis (see below). A second design using only tarping and control treatments within replicate blocks was conducted at AHL at an elevation 0.5 m higher than the mid-elevation plots to explore whether the effects of tarping varied with elevation and tidal inundation. The first and second experimental design at AHL were conducted concurrently to allow for direct comparisons.
A third design, conducted only at AHL, compared the effectiveness of herbicide to tarping in eliminating L. ramosissimum. We used a randomized block design with five replicate blocks, each containing three different treatment plots measuring 4.3 by 4.6 m (Table 1). The treatments were tarping, herbicide, and an unmanipulated control. To evaluate if the duration of tarping could affect the results, we removed half of the tarp after a tarping duration of 15 weeks (15-week-tarp) and the remaining half after a duration of 20 weeks (20-week-tarp). The herbicide treatment consisted of two separate applications of Telar®, chlorsulfuron benzenesulfonamide, conducted at the initiation of the experiment and 28 weeks later. Telar® is an herbicide that stops cell division in plant roots and shoots, which causes plants to stop growing (Washington State Department of Transportation 2017). Telar® was applied by California Department of Fish and Wildlife at the maximum label rate of 85 g per 0.40 hectare.
The percent cover of all plant species was sampled using point contact (Manly et al. 2015) by recording species intercepts at points uniformly spaced in a grid within each treatment plot. There were 55 sampling points for the first and second experimental design and 140 sampling points for the third experimental design. Percent cover of Limonium species and all native species within a plot was calculated as the total count of either category divided by the number of points sampled within a plot. Due to multiple species encountered at a point intercept, it was possible to achieve a total plant percent cover greater than 100 percent within a plot. Percent cover was measured immediately prior to initiating treatments, and during and after treatments had concluded. Follow-up sampling time intervals varied among the wetlands due to logistical constraints. We characterized the wilted dark brown Limonium resulting from tarping and the stiff light brown Limonium resulting from herbicide as dead; only green and obviously alive Limonium are reported. We also quantified flowering of L. ramosissimum as part of experimental design 3 when we sampled in September 2017. Table 1 shows the number of replicates, duration of tarping, and timing of sampling for each site and experimental design.
Data analysis
We used a mixed model repeated measures design with eradication treatment as a fixed factor, plots as the repeated measure, and duration (weeks) as a random factor at all the sites. We also tested for differences between treatments and controls at the end of each experimental period using Tukey’s HSD test to adjust for multiple comparisons with an experiment wise Type 1 error of 0.05. All data analyses were performed in JMP Pro 16.0 (JMP 1989–2023).
Results
All non-herbicide treatments in the mid-marsh (selective removal, scraping, tarping) were initially effective in reducing the cover of Limonium to near zero, but follow-up sampling revealed differences among treatments and experiments in the successful longer-term suppression of Limonium, and in negative effects on native species. There was a significant treatment by time (week) interaction for cover of Limonium at each site.
Selective removal initially reduced cover of Limonium at all sites but then cover increased at SDR such that 24 weeks after treatment Limonium was statistically comparable to the controls (Table S1a-b; Fig. 3a), whereas cover of Limonium remained low at AHL and CSM (after 55 and 29 weeks, respectively) (Tables S2a-b, S3a-b; Figs. 4a, 5a). Selectively removing Limonium largely avoided impacts to native species, having no significant effect on the cover of native species at any site initially or over time (Tables S1c-d, S2c-d and S3c-d; Figs. 3b, 4b, 5b).

Mean percent cover in response to experimental treatments at San Diego River Mouth between March 2013 and March 2014 (n = 5). Cover associated with 0 months is pre-treatment. The same letter at treatment means at each sampling point denote means that are not significantly different. a. The % cover of Limonium duriusculum. b. The % cover of Salt Marsh Natives

Mean percent cover in response to experimental treatments in mid-elevation salt marsh at Agua Hedionda Lagoon between October 2014 and November 2015 (n = 3). Cover associated with 0 months is pre-treatment. The same letter at treatment means at each sampling point denote means that are not significantly different. a. The % cover of Limonium ramosissimum. b. The % cover of Salt Marsh Natives

Mean percent cover in response to experimental treatments at Carpinteria Salt Marsh between November 2014 and June 2015 (n = 4). Cover associated with 0 months is pre-treatment. The same letter at treatment means at each sampling point denote means that are not significantly different. a. The % cover of Limonium duriusculum. b. The % cover of Salt Marsh Natives
The scraping treatment, which removed all surface vegetation and some surface sediment, produced an immediate reduction in Limonium cover at all sites comparable to the selective removal treatment (Tables S1a-b, S2a-b, S3a-b; Figs. 3a, 4a, 5a). However, scraping had a significant negative effect on native species at one of three sites (AHL), with native cover only ~ 65% of that in the control 55 weeks after being scraped (Table S2c-d; Fig. 4b).
The tarping treatment was also effective within mid-elevation salt marsh in reducing Limonium cover, which remained low (first experimental design: 0.5%—SDR, 0.0%—AHL, and 1.4%—CSM; third experimental design: 3%—AHL) through the conclusion of these experiments (Tables S1a-b, S2a-b, S3a-b, S4a-b; Figs. 3a, 4a, 5a, 6a). Tarping would breakdown or dissolve the biomass of Limonium. Tarping for a duration as short as 10 weeks at SDR and 13 weeks at AHL reduced cover of Limonium to zero (Tables S1a-b, S2a-b; Figs. 3a, 4a). However, the effect of tarping on native vegetation was more variable, influenced by both initial cover and tarping duration (Tables S1c-d, S2c-d, S3c-d, S4c-d; Figs. 3b, 4b, 5b, 6b). When initial cover of natives was relatively low at AHL (i.e., below 50%), 15 and 20 weeks of tarping did not negatively affect cover of natives (i.e., was comparable to the control) and ultimately benefited native cover (i.e., had a significantly higher cover than the control) after the tarps had been removed (Table S4d, Fig. 6b). However, increasing the duration of tarping, such as the 30 weeks of tarping at CSM (Table S3c-d; Fig. 5b) and 55 weeks at AHL (Table S2c-d; Fig. 4b) did negatively affect cover of natives (Tables S2c-d, S3c-d; Figs. 4b, 5b); We did not sample at these locations after the removal of tarps, so we do not know whether or how long it took for natives to recover. When initial cover of natives was relatively high (i.e., above 70%), a tarping duration as low as 10 weeks at SDR (Table S1c-d; Fig. 3b) and somewhere between 13 and 18 weeks at AHL (Table S2c-d; Fig. 4b) resulted in an initial reduction of native cover that recovered within 12 weeks of removing the tarps (i.e., comparable to the control) at SDR (Table S1c-d; Fig. 3b) and within 5 weeks of removing the tarps at AHL (Tables S2c-d; Fig. 4b).

Mean percent cover in response to experimental treatments at Agua Hedionda Lagoon between December 2016 and June 2018 (n = 5). Cover associated with 0 months is pre-treatment. Herbicide treatments conducted December 2016 and June 2017. The same letter at treatment means at each sampling point denote means that are not significantly different. a. The % cover of Limonium ramosissimum. b. The % cover of Salt Marsh Natives
Comparison of the tarping results at the high-elevation salt marsh with the mid-elevation salt marsh at AHL suggests that elevation or the associated tidal inundation frequency/duration could affect the timeline of Limonium reduction by tarping. A longer tarping duration was required to reduce Limonium cover to 0% in the high-elevation salt marsh (occurred between 18 and 55 weeks) (Tables S5a-b; Fig. 7a) compared with the mid-elevation salt marsh (occurred within 13 weeks) (Tables S2a-b; Fig. 4a). However, keeping the tarps in place for too long, such as 55 weeks, also significantly reduced the cover of native species, which was comparable to that of Limonium at the end of the experiment (Tables S5a-d; Figs. 7a, b).

Mean percent cover in response to experimental treatments at Agua Hedionda Lagoon in high-elevation salt marsh between October 2014 and November 2015 (n = 3). Cover associated with 0 months is pre-treatment. The same letter at treatment means at each sampling point denote means that are not significantly different. a. The % cover of Limonium ramosissimum. b. The cover of Salt Marsh Natives
Two applications of herbicide were less successful than tarping in reducing the cover of Limonium (Tables S4a-b; Fig. 6a). From 16 to 81 weeks after the initiation of the herbicide treatment, cover of Limonium averaged ~ 26%, significantly higher than tarping treatments where cover remained 5% or less (Table S4b; Fig. 6a). Herbicide treatments did not breakdown the biomass of Limonium even when it appeared dead such that the Limonium continued to take up physical space. Herbicide’s negative impact on native species was not statistically significant (i.e., compared to controls); however, cover of natives in the herbicide treatment was significantly lower than in the tarping treatments at the conclusion of the experiment (Table S4d; Fig. 6b).
Unexpectedly, in June 2017, a little more than six months following the initiation of the herbicide experiment at AHL (third experimental design), we observed flowering of L. ramosissimum in the herbicide treatment when no flowering was observed in the control or tarping treatments, or outside the experimental plots. When we measured cover of flowering plants three months later (September 2017), flowering L. ramosissimum in herbicide plots averaged 10% cover and significantly higher than the non-herbicide and control treatments (Table S6; Fig. 8a, b).

Mean percent cover of non-flowering Limonium ramosissimum and flowering Limonium ramosissimum within each treatment 39 weeks after initiation of all treatments at Aqua Hedionda Lagoon. The same letter at treatment means at each sampling point denote means that are not significantly different. a. The % cover of non-flowering of Limonium ramosissimum. b. The % cover of flowering Limonium ramosissimum
Discussion
Our experiments demonstrated that the non-herbicide treatments initially suppressed the cover of non-native Limonium to near zero, but the treatments varied in their effectiveness over time, impact on native species, and practicality. Selective removal through the manual hand pulling of Limonium had a minimal effect on native species, but this treatment did not lead to a consistent suppression of Limonium over time. For example, selective removal was effective at suppressing Limonium cover over time at AHL and CSM, whereas Limonium re-established to achieve high cover within one-year post-treatment at SDR. One explanation for this inconsistency is that selective removal did not remove enough root stock to suppress resprouting. However, the same hand pulling technique was used at all sites, suggesting that this was unlikely. More likely, selective removal did not reduce the non-native seed bank, which may have differed between sites, enabling the sprouting of new Limonium plants over time. Consequently, surveillance and selective removal would need to be conducted over multiple years to fully suppress Limonium which is a typical recommendation for the control of many species of non-native plants (DiTomaso et al. 2013).
Selective removal is also very time and labor intensive in areas of high Limonium infestation, and requires removal of biomass from the marsh, which may not be feasible on a large scale. It was the primary technique used to remove L. duriusculum from the SDR by the City of San Diego as a mitigation project. The effort greatly reduced L. duriusculum; however, individuals continued to be observed after four years of effort indicating that removal efforts needed to continue (Tierra Data Inc 2023). Selective removal may be practical in small areas with a low infestation, where land managers will continue to visit the site to remove future infestations, and Limonium plants are obvious to personnel removing it.
Scraping was effective in reducing the percent cover of Limonium to, or near, zero for up to a year post-treatment. However, the potential longer-term negative impacts of this treatment on native species (e.g., at AHL) make scraping an undesirable treatment alternative. In addition, removing soil creates a disturbance that could affect the moisture and nutrient content of the remaining soil and facilitate the establishment of other invasive plant species (Zedler and Kercher 2004; Ehrenfeld 2008). Scraping is slow, labor intensive, generates significant mass of soil and vegetation for disposal, and is probably not feasible to apply on a large scale without heavy equipment. Consequently, this treatment is not recommended on a large scale where native vegetation is desirable.
Our experiments demonstrate that at least 10 weeks of tarping can reduce the cover of living Limonium to near zero. Furthermore, tarped Limonium decompose over time, thereby opening space for the recruitment and recovery of native species. Our results show that once Limonium is removed, native plants can resprout within the tarping treatment area in the absence of additional management effort. Tarping, also known as solarization or plastic mulching, has been used in horticulture to control weeds, and larger non-native plant species such as Carpobrotus edulis L. (iceplant) (DiTomaso et al. 2013; Tarara 2000; Kubalek et al. 2022). Tarping kills plants through increasing the humiture and soil temperature (Horowitz et al. 1983). Though studies have found that clear plastic increases soil temperatures more and will kill more seeds than black plastic (Horowitz et al. 1983; Standifer et al. 1984), we do not recommend using clear plastic to control invasive plants in salt marshes without further study because of concern that the higher temperatures generated under clear plastic could also negatively affect native plants and the native seed bank. Further, black tarps outperform clear tarps in maritime regions characterized by persistent cloud cover and moderate temperatures, such as in California estuaries (Kubalek et al. 2022).
Our data suggest that the duration of tarping needs to balance the time necessary to eliminate Limonium with impacts to native species, and that tarping duration may be adjusted depending on elevation and time of year. Mortality of Limonium in AHL with tarping occurred more slowly at higher or drier elevations, suggesting that a longer duration of tarping may be necessary at higher elevations. We postulate that the difference in mortality of Limonium between mid- and high-elevation salt marsh may result from increased stress through the interaction of temperature with the higher soil moisture associated with a greater frequency/duration of tidal inundation at lower elevations. We also conducted our tarping experiments during the winter and spring when air temperatures are cooler. If tarping is conducted when temperatures are warmer, such as summer and fall, a shorter duration of tarping could be effective. Because of possible elevation and air temperature effects on tarping success, we suggest tarping for a duration of at least 12 to 20 weeks, to be modified based on monthly visits to the treatment sites to evaluate whether Limonium is eliminated, and the tarps can be removed.
At AHL, efforts to control L. ramosissimum prior to our experiments centered on the intermittent application of Telar® (chlorsulfuron benzenesulfonamide) by the California Department of Fish and Wildlife. The Telar® treatment was not as effective in suppressing L. ramosissimum as the other treatments, either initially or over time, and native species did not recover from Limonium infestations when herbicide were applied. Treatment by herbicide left Limonium biomass in place, which although brown, may not be completely dead. Indeed, we observed these ‘dead’ plants sprouting new green vegetation. The resprouting of L. ramosissimum following herbicide treatment suggests that the herbicide either did not contact all the living L. ramosissimum tissue or that this plant is resistant to the application of this herbicide. In addition, while Limonium may appear dead, it continues to physically occupy space, potentially inhibiting the recruitment and recovery of native vegetation. Limonium produces copious seeds (Hubbard and Page 1997). The presence of an extensive seed bank in the soil also requires that any herbicide be reapplied with possible negative unintended consequences to marsh biota. Some herbicides cannot be used below the mean high tide line, areas often heavily infested with L. ramosissimum, which also limits the general usefulness of chemical control agents. Application of herbicide also had the unintended consequence of stimulating flowering in L. ramosissimum thereby producing more Limonium seed. We speculate that stress associated with the herbicide induced the L. ramosissimum to flower earlier than elsewhere (Takeno 2016).
In conclusion, our findings support the use of tarping as the primary management method to control or eradicate dense, near monospecific infestations of Limonium in salt marshes. We suggest that a combination of tarping and selective removal be used to control Limonium in areas of lower infestation. Such an effort is currently being implemented in several southern California coastal wetlands (e.g., Agua Hedionda Lagoon, Upper Newport Bay, Huntington Beach Wetlands, Carpinteria Salt Marsh, and Sweetwater Marsh National Wildlife Refuge). Based on our findings and observations, specific recommendations include: (1) tarp dense stands of Limonium for a duration of 12 to 20 weeks with modifications appropriate for the time of year and tidal elevation; (2) use scraping or selective removal where Limonium is sparse or covers a small area; (3) place removed individuals, particularly flowering individuals, in closed plastic bags immediately and remove the bag from the site to prevent release of seeds or re-rooting; and (4) monitor the site following treatment and conduct control treatments to address Limonium sprouting from the sea bank to maintain eradication efforts.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Archbald G, Boyer KE (2014a) Distribution and invasion potential of Limonium ramosissimum subsp. Provincial in San Francisco Estuary salt marshes. San Franc Estuary Watershed Sci 12:1–22
Archbald G, Boyer KE (2014b) Potential for spread of Algerian sea lavender (Limonium ramosissimum subsp. provincial) in tidal marshes. Inv Plant Sci Manage 7:454–463
Boyer KE, Burdick AP (2010) Control of Lepidium latifolium (perennial pepperweed) and recovery of native plants in tidal marches of the San Francisco Estuary. Wetlands Ecol Manage 18:731–743
Burdick DM, Moore GE, Boyer KE (2021) Impacts of exotic and native species invading tidal marshes. In: FitzGerald DM, Hughes ZJ (eds) Salt marshes: Function, Dynamics, and Stresses. Cambridge University Press, Cambridge, pp 367–387
DiTomaso JM, Kyser GB (2013) Weed control in natural areas in the western United States. Weed Research and Information Center, University of California, p 544
Ehrenfeld JG (2008) Exotic invasive species in urban wetlands: environmental correlates and implications for wetland management. J Appl Ecol 45:1160–1169
Gratton C, Denno DF (2006) Arthropod food web restoration following removal of an invasive wetland plant. Ecol Appl 16(2):622–631
Horowitz M, Regev Y, Herzlinger G (1983) Solarization for weed control. Weed Sci 31:170–179
Hubbard DM, Page HM (1997) Biology and control of invasive sea lavender, Limonium ramosissium in Carpinteria Salt Marsh, California. Unpublished report submitted to U.S. Fish and Wildlife Service, Ventura Field Office, 2493 Portola Road, Suite B, Ventura, California
JMP ®, Version 16.0 (1989–2023) SAS Institute Inc., Cary, NC
Johnson D, Brooks AJ (2018) Controlling invasive sea lavender in Carpinteria Salt Marsh. Dispatch 26(1):4–5
Kubalek R, Granastein D, Collins D, Miles C (2022) Review of tarping and a case study on small-scale organic farms. HortTechnology 32:119–128
Levin LA, Neira C, Grosholz ED (2006) Invasive cordgrass modifies wetland trophic function. Ecology 87(2):419–432
Manly BFJ, Navarro Alberto JA (2015) Introduction to ecological sampling. Chapman and Hall/CRC applied environmental statistics book 10, 1st edn. CRC Press, Boca Raton, Florida
Stalker J (2023) Persistence: Marin Audubon’s work to keep invasive sea lavender from invading Marin County tidal wetlands. Dispatch 30(3):10–11
Standifer LC, Wilson PW, Porche-Sorbet R (1984) Effects of solarization on soil weed seed populations. Weed Sci 32:569–573
Takeno K (2016) Stress-induced flowering: the third category of flowering response. J Exp Bot 67:4925–4934
Tarara JM (2000) Microclimate modification with plastic mulch. HortScience 35(2):169–180
Tierra Data Inc (2023) U.S. Fish and wildlife service FWS-SDG-09B0224-13I0118/18F0927 Annual mitigation maintenance and monitoring report for the west mission bay drive bridge replacement project, November 1, 2021 through October 31, 2022
United States Fish and Wildlife Service (2020) Chloropyron maritimum subsp. maritimum [Cordylanthus maritimus subsp. maritimus (salt marsh bird’s beak)] 5-Year Review, p. 20
United States Fish and Wildlife Service (2023) General provisions; revised list of migratory birds. Final rule. July 31, 2023. Federal Register, 88(145), 49310–49355.
Whitcomb A (2016) Protecting San Francisco Bay salt marshes from invasive sea lavender. Dispatch 24(3):4–5
Washington State Department of Transportation (2017) Chlorsulfuron. Roadside Vegetation Management Herbicide Fact Sheet.
Zedler JB, Kercher S (2004) Causes and consequences of invasive plants in wetlands: opportunities, opportunists, and outcomes. Crit Rev Plant Sci 23(5):431–452
Zembal RS, Hoffman M, Patton RT (2015) A survey of the Belding’s Savannah sparrow (Passerculus sandwichensis beldingi) in California, 2015. California Department of Fish and Wildlife, Wildlife Branch, Nongame Wildlife Program Report, 2015- 02, Sacramento, CA, p. 20
Acknowledgements
The authors would like to thank J. Ekhoff, G. Penaflor, C. Navarro, and T. J. Gaven (California Department of Fish and Wildlife) and B. Miller (City of San Diego) for providing access to lands they manage. We also thank K. Lunneberg, L. Rodman, S. Richter (Agua Hedionda Lagoon Foundation), J. Ukegawa (Carlsbad Strawberry Company, Inc.), R. Dhu (San Diego River Park Foundation), C. Winchell (U.S. Fish and Wildlife Service), and J. Hoesterey and R. Johnson (Marine Science Institute, University of California, Santa Barbara) for field assistance. This work was supported in part by funding from the U.S. Fish and Wildlife Service Coastal Program (Agreement Numbers 81430BJ002 and F17AC00190). The findings and conclusions in this article are those of the author(s) and do not necessarily represent the views of the U.S. Fish and Wildlife Service (USFWS). The authors complied with USFWS 212 FW 1 Standards of Conduct while conducting all aspects of this work.
Funding
This work was supported by the U.S. Fish and Wildlife Service Coastal Program (Agreement Numbers 81430BJ002 and F17AC00190).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design, material preparation, and data collection. Data analyses were performed by Dr. Stephen C. Schroeter. The first draft of the manuscript was written by Carolyn Lieberman and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lieberman, C.H., Schroeter, S.C., Hormick, P. et al. Evaluation of techniques for controlling non-native sea lavenders in California coastal salt marshes. Wetlands Ecol Manage (2024). https://doi.org/10.1007/s11273-024-09994-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11273-024-09994-z