
Abstract
Planting various hybrids of larch species in commercial forestry has gained increased importance around the world. However, there are uncertainties about the genetic purity of hybrid larch seed lots due to mixture of pure and hybrid seeds in the outputs of commercial hybrid larch seed orchards. Thus, the aim of the study was to evaluate the feasibility of visible + near infrared (Vis + NIR) spectroscopy to distinguish European × Japanese larch (Larix × eurolepis) from pure European (Larix decidua) and Japanese (Larix kaempferi) larch seeds. Vis + NIR reflectance spectra were obtained using single seeds, and discriminant models were developed with partial-least square-discriminant analysis (PLS-DA) and orthogonal projections to latent structures-discriminant analysis (OPLS-DA) on both raw and pre-treated spectra. The best PLS-DA model assigned L. × eurolepis and L. decidua seeds to their respective classes with 100 % accuracy while that of L. kaempferi with 97 % accuracy. The best OPLS-DA model resulted in 100 % classification accuracy of pure and hybrid larch seeds. The origins of spectral differences between pure and hybrid larch seeds were attributed to differences in color, seed moisture content, fatty acids and proteins. In conclusion, the result demonstrates the feasibility of Vis + NIR spectroscopy as a powerful non-destructive method for verification of hybrid larch seeds, which is of great importance in maintaining the genetic purity of hybrid seed lots and for monitoring natural hybridization in commercial open pollinated mixed species seed orchard.
Similar content being viewed by others

Introduction
Planting various hybrids of larch species in commercial forestry has gained increasing importance around the world, owing to their heterotic vigor (Pâques 2004; Baltunis et al. 1998). In Sweden, for example, the interest in growing Larix eurolepis Henry—a hybrid of European larch (Larix decidua Mill.) and Japanese larch [Larix kaempferi (Lamb.) Carr.]—has increased over the past decades. This is much due to a greater focus on climate adaptation in forestry. Future climate change will probably alter the growing conditions in Scandinavia in a way that makes forestry with high productivity exotic species more attractive. Using several species, instead of the few traditional ones, Picea abies (L.) Karst and Pinus sylvestris L., is a way to spread the risk of an unknown future. Hybrid larch offers good growth, relatively short rotation age and show high wind stability at older ages (Ekö et al. 2004).
Commercial seeds of hybrid larch are produced in seed orchards, which normally include a number of grafts of one L. kaempferi clone (maternal parent) and of several L. decidua clones (paternal parents). There are also examples where the species of the maternal and paternal clones are the reversed. To get as high proportion of hybrid seed as possible, seeds are normally collected only from the maternal clones, while the paternal clones only are used for pollination (Hannerz et al. 1993). Still the seeds from seed orchards of this design show varying hybrid seed proportions over years, as shown in a French study (Acheré et al. 2002) where it varied between 2 and 67 %. This may be caused by poor synchronization in flowering among the two species in combination with external pollen from nearby larch plantations, including hybrids or the same species as the maternal clone (Pâques 2000). Another reason may be that the maternal clone is not self-sterile, i.e. producing self-pollinated seeds (Pâques et al. 2013). There are also examples of hybrid seed orchard designs where several maternal clones are included from which seeds are collected. However, the hybrid seed proportion from such designs is likely to be lower since the maternal trees are able to pollinate each other, producing viable pure parental seed. For instance, the English NT23 has such a design and has shown a hybrid rate of 15 % (Helmersson et al. 2015). The varying proportion of hybrid seeds results in problems both for nursery managers and larch growers (individual forest farmers or companies).
As seeds can easily be transferred between countries or between places within countries, there could be a high risk to the maintenance of the genetic purity of seed lots as the transaction practice hitherto is based on “trust-on-labels”. For instance, the EU regulations demands that hybrid seed producers have to provide information about the hybrid proportion (Pâques 2009). In the past, several approaches have been used to distinguish hybrid larch from its pure parental species, including morphological traits (Gauchat and Pâques 2011), cytogenetic analysis (Zhang et al. 2010), leaf fatty acid compositions (Sato et al. 2008) and molecular markers (Acheré et al. 2004; Pâques 2009). Rogueing of seedlings in the nursery is also used in practical operations, as the seedlings of pure parental species grow slowly compared to the seedlings of the hybrid. Although these methods allow identification of larch species and their hybrids, they have mostly limited application in certification of hybrid larch seed lots. Some of the approaches are destructive, but all are considered expensive.
The aim of this study was to evaluate the feasibility of near infrared spectroscopy (NIRS) as cost-effective, technically less complex and non-destructive method for identifying hybrid larch seeds from that of pure parental species. NIRS is a widely recognized analytical technique that measures moisture and chemical composition of biological materials based on absorption of near infrared radiation by bonds between light atoms, such as C–H, O–H and N–H that result in overtones and combination bands detectable in the 780–2500 nm wavelength region (Workman and Weyer 2012). NIRS has been proven successful for authentication of melon genotypes (Seregély et al. 2004), separation of Eucalyptus globulus genotypes (Castillo et al. 2008), hybrids of Eucalyptus (Humphreys et al. 2008) and pine (Espinoza et al. 2012) species as well as detection of introgression in Coffea arabica cultivars (Bertrand et al. 2005). To our knowledge, no study has been carried out to identify hybrid larch seeds using NIRS, which is of paramount importance for monitoring natural hybridization in commercial open pollinated mixed species seed orchard.
Materials and methods
Seed samples
Seeds of L. decidua, L. kaempferi and their hybrid (L. × eurolepis) were obtained from clonal archive at the research station of the Swedish Forest Research Institute at Ekebo, Sweden (55°57′N, 13°07′E, 80 m). Seeds of L. decidua of known maternal (D02V983) and paternal (S21K9780044) clones, and similarly L. × eurolepis (S21K9580102 × S21K9580032), were produced by controlled pollination. Seeds of L. kaempferi were produced by open pollination of known maternal clone (S08N1001) but unknown paternal clone. Seeds of both L. decidua and L. × eurolepis were produced in 2010 and that of L. kaempferi were produced in 1995. The seeds were stored in a freezer (−4 °C) from the time of harvest. A total of 336 seed samples, 112 samples per species, were randomly drawn from the total seed lots of each family to serve as working sub-samples for NIR analysis.
Spectral acquisition
NIR reflectance spectra of single seeds, expressed in the form of log (1/Reflectance), were collected with XDS Rapid Content Analyzer (FOSS NIRSystems, Inc.) from 400 to 2498 nm at an interval of 0.5 nm. Individual seeds were directly placed at the center of the scanning glass window of the instrument with 9 mm aperture at stationary module and then covered with the instrument’s lid that had a black background. Prior to collecting the NIR spectra of single seed, reference reflectance measurement was taken using the standard built-in reference of the instrument. In addition, reference measurements were taken after every 20 scans to reduce the effects of possible instrumental “drift”. For every seed, 32 monochromatic scans were made and the average value recorded.
Spectral data pre-treatment
NIR data are not usually amenable for direct analysis due to unwanted systematic variation that is not correlated with the chemical composition of the sample caused by instrumental drift, path length differences, base line shift and light scattering. This systematic noise in the spectra may be removed from the original NIR spectra to enhance signal to noise ratio using spectral data pre-treatment techniques; the most common ones in NIR spectroscopy being multiplicative signal correction (MSC), standard normal variate transformation (SNV), first and second derivatives (Rinnan et al. 2009). The MSC and SNV approaches reduce the scatter effect originating from sample physical variability while derivatives remove multiplicative effects in the spectra thereby reducing peak overlap and particle size between the samples. In this study, the original spectral data were pre-treated using these techniques, and the performance of the multivariate classification models were evaluated.
Multivariate classification modelling
Multivariate classification models were derived by partial least squares-discriminant analysis (PLS-DA) and bi-orthogonal projection to latent structures-discriminant analysis (O2PLS-DA). PLS-DA is a regularized version of linear discriminant analysis, in which a matrix of dummy variables that indicate class membership was used as regressand and the digitized NIR spectra as a descriptor matrix (Næs et al. 2002). Unlike PLS-DA modelling, O2PLS-DA modelling is a two-step approach where it first removes spectral variations that have no correlation with the classes and then fit the discriminant models based on predictive spectral variation. The spectral filtering step in O2PLS-DA was unique from the spectral treatments mentioned above as it took the response variable into account in its algorithm and removed more general types of interferences in the spectra by removing components orthogonal to the response variable calibrated against. To this end, the O2PLS-DA modelling approach used the information in the categorical response matrix Y (a matrix of dummy variables in our case) to decompose the X matrix (the spectral data) into three distinct parts: (1) the predictive score matrix and its loading matrix for X, (2) the corresponding Y-orthogonal score matrix and loading matrix of Y-orthogonal components, and (3) the residual matrix of X (Trygg and Wold 2003). Components orthogonal to the response variable containing unwanted systematic variation were then subtracted from the original spectral data to produce a filtered descriptor matrix. The final discriminant model was then computed using the filtered predictive spectral variations only.
Prior to modelling, the data set was divided into calibration and prediction sets. The calibration set was composed of 225 individual seeds (75 individual seeds per seed lot) and the prediction set was composed of 111 individual seeds (37 individual seeds per seed lot). Classification models were developed using absorbance values as regressor and a Y-matrix of dummy variables (1 for member of a given class, 0.0 otherwise) as regressand. The models were first fitted on both visible and NIR regions (400–2500 nm) and then on NIR region (780–2500 nm) only, and the performance of these models were compared. All calibrations were developed on mean-centered data sets and the number of significant model components were determined by a seven-segment cross validation (a default setting). A component was considered significant if the ratio of the prediction error sum of squares (PRESS) to the residual sum of squares of the previous dimension (SS) was statistically smaller than 1.0 (Eriksson et al. 2006). Finally, the computed models were used to classify samples in the prediction set, and seeds were considered as member of a given class if the predicted value was greater than an acceptance threshold (≥0.5) and all others were considered as non-members. Classification accuracy, expressed in percentage, was computed as the proportion of seeds predicted correctly as member of a given class to the total number of seeds in the prediction set for that class.
To analyze absorption bands that accounted for the discrimination of pure and hybrid larch seeds, a parameter called variable influence on projection (VIP) was computed as follows.
VIP for A components and K variables is a weighted sum of squares of the PLS weights (w) for a given component a and k variable, taking into account the amount of explained Y-variance (SSY) of a component, and SSY0 and SSYA are sum of squares of the response variable Y before and after extracting A number of components, respectively. Its major advantage is that there will be only one VIP-vector, summarizing all components and Y-variables, thereby enabling absorption bands that influence the discriminant models to be identified. As a rule, predictors with VIP value greater than 1.0 have a strong influence on the model, but a cut-off around 0.7–0.8 has been suggested to discriminate between relevant and irrelevant predictors (Eriksson et al. 2006). All calculations were performed using Simca-P + software (Version 13.0.0.0, Umetrics AB, Sweden).
Results
Spectral profile
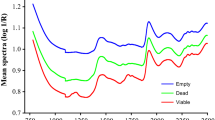
The average reflectance spectra, in the form of log (1/R), of Japanese and European larch seeds and their hybrid showed a similar profile with one major absorption peak in the visible region at 460 nm and three major peaks in the NIR region around 1470, 1924 and 2100 nm (Fig. 1). Absorbance values in the visible range were slightly higher for seeds of L. × eurolepis than seeds of the other two species. In the NIR region, seeds of L. decidua had higher absorbance values than seeds of L × eurolepis, which in turn had higher absorbance values than seeds of L. kaempferi. As a whole, the Vis + NIR spectra provided sufficient information to develop classification models.

Average raw spectra of L. decidua, L. kaempferi and their interspecific hybrid (L. × eurolepis) in reflectance spectroscopy mode
Classification of pure and hybrid larch seeds by PLS-DA modelling
PLS-DA models developed using Vis + NIR spectral range (400–2500 nm) required 9–15 significant components (A) to describe 91–94 % of the class variation (R2Y) in the calibration set, depending on the data set (Table 1A). The prediction power of the models according to cross-validation (\({{\text{Q}}^{2}}_{\text{cv}}\)) ranged from 85 to 87 %. For samples in the prediction set, the accuracy of predicted class membership for L × eurolepis was 100 % across all data sets, except the 2nd derivative data set where one seed sample was rejected as a non-member. Similarly, the accuracy of predicted class membership for L. decidua seeds was 97–100 %; and that of L. kaempferi was 95–97 %.
When the PLS-DA models were developed using the NIR region alone (780–2500 nm), the number of significant components to build the model was slightly lower than the previous models built using Vis + NIR region. However, the computed models still explained 86–94 % of the class variation for the calibration set with 80–87 % prediction ability according to cross-validation (Table 1B). For samples in the prediction set, the classification accuracy of pure and hybrid larch seeds did not change much compared to the model built using Vis + NIR region, except the 1st derivative data set that resulted in 13 % less classification accuracy for L. kaempferi (cf. 84 % in NIR and 97 % in Vis + NIR, Table 1).
Classification of pure and hybrid larch seeds by O2PLS-DA modelling
The O2PLS-DA models developed using the Vis + NIR had two predictive and 7–14 Y-orthogonal components, depending on the data set (e.g. A = 2 + 10 for untreated data set, Table 2). The predictive spectral variation (R2XP) accounted for 9–46 % of the total spectral variation of the pure and hybrid seed classes while the Y-orthogonal spectral variation (R2Xo) constituted 47–82 %, depending on the data set (Table 2A). The predictive spectral variations (R2XP), in turn, modelled more than 90 % of the variation between pure and hybrid seed classes (R2Y) in the calibration set for all but untreated data sets, with 83–90 % prediction accuracy (\({{\text{Q}}^{2}}_{\text{cv}}\)) according to cross validation (Table 2A). For models fitted using the NIR region alone, the two components were also required to build the models, but the Y-orthogonal components were slightly lower than the full spectrum models (Table 2B). The predictive spectral variation ranged from 5 to 28 %, which in turn modelled 77–90 % of the class variation with 74–88 % classification accuracy according to cross-validation (Table 2B). The modelled class variation (R2Y) and the predictive ability of the model (\({{\text{Q}}^{2}}_{\text{cv}}\)) were larger for pre-treated than untreated data sets, particularly for SNV-treated data set, irrespective of the wavelength region. As a whole, the model statistics showed that the NIR region alone contained substantial information that allowed hybrid larch seeds to be discriminated from pure parental larch seeds.
The O2PLS-DA models computed using SNV-treated data sets consistently assigned L. decidua and L. kaempferi seeds in the prediction set to their respective classes with 100 % accuracy in both Vis + NIR and NIR regions, while the classification accuracy for L × eurolepis seeds was 97 % in the NIR region and 100 % in Vis + NIR region (Fig. 2). The model fitted on MSC-treated data set also resulted in complete recognition of pure and hybrid seeds in the Vis + NIR region, as also the model fitted on 1st derivative data set, except for the L. kaempferi seeds that were classified with 97 % accuracy (Table 3). In the NIR region alone, the classification accuracy was slightly less, particularly for MSC-treated data set where the accuracy was 11, 8 and 3 % less than that in the Vis + NIR region for L. decidua, L. kaempferi and L × eurolepis, respectively.

Class membership of L. decidua (a), L. × eurolepis (b) and L. kaempferi (c) seeds in the prediction set (n = 37 for each seed lot) predicted by O2PLS model developed using SNV-transformed data set according to their class. Note that the red dashed line is threshold for classification. (Color figure online)
To get a better insights into the modelling process, score and loading plots for O2PLS-DA model fitted on SNV-treated data set were further examined (note that this model had consistently higher prediction accuracy in both Vis + NIR and NIR regions than models fitted on other data sets). The score plot (t[1] vs. t[2]) showed a clear separation of L. decidua seed lot from the other two seed lots along the first predictive component, while L × eurolepis seed lot was clearly separated from the pure larch seed lots along the second component (Fig. 3a). The corresponding predictive loading plot for the first component revealed that one sharp peak at 410 nm and four broad absorption bands in 1409–1630, 1886–1996, 2019–2190 and 2230–2410 nm that were useful to discriminate L. decidua seed lot from the other seed lots (Fig. 3b). The loading plot for the second predictive component also showed one sharp peak at 460 nm and two broad absorption bands in 840–1190 and 1217–1620 nm that were mainly accounted for discriminating L × eurolepis seed lot from the pure parental seed lots, while an absorption peak at 638 nm was mainly accounted for discriminating L. kaempferi from the L × eurolepis seed lot (Fig. 3c).

Score plot for the first two predictive components (t1 vs. t2) of O2PLS-DA model built using SNV-transformed spectra, depicting clear-cut separation of seeds of European (green circle), Japanese (red triangle) and hybrid (blue inverted triangle) larch (a) together with loading plots for the first (b) and second predictive components (c) showing absorption bands accounted for class discrimination. Note that the ellipse in a is 95 % confidence interval according to Hotelling’s T 2 test (a multivariate generalization of Student’s t test). (Color figure online)
To determine which phenomena were irrelevant for discriminating seeds of pure and hybrid larch and in which spectral region they occurred, the score and loading plots of the first two Y-orthogonal components, accounting for 52 % of the spectral variation, were examined. The score plots for the first two Y-orthogonal components showed few samples fell outside of the 95 % confidence ellipse (Fig. 4a) according to Hotelling’s T 2 test (a multivariate generalization of Student’s t test), but excluding these outlying samples did not improve the model. The corresponding loading plots showed that small peaks centered at 415, 687, 1456, 1929 and 2111 nm in the first component (Fig. 4b); and peaks centered at 484, 771, 1453, 1929 and 2110 nm in the second component (Fig. 4c) were accounted for spectral variations uncorrelated to class discrimination. Thus, the sources of irrelevant spectral variation explained by the model were related to divergence in color, moisture content and storage reserves among individual seeds within each class, which in turn was related with individual seed size variation.

Orthogonal Score plot of O2PLS-DA model built using SNV-treated spectra, showing few outliers of European (green circle), Japanese (red triangle) and hybrid (blue inverted triangle) larch seeds (a) together with the corresponding loading plots for the first (b) and second orthogonal components (c) showing absorption bands. Note that the ellipse is 95 % confidence interval according to Hotelling’s T 2 test. (Color figure online)
Absorption bands accounted for classification of pure and hybrid larch seeds
The VIP plot shows that absorption bands in 400–750 nm, with two major peaks centered around 460 and 638 nm and two shoulder peaks in the vicinity of 415 and 687 nm had a strong influence on the discrimination of pure and hybrid larch seeds (VIP > 1; Fig. 5). In the NIR region, absorption bands in 1890–2201 and 2245–2500 nm, with peaks centered at 1929, 2098, 2332 and 2490 nm also accounted for class discrimination. Other NIR regions of interest that helped improve class discrimination appeared in the 860–1380, 1410–1505 and 2240–2388 nm (VIP = 0.81 − 1.0).

VIP plot for the O2PLS model built on SNV-treated data set in 400–2500 nm wavelength region. The threshold of significant contribution in model building is shown by red dashed line. (Color figure online)
Discussion
The results demonstrated that pure and hybrid larch seeds can be successfully distinguished by Vis + NIR spectroscopy and multivariate modelling. PLS-DA models fitted on both raw and pre-treated spectral data sets resulted in comparable classification accuracy for samples in the prediction set. However, spectral pre-treatments slightly reduced the number of components needed to build the models, which might be attributed to the removal of scatter effect to some extent (Rinnan et al. 2009). On the contrary, the O2PLS-DA modelling approach needed only two predictive components to successfully classify pure and hybrid larch seeds. In particular, the classification model fitted on SNV-treated data set had excellent prediction ability (Q2) compared to other data sets (note that a model with Q2 ≥ 0.9 is considered excellent sensu Eriksson et al. 2006). This indicates that not only the scatter effect but also other sources of spectral noise that are not correlated with class discrimination exist.
Analyses of score and loading plots of Y-orthogonal components revealed some outlying samples from each seed lot, and absorption peaks at 416, 680, 1456, 1930 and 2110 nm that accounted for spectral variation uncorrelated with class discrimination (Fig. 4b, c). Absorption bands in 1400–1500 and 1900–2000 nm with peaks at 1450 and 1940 nm were likely attributed to the presence of water due to O–H stretch first overtone and combination bands involving O–H stretch and O–H (Osborne et al. 1993; Tigabu and Oden 2003). The absorption band in 2050–2200 nm is characteristic of CH2 stretch-bending combinations and correlates positively with fatty acid composition (Hourant et al. 2000). Apparently differences in color, moisture content and fatty acids among individual seeds within each seed lot can possibly cause spectral variations that have no correlation to class discrimination. By extracting these spectral variations irrelevant to class discrimination, the O2PLS-DA modelling approach resulted in dimensionally less complex and parsimonious models (A = 2 for O2PLS-DA vs. A = 15 for PLS-DA). Dimensional complexity is an important factor in the interpretation of multivariate analysis and parsimonious models with few components are often highly preferred (Trygg and Wold 2003).
In the visible region, absorption maxima that were accounted for discriminating L. decidua, L × eurolepis and L. kaempferi seeds appeared at 410, 460 and 638 nm, respectively (Figs. 3b, c, 5). Apparently seeds of L. kaempferi appear to be more red-brownish than L. decidua and L. × eurolepis seeds, which in turn vary slightly in color. As the seed coat and the megagametophyte (storage organ) account for more than half of the total seed mass and are of maternal origin, the chemical composition of the seed coat would presumably be influenced more by the genotype of the maternal than paternal parents. It should be noted that the maternal parent for the hybrid larch in the present study was L. decidua while the paternal parent was L. kaempferi. Many conifers also exhibit genotypic variation in seed physical traits, such as surface structure of seeds (Tillman-Sutela et al. 1998), seed size and germinability (Mamo et al. 2006) as well as qualitative color characteristics of the seed coat (Tillman-Sutela and Kauppi 1995); thus color variation might be expected among seed lots investigated in the present study. Our finding for Larix agrees with previous studies that have demonstrated the efficacy of the visible region for classifying wheat kernels according to their color (Wang et al. 1999) and in identifying seed origin and parents of P. sylvestris L. trees (Tigabu et al. 2005).
In NIR region, absorption bands in 1409–1630, 1886–1996, 2019–2190 and 2230–2410 nm were highly relevant for discriminating L. decidua seeds from L × eurolepis and L. kaempferi seeds (Figs. 3b, 5). The 1409–1630 nm region of the NIR reflectance spectra presents two broad peaks at 1480 and 1550 nm, which corresponds to first overtone of O–H and N–H and combination band of C–H vibration of various functional groups; notably ROH, starch, H2O and protein moieties reported in other studies (Workman and Weyer 2012). The absorption band in 1900–2000 nm with absorption peak centered at 1929 nm may arise from C=O stretch second overtone, combination of O–H stretch and HOH deformation, and O–H bend second overtone. The results of previous studies have shown that molecular moieties of protein, starch and water show overlapping absorption peaks in this region (Shenk et al. 2001; Workman and Weyer 2012). The absorption bands in 2019–2190 and 2230–2410 nm are characteristic of CH2 stretch-bend combinations as well as other vibrational modes of molecular bonds (Workman and Weyer 2012). Several fatty acids, notably polyunsaturated fatty acids, in several oil crops have shown positive correlation to absorption bands in these regions (Hourant et al. 2000; Osborne et al. 1993). Tigabu and Oden (2003) also found correlations between absorbance values in these spectral regions and major fatty acids as a basis for discrimination of viable and empty seeds of Pinus patula Schiede and Deppe.
Thus, NIR spectroscopy appears to have detected differences in chemical compounds between the two species and the hybrid, probably reserve compounds like lipids and proteins as well as moisture content differences, thus allowing L. decidua seeds to be distinguished from seeds of L × eurolepis and L. kaempferi. Fatty acids such as linoleic, Δ5-olefinic, pinolenic and oleic acids were the major composition in seeds of larch species that contributed to discriminating filled-viable, empty and insect-attacked seeds of three larch species in another study (Tigabu and Oden 2004). It should be noted that lipids are the dominant storage reserve compounds in the seeds of many conifers including those of larch; and the major fatty acids include linoleic, Δ5-olefinic, pinolenic and oleic acids that account for 43.1, 30.6, 27.4 and 18.8 % of the total fatty acids, respectively in L. decidua seeds while linoleic acid accounts for 45.5 %, Δ5-olefinic acid for 28.9 %, pinolenic acid for 25.8 % and oleic acids for 18.4 % of the total fatty acids in seed lipids of L. kaempferi (Wolff et al. 1997, 2001).
Absorption bands in 840–1190 and 1217–1620 nm regions allowed discrimination between L × eurolepis seed lots and the pure parental seed lots. The absorption bands in these regions are characteristic of the third overtone of C–H stretching vibration, combination of N–H second overtone stretching vibration and C–H stretch and deformation (Osborne et al. 1993; Shenk et al. 2001; Workman and Weyer 2012). Functional groups responsible for absorption in these regions are mainly CH3, CH2, ArNH2 (aromatic amino acids) and NH2, which are common molecular moieties of fatty acids and proteins Thus, it appears that NIR spectroscopy has utilized differences in fatty acids and proteins as a basis for discriminating seeds of L × eurolepis from L. kaempferi and L. decidua. This divergence in seed storage reserves between hybrid and pure parental (particularly L. kaempferi) seeds is expected because the contribution of the paternal parent (which is L. kaempferi in this study) to the total seed mass is much lower than that of the maternal parent. The embryo (a smaller fraction of the seed mass) is derived from both parents while more than half of the seed mass is of maternal origin. Maternal variation in seed storage reserves is also evident as reproductive allocation in plants is generally governed by the genetic constitution (see review, Bazzaz et al. 2000). Tigabu et al. (2005) have found maternal variation in storage reserves as the basis for identifying among maternal parents of Scots pine using NIR spectra.
Conclusions
Classification of pure and hybrid larch seeds using NIR spectroscopy was successful. Thus, the results highlight the feasibility of NIR spectroscopy as a non-destructive method for verification of hybrid larch seeds. The technique is rapid and efficient as it takes ca. 2 min to scan a single seed, non-destructive and there is no need for sample preparation. Moreover, the OPLS-DA modelling approach results in the development of parsimonious models that provide additional information that allow within-class variation to be explained. From practical viewpoint, the technique can offer a unique opportunity for seed orchard managers to rapidly estimate the hybrid seed yield from open pollinated mixed species seed orchards. Breeders can also benefit from use of the NIR technique to assess the efficiency of artificial pollination in seed orchard management research. As a whole, NIR spectroscopy is a very promising method which should be verified using seeds from many different families before recommending its use in operational practice.
References
Acheré V, Rampant PF, Benoît V, Leplé J, Pâques L E, Prat D (2002) Characterization of hybrids between Larix decidua and Larix kaempferi by molecular markers. In: Improvement of larch (Larix Sp.) for better growth, stem form and wood quality. Proceedings of an international symposium, INRA, Unité d’Amélioration, Gap (Hautes-Alpes)-Auvergne & Limousin, France, 16–21 September, 2002, pp 176–186
Acheré V, Faivre-Rampant P, Pâques LE, Prat D (2004) Chloroplast and mitochondrial molecular tests identify European × Japanese larch hybrids. Theor Appl Genet 108:1643–1649. doi:10.1007/s00122-004-1595-y
Baltunis BS, Greenwood MS, Eysteinsson T (1998) Hybrid vigor in Larix: growth of intra- and interspecific hybrids of Larix decidua, L. laricina, and L. kaempferi after 5 years. Silvae Genet 47:288–293
Bazzaz FA, Ackerly DD, Reekie EG (2000) Reproductive allocation in plants. In: Fenner M (ed) Seeds: the ecology of regeneration in plant communities. Wallingford, New York, pp 1–29
Bertrand B, Etienne H, Lashermes P, Guyot B, Davrieux F (2005) Can near-infrared reflectance of green coffee be used to detect introgression in Coffea arabica cultivars? J Sci Food Agric 85:955–962. doi:10.1002/jsfa.2049
Castillo R, Otto M, Freer J, Valenzuela S (2008) Multivariate strategies for classification of Eucalyptus globulus genotypes using carbohydrates content and NIR spectra for evaluation of their cold resistance. J Chemom 22:268–280. doi:10.1002/cem.1126
Ekö PM, Larsson-Stern M, Albrektson A (2004) Growth and yield of hybrid larch (Larix × eurolepis A. Henry) in southern Sweden. Scand J For Res 19:320–328. doi:10.1080/02827580410024151
Eriksson L, Johansson E, Kettaneh-Wold N, Trygg J, Wikström C, Wold S (2006) Multi- and megavariate data analysis. Umetrics Academy, Umeå
Espinoza JA, Hodge GR, Dvorak WS (2012) The potential use of near infrared spectroscopy to discriminate between different pine species and their hybrids. J Near Infrared Spectrosc 20:437–447. doi:10.1255/jnirs.1006
Gauchat ME, Pâques LE (2011) Indirect prediction of bud flushing from ex situ observation in hybrid Larch (Larix decidua × L. kaempferi) and their parents. Environ Exp Bot 70:121–130. doi:10.1016/j.envexpbot.2010.08.001
Hannerz M, Hajek J, Stener L-G, Werner M (1993) Lärkfröplantager i Sverige. Resultat nr 8. Skogforsk (In Swedish, English summary)
Helmersson A, Westin J, Stener L-G (2015) Larch—state of knowledge, regeneration material and tree breeding possibilities. Skogforsk (In Swedish, English summary)
Hourant P, Baeten V, Morales MT, Meurens M, Aparicio R (2000) Oil and fat classification by selected bands of near-infrared spectroscopy. Appl Spectrosc 54:1168–1174. doi:10.1366/0003702001950733
Humphreys JR, O’Reilly-Wapstra J, Harbard J, Davies NW, Griffin AR, Jordan GR, Potts BM (2008) Discrimination between seedlings of Eucalyptus globulus, E. nitens and their F1 hybrid using near-infrared reflectance spectroscopy and foliar oil content. Silvae Genet 57:262–269
Mamo N, Mihretu M, Fekadu M, Tigabu M, Teketay D (2006) Variation in seed and germination characteristics among Juniperus procera populations in Ethiopia. For Ecol Manag 225:320–327. doi:10.1016/j.foreco.2006.01.026
Næs T, Isaksson T, Fearn T, Davies T (2002) A user friendly guide to multivariate calibration and classification. NIR Publications, Chichester
Osborne BG, Fearn T, Hindle PT (1993) Practical NIR spectroscopy with applications in food and beverage analysis. Harlow, New York
Pâques LE (2000) Interspecific hybridisation in larch: the long way to get outstanding varieties. In: Dungey HS, Dieters MJ, Nikles DG (Comp) Hybrid breeding and genetics of forest trees. Noosa, Queensland, pp 373–385
Pâques LE (2004) Role of European and Japanese larch in the genetic control of growth, architecture and wood quality traits in interspecific hybrids (Larix × eurolepis Henry). Ann For Sci 61:25–33. doi:10.1051/forest:2003081
Pâques LE (2009) Purity control of seed lots or seedling batches of hybrid larch (Larix decidua × L. kaempferi). http://treebreedex.eu/
Pâques LE, Foffová E, Heinze B, Lelu-Walter M-A, Liesebach M, Philippe G (2013) Larches (Larix Sp.). In: Pâques LE (ed) Forest tree breeding in Europe, vol 25. Springer, Netherlands, pp 13–122
Rinnan Å, Van Den Berg F, Engelsen SB (2009) Review of the most common pre-processing techniques for near-infrared spectra. TrAC-Trends Analyt Chem 28:1201–1222. doi:10.1016/j.trac.2009.07.007
Sato M, Seki K, Kita K, Moriguchi Y, Yunoki K, Kofujita H, Ohnishi M (2008) Prominent differences in leaf fatty acid composition in the F1 hybrid compared with parent trees Larix gmelinni var. japonica and L. kaempferi. Biosci Biotechnol Biochem 72:2895–2902. doi:10.1271/bbb.80386
Seregély Z, Deák T, Bisztray GD (2004) Distinguishing melon genotypes using NIR spectroscopy. Chemom Intell Lab Sys 72:195–203. doi:10.1016/j.chemolab.2004.01.013
Shenk JS, Workman JJ, Westerhaus MO (2001) Application of NIR spectroscopy to agricultural products. In: Burns DA, Ciurczak EW (eds) Handbook of near-infrared spectroscopy, 2nd edn. Marcel Dekker, New York, pp 419–474
Tigabu M, Oden PC (2003) Discrimination of viable and empty seeds of Pinus patula Schiede & Deppe with near-infrared spectroscopy. New For 25:163–176. doi:10.1023/A:1022916615477
Tigabu M, Oden PC (2004) Simultaneous detection of filled, empty and insect-infested seeds of three Larix species with single seed near-infrared transmittance spectroscopy. New For 27:39–53. doi:10.1023/A:1025037408922
Tigabu M, Oden PC, Lindgren D (2005) Identification of seed sources and parents of Pinus sylvestris L. using visible–near infrared reflectance spectra and multivariate analysis. Trees 19:468–476. doi:10.1007/s00468-005-0408-5
Tillman-Sutela E, Kauppi A (1995) The significance of structure for the imbibition in seeds of the Norway spruce, Picea abies (L.) Karst. Trees 9:269–278. doi:10.1007/BF00202017
Tillman-Sutela E, Kauppi A, Sahlén K (1998) Effect of disturbed photoperiod on the surface structures of ripening Scots pine (Pinus sylvestris L.) seeds. Trees 12:499–506. doi:10.1007/PL00009731
Trygg J, Wold S (2003) O2-PLS, a two-block (X–Y) latent variable regression (LVR) method with an integral OSC filter. J Chemom 17:53–64. doi:10.1002/cem.775
Wang D, Dowell FE, Lacey RE (1999) Single wheat kernel color classification using near-infrared reflectance spectra. Cereal Chemom 76:30–33. doi:10.1094/CCHEM.1999.76.1.30
Wolff RL, Comps B, Marpeau AM, Deluc LG (1997) Taxonomy of Pinus species based on the seed oil fatty acid composition. Trees 12:113–118. doi:10.1007/PL00009698
Wolff RL, Lavialle O, Pédrono F, Pasquier E, Deluc LG, Marpeau AM, Aitzetmüller K (2001) Fatty acid composition of Pinaceae as taxonomic markers. Lipids 36:439–451. doi:10.1007/s11745-001-0741-5
Workman J, Weyer L (2012) Practical guide and spectral atlas for interpretive near-infrared spectroscopy. Boka Raton, Florida
Zhang SG, Yang WH, Han SY, Han BT, Li MX, Qi LW (2010) Cytogenetic analysis of reciprocal hybrids and their parents between Larix leptolepis and Larix gmelinii: implications for identifying hybrids. Tree Genet Genomes 6:405–412. doi:10.1007/s11295-009-0258-1
Acknowledgments
The Karl Erik Önnesjös Foundation is acknowledged for financial support of this research. The anonymous reviewers and the associate editor are gratefully acknowledged for their valuable comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Farhadi, M., Tigabu, M., Stener, LG. et al. Feasibility of visible + near infrared spectroscopy for non-destructive verification of European × Japanese larch hybrid seeds. New Forests 47, 271–285 (2016). https://doi.org/10.1007/s11056-015-9514-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-015-9514-4