
Abstract
Neural mass models are successful in modeling brain rhythms as observed in macroscopic measurements such as the electroencephalogram (EEG). While the synaptic current is explicitly modeled in current models, the single cell electrophysiology is not taken into account. To allow for investigations of the effects of channel pathologies, channel blockers and ion concentrations on macroscopic activity, we formulate neural mass equations explicitly incorporating the single cell dynamics by using a bottom-up approach. The mean and variance of the firing rate and synaptic input distributions are modeled. The firing rate curve (F(I)-curve) is used as link between the single cell and macroscopic dynamics. We show that this model accurately reproduces the behavior of two populations of synaptically connected Hodgkin-Huxley neurons, also in non-steady state.










Similar content being viewed by others

Notes
The simulation code is available from modelDB (Hines et al. 2004), accession nr. 155130 and Researchgate www.researchgate.net/profile/Bas-Jan_Zandt
Available from ModelDB, accession nr. 154739.
References
Allen, C., & Stevens, C.F. (1994). An evaluation of causes for unreliability of synaptic transmission. Proceedings National Academy Science USA, 383 (10), 380–10.
Amit, D., & Brunel, N. (1997). Dynamics of a recurrent network of spiking neurons before and following learning Network Computation in Neural Systems.
Baladron, J., Fasoli, D., Faugeras, O., Touboul, J. (2012). Mean-field description and propagation of chaos in networks of hodgkin-huxley and fitzhugh-nagumo neurons. Journal Mathematics Neuroscience, 2 (1), 10. doi:10.1186/2190-8567-2-10.
Bazhenov, M., Timofeev, I., Frhlich, F., Sejnowski, T.J. (2008). Cellular and network mechanisms of electrographic seizures. Drug Discovery Today Dis Models, 5 (1), 45–57. doi:10.1016/j.ddmod.2008.07.005.
Bhattacharya, B.S., Coyle, D., Maguire, L.P. (2011). A thalamo-cortico-thalamic neural mass model to study alpha rhythms in Alzheimer’s disease. Neural networks : the official journal of the International Neural Network Society, 24 (6), 631–45. doi:10.1016/j.neunet.2011.02.009.
Chizhov, A., & Graham, L. (2007). Population model of hippocampal pyramidal neurons, linking a refractory density approach to conductance-based neurons. Physical Review E, 75(1) (011), 924. doi:10.1103/PhysRevE.75.011924.
De Schutter, E. (2010). Computational Modeling Methods for Neuroscientists. Mit Press, chap, 6. URL http://books.google.nl/books/about/Computational_Modeling_Methods_for_Neuro.html?id=20RvPgAACAAJ&pgis=1.
Deco, G., Jirsa, V.K., Pa, Robinson, Breakspear, M., Friston, K. (2008). The dynamic brain: from spiking neurons to neural masses and cortical fields. PLoS computational biology, 4(8) (e1000), 092. doi:10.1371/journal.pcbi.1000092.
Dreier, J.P. (2011). The role of spreading depression, spreading depolarization and spreading ischemia in neurological disease. Nature medicine, 17 (4), 439–47. doi:10.1038/nm.2333.
Faugeras, O., Touboul, J., Cessac, B. (2009). A constructive mean-field analysis of multi-population neural networks with random synaptic weights and stochastic inputs. Frontiers in computational neuroscience, 3 (February), 1. doi:10.3389/neuro.10.001.2009.
Fröhlich, F., Bazhenov, M., Iragui-Madoz, V., Sejnowski, T.J. (2008). Potassium dynamics in the epileptic cortex: new insights on an old topic. Neuroscientist, 14 (5), 422–433. doi:10.1177/1073858408317955.
Galtier, M.N., & Touboul, J. (2013). Macroscopic equations governing noisy spiking neuronal populations with linear synapses. PLoS One, 8(11) (e78), 917. doi:10.1371/journal.pone.0078917.
Grefkes, C., & Fink, G.R. (2011). Reorganization of cerebral networks after stroke: new insights from neuroimaging with connectivity approaches. Brain : a journal of neurology, 134 (Pt 5), 1264–76. doi:10.1093/brain/awr033.
Hindriks, R., & Putten, van MJaM (2012). Meanfield modeling of propofol-induced changes in spontaneous EEG rhythms. NeuroImage, 60 (4), 2323–34. doi:10.1016/j.neuroimage.2012.02.042.
Hines, M.L., Morse, T., Migliore, M., Carnevale, N.T., Shepherd, G.M. (2004). Modeldb: A database to support computational neuroscience. Journal Computational Neuroscience, 17 (1), 7–11. doi:10.1023/B:JCNS.0000023869.22017.2e.
Hocepied, G., Legros, B., Van Bogaert, P., Grenez, F., Nonclercq, A. (2013). Early detection of epileptic seizures based on parameter identification of neural mass model. Computer Biology Medicine, 43 (11), 1773–1782. doi:10.1016/j.compbiomed.2013.08.022.
Holt, G. (1997). A critical reexamination of some assumptions and implications of cable theory in neurobiology. PhD thesis: California Institute of Technology. URL http://lnc.usc.edu/holt/papers/thesis/.
Hutt, A. (2012). The population firing rate in the presence of GABAergic tonic inhibition in single neurons and application to general anaesthesia. Cognitive neurodynamics, 6 (3), 227–37. doi:10.1007/s11571-011-9182-9.
Hutt, A. (2013). The anesthetic propofol shifts the frequency of maximum spectral power in eeg during general anesthesia: analytical insights from a linear model. Front Computational Neuroscience, 7, 2. doi:10.3389/fncom.2013.00002.
Izhikevich, E.M. (2007). Dynamical Systems in Neuroscience The Geometry of Excitability and Bursting: Computational neuroscience, vol First. MIT Press. URL http://www.amazon.com/dp/0262514206.
Jansen, B.H., & Rit, V.G. (1995). Electroencephalogram and visual evoked potential generation in a mathematical model of coupled cortical columns. Biology Cybernetics, 73 (4), 357–366.
Liley, D.T.J., Cadusch, P.J., Dafilis, M.P. (2002). A spatially continuous mean field theory of electrocortical activity. Network (Bristol England), 13 (1), 67–113.
Manwani, A., & Koch, C. (1999). Detecting and estimating signals in noisy cable structure, i: neuronal noise sources. Neural Computation, 11 (8), 1797–1829.
Marreiros, A.C., Daunizeau, J., Kiebel, S.J., Friston, K.J. (2008). Population dynamics: variance and the sigmoid activation function. NeuroImage, 42 (1), 147–57. doi:10.1016/j.neuroimage.2008.04.239.
Meisler, M.H., & Kearney, J.A. (2005). Sodium channel mutations in epilepsy and other neurological disorders. Journal Clinical Investigation, 115 (8), 2010–2017. doi:10.1172/JCI25466.
Moran, R., Pinotsis, D.A., Friston, K. (2013). Neural masses and fields in dynamic causal modeling. Front Computational Neuroscience, 7, 57. doi:10.3389/fncom.2013.00057.
Ostojic, S., & Brunel, N. (2011). From spiking neuron models to linear-nonlinear models. PLoS computational biology, 7(1) (e1001), 056. doi:10.1371/journal.pcbi.1001056.
van Putten M.J., & Zandt B.J. (2013). Neural Mass modeling for predicting seizures. Clinical Neurophysiology. doi:10.1016/j.clinph.2013.11.013.
Robinson, P.A., Rennie, C.J., Wright, J.J., Bahramali, H., Gordon, E., Rowe, D.L. (2001). Prediction of electroencephalographic spectra from neurophysiology. Physics Review E Statistics Nonlinear Soft Matter Physics, 63(2 Pt 1) (021), 903.
Pa, Robinson, Wu, H., Kim, J.W. (2008). Neural rate equations for bursting dynamics derived from conductance-based equations. Journal of theoretical biology, 250 (4), 663–72. doi:10.1016/j.jtbi.2007.10.020.
Schevon, C.A., Ng, S.K., Cappell, J., Goodman, R.R., McKhann, G Jr, Waziri, A., Branner, A., Sosunov, A., Schroeder, C.E., Emerson, R.G. (2008). Microphysiology of epileptiform activity in human neocortex. Journal Clinical Neurophysiol, 25 (6), 321–330. doi:10.1097/WNP.0b013e31818e8010.
Shriki, O., Hansel, D., Sompolinsky, H. (2003). Rate models for conductance-based cortical neuronal networks. Neural computation, 15 (8), 1809–41. doi:10.1162/08997660360675053.
Somjen, G. (2004). Ions in the Brain: Normal Function, Seizures and Stroke. USA: Oxford University Press. http://books.google.nl/books?id=yaNrAAAAMAAJ.
Somjen, G.G. (2001). Mechanisms of spreading depression and hypoxic spreading depression-like depolarization. Physiology Reviews, 81 (3), 1065–1096.
Stead, M., Bower, M., Brinkmann, B.H., Lee, K., Marsh, W.R., Meyer, F.B., Litt, B., Van Gompel, J., Worrell, G.A. (2010). Microseizures and the spatiotemporal scales of human partial epilepsy. Brain, 133 (9), 2789–2797. doi:10.1093/brain/awq190.
Tjepkema-Cloostermans, M.C., Hindriks, R., Hofmeijer, J., van Putten MJAM (2013). Generalized periodic discharges after acute cerebral ischemia: Reflection of selective synaptic failure?. Clinical Neurophysiol. doi:10.1016/j.clinph.2013.08.005.
Touboul, J., Hermann, G., Faugeras, O. (2012). Noise-induced behaviors in neural mean field dynamics. SIAM Journal on Applied Dynamical Systems, 11 (1), 49–81. doi:10.1137/110832392. http://epubs.siam.org/doi/pdf/10.1137/110832392.
Victor, J.D., Drover, J.D., Conte, M.M., Schiff, N.D. (2011). Mean-field modeling of thalamocortical dynamics and a model-driven approach to EEG analysis. Proceedings of the National Academy of Sciences of the United States of America 108 Suppl, 15, 631–8. doi:10.1073/pnas.1012168108.
Visser, S., & Van Gils, S. (2014). Lumping Izhikevich neurons. EPJ Nonlinear Biomedical Physics, 2 (1), 6. doi:10.1140/epjnbp19. URL http://www.epjnonlinearbiomedphys.com/content/2/1/6.
Wilson, H.R., & Cowan, J.D. (1972). Excitatory and inhibitory interactions in localized populations of model neurons. Biophysical journal, 12 (1), 1–24. doi:10.1016/S0006-3495(72)86068-5.
Zandt, B.J., ten Haken, B., van Dijk, J.G., van Putten, M J.A.M. (2011). Neural dynamics during anoxia and the ”wave of death”. PLoS One, 6(7) (e22), 127. doi:10.1371/journal.pone.0022127.
Ziburkus, J., Cressman, J.R., Barreto, E., Schiff, S.J. (2006). Interneuron and pyramidal cell interplay during in vitro seizure-like events. Journal Neurophysiology, 95 (6), 3948–3954. doi:10.1152/jn.01378.2005.
Acknowledgements
This work was financially supported by Ministerie van Economische Zaken, Provincie Overijssel and Provincie Gelderland through the ViPBrainNetworks project. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Action Editor: Michael Breakspear
Appendices
Appendix A: Distribution of the synaptic input dynamics and distribution of the synaptic input in a network of spiking cells
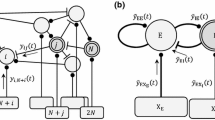
A network with excitatory and inhibitory neurons, contains four sets of synaptic connections (e →e, e →i, i →e and i →i). We describe the total synaptic conductance G n induced by one of these sets on a postsynaptic neuron n (see Fig. 11). First, the distribution of G is derived from a synaptic connection matrix. We denote the number of cells in the presynaptic population with M, and that in the postsynaptic one with N. This yields a maximum of N×M synapses in the set. The presence and strength of the synaptic connections is described with an N×M matrix W. W n,m =1 denotes a synaptic connection from cell m to n of average weight, while 0 stands for no connection. This system is equal to that analyzed by Faugeras et al. (Faugeras et al. 2009), however we do not constrain our analysis to a normal distribution of W n,m .

Sketch of the synaptic variables. One synaptic population is shown, with presynaptic single cell firing rates f m , synaptic conductances g m and summed synaptic conductances G n
We assume the states of the synapses originating from the same presynaptic cell m are all equal, except for their weight. They are therefore described with the same variable g m (t). The conductance of the synapse g m (t) is described as the convolution of the impulse response H with the spike rate f m (t) of cell m:
The total synaptic conductance G n of cell n induced by one presynaptic population, is calculated as the linear sum of all synapses, using the connection matrix:
Assuming the firing rates and synaptic strengths are uncorrelated, and defining the weighted number of connections neuron n receives as
the mean of the total synaptic conductance \(\bar {g}\) is readily calculated:
, using bars to denote the mean values.
This derivation may seem an unnecessarily elaborate way of obtaining the familiar Eq. (28), but it sets the stage for deriving the variance of G.
To calculate the variance of G n Eq. (26) over the N postsynaptic cells , the terms W n,m and g m are written as their mean plus a deviation:
and similar for g. With these expressions, the variance of G n Eq. (26) is calculated:
The subscript n denotes the variance is calculated over n. The first two terms are zero, since the sums over the deltas are per definition zero. Hence,
Assuming the synaptic weights and synaptic activations are uncorrelated and using the standard rules for the variance of products and sums:
noting that \(M\bar {W} = N_{\text {syn}}\) (equation 27), \(\text {var}_{m}(g_{m}) \equiv {\sigma _{g}^{2}}\) and defining
this is results in:
The first term is the variance due to differences in the weighted number of synaptic connections N syn received by the cells. The second term is the variance due to different synaptic activations caused by differences in presynaptic spike rates. \(N_{\text {syn}}^{\prime }\) can be interpreted as the average number of connections a postsynaptic neuron does not share with a random other cell of its population. For example, in the case where all cells are connected with probability p it is equal to M p−M p 2, where M p is equal to the average number of connections per neuron, and M p 2 the average number of connections shared with another neuron. Note that these expressions do not require the number of synapses to approach infinity.
Now the task to describe the mean and variance of g(t) is left. To calculate σ g over the individual synapses, we need to make some assumptions on the distribution of the spikes in the input. Amit and Brunel calculated this variance in the steady state, assuming Poisson (shot noise) statistics (Amit and Brunel 1997). We will show a similar approach can be used for dynamic input and describe statistics for periodically generated action potentials, i.e. with regular intervals. These describe the firing rates observed in our spiking network model better.
There is a fundamental difference between calculating the distribution of the conductance induced by a Poisson process or by a periodical process. The conductance of a synapse receiving action potentials generated by a Poisson process with a known rate, is a stochastic signal. In contrast, the conductance of a synapse receiving action potentials periodically at a known rate is deterministic (assuming the synaptic integration smooths the input sufficiently, such that the phase of the input is irrelevant). Therefore, the variance of the synaptic conductances is induced by the variance of the firing rates themselves, rather than by the realization of the spike generation process. We calculate this variance now for time dependent firing rates.
The firing rates f(t) are considered to be distributed over the presynaptic neurons, with mean \(\bar {f}(t)\) and standard deviation σ f (t). We consider the inputs to be smooth continuous functions (rates) rather than delta pulse trains, as is common in neural mass modeling.
The mean of the synaptic activation is trivial to calculate from Equation (25):
where the brackets denote the expectancy.
To calculate the variance of g, we use hats to denote the deviation from the expectancy of f and g, i.e.
and explicitly write the convolutions as integrals:
The expectancy is replaced by the (auto)correlation coefficient C ac:
In our simulations of spiking networks it was observed that \(\hat {f}\) is highly correlated with itself over periods much longer than the duration of H, i.e. cells tend to fire faster or slower than the population average over longer periods of time. In that case C ac≈1 and the right hand side of Equation (44) is equal to the squared convolution with the variance (c.f. Equations (40-42)),
which is used for our simulations.
For cases where \(\hat {f}\) does fluctuate faster (typically when var (N syn) is low), Equation (44) can be simplified in another way. It is reasonable to assume the autocorrelation coefficient depends only on the time difference u−u ′, while furthermore σ f fluctuates slowly compared to the synaptic time constant. In that case, the autocorrelation coefficient can be effectively replaced with a correction constant C between 0 and 1. The expression for σ g becomes:
where
However, deriving a closed expression for the autocorrelation coefficient C ac(u−u ′) in a recurring network is complicated and outside the scope of this work.
Appendix B: Approximating a synaptic conductance with a constant current
We show how a synaptically induced (excitatory) current can be approximated with a constant input. This approach can be easily extended to include a second (inhibitory) conductance as well. First, we write the dynamics of a Hodgkin-Huxley (HH) model with the added synaptic conductance as:
Here I HH is the sum of all gated and leak currents in the HH model, g syn is the synaptic conductance and E syn the reversal potential of the synapse. The synaptic current was split in a constant current and a leak current or shunt. This distinction is artificial and therefore V ∗ is an arbitrary voltage, which we are free to choose. Our aim is to choose it such, that neglecting the shunting term changes the firing rate minimally. For example, when the (type 1) HH neuron is firing action potentials, the region around the threshold voltage V th is traversed relatively slowly, due to the ghost of the saddle-node bifurcation (Izhikevich 2007). The time taken to traverse this region largely determines the firing rate. Therefore, for continuously firing HH neurons, an accurate approximation for the firing rate is obtained when V ∗ is set to V th, such that the shunting term is small in that region.
Choosing an optimal V ∗ minimizes the error in the approximation, but does not guarantee that it is accurate. In order for the approximation to be accurate, the synaptic conductance should be small compared to the total (leak and gated) conductance. For our single cell models, the approximation was empirically validated.
Note that the optimal value for V ∗ depends on the specific single cell model used. However, we found that also the firing rate of our type 2 neuron was approximated reasonably when V ∗ was set to V th, with an accuracy depending on the relative magnitude of the inhibitory conductance.
Appendix C: Detailed results
We show the results of the simulated networks, as described in the main body of the paper, in more detail. Results are shown for networks of type 1 neurons with weak feedback (Fig. 12), with strong feedback (Fig. 13) and type 2 neurons (Fig. 14). The NMM model describes the dynamics of the network of neurons very well, both for the synaptic conductances (mean and variance) and the firing rates (mean and variance). The fluctuations in the standard deviations are caused by the small size of the modeled network. These are stochastic in nature and hence are not reproduced by the NMM.

Comparison of our new NMM with a detailed network model. The reaction to a double step in input rate of the network model (full lines and dots) is compared to that of the NMM (blue dashed lines). The mean and standard deviation of three synaptic currents are displayed, as well as those of the firing rates of the excitatory and inhibitory populations

Simulations of a network with strong feedback. The reaction to a double step in input rate of the new NMM (blue dashed lines) is compared to that of the network model (full lines and dots). The mean and standard deviation of three synaptic currents are displayed, as well as the firing rates of the excitatory and inhibitory populations. The standard deviations of the firing rates of the network model are not shown since they are dominated by artifacts

The dynamics of two populations of type 2 spiking neurons. The reaction to a double step in input rate of the NMM (blue dashed lines) is compared to that of the network model (full lines). The mean and standard deviation of three synaptic currents are displayed, as well as those of the firing rates of the excitatory and inhibitory populations
Rights and permissions
About this article

Cite this article
Zandt, BJ., Visser, S., van Putten, M.J.A.M. et al. A neural mass model based on single cell dynamics to model pathophysiology. J Comput Neurosci 37, 549–568 (2014). https://doi.org/10.1007/s10827-014-0517-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10827-014-0517-5