
Abstract
Model organisms are central to contemporary biology and studies of embryogenesis in particular. Biologists utilize only a small number of species to experimentally elucidate the phenomena and mechanisms of development. Critics have questioned whether these experimental models are good representatives of their targets because of the inherent biases involved in their selection (e.g., rapid development and short generation time). A standard response is that the manipulative molecular techniques available for experimental analysis mitigate, if not counterbalance, this concern. But the most powerful investigative techniques and molecular methods are applicable to single-celled organisms (‘microbes’). Why not use unicellular rather than multicellular model organisms, which are the standard for developmental biology? To claim that microbes are not good representatives takes us back to the original criticism leveled against model organisms. Using empirical case studies of microbes modeling ontogeny, we break out of this circle of reasoning by showing: (a) that the criterion of representation is more complex than earlier discussions have emphasized; and, (b) that different aspects of manipulability are comparable in importance to representation when deciding if a model organism is a good model. These aspects of manipulability harbor the prospect of enhancing representation. The result is a better understanding of how developmental biologists conceptualize research using experimental models and suggestions for underappreciated avenues of inquiry using microbes. More generally, it demonstrates how the practical aspects of experimental biology must be scrutinized in order to understand the associated scientific reasoning.





Similar content being viewed by others


Notes
“The motivation for their study is not simply to understand how that particular animal develops, but to use it as an example of how all animals develop” (Slack 2006, 61). “Basic research aimed at understanding the general principles of life uses model organisms such as the frog, chicken, fish, and mouse” (Nüsslein-Volhard 2006, 138).
Much of the funding for developmental biology in the United States comes from the National Institutes of Health. A similar pattern holds in other national contexts. For example, the Welcome Trust makes the connection explicit: “Yeast, fruit flies, nematode worms and mice: just some of the model organisms whose study is bringing insights into the role of genes in the human body” (http://genome.wellcome.ac.uk/node30073.html, our emphasis). It is difficult to escape questions of human biomedical applicability when assessing model organism use, but we will not explore it here (see Ankeny and Leonelli 2011; Bolker 2009 for further discussion).
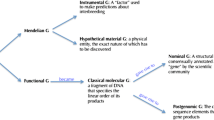
There is an ambiguity in talk of model organisms “representing” other organisms. One possibility is that this representing is done indirectly through abstract models (“representations”) constructed from data derived from the study of developmental model organisms. Another possibility is that the material entity itself is a “direct” representation of other organisms. Although we think both possibilities have merit in understanding the reasoning practices of developmental biology, our argument does not depend on its resolution one way or another. Therefore, we will continue using the phrasing of model organisms representing other organisms.
“The past two decades have brought major breakthroughs in our understanding of the molecular and genetic circuits that control a myriad of developmental events in vertebrates and invertebrates. These detailed studies have revealed surprisingly deep similarities in the mechanisms underlying developmental processes across a wide range of bilaterally symmetric metazoans … [these] comparisons have defined a common core of genetic pathways guiding development” (Bier and McGinnis 2003, 25, our emphasis).
Hickson et al. (2009) argue that metastatic colonization in cancer cells is controlled by a quorum-sensing mechanism directly akin to bacterial cell–cell communication.
“For a large number of problems there will be some animal of choice or a few such animals on which it can be most conveniently studied” (Krogh 1929, 202).
Note that this distinction is not identical to Bolker’s two modes of representation: exemplary and surrogate (Bolker 2009). Both general and Krogh-principle model organisms can instantiate exemplary modes (serving basic research by exemplifying a larger group) and surrogate modes (providing indirect experimental access phenomena that are difficult to work with).
Representation often takes a back seat in descriptions of the advantages of model organisms, such as in the Welcome Trust’s account of zebrafish, where the preferable model features are almost all manipulation related: “The zebrafish comes close to being the ideal model organism for vertebrate development because it appears to combine the best features of all the other models. … zebrafish embryos develop externally and can be viewed and manipulated at all stages. … zebrafish development is more rapid than in the frog, the organisation of the embryo is simpler and … the embryo is transparent. Like the mouse, the zebrafish is amenable to genetic analysis and has a similar generation interval (2–3 months). However, zebrafish are smaller than mice and they produce more offspring in a shorter time. … It is easy to induce new mutations in zebrafish and large-scale screens have been carried out to identify mutations causing defects in particular biological processes, such as the developing nervous system” (http://genome.wellcome.ac.uk/doc_WTD020806.html).
“Neither the frog nor the chicken is favoured for its genetic amenability. …An additional disadvantage of Xenopus laevis for genetic analysis is that the species is tetraploid. …The predominant reason for the use of Xenopus and chickens as models is that they produce large, robust embryos whose development occurs outside the body of the mother. …the embryos are much more accessible than those of mammals. …The accessibility of the embryos means they can be surgically manipulated or treated with proteins and chemicals that interfere with normal development” (http://genome.wellcome.ac.uk/doc_WTD020805.html).
“Because mice do not require Dmrt1 for primary sex determination, they may not be typical vertebrates, but the genetic and molecular tools that are available in the mouse have allowed a detailed dissection of Dmrt1 function” (Matson and Zarkower 2012, 169, our emphasis).
“One of the most attractive features of C. elegans is that it can be handled like a microbe. Large numbers of worms can be maintained inexpensively on lawns of bacteria growing on standard agar plates, but viable cultures can be stored as frozen stocks and then revived when required” (http://genome.wellcome.ac.uk/doc_WTD020809.html, our emphasis).
Others have noted this possibility but tended to be pessimistic because of our lack of knowledge: “Many problems in eukaryotic cell biology can be most easily studied in unicellular organisms, such as yeast…. Other problems, however, currently can be studied meaningfully only in intact animals. This may be because we do not know how to mimic crucial aspects of the organismal environment in vitro, because cell–cell interactions play an important role, or because the process under study involves a behavior that is not currently understood in terms of the properties of individual cells. Examples include pattern formation in the embryo and the development and function of organ systems” (Rubin 1988, 1453, our emphasis).
These kinds of questions are pertinent apart from microbial models. Developmental biology already has synthetic models that break continuity with natural organisms, such as tissue explants from Xenopus (Sive et al. 2007). “Keller” explants are portions of dorsal mesendoderm and ectoderm that must be dissected out of intact embryos, assembled into a non-natural rectangular shape, flattened (‘sandwiched’) between a slide and cover slip (or equivalent), and cultured in vitro. They facilitate the observation of convergent extension, a type of morphogenesis, relevant to the developmental phenomenon of gastrulation. This suggests further treatment of what it means for something to be a “synthetic” model of development and the different ways one achieves this result (e.g., de novo construction versus extraction from a living entity).
We owe special thanks to an anonymous referee for stressing this point explicitly.
This also has been demonstrated experimentally in Dictyostelium: “the single-cell bottleneck is a powerful stabilizer of cellular cooperation in multicellular organisms” (Kuzdzal-Fick et al. 2011, 1548).
There are precedents in the history of philosophy of science that elevate manipulative capacities as a mark of epistemic worthiness, such as Bacon’s notion of maker’s knowledge (Pérez-Ramos 1989), but the intended scope of our remarks is with respect to the critical literature surrounding model organisms over the past three decades.
The same strategy applies to models of evolution: a regular movement between microbial and metazoan models of evolution is a potent methodology for the dissection of the mechanics of evolutionary processes because it maximizes experimental manipulation in conjunction with representational calibration derived from multiple model organism types.
References
Ackermann M, Stearns SC, Jenal U (2003) Senescence in a bacterium with asymmetric division. Science 300:1920
Alao JP (2007) The regulation of cyclin D1 degradation: roles in cancer development and the potential for therapeutic invention. Mol Cancer 6:24
Ankeny RA (2009) Model organisms as fictions. In: Suárez M (ed) Fictions in science: philosophical essays on modeling and idealization. Routledge, Tayor & Francis Group, New York, pp 193–204
Ankeny RA, Leonelli S (2011) What’s so special about model organisms? Stud Hist Philos Sci 42:313–323
Bier E, McGinnis W (2003) Model organisms in the study of development and disease. In: Epstein CJ, Erickson RP, Wynshaw-Boris A (eds) Molecular basis of inborn errors of development. Oxford University Press, New York, pp 25–45
Bolker JA (1995) Model systems in developmental biology. BioEssays 17:451–455
Bolker JA (2009) Exemplary and surrogate models: two modes of representation in biology. Perspect Biol Med 52:485–499
Botstein D, Fink GR (1988) Yeast: an experimental organism for modern biology. Science 240:1439–1443
Botstein D, Fink GR (2011) Yeast: an experimental organism for 21st century biology. Genetics 189:695–704
Brigandt I, Love AC (2012) Reductionism in biology. In: Zalta EN (ed) The stanford encyclopedia of philosophy. http://plato.stanford.edu/entries/reduction-biology/
Brun YV, Shimkets LJ (eds) (2000) Prokaryotic development. American Society for Microbiology, Washington, DC
Chen ZH, Schaap P (2012) The prokaryote messenger c-di-GMP triggers stalk cell differentiation in Dictyostelium. Nature 488:680–683
Collins J (2012) Synthetic biology: bits and pieces come to life. Nature 483:S8–S10
Crotty DA, Gann A (2009) Emerging model organisms: a laboratory manual, vol 1. Cold Spring Harbor Laboratory Press, Cold Spring Harbor
Davies JA (2005) Mechanisms of morphogenesis: the creation of biological form. Elsevier Academic Press, San Diego
Davis RH (2004) The age of model organisms. Nat Rev Genet 5:69–76
Dayel MJ, Alegado RA, Fairclough SR, Levin TC, Nichols SA, McDonald K, King N (2011) Cell differentiation and morphogenesis in the colony-forming choanoflagellate Salpingoeca rosetta. Dev Biol 357:73–82
Ereshefsky M, Pedroso M (2013) Biological individuality: the case of biofilms. Biol Philos. doi:10.1007/s10539-012-9340-4
Erwin DH, Valentine JW (2013) The Cambrian explosion: the construction of animal biodiversity. Roberts, Greenwood
Escalante AE, Inouye S, Travisano M (2012) A spectrum of pleiotropic consequences in development due to changes in a regulatory pathway. PLoS ONE 7:e43414
Fairclough SR, Dayel MJ, King N (2010) Multicellular development in a choanoflagellate. Curr Biol 20:R875–R876
Fiers W, Contreras R, Duerinck F, Haegeman G, Iserentant D, Merregaert J, Min Jou W, Molemans F, Raeymaekers A, Van den Berghe A, Volckaert G, Ysebaert M (1976) Complete nucleotide sequence of bacteriophage MS2 RNA: primary and secondary structure of the replicase gene. Nature 260:500–507
Foster KR (2011) The sociobiology of molecular systems. Nat Rev Genet 12:193–203
Frank SA (2007) Dynamics of cancer: incidence, inheritance, and evolution. Princeton University Press, Princeton
Frankino WA, Raff RA (2004) Evolutionary importance and pattern of phenotypic plasticity. In: DeWitt TJ, Scheiner SM (eds) Phenotypic plasticity: functional and conceptual approaches. Oxford University Press, New York, pp 64–81
Frigg R, Hartmann S (2012) Models in science. In: Zalta EN (ed) The stanford encyclopedia of philosophy. http://plato.stanford.edu/archives/fall2012/entries/models-science/
Gajewski K, Schulz RA (2002) Comparative genetics of heart development: conserved cardiogenic factors in Drosophila and vertebrates. In: Ostadal B, Nagano M, Dhalla NS (eds) Cardiac development. Kluwer, Boston, pp 1–23
Gerhart J, Kirschner M (2007) The theory of facilitated variation. Proc Natl Acad Sci USA 104:8582–8589
Gilbert SF (2010) Developmental biology, 9th edn. Sinauer, Sunderland
Grosberg RK, Strathmann RR (2007) The evolution of multicellularity: a minor major transition? Ann Rev Ecol Evolut Syst 38:621–654
Gu Z, David L, Petrov D, Jones T, Davis RW, Steinmetz LM (2005) Elevated evolutionary rates in the laboratory strain of Saccharomyces cerevisiae. Proc Natl Acad Sci USA 102:1092–1097
Herron MD, Hacket JD, Aylward FO, Michod RE (2009) Triassic origin and early radiation of multicellular volvocine algae. Proc Natl Acad Sci USA 106:3254–3258
Hickson J, Yamada SD, Berger J, Alverdy J, O’Keefe J, Bassler B, Rinker-Schaeffer C (2009) Societal interactions in ovarian cancer metastasis: a quorum-sensing hypothesis. Clin Exp Metastasis 26:67–76
Holley SA, Geisler R, Nüsslein-Volhard C (2000) Control of her1 expression during zebrafish somitogenesis by a Delta-dependent oscillator and an independent wave-front activity. Genes Dev 14:1678–1690
Huntley S, Hamann N, Wegener-Feldbrügge S, Treuner-Lange A, Kube M, Reinhardt R, Klages S et al (2011) Comparative genomic analysis of fruiting body formation in Myxococcales. Mol Biol Evol 28:1083–1097
Inouye M, Inouye S, Zusman DR (1979) Biosynthesis and self-assembly of protein S, a development-specific protein of Myxococcus xanthus. Proc Natl Acad Sci USA 76:209–213
Inouye S, Soberon X, Franceschini T, Nakamura K, Itakura K, Inouye M (1982) Role of positive charge on the amino terminal region of the signal peptide in protein secretion across the membrane. Proc Natl Acad Sci USA 79:3438–3441
Inouye S, Nariya H, Munoz-Dorado J (2008) Protein Ser/Thr kinases and phosphotases in Myxococcus xanthus. In: Whitworth DE (ed) Myxobacteria: multicellularity and differentiation. American Society for Microbiology, Washington, DC, pp 191–210
Insall R, Nayler O, Kay RR (1992) DIF-1 induces its own breakdown in Dictyostelium. EMBO J 11:2849–2854
Jenner RA, Wills MA (2007) The choice of model organisms in evo-devo. Nat Rev Genet 8:311–319
Kampichler C, Bruckner A, Kandeler E (2001) Use of enclosed model ecosystems in soil ecology: a bias towards laboratory research. Soil Biol Biochem 33:269–275
Kilfoil ML, Lasko P, Abouheif E (2009) Stochastic variation: from single cells to superorganisms. HFSP J 3:379–385
Kirby ML (1999) Contribution of neural crest to heart and vessel morphology. In: Harvey RP, Rosenthal N (eds) Heart development. Academic Publishers, San Diego, pp 179–193
Kraemer SA, Toups MA, Velicer GJ (2010) Natural variation in developmental life-history traits of the bacterium Myxococcus xanthus. FEMS Microbiol Ecol 73:226–233
Krogh A (1929) The progress of physiology. Science 70:200–204
Kuzdzal-Fick J, Fox SA, Strassmann JE, Queller DC (2011) High relatedness is necessary and sufficient to maintain multicellularity in Dictyostelium. Science 334:1548–1551
Lennox ES (1955) Transduction of linked genetic characters of the host by bacteriophage P1. Virology 1:190–206
Lenski RE, Travisano M (1994) Dynamics of adaptation and diversification: a 10,000-generation experiment with bacterial populations. Proc Natl Acad Sci USA 91:6808–6814
Lichius A, Lord KM, Jeffree CE, Oborny R, Boonyarungsrit P, Read ND (2012) Importance of MAP kinases during protoperithecial morphogenesis in Neurospora crassa. PLoS ONE 7:e42565
Liu C, Fu X, Liu L, Ren X, Chau CKL, Li S, Xiang Lu et al (2011) Sequential establishment of stripe patterns in an expanding cell population. Science 334:238–241
Lord KM, Read ND (2011) Perithecium morphogenesis in Sordaria macrospora. Fungal Genet Biol 48:388–399
Losick R, Shapiro L (1984) Microbial development. Cold Spring Harbor Laboratory Press, Cold Spring Harbor
Love AC (2008) Explaining the ontogeny of form: philosophical issues. In: Plutynski A, Sarkar S (eds) The blackwell companion to philosophy of biology. Blackwell Publishers, Malden, pp 223–247
Love AC (2010) Idealization in evolutionary developmental investigation: a tension between phenotypic plasticity and normal stages. Philos Trans R Soc B Biol Sci 365:679–690
Love AC (2013) Theory is as theory does: scientific practice and theory structure in biology. Biol Theory. doi:10.1007/s13752-012-0046-2
Lynch VJ (2009) Use with caution: developmental systems divergence and potential pitfalls of animal models. Yale J Biol Med 82:53–66
Malumbres M, Barbacid M (2001) Milestones in cell division: to cycle or not to cycle: a critical decision in cancer. Nat Rev Cancer 1:222–231
Malumbres M, Barbacid M (2009) Cell cycle, CDKs and cancer: a changing paradigm. Nat Rev Cancer 9:153–166
Matson CK, Zarkower D (2012) Sex and the singular DM domain: insights into sexual regulation, evolution and plasticity. Nat Rev Genet 13:163–174
Maxam AM, Gilbert W (1977) A new method for sequencing DNA. Proc Natl Acad Sci USA 74:560–564
McFall-Ngai MJ (2002) Unseen forces: the influences of bacteria on animal development. Dev Biol 242:1–14
McGary KL, Park TJ, Woods JO, Cha HJ, Wallingford JB, Marcotte EM (2010) Systematic discovery of nonobvious human disease models through orthologous phenotypes. Proc Natl Acad Sci USA 107:6544–6549
Merlo P, Maley R (2006) Cancer as an evolutionary and ecological process. Nat Rev Cancer 6:924–935
Metscher BD, Ahlberg PE (1999) Zebrafish in context: uses of a laboratory model in comparative studies. Dev Biol 210:1–14
Minelli A (2011) Animal development, an open-ended segment of life. Biol Theory 6:4–15
Mizuno T, Chou MY, Inouye M (1984) A unique mechanism regulating gene expression: translational inhibition by a complementary RNA transcript (micRNA). Proc Natl Acad Sci USA 81:1966–1970
Moir A (2006) How do spores germinate? J Appl Microbiol 101:526–530
Nigg S (2001) Mitotic kinases as regulators of cell division and its checkpoints. Nat Rev Mol Cell Biol 2:21–32
Nüsslein-Volhard C (2006) Coming to life: how genes drive development. Kales Press, Carlsbad
Palmeirim I, Henrique DA, Ish-Horowicz D, Pourquié O (1997) Avian hairy gene expression identifies a molecular clock linked to vertebrate segmentation and somitogenesis. Cell 91:639–648
Pérez-Ramos A (1989) Francis Bacon’s idea of science and the maker’s knowledge tradition. Oxford University Press, New York
Queller DC, Strassmann JE (2009) Beyond society: the evolution of organismality. Philos Trans R Soc Lond B 364:3143–3155
Rao CV, Wolf DM, Arkin AP (2002) Control, exploitation and tolerance of intracellular noise. Nature 420:231–237
Ratcliff WC, Pentz J, Travisano M (2013) Tempo and mode of multicellular adaptation in experimentally-evolved Saccharomyces cerevisiae. Evolution
Ratcliff WC, Denison RF, Borrello M, Travisano M (2012) Experimental evolution of multicellularity. Proc Natl Acad Sci USA 109:1595–1600
Rebolleda-Gomez M, Ratcliff WC, Travisano M (2012) Adaptation and divergence during experimental evolution of multicellular Saccharomyces cerevisiae. Artif Life 13:99–104
Rubin GM (1988) Drosophila melanogaster as an experimental organism. Science 240:1453–1459
Saiki RK, Gelfand DH, Stoffel S, Scharf SJ, Higuchi R, Horn GT, Mullis KB, Erlich HA (1988) Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 239:487–491
Saito TT, Kato A, Kay RR (2008) DIF-1 induces the basal disc of the Dictyostelium fruiting body. Dev Biol 317:444–453
Sanger F, Nicklen S, Coulson AR (1977) DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA 74:5463–5467
Schaap P (2011) Evolutionary crossroads in developmental biology: Dictyostelium discoideum. Development 138:387–396
Schröter C, Ares S, Morelli L, Isakova A, Hens K, Soroldoni D, Gajewski M et al (2012) Topology and dynamics of the zebrafish segmentation clock core circuit. PLoS Biol 10:e1001364
Schulze J, Schierenberg E (2011) Evolution of embryonic development in nematodes. EvoDevo 2:18
Schultz D, Wolynes PG, Jacob EB, Onuchic JN (2009) Deciding fate in adverse times: sporulation and competence in Bacillus subtilis. Proc Natl Acad Sci USA 106:21027–21034
Shapiro JA (1998) Thinking about bacterial populations as multicellular organisms. Ann Rev Microbiol 52:81–104
Sive HL, Grainger RM, Harland RM (2007) Xenopus laevis Keller explants. Cold Spring Harb Protoc 6:pdb.prot4749
Slack JMW (2006) Essential developmental biology, 2nd edn. Blackwell Publishing, Malden
Slack JMW (2009) Emerging market organisms. Science 323:1674–1675
Sommer RJ (2009) The future of evo-devo: model systems and evolutionary theory. Nat Rev Genet 10:416–422
Sprouffske K, Merlo LMF, Gerrish PJ, Maley CC, Sniegowski PD (2012) Cancer in light of experimental evolution. Curr Biol 22:R762–R771
Steel DP (2008) Across the boundaries: extrapolation in biology and social science. Oxford University Press, New York
Stewart EJ, Madden R, Paul G, Taddei F (2005) Aging and death in a organism that reproduces by morphologically symmetric division. PLoS Biol 3:e45
Taylor AL, Trotter CD (1972) Linkage map of Escherichia coli strain K-12. Bacteriol Rev 36:504–524
Travisano M, Velicer G (2004) Strategies of microbial cheater control. Trends Microbiol 12:72–78
Ueki T, Inouye S, Inouye M (1996) Positive-negative KG cassettes for construction of multi-gene deletions using a single drug marker. Gene 183:153–157
Weber M (2005) Philosophy of experimental biology. Cambridge University Press, New York
Whitworth DE (ed) (2008) Myxobacteria: multicellularity and differentiation. American Society for Microbiology, Washington, DC
Williams JG (2006) Transcriptional regulation of Dictyostelium pattern formation. EMBO Rep 7:694–698
Winzeler EA, Shoemaker DD, Astromoff A, Liang H, Anderson K, Andre B, Bangham R et al (1999) Functional characterization of the S. cerevisiae genome by gene deletion and parallel analysis. Science 285:901–906
Zhao Y, Potter SS (2002) Functional comparison of the Hoxa4, Hoxa10, and Hoxa11 homeoboxes. Dev Biol 244:21–36
Acknowledgments
We benefited from the feedback and critical comments provided by Ana Escalante, Brian Hall, Masayori Inouye, Sabina Leonelli, Katherine Liu, Maureen O’Malley, Maria Rebolleda-Gomez, Pauline Schaap, Beckett Sterner, Greg Velicer, and anonymous reviewers for the journal. Their assistance should not be interpreted as an endorsement of our argument. Special thanks to participants of the symposium “Molecules, Organisms, Systems: Developing Multilevel Integrated Insights into Biological Processes” at the 23rd biennial meeting of the Philosophy of Science Association (San Diego, November 2012), where a subset of this material was presented and discussed, and to participants of a colloquium presentation of this manuscript at the Department of History and Philosophy of Science, Indiana University (January 2013). ACL is grateful to Matthew Spates for helpful discussion and research assistance. MT dedicates this article to Sumiko Inouye, a fellow developmental microbiologist and close friend, who will be greatly missed.
Author information
Authors and Affiliations
Corresponding author
Additional information
Order of authorship is alphabetical.
Rights and permissions
About this article
Cite this article
Love, A.C., Travisano, M. Microbes modeling ontogeny. Biol Philos 28, 161–188 (2013). https://doi.org/10.1007/s10539-013-9363-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10539-013-9363-5