
Abstract
The ruff Philomachus pugnax, a lekking shorebird wintering in Africa and breeding across northern Eurasia, declined severely in its western range. Based on a capture-mark-resighting programme (2004–2011) in the westernmost staging area in Friesland (the Netherlands), we investigated changes in apparent annual survival in relation to age and sex to explore potential causes of decline. We also related temporal variation in apparent survival to environmental factors. We used the Capture-Mark-Recapture multievent statistical framework to overcome biases in survival estimates after testing for hidden heterogeneity of detection. This enabled the estimation of the probability to belong to high or low detectability classes. Apparent survival varied between years but was not related to weather patterns along the flyway, or to flood levels in the Sahel. Over time, a decline in apparent survival is suggested. Due to a short data series and flag loss in the last period this cannot be verified. Nevertheless, the patterns in sex-specific detectability and survival lead to new biological insights. Among highly detectable birds, supposedly most reliant on Friesland, males survived better than females (\(\bar{\phi }_{\text{HD males}}\) = 0.74, range 0.51–0.93; \(\bar{\phi }_{\text{HD females}}\) = 0.51, range 0.24–0.81). Among low detectable birds, the pattern is reversed (\(\bar{\phi }_{\text{LD males}}\) = 0.64, range 0.37–0.89; \(\bar{\phi }_{\text{LD females}}\) = 0.73, range 0.48–0.93). Probably the staging population contains a mixture of sex-specific migration strategies. A loss of staging females could greatly affect the dynamics of the western ruff population. Further unravelling of these population processes requires geographically extended demographic monitoring and the use of tracking devices.
Similar content being viewed by others


Introduction
Increasing human pressures on habitats are driving many migratory species in decline (Berger 2004; Sanderson et al. 2006; Wilcove and Wikelski 2008; Limburg and Waldman 2009). Despite their great ability to move, due to specific habitat requirements, tight time schedules and density dependent processes, long-distance migrants are vulnerable to global change (Alerstam et al. 2003; Piersma 2007; Taylor and Norris 2007). An understanding of the resilience of migratory species, and the design of effective conservation measures for their protection, requires the identification of the reasons for population change. An essential step is to address the variation in demographic processes (survival, recruitment and movements) determining population growth rate (Caswell 2001). The challenge to do this properly is considerable when studying long-distance migrants because: (1) the variation in demographic processes that may be related to immediate or delayed effect(s) of event(s) encountered in separated areas and times during the species migratory cycle (Harrison et al. 2011; Juillet et al. 2012) and (2) the degree of migratory connectivity will influence the population dynamics (Webster et al. 2002; Taylor and Norris 2010). Capture-Mark-Recapture (CMR) methods based on the long-term monitoring of marked individuals constitute a versatile approach to assess population change. Taking into account imperfect detection of marked individuals, it allows the estimation of demographic rates in a robust way and provides a basis for inference on potential drivers (Lebreton et al. 1992; Gimenez et al. 2008).
The ruff (Philomachus pugnax) is a long-distance migrating shorebird found across the African and Eurasian continents (van Rhijn 1991). Since the late 20th century, ruffs showed a strong population decline over their westernmost migration route, the route that connects the inland wetlands of West-Africa to the wet grasslands of northwestern Europe and the tundras north and east of it (Zöckler 2002a; Rakhimberdiev et al. 2010; Verkuil et al. 2012). Breeding ruffs first vanished from temperate latitudes partly because of the loss of wet grasslands habitat following European agriculture intensification (Thorup 2006), but their decline also spread into the sub-arctic region of Scandinavia (Väisänen et al. 2005; Øien and Aarvak 2010; Lindström and Green 2013). The latter suggests that global factors, including climate warming (Zöckler 2002b; Ims et al. 2008; Virkkala and Rajasärkkä 2011) may play a role. At the main staging area of ‘western’ ruffs, in the Netherlands, peak roost counts of the spring passage population have declined from >20,000 in the late 1990s to fewer than 5000 in 2010 (Verkuil et al. 2012; Fig. 1b). In concert with a decline in numbers, the daily body mass increments of staging ruffs decreased between 2001 and 2008, suggesting a decrease in staging habitat quality as a driver of this loss (Verkuil et al. 2012). Changes occurring on the African nonbreeding grounds, including the repeated droughts, high hunting pressure and increasing human pressure in the Sahel region could be involved as well (Zwarts et al. 2009). Recent studies suggested the occurrence of a massive redistribution towards a more eastern flyway which would have contributed to the decline in numbers of western ruffs (Rakhimberdiev et al. 2010; Verkuil et al. 2012).

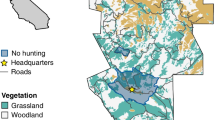
a Study area in southwest Friesland, the Netherlands. Black circles indicate main night-time roosts along the IJsselmeer lakeshore used by staging ruffs. Green (light grey area with grey outline) and red (dark grey) areas represent areas of intensively and extensively managed grasslands, respectively. Most used polders (between 2004 and 2011) are outlined by a black line. b Maximum number of ruffs counted over roosts along the IJsselmeer lakeshore specified in (a) from July to June but here relevant to spring migration season hosting highest number of migrating ruffs—www.sovon.nl. The grey area represents 95 % confidence interval. c Number of male and female ruff vs. total number of individuals captured per spring between 2004 and 2011 in the study area
The demographic processes underlying ruff population changes remain to be investigated, and given the strong sexual dimorphism (males are much larger, and more ornamented, than females) this requires a sex-specific approach. The ruff is a lekking species (Hogan-Warburg 1966; Widemo 1998) in which males and females differ not only by size but also by their ecology and migratory behaviour (van Rhijn 1991). The demographic processes and density-dependent processes relative to the spatio-temporal distributions of the two sexes are important determinants of their population dynamics (van Rhijn 1991; Höglund 1996; Anthony and Blumstein 2000; Gascoigne et al. 2009).
As a first step in the investigation of the demography of the western migrant ruff population, we focused on survival probability. For shorebird species, which are relatively long-lived with an early maturity but a highly variable recruitment due to the unpredictability of their breeding environments especially at high latitude (see e.g., Aharon-Rotman et al. 2015), adult survival is likely an important determinant of population growth rate (Sæther and Bakke 2000). Based on a capture-mark-resight dataset collected during eight successive spring migration seasons (March–April 2004–2011) in the staging area of the Netherlands in the province of Friesland, we used the CMR statistical framework to investigate annual variation in ruff “apparent” survival probability (i.e., product of survival and fidelity) as a function of sex and age. Further, we explored relationships between annual variation in apparent survival and environmental conditions encountered during the entire annual cycle along the East-Atlantic migratory flyway.
Methods
Field methods and data collection
Our study area in southwest Friesland, the Netherlands, is situated along the northeast shores of Lake IJsselmeer and covers over 100 km2 of agricultural habitat, mainly intensively managed grasslands for dairy farming along with traditionally managed wet herb-rich grasslands (Verkuil and de Goeij 2003; Groen et al. 2012, Fig. 1a). The passage of northward migrating ruffs starts in mid-March and lasts into May. From the 2nd week of March numbers build up to peak in the first half of April. Males stay for about three weeks (Verkuil et al. 2010), during which they undertake nuptial moult and gain body mass for the flight towards their breeding grounds (Jukema and Piersma 2000; Verkuil et al. 2010). Migration of subadults (first year of age) starts in early April and females (called ‘reeves’) only appear in mid-April (Wymenga 1999). Females are also much less numerous than males and due to insufficient resightings data, their staging duration could not be determined (see Verkuil et al. 2010). By the second week of May all ruffs have left the staging area.
Ruffs were captured with traditional, partially wind-driven, “wilsternets” (equivalent to a 20 m long and 3 m high clap net; Piersma et al. 2005). Age was determined from leg colour [grey/green in subadults (first year bird); orange/pink in adults] and the presence of brown (buff) middle cover tertiary feathers retained from juvenile plumage (Glutz von Blotzheim et al. 1975). As females are a third smaller than males, most birds could easily be assigned a sex, while the fæders, female-mimicking males, were discriminated using wing length (Jukema and Piersma 2006). At last, each captured ruff was marked with a metal ring and a unique combination of 4 colour-rings and a flag before release.
Observations of colour-ringed individuals were made with telescopes (20–60 × 65 mm and 20–60 × 80 mm telescopes) from cars parked along country roads between agricultural fields or in nature reserves. Each spring, two teams of two persons were looking for colour-ringed individuals for up to 10 h a day, 6 days a week. The southern, central and northern parts of our study site were visited alternatively in order to achieve complete coverage every 2 days.
Our observation effort spread during the entire passage period (i.e., 15 March–15 May). In this way we guaranteed a good and consistent coverage of the passage population throughout the years of study. Staging ruffs were not evenly spread across the study area, and some subareas were used more than others, in particular coastal polders close to roost sites (see Fig. 1a). Theses sub-areas necessarily required more time. However, as ruffs are very mobile, turnover of individuals is usually high. Also, all observations for which the colour-ring combination was not identified with assurance were excluded from the analyses.
Between 2004 and 2010 we marked 5026 ruffs. Among them were 4210 males of which 659 (15 %) were marked as subadult and 3551 as adult; and 816 females of which 157 (18 %) were subadult and 659 adult. From 2004 to 2011, 23 % of the marked females (n = 187) and 42 % of the marked males (n = 1769) were seen at least once after release. Tag loss (loss of the flag) was very limited until 2009. In 2010, the loss of flag became more common and we started recording it carefully. The 14 cases individuals without a flag were identified by reading their metal ring with a telescope indicated that flags were not lost immediately after ringing, but suggest a gradual loss. Degradation or heterogeneous quality of the plastics may be involved. Loss of colour-rings hardly ever occurred. Observations of birds which had lost their coloured flag comprised about 5 % (100 observations of marked ruffs with no flag on a total of 2138 observations) and 10 % (206 observations of marked ruffs with no flag on a total of 2146 observations) of our sightings in the spring seasons of 2010 and 2011, respectively. However as a same bird could be observed more than one time, the proportion of birds that have lost their flag must be lower. In the absence of physical recaptures to individually verify the loss of flags using information on the metal rings, tag-loss cannot be easily handled (see Juillet et al. 2010). We must therefore acknowledge that our estimates of apparent survival are likely negatively biased especially at the end of the study (see Arnason and Mills 1981).
Statistical modelling
We defined four groups: males and females, either marked as adults or marked as subadults. The encounter history of each individual was coded ‘1’ when the individual was either captured or subsequently observed during a given encounter occasion (i.e., here a spring migration period); ‘0’ when the individual was not observed.
Goodness of fit tests (GOF—see Pradel et al. 2005) performed in software U-CARE (Choquet et al. 2009a) indicated a slight lack of fit between the general time- and group-dependent Cormack-Jolly-Seber models (Cormack 1964; Jolly 1965; Seber 1965) and our data (overall test, χ 2 = 121.2, P = 0.042, \(\hat{c}\) = 1.263). Significant trap-dependence, in this case ‘trap-happiness’ (i.e., individuals recently captured or observed were more likely to be re-observed subsequently—overall Test 2.CT, P < 0.001), was detected particularly among males (Test 2.CT—males marked as subadult, P = 0.018; marked as adult, P = 0.001). The test for transience was significant for adult males (i.e., presence of transients: individuals marked and never re-observed –Test 3.SR, P = 0.016). In females, all test statistics were non-significant, but the sparse amount of data could have limited the power to detect such biases (see Pradel et al. 1997). Results of GOF tests indicated overall non-homogeneous detection (Crespin et al. 2008). Such hidden heterogeneity in detection among marked ruffs may be related to our sampling or linked to the behaviour of the birds.
To account for uncertainty on heterogeneity of detection and to overcome biases in survival estimates, we adopted multievent models (Pradel 2005; Crespin et al. 2008; Pradel and Sanz-Aguilar 2012). We considered two live states, one being individuals alive and at high probability of detection (highly detectable, HD) and the other, individuals alive at low probability of detection (low detectable, LD). A third state is reserved for dead individuals that cannot be observed. Upon initial capture, each individual has a probability (π) to belong to a given detectability class. This probability relates to the overall encounter history of the individuals, depending on whether individuals have been frequently or sporadically resighted across the study period. Each spring migration period, individuals can be either observed or not (i.e., the events) with a detection probability (p) conditional on the underlying states (alive and either of high or of low detectability). Thus, birds assigned to the high detectability class with a probability π HD, shared a high probability of detection p HD and, inversely, birds assigned to the low detectability class with a probability (1 − π HD), shared a low probability of detection p LD. Individuals were assumed to survive between years t and t + 1 with a survival probability (ϕ) which could be state-specific (survival probabilities in state HD or LD). Transitions between states were not considered (i.e., no transitions between detectability classes).
Differences in detectability may be linked to unrecognized behavioural features of the birds, features which themselves might be related to their survivorship or fidelity to Friesland. For this reason we integrated detectability classes into the modelling of the apparent survival probability. We also defined two age classes: adults (age ≥ 2 years—noted ad) and subadults (first year—noted 1y). Birds marked as subadults may have different propensities to re-use Friesland and/or to survive the year following marking than adults. Subadults may only be “prospectors” on breeding grounds (e.g., Kentie et al. 2014). According to van Rhijn (1991) only a small proportion of males and females will reproduce when one year old. First year male ruffs refuel less rapidly (Karlionova et al. 2008) and develop less elaborated nuptial plumage than adults, behave mostly as marginal males and are less involved in displays at leks and have lower mating chances (van Rhijn 1991). We thus considered age as an additive effect. In view of the distinctiveness of males and females, we consider an effect of sex in combination with age and detectability classes.
At last, our most complex model assumed a non-null transience probability for adult male τ(ad males). The probability of membership of a given detectability class (π) presented the same temporal trends and variation in adult males, females (sex ad) and subadults \(\pi \left( {sex_{\text{ad}} + 1y + t} \right).\) The apparent survival probabilities (ϕ) varied independently over years between high and low detectable individuals (H × t) while sex and age effects were modeled as additive. Adult males and females in both detectability class were allowed to have different intercepts (H × sex ad), but no different trends over years (i.e., low number of female resightings). Subadults were considered independently \(\phi \left( {H \; \times \; \left( {sex_{\text{ad}} \; + \; t} \right) \; + \; 1y} \right)\). The detection probabilities of high and low detectable individuals varied independently over years (H × t), the sex effect was modeled as additive with different intercepts for males and females in both detectability \(p\left( {H \; \times \; \left( {t\; + \; sex} \right)} \right)\), but no differences in temporal pattern. Our most complex model was then \(\left\{ {\tau \left( {ad_{\text{males}} } \right),\pi \left( {sex_{\text{ad}} \; + \; 1y\; + \; t} \right),\phi \left( {H \; \times \; \left( {sex_{\text{ad}} + t} \right) \; + \; 1y} \right),p\left( {H\; \times \; \left( {sex\; + \; t} \right)} \right)} \right\}\) and constituted our starting point. Keeping the apparent survival probability time dependent (our parameter of interest), we followed a step-down approach (Lebreton et al. 1992) and consecutively modelled the transience parameter, the probability of detection and the probability of membership in a given detectability class. As the last step we modelled apparent survival probabilities.
Environmental variables
We tested the influence of environmental covariates on temporal variation of apparent survival probabilities. We considered local weather and regional climate indices as covariates of the survival probability. Foraging conditions in Friesland are crucial for staging ruffs to combine refuelling and the development of nuptial moult (Jukema et al. 2001). To fuel up, ruffs rely on soil-dwelling invertebrates (van Rhijn 1991; Verkuil and de Goeij 2003; J. Onrust, unpublished data) made less available by the intensive management of grasslands (Vickery et al. 2001). In absence of comprehensive data on the yearly variations in invertebrate prey availability for different land use categories, we assumed that in this intensively managed landscape foraging conditions for ruffs will mainly be modulated by weather conditions. Probing for and finding soil-dwelling invertebrates will get harder as the vegetation grows and as the soil dries during the spring season. Hence, we assumed that both dry and early springs might negatively affect refuelling rates and thereby influence survival. Growing degree day (GDD), the sum of average daily temperature relative to a temperature threshold of 0 °C, has been shown to be a relevant measure of the onset of spring growth of graminoids in the Netherlands (van Wijk et al. 2012). Here we used the cumulative values of GDD from 1 January to 15 March as a measure of seasonal advancement at the moment that ruffs started to stage in the study area. To characterize wet and dry springs we used the total amount of precipitation during spring stopover period (from 15 March to 15 May). Both precipitation and temperature data were obtained from Stavoren (52°52′56.8″N; 5°21′58.0″E) situated in the south of our study area (www.knmi.nl).
To indicate the regional weather pattern over most of the East Atlantic migration flyway, we used a principal component (PC) based North Atlantic Oscillation (Hurrell 2003), provided by NCAR’s Climate Analysis Section (http://climatedataguide.ucar.edu/guidance/hurrell-north-atlantic-oscillation-nao-index-pc-based). Upon arrival on the breeding grounds, snow cover will make food scarce or unavailable increasing risk of starvation (Meltofte et al. 2007; Buehler and Piersma 2008). We used NAO average values from December through March in the winter preceding breeding (pre-breeding NAO or PBNAO) as a measure of spring snow melt conditions (Stenseth et al. 2003; Kausrud et al. 2008). A small number of ruffs does not reach Africa but remains in Europe in winter, mainly in the Netherlands, Belgium and the United Kingdom (Prater 1973; Castelijns et al. 1994; Girard 2009; L.E. Schmaltz, unpublished data). These birds could represent a non-negligible proportion of the remaining spring passage population of ruffs in Friesland. Thus, we included winter NAO values from October to January reflecting mild and wet winter (positive NAO) vs. cold and dry winter (negative NAO) over northern Europe (Stenseth et al. 2003).
For ruffs wintering in the Sahelian floodplains of West-Africa, dry winters will cause small extents of open water areas to shrink more rapidly and hence force birds to concentrate on fewer feeding and roosting areas (Kone et al. 2002). During the pre-migratory period, this will lead to higher competition and hunting pressure (Zwarts et al. 2009). As a covariate we considered the maximum flood extent calculated from aerial pictures made every winter from 2004–5 to 2010–11 from the inner Niger Delta floodplain (L. Zwarts, personal communication), the main wintering area for western migrant ruffs.
We used principal component analysis (PCA) of the 5 environmental variables to combine their variation in uncorrelated, so-called ‘synthetic’, covariates which allowed us to reduce the number of environmental covariates and prevent collinearity issues (Joliffe 2002; Juillet et al. 2012). This analysis was performed in software R (R Development Core Team 2008) using the ADE-4 package (Dray and Dufour 2007). We investigated the relationships between the synthetic covariates and annual variation in survival within our best supported time dependent survival model. All models of our analysis were fitted in software E-SURGE (Choquet et al. 2009b, version 1.8.5). Model selection was based on Akaike’s information criterion (QAICc) corrected for overdispersion and small sample sizes (Burnham and Anderson 2002). To confirm that deviance did not settle on local minima during maximum likelihood procedure, we ran each model with 3 set of different initial values (Choquet and Nogué 2011).
Results
Model selection on the transience parameter (τ), the probability of membership of a given detectability class (π) and the probability of detection (p)
A transience effect did not improve the fit of our general model to the data and thus was not retained (Table S1 in Electronic Supplementary Material (ESM)). The probability of membership in a high detectability class varied between years, in parallel in subadult and adult males and females over the years (Table S1 in ESM, model selection on π, model 1 \(\pi \left( {sex_{\text{ad}} \; + \; 1y \; + \; t} \right)\)). Subadults had the highest probability to be assigned to the high detectability class, then adult males and finally adult females (Fig. 2a). In the multievent approach, the probability to belong to the high detectability class provide an estimate of the actual proportion of high detectable individuals within newly marked birds (Pradel 2005). The probability of membership in a high detectability class showed an increase with time (Fig. 2a), which would indicate an increase of the fraction of highly detectable birds within newly captured individuals in the course of this study. However, we acknowledge that this interpretation has to be taken with prudence. As our study is relatively short, intervals of confidence become rapidly larger (i.e., the capture histories of newly marked birds getting shorter) and assignment to either detectability class less reliable with the years.

Annual variation in probability of membership in the High Detectability class (π HD) (a), in the probability of detection (p) (b), and in the probability of apparent survival (ϕ) in females (c) and males (d) ruffs migrating through Friesland (the Netherlands) upon northward migration from 2004 to 2011. Estimates are based on the best supported model \(\left\{ {\pi \left( {sex_{\text{ad}} + 1y + \;t} \right),\phi \left( {H \times sex + \; t} \right),\;p\left( {H + sex + t} \right)} \right\}\) (model 8—Table 1). The error bars show 95 % confidence intervals
Detection probabilities varied with years in the same way between males and females (Table S1 in ESM, model selection on p, model 3 p(H + sex + t)), so that detection probabilities were much higher for males (\(\bar{p}_{\text{HD}}\) = 0.68, range 0.50–0.75; \(\bar{p}_{\text{LD}}\) = 0.21, range 0.18–0.33; Fig. 2b) than for females (\(\bar{p}_{\text{HD}}\) = 0.47, range 0.27–0.62; \(\bar{p}_{\text{LD}}\) = 0.08, range 0.04–0.15; Fig. 2b). In both sexes, the difference in detection probabilities between the two detectability classes was relatively important.
Model selection on the apparent survival probability (ϕ)
The best fitted parameterization on apparent survival probability indicated inter-annual variation (Table 1, model 8 ϕ(H × sex + t)). Apparent survival estimates decrease strongly at the end of the study period (Fig. 2c, d). A linear trend in survival probability was not supported (Table 1). Individuals at high and low detectability exhibited quantitative differences in survival, specific in opposite way for each sex (i.e., different intercepts—H × sex). According to the best supported model, the apparent survival probability of males at high detectability was higher than for males at low detectability (Fig. 2d—\(\bar{\phi }_{\text{HD males}}\) = 0.74, range: 0.51–0.93; \(\bar{\phi }_{\text{LD males}}\) = 0.64, range: 0.37–0.89), but the inverse was found for females (Fig. 2c—\(\bar{\phi }_{\text{HD females}}\) = 0.51, range 0.24–0.81; \(\bar{\phi }_{\text{LD females}}\) = 0.73, range 0.48–0.93). We only found slight support for age differences in survival. The second best model distinguishing subadults from adult males and females fitted equally well (model 8 vs. model 6, ΔQAICc = 0.88, Table 1).
That variation in apparent annual survival between individuals of high and low detectability were independent was not strongly supported (model 8 vs. model 3, ΔQAICc ≥ 4.37, Table 1). Models considering no difference in survival between individuals at high and low detectability poorly represented our data (ΔQAICc ≥ 10.79, Table 1). Finally, our best fitting model (model 8 \(\left\{ {\pi \left( {sex_{\text{ad}} + 1y + t} \right),\phi \left( {H \times sex + t} \right),p\left( {H + sex + t} \right)} \right\}\), Table 1), which included heterogeneity in detection probability, received much better support than an equivalent model under a traditional Cormack-Jolly-Seber model accounting for trap-dependence on males (ΔQAICc = 86.21, Table 1).
Environmental variables
In the analysis of environmental variables, we retained the two first principal components from the PCA (Eigenvalues >1, see Fig. S1 in ESM). The first and second principal component (F1, F2) accounted for 50 and 30 % of the total variation in the 5 covariates, respectively. Pre-breeding NAO index, winter NAO index and growing degree day were positively correlated and contributed the most to F1 (see Table S2 and Fig. S1 in ESM). Positive values of F1 were thus associated to an early onset of spring and early snow melt on the breeding grounds, and wet and mild winter conditions over northwestern Europe. F2 was positively associated to wet spring conditions on the staging site, but lower flood level in Niger during winter (only relatively to the study period—Table S2 and Fig. S1 in ESM). Hereafter, F1 and F2 were noted respectively as GDDspr/NAOs and PPspr/Flood. Incorporation of the synthetic covariates in our best model (model 8) did not improve the fit to our data and actually received less support than a ‘constant’ model (Table 2). A model which combined the additive effects of both F1 and F2 did not provide better fits.
Discussion
This study examined the year to year variation in apparent survival of western migrant ruffs of different age and sex staging in Friesland, the Netherlands, during northward migration. Using a capture-mark-resight approach, we found evidence for heterogeneity in the detection probability among colour-marked ruffs. We used multievent models integrating high and low detectability classes which allowed us to overcome bias and to investigate the biological meaning of heterogeneous detection. Since 2004 apparent survival probability of western migrant ruffs varied between years. Overall, apparent survival differed according to sex and the detectability of individuals. The males in the high detectability class had a higher apparent survival than the males in the low detectability class, whereas in the females this pattern was reversed.
Annual variation in apparent survival probability of western ruffs could not be explained by the local spring weather conditions, by the broad climate index across the flyway, or by the Inner Niger Delta flood level. During our study, the flood levels of the Inner Niger Delta were relatively high compared to levels during the droughts in the 1980s (L. Zwarts, personal communication) which were responsible of mass mortality events (Zwarts et al. 2009). This might explain why we found no relationship between flood extent and apparent survival within our study period. Factors explaining variation in apparent survival thus remain unknown. The general insights on human-induced habitat changes causing the decline in Afro-Palearctic migrants are there (Vickery et al. 2014), but to fully establish these relationships we need more targeted ecological data.
Survival varied between years and suggest a decline over time. We acknowledge that this pattern to some extent was linked to flag loss. However, the loss of flags likely happened gradually, as the plastic wears out with time. A higher mortality and/or that less bird return to Friesland in the end of our study period may be also involved. The latter would be consistent with previous studies suggesting redistribution of western ruffs towards more eastern migratory routes (Rakhimberdiev et al. 2010; Verkuil et al. 2012).
During the analysis of apparent annual survival it was necessary to allow for detectability differences between groups of animals. This significantly improved the fit of the models and also uncovered biologically interesting patterns. There are several ideas about how behaviour may affect detectability: (1) the ranging pattern of individuals relative to the study area may consistently differ between birds (e.g., Fearnbach et al. 2012). Although our observation effort covers the core staging area of ruffs in Friesland, individuals with ‘home ranges’ edging our study site would have smaller chances to be resighted than birds foraging exclusively within its limits. (2) the overall distribution of staging ruffs may have concentrated in certain areas (see Fig. 1a, L.E. Schmaltz, unpublished data). Therefore the ruffs were easier to find for the observers thereby increasing overall detectability of the birds. Such interactions between ecologically relevant behaviours of the birds and our way to detect them could explain an increase of the fraction of highly detectable birds within newly captured individuals during the study. (3) intrinsic behavioural features such as migration strategies may differ between staging ruffs. In long-distant migrants, a staging area typically harbours a mixture of individuals with different wintering area, timing of migration and breeding latitudes (e.g., Duijns et al. 2012; Franks et al. 2012), and/or staging duration (e.g., Dierschke et al. 2004). For instance, in a population of western gray whales (Eschrichtius robustus) monitored from feeding grounds upon migration (i.e., equivalent to a staging area), detectability differences between individuals were related to their residence time in the study area (Bradford et al. 2006). Our three explanations for detection heterogeneity are non-mutually exclusive. We conclude that detectability difference should overall be related to the degree of site use, either in space (relatively to the study area) and/or time (fidelity, staging duration).
Among the birds the most reliant on Friesland (i.e., the class of highly detectable individuals), for which apparent survival probability approaches most true survival, adult females survive much less than adult males. In accordance, between 2004 and 2011, the numbers of females captured decreased faster (with 53 %, see Fig. 1c) than the numbers of males (with 42 %). A lower female survival is unexpected in ruffs, as in many other polygynous vertebrate species males usually have a higher mortality than females (Trivers 1972). Together with a loss of females, this hints at female-specific vulnerability to conditions encountered somewhere along the western route. Zwarts et al. (2009) warned, for instance, that the tens of thousands of ruffs hunted in Mali for commercial purpose are mainly females that face higher risk due to their delayed departure compare to males coinciding with lower water levels and therefore higher hunting pressure (Kone et al. 2002). A selective loss of females would have severe repercussion on recruitment, while perturbation of local operational sex ratio could also expose the population of western ruffs, already much reduced in size, to higher risk of local extinction (Courchamp et al. 2008; Lee et al. 2011).
Interestingly, unlike females, males in the high detectability class, which we believe are most reliant on Friesland, had a higher apparent survival than the low detectable males that would be less reliant on Friesland. These inverse survival differences according to detectability and sexes are intriguing but the lack of information outside our study area limit further interpretation. Nevertheless it suggests that males and females with diverse migratory patterns and fidelity to Friesland, might contribute differently to the population dynamics of western ruffs.
Staging areas as Friesland are the crossroads where very different individuals with respect to origin, destination, behaviour and fidelity pass through. To monitor the survival of a migratory species from a single staging area is certainly challenging: emigration and mortality will always be confounded and the staging population might be very heterogeneous. Our results point at sex-specific behavioural variability among ruffs staging in Friesland associated with different apparent survival probabilities. Future work is needed to better characterize the apparent survival of these staging ruffs and investigate the influence of their migratory features. The low survival of highly detectable females and their concomitant decrease in numbers in Friesland might greatly affect the population dynamics of western ruffs and are in need to be investigated further. To unravel population processes at play, efforts should be made to collect data on female selective harvesting in the Sahel, to study reproductive success and widen our knowledge on migratory connectivity, inter-flyway movements and sex-specific dispersal aptitudes. To achieve such goals, the capture-mark-resight monitoring should be extended to multiple staging sites, combined to a multi-year satellite telemetry study conducted in both males and females as well as the establishment of throughout breeding monitoring. As ruffs are a very opportunistic species highly dependent on shallow waterbodies and soft substrates during the entire life cycle (van Rhijn 1991), all efforts to preserve these habitat characteristics throughout its migratory flyway will benefit the species. Such large scale and integrative approach should be a priority to protect and manage this widespread migrating species with weak connectivity in times of rapid global change.
References
Aharon-Rotman Y, Soloviev M, Minton C, Tomkovich P, Hassell C, Klaassen M (2015) Loss of periodicity in breeding success of waders links to changes in lemming cycles in Arctic ecosystems. Oikos 124:861–870
Alerstam T, Hedenström A, Åkesson S (2003) Long-distance migration: evolution and determinants. Oikos 2:247–260
Anthony LL, Blumstein DT (2000) Integrating behaviour into wildlife conservation: the multiple ways that behaviour can reduce N e. Biol Conserv 95:303–315
Arnason AN, Mills KH (1981) Bias and loss of precision due to tag loss in Jolly-Seber estimates for mark–recapture experiments. Can J Fish Aquat Sci 38:1077–1095
Berger J (2004) The last mile: how to sustain long-distance migration in mammals. Conserv Biol 18:320–331
Bradford A, Wade P, Weller D, Burdin A, Ivashchenko Y, Tsidulko G, VanBlaricom GR, Jr Brownell R L (2006) Survival estimates of western gray whales Eschrichtius robustus incorporating individual heterogeneity and temporary emigration. Mar Ecol- Prog Ser 315:293–307
Buehler DM, Piersma T (2008) Travelling on a budget: predictions and ecological evidence for bottlenecks in the annual cycle of long-distance migrants. Phil Trans R Soc B-Biol Sci 363:247–266
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Castelijns H (1994) Black-tailed godwit Limosa limosa islandica and ruff Philomachus pugnax winter in increasing numbers in Zeeuws-Vlaanderen (SW-Netherlands). Limosa 67:113–115 (in Dutch with English abstract)
Caswell H (2001) Matrix population models: construction, analysis and interpretation, 2nd edn. Sinauer Associates, Sunderland
Choquet R, Nogué E (2011) E-SURGE 1-8 user’s manual. CEFE, Montpellier
Choquet R, Lebreton J-D, Gimenez O, Reboulet A-M, Pradel R (2009a) U-CARE: utilities for performing goodness of fit tests and manipulating CApture-REcapture data. Ecography 32:1071–1074
Choquet R, Rouan L, Pradel R (2009b) Program E-SURGE: a software application for fitting multievent models. In: Thomson DL, Cooch EG, Conroy MJ (eds) Modeling demographic processes in marked populations. Springer, US, New York, pp 845–865
Cormack RM (1964) Estimates of survival from the sighting of marked animals. Biometrika 51:429–438
Courchamp F, Berec L, Gascoigne J (2008) Allee Effects in ecology and conservation. Oxford University Press, New York
Crespin L, Choquet R, Lima M, Merritt J, Pradel R (2008) Is heterogeneity of catchability in capture–recapture studies a mere sampling artifact or a biologically relevant feature of the population? Popul Ecol 50:247–256
Dierschke V, Mendel B, Schmaljohann H (2004) Differential timing of spring migration in northern wheatears Oenanthe oenanthe: hurried males or weak females? Behav Ecol Sociobiol 57:470–480
Dray S, Dufour A (2007) The Ade4 package: implementing the duality diagram for ecologists. J Stat Softw 22:1–20
Duijns S, Jukema J, Spaans B, van Horssen P, Piersma T (2012) Revisiting the proposed leap-frog migration of bar-tailed godwits along the East-Atlantic flyway. Ardea 100:37–43
Fearnbach H, Durban J, Parsons K, Claridge D (2012) Photographic mark-recapture analysis of local dynamics within an open population of dolphins. Ecol Appl 22:1689–1700
Franks SE, Norris DR, Kyser TK, Fernández G, Schwarz B, Carmona R, Colwell MA, Sandoval JC, Dondua A, River Gates H, Haase B, Hodkinson DJ, Jiménez A, Lanctot RB, Ortego B, Sandercock BK, Sanders F, Takekawa JY, Warnock N, Ydenberg RC, Lank DB (2012) Range-wide patterns of migratory connectivity in the western sandpiper Calidris mauri. J Avian Biol 43:155–167
Gascoigne J, Berec L, Gregory S, Courchamp F (2009) Dangerously few liaisons: a review of mate-finding Allee effects. Popul Ecol 51:355–372
Gimenez O, Viallefont A, Charmantier A, Pradel R, Cam E, Brown CR, Anderson MD, Bomberger Brown M, Covas R, Gaillard J-M (2008) The risk of flawed inference in evolutionary studies when detectability is less than one. Am Nat 172:441–448
Girard O, Scott D, Dodman T (2009) Ruff, Philomachus pugnax. In: Delany S, Scott D, Dodman T, Stroud S (eds) An atlas of wader populations in Africa and western Eurasia. Wageningen, Wetlands International
Glutz von Blotzheim UN, Bauer KM, Bezzel E (1975) Handbuch der vögel mitteleuropas: Charadriiformes (1. Teil), Volume 6. Akademische Verlag, Wiesbaden (in German)
Groen NM, Kentie R, de Goeij P, Verheijen B, Hooijmeijer JCEW, Piersma T (2012) A modern landscape ecology of black-tailed godwits: habitat selection in southwest Friesland, the Netherlands. Ardea 100:19–28
Harrison XA, Blount JD, Inger R, Norris DR, Bearhop S (2011) Carry-over effects as drivers of fitness differences in animals. J Anim Ecol 80:4–18
Hogan-Warburg AJ (1966) Social behavior of ruff, Philomachus pugnax. Ardea 54:109–229
Höglund J (1996) Can mating systems affect local extinction risks? Two examples of lek-breeding waders. Oikos 77:184–188
Hurrell JW, Kushnir Y, Ottersen G, Visbeck M (2003) The North Atlantic oscillation: climatic significance and environmental impact. American Geophysical Union, Washington
Ims RA, Henden JA, Killengreen ST (2008) Collapsing population cycles. Trends Ecol Evol 23:79–86
Jolliffe IT (2002) Principal component analysis, 2nd edn. Springer, New York
Jolly GM (1965) Explicit estimates from capture-recapture data with both death and immigration-stochastic model. Biometrika 52:225–247
Juillet C, Choquet R, Gauthier G, Pradel R (2010) A capture–recapture model with double-marking, live and dead encounters, and heterogeneity of reporting due to auxiliary mark loss. J Agr Biol Environ Stat 16:88–104
Juillet C, Choquet R, Gauthier G, Lefebvre J, Pradel R (2012) Carry-over effects of spring hunt and climate on recruitment to the natal colony in a migratory species. J Appl Ecol 49:1237–1246
Jukema J, Piersma T (2000) Contour feather moult of ruffs Philomachus pugnax during northward migration, with notes on homology of nuptial plumages in scolopacid waders. Ibis 142:289–296
Jukema J, Piersma T (2006) Permanent female mimics in a lekking shorebird. Biol Lett 2:161–164
Jukema J, Wymenga E, Piersma T (2001) Stopping over in SW Friesland: fattening and moulting in ruffs Philomachus pugnax during northward migration in the Netherlands. Limosa 74:17–26 (in Dutch with English abstract)
Karlionova N, Meissner WW, Pinchuk P (2008) Differential development of breeding plumage in adult and second-year male ruffs Philomachus pugnax. Ardea 96:39–45
Kausrud KL, Mysterud A, Steen H, Vik JO, Østbye E, Cazelles B, Framstad E, Eikeset AM, Mysterud I, Solhøy T, Stenseth NC (2008) Linking climate change to lemming cycles. Nature 456:93–97
Kentie R, Both C, Hooijmeijer JCEW, Piersma T (2014) Age-dependent dispersal and habitat choice in black-tailed godwits Limosa limosa limosa across a mosaic of traditional and modern grassland habitats. J Avian Biol 45:396–405
Kone B, Diallo M, Fofana B (2002) Exploitation des oiseaux d’eau. In: Wymenga E, Kone B, van der Kamp J, Zwarts L (eds) Delta intérieur du fleuve Niger: Ecologie et gestion durable des ressources naturelles. A&W/Wetlands International/Rijkswaterstaat, Veenwouden, pp 201–207 (in French)
Lebreton J-D, Burnham KP, Clobert J, Anderson DR (1992) Modeling survival and testing biological hypotheses using marked animals: a unified approach with case studies. Ecol Monogr 62:67–118
Lee AM, Saether BE, Engen S (2011) Demographic stochasticity, allee effects, and extinction: the influence of mating system and sex ratio. Am Nat 177:301–313
Limburg KE, Waldman JR (2009) Dramatic declines in North Atlantic diadromous fishes. Bioscience 59:955–965
Lindström Å, Green M (2013) Monitoring population changes of birds in Sweden. Annual report for 2012. Departement of Ecology, Lund University (in Swedish)
Meltofte H, Piersma T, Boyd H, McCaffery B, Ganter B, Golovnyuk VV, Graham K, Gratto-Trevor CL, Morrison RIG, Nol E, Rösner H-U, Schamel D, Schekkerman H, Soloviev MY, Tomkovich PT, Tracy CM, Tulp I, Wennerberg L (2007) Effects of climate variation on the breeding ecology of Arctic shorebirds. Meddelelser om Grønland Bioscience 59. Danish Polar Center 2007, Copenhagen
Øien IJ, Aarvak T (2010) Brushanen forsvinner. Vår Fuglefauna 33:162–173 (in Norwegian)
Piersma T (2007) Using the power of comparison to explain habitat use and migration strategies of shorebirds worldwide. J Ornithol 148:S45–S59
Piersma T, Rogers KG, Boyd H, Bunskoeke EJ, Jukema J (2005) Demography of Eurasian Golden Plovers Pluvialis apricaria staging in the Netherlands, 1949–2000. Ardea 93:49–64
Pradel R (2005) Multievent: an extension of multistate capture-recapture models to uncertain states. Biometrics 61:442–447
Pradel R, Sanz-Aguilar A (2012) Modeling trap-awareness and related phenomena in capture-recapture studies. PLoS One 7:e32666
Pradel R, Hines JE, Lebreton J-D, Nichols JD (1997) Capture-recapture survival models taking account of transients. Biometrics 53:60–72
Pradel R, Gimenez O, Lebreton J-D (2005) Principles and interest of GOF tests for multistate capture—recapture models. Anim Biodivers Conserv 28:189–204
Prater AJ (1973) The wintering population of ruffs in Britain and Ireland. Bird Study 20:245–250
R Development Core Team (2008) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rakhimberdiev E, Verkuil YI, Saveliev AA, Väisänen RA, Karagicheva J, Soloviev MY, Tomkovich PS, Piersma T (2010) A global population redistribution in a migrant shorebird detected with continent-wide qualitative breeding survey data. Divers Distrib 17:144–151
Sæther BE, Bakke Ø (2000) Avian life history variation and contribution of demographic traits to the population growth rate. Ecology 81:642–653
Sanderson FJ, Donald PF, Pain DJ, Burfield IJ, van Bommel FPJ (2006) Long-term population declines in Afro-Palearctic migrant birds. Biol Conserv 131:93–105
Seber GAF (1965) A note on multiple-recapture census. Biometrika 52:249–259
Stenseth NC, Ottersen G, Hurrell JW, Mysterud A, Lima M, Chan K-S, Yoccoz NG, Ådlandsvik B (2003) Studying climate effects on ecology through the use of climate indices: the North Atlantic Oscillation, El Niño Southern Oscillation and beyond. Proc R Soc B-Biol Sci 270:2087–2096
Taylor CM, Norris DR (2007) Predicting conditions for migration: effects of density dependence and habitat quality. Biol Lett 3:280–284
Taylor CM, Norris DR (2010) Population dynamics in migratory networks. Theor Ecol 3:65–73
Thorup O (2006) Breeding waders in Europe 2000. Int Wader Stud 14:1–142
Trivers RL (1972) Parental investment and sexual selection. In: Campbell B (ed) Sexual selection and the descent of man 1871–1971. Aldine, Chicago, pp 136–179
Väisänen RA (2005) Monitoring population changes of 86 land bird species breeding in Finland in 1983–2005. Linnut-Vuosikirja 2005:83–98 (in Finnish with English abstract)
van Rhijn JG (1991) The ruff: individuality in a gregarious wading bird. T. and A.D Poyser, London
van Wijk RE, Kölzsch A, Kruckenberg H, Ebbinge BS, Müskens GJDM, Nolet BA (2012) Individually tracked geese follow peaks of temperature acceleration during spring migration. Oikos 121:655–664
Verkuil Y, de Goeij P (2003) Do Reeves make different choices? Meadow selection by spring staging ruffs Philomachus pugnax in southwest Friesland. Limosa 76:157–168 (in Dutch with English abstract)
Verkuil YI, Wijmenga JJ, Hooijmeijer JCEW, Piersma T (2010) Spring migration of ruffs Philomachus pugnax in Fryslân: estimates of staging duration using resighting data. Ardea 98:21–33
Verkuil YI, Karlionova N, Rakhimberdiev EN, Jukema J, Wijmenga JJ, Hooijmeijer JCEW, Pinchuk P, Wymenga E, Baker AJ, Piersma T (2012) Losing a staging area: eastward redistribution of Afro-Eurasian ruffs is associated with deteriorating fuelling conditions along the western flyway. Biol Conserv 149:51–59
Vickery JA, Tallowin JR, Feber RE, Asteraki EJ, Atkinson PW, Fuller RJ, Brown VK (2001) The management of lowland neutral grasslands in Britain: effects of agricultural practices on birds and their food resources. J Appl Ecol 38:647–664
Vickery JA, Ewing SR, Smith KENW, Pain DJ, Bairlein F, Gregory RD, Skorpilov J (2014) The decline of Afro-Palaearctic migrants and an assessment of potential causes. Ibis 156:1–22
Virkkala R, Rajasärkkä A (2011) Northward density shift of bird species in boreal protected areas due to climate change. Boreal Environment Research 16(Suppl. B):2–13
Webster MS, Marra PP, Haig SM, Bensch S, Holmes RT (2002) Links between worlds: unraveling migratory connectivity. Trends Ecol Evol 17:76–83
Widemo F (1998) Alternative reproductive strategies in the ruff, Philomachus pugnax: a mixed ESS? Anim Behav 56:329–336
Wilcove DS, Wikelski M (2008) Going, going, gone: is animal migration disappearing? PLoS Biol 6:e188
Wymenga E (1999) Migrating ruffs Philomachus pugnax through Europe, spring 1998. Wader Study Group Bull 88:43–48
Zöckler C (2002a) A comparision between tundra and wet grassland breeding waders with special reference to the ruff (Philomachus pugnax). Fed Agency Nat Conserv, Bonn
Zöckler C (2002b) Declining Ruff Philomachus pugnax populations: a response to global warming? Wader Study Group Bull 97:19–29
Zwarts L, Bijlsma RG, van der Kamp J, Wymenga E (2009) Living on the edge: wetlands and birds in a changing Sahel. KNNV Publishing, Zeist
Acknowledgments
We thank the Frisian “wilsternetters” Albert Anne Mulder, Doede Anne Mulder, Eeltje Anne Mulder, Fons Baarsma, Jappie Boersma, Cees Dekker, Piet Feenstra, Albert Hendrik Mulder, Doede Hendrik Mulder, Douwe de Jager, Bauke de Jong, Joop Jukema, Bauke Kuipers, Willem Louwsma, Catharinus Monkel, Rein Mulder, Jaap Strikwerda, Fokke Tuinstra, Bram van der Veen, F. van der Veen, Arend Veenstra, Sierd Visser, Willem Visser, Piet Vlas, Douwe van der Zee, and Rinkje van der Zee for their dedication to catch Ruffs since 2004. We thank all the volunteers and independent observers who contributed to colour-mark and/or resight ruffs over the years. We thank Olaf Klaassen, Jelle Postma and Chris van Turnhout (all SOVON) for making available updated information on ruff roost-site counts. This study was carried out under license of Animal Experimentation Committee (DEC) of the University of Groningen in accordance to the Dutch laws. This work was financially supported by an Ubbo Emmius PhD grant from the University of Groningen, supplemented by the Province of Friesland and a start-up grant of the University of Groningen to TP, and by smaller grants of the GUF/Gratama Foundation (Project 04.05) and the Schure-Beijerinck Popping Foundation (SBP/JK2006-39 and SBP/JK2007-34) to YIV and JCEWH.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Schmaltz, L.E., Juillet, C., Tinbergen, J.M. et al. Apparent annual survival of staging ruffs during a period of population decline: insights from sex and site-use related differences. Popul Ecol 57, 613–624 (2015). https://doi.org/10.1007/s10144-015-0511-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10144-015-0511-4