
Abstract
Infection with herpes simplex viruses is one of the most ancient diseases described to affect humans. Infection with these viruses produces vexing effects to the host, which frequently recur. Infection with herpes simplex viruses is lifelong, and currently there is no vaccine or drug to prevent or cure infection. Prevalence of herpes simplex virus 2 (HSV-2) infection varies significantly depending on the geographical region and nears 20 % worldwide. Importantly, HSV-2 is the first cause of genital ulcers in the planet. HSV-2 affects approximately 500 million people around the globe and significantly increases the likelihood of acquiring the human immunodeficiency virus (HIV), as well as its shedding. Thus, controlling HSV-2 infection and spread is of public health concern. Here, we review the diseases produced by herpes simplex viruses, the factors that modulate HSV-2 infection, the relationship between HSV-2 and HIV and novel therapeutic and prophylactic microbicides/antivirals under development to prevent infection and pathological outcomes produced by this virus. We also review mutations associated with HSV-2 resistance to common antivirals.
Similar content being viewed by others


Introduction
Humans and herpes simplex viruses (HSV) have interacted with each other for tens of thousands of years, as recently evidenced by phylogenetic analyses that relate HSV serotypes to the intercontinental migration of our ancestors [1]. Noteworthy, allusions to the diseases produced by these viruses date back to 2,000 years BC and the word herpes to ~400 BC by Hippocrates, who referred to the pathological outcomes produced by these viruses with the term herpein (to crawl in Greek), likely because of the pain that crawled through the skin of those affected [2, 3]. However, in the following centuries, the term herpes became widely used to describe skin lesions produced by numerous agents and conditions that not necessarily related to HSV. A more detailed description of the effects produced by HSV came hundreds of years later with the publication of De Morbis Venereis by John Astruc in 1736 that narrated with detail the pathological manifestations of herpes genitalis [4]. Later studies would describe the recurrent character of the disease, suggesting persistent infection [5, 6]. However, regardless of the early perception that herpetic diseases were transmissible from person to person, the infectious character of these pathologies only became evident at the end of the ninth century and beginning of the twentieth century. Experimental work performed by J. Vidal in humans [7] and Gruter in rabbits [8] demonstrated for the first time that herpes could be transmitted between individuals with material obtained from herpetic vesicles. Although these findings warned people about the infectivity of this disease, its threat has been somewhat underestimated since then with lesions commonly being described as “cold sores” and “fever blisters” in the oral area. Regretfully, these popular terms undervalue the persistent nature of HSV. Currently, the term herpes has found again common use to appropriately indicate disease produced by two closely related, but different viruses: herpes simplex virus 1 (HSV-1) and herpes simplex virus 2 (HSV-2). Both HSV-1 and HSV-2 produce similar lesions that recur with variable frequency in skin and mucosae throughout the individual’s life and latently infect neurons of the trigeminal or dorsal root ganglia [9]. These neurotropic characteristics of HSV and their capacity to escape the host immune response make these viruses successful pathogens, as evidenced by their capacity to persist within individuals and remain at a high prevalence in the population [10].
Clinical manifestations of HSV-2
Infection with HSV-1 or HSV-2 generally requires intimate contact with an infected individual (e.g., sexual intercourse and saliva exchange). However, direct contact of infected tissues with mucosae (e.g., eyes, genitalia) or eroded skin (e.g., with abrasions, microlesions) also provides entry portals for HSV in the host [11]. Although herpetic lesions in the oro-facial area are commonly attributed to HSV-1 and herpetic lesions in the genital area are associated with HSV-2, infections at both sites can occur with these two viruses [12, 13].
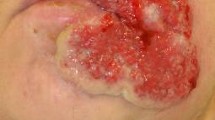
Common pathologies produced by HSV include gingivostomatitis, herpes labialis and herpes genitalis, which are generally characterized by vesicular ulcers, although these may be less evident in the genital area [13–16]. Herpetic lesions contain high amounts of virions and infiltrating leukocytes that combat infection and last 2–21 days followed by the establishment of scabs that are resolved without scarification. HSV-1 and HSV-2 can also result in multiple other skin diseases, such as eczema herpeticum in individuals with atopic dermatitis that can be life-threatening for infants [17, 18], herpetic whitlow, which manifests as lesions at the finger tips [19, 20] and herpes gladiatorum [11, 21], an herpetic condition facilitated by eroded skin and frequently observed in individuals that practice contact sports such as wrestling (Fig. 1).

Clinical manifestations produced by herpes simplex viruses: the sites of infection by herpes simplex viruses include brain (meningitis, encephalitis), infections in the ocular area (eyes: keratitis, retinitis and conjunctivitis; mouth: lips and gingivostomatitis; facial nerves: facial paralysis), torso, limbs and genitals (neck, arms, hands, fingers, legs, genitals) and internal organs, such as lungs, kidneys and liver. The most frequent HSV serotype isolated for each pathology is indicated
Besides skin-related diseases, HSV are also associated with ocular complications, mainly epithelial or stromal keratitis (corneal infection with HSV) (Fig. 1). It is estimated that HSV is the first cause of blindness due to infection in the Western world [22, 23]. Although HSV-2 infects human corneal fibroblasts in vitro [24], as well as mouse corneas in the animal model [25], blindness by HSV in humans is generally attributed to HSV-1 [26–28]. Paradoxically, HSV infection of the retina (retinitis) is more frequently ascribed to HSV-2 in humans (Fig. 1) [29]. Similarly, acute necrotic retinitis is also frequently credited to HSV-2 [30, 31]. Other typical manifestations of HSV in the ocular area include eyelid infections and conjunctivitis (Fig. 1). These observations suggest that although both viruses share similarities in their capacity to infect cells derived from the ocular region, they may be isolated at slightly different frequencies depending on the disease produced.
A particularly relevant characteristic of HSV is that they also infect the central nervous system, which occurs more frequently in neonates than adults as a result of congenital infection from the mother to the newborn at the moment of birth. HSV infection of neonates can result in encephalitis and meningitis leading to severe neurological damage, such as mental retardation and to significant sequelae, as well as more dramatically death [32, 33]. This type of infection also occurs in adults, with HSV currently being the primary cause of viral encephalitis and meningitis worldwide [34, 35]. Although HSV-2 is capable of infecting the nervous system (0.016 % of cases), the main causative agent of encephalitis and meningitis in infants and adults is HSV-1 [35–37]. However, the number of cases of encephalitis and meningitis produced by HSV, as well as the specific virus producing these diseases, vary significantly between studies.
Another neurological illness associated with HSV is Mollaret’s syndrome (aseptic meningitis, recurrent benign leukocyte meningitis), in which one-third of the individuals suffer recurrent meningitis with spasm episodes, diplopia, pathological reflexes, cranial nerve paralysis, hallucinations and even coma (Fig. 1) [38–40]. This rare disease is characterized by aseptic inflammation of the meninges, which consists in leukocyte infiltration and the presence of Mollaret’s cells (giant mononuclear cells) in the inflamed tissue. Unlike encephalitis in adults, which is mainly produced by HSV-1, Mollaret’s syndrome is mainly attributed to HSV-2 [38, 39, 41]. Less clear is the participation of HSV in Bell’s palsy, a form of facial paralysis with unknown etiology (Fig. 1). Though HSV-1 is frequently referred as one of the causative agents of this disease [42, 43], other viruses, such as the human herpesvirus-6, have also been proposed to be involved in the onset of this form of facial paralysis [44, 45].
Although HSV infection is generally associated with skin, mucosal membrane and central nervous system pathologies, these viruses can also infect other tissues producing severe damage. Indeed, HSV infect a broad range of organs in the body, although in lower frequencies than the skin, mucosal membranes and eyes. Indeed, HSV can infect the liver [46, 47] and kidneys [47], and produce sepsis [46] (Fig. 1). These conditions rarely occur in immunocompetent individuals and usually predominate in immunocompromised and transplanted patients, as well as infants. Furthermore, HSV can also infect the lungs producing acute pulmonary damage (acute lung injury, ALI) and acute respiratory distress syndrome (ARDS) in both immunocompetent and immunocompromised individuals [47–49]. A recent study detected HSV-1 in 13 % of bronchoalveolar lavages obtained from patients with ALI/ARDS that were mainly intubated [50]. However, pulmonary infection with HSV is more frequently described in immunosuppressed individuals, such as those with advanced human immunodeficiency virus (HIV) infection, those undergoing chemotherapy and persons transplanted with solid organs or bone marrow [51–53].
Taken together, HSV produces a wide range of diseases in humans that can affect several organs. Although HSV-1 and HSV-2 may be isolated at different frequencies from the infected tissues, it is important to bear in mind that both viruses can produce similar diseases at these sites.
Incidence and prevalence of HSV-2
It is estimated that approximately 500 million people are currently infected with HSV-2 worldwide and that more than 20 million new cases arise each year [54]. The prevalence of HSV-2 varies significantly depending on the global region, ranging from ~5 % for countries such as Spain and up to 70 % for Sub-Saharan Africa [10, 54–56]. The United States currently displays a prevalence nearing ~16 %, which is below the global estimated average: 23.6 % [54, 57, 58]. For Europe, prevalence of HSV-2 varies significantly between countries ranging from ~5 % for England and Spain, ~20–30 % for countries such as Sweden, Denmark and Germany and up to 40 % for Turkey [56]. It is important to point out that the populations evaluated in these studies can vary significantly, as well as the methods used for determining seroprevalence (e.g., enzyme-linked immunosorbent assay vs. immunoblot) [56]. Remarkably, HSV-2 prevalence studies are rarely updated, and thus, prevalence values can significantly vary with time, as recently observed in a study that assessed pregnant women in Switzerland and reported prevalence values that jumped from 10 to 20 % in a 15-year period [59]. On the other hand, countries belonging to Asia and Oceania display prevalence rates ranging from ~5 % for New Zealand and Japan, ~10 % for Australia and the Philippines and up to ~20 % for China [56, 60]. For Latin America, there are relatively few epidemiological studies assessing the prevalence of HSV-2. One study suggests nearly 54 million cases for South America and the Caribbean [54]. A recent study in Peru determined that the prevalence of HSV-2 nears 15 % within 18- to 29-year-old individuals [61]. Brazil, on the other hand, displays a prevalence of HSV-2 close to 11 % [62], although certain studies suggest higher values [63, 64].
Noteworthy, the prevalence of HSV-2 infection is generally higher in women than in men with 50 % more cases occurring in the former group [54, 57, 65]. Although the reasons for these differences are unclear, it has been proposed that men might be more resistant to HSV-2 infection than women because of increased keratinization of the skin at the external surfaces of the genitalia.
Because HSV-2 infection is associated with susceptibility to HIV and increased shedding of the latter (see below), determining the prevalence of HSV-2 in the population is of public health relevance. Indeed, such epidemiological information could also help determine the effectiveness of sex education policies in different countries.
Although herpetic lesions at the oro-facial area (e.g., herpes labialis) are usually caused by HSV-1 and those occurring at the genitalia (herpes genitalis) are generally associated to HSV-2, HSV-1 is progressively emerging as the most common cause of primary genital herpes worldwide. Indeed, many studies have reported that HSV-1 may account for up to 80 % of cases of first-episode genital and anogenital herpes [12, 66–73]. Interestingly, a gradual transition from HSV-2 to HSV-1 as the leading causes of primary genital herpes can be observed in numerous recent studies [74–76]. Such a tendency would be explained in part by changes in the sexual behavior of the populations during the last decades [77]. On the other hand, up to 10 % of primary oro-facial herpes infections could be caused by HSV-2 [71]. However, this value may be underestimated because of infrequent serotypification or diagnosis of herpes labialis, which is shown in recent epidemiological studies [12]. Importantly, the capacity of both HSV-1 and HSV-2 to infect common tissues relies on the fact that the target receptors for these viruses are the same [78–80].
Because HSV-1 and HSV-2 remain latent in the host, variations in their reactivation/recurrence rates could significantly impact the frequencies at which each virus is isolated from a specific location in time. That is, increased incidence of a specific herpes simplex virus at a particular site might not necessarily relate to higher recurrences at that place in time, as this virus might recur there to a lesser extent than the other virus [81]. Indeed, despite the fact that HSV-1 is frequently isolated in primary genital infection, recurrences by this virus represent less than 10 % of the total cases of genital herpes [82, 83]. This apparent discrepancy would be explained by the low reactivation capacity of HSV-1 in the genital area, as compared to HSV-2 [81]. Effectively, it has been observed that HSV-1 reactivates significantly less in the genital area than HSV-2, the latter recurring 4–5 times a year, up to 6 times more than HSV-1 at this location [81, 84]. Inversely, it has been reported that HSV-2 recurs significantly less than HSV-1 at the oro-facial area [81]. Altogether, these findings provide a possible explanation for the different frequencies at which HSV-1 and HSV-2 are observed at specific anatomical sites during recurrences.
Infection with HSV does not always lead to symptoms or recurrences. However, 20–50 and 80–90 % of individuals that manifest primary symptomatic infections with HSV-1 and HSV-2, respectively, will develop genital recurrences during the first year of infection, while the remaining will rarely display any signs of disease [84]. Importantly, it has been observed that the frequency of future recurrences can be associated, at least in part with the severity of the primary infection episode [84]. On the other hand, the frequency and severity of recurrences by HSV tend to decreases in time, up to half the number of episodes over periods of 4 years [84]. The incidence of HSV-2 also varies greatly depending on the age of the individuals and is more pronounced once sexual activity is initiated. Currently, the highest infection rates in adults occur between ages 15 and 20 [54], although peak incidences at slightly higher ages are commonly observed in some populations [85].
The incidence of symptomatic neonate infection with HSV varies largely depending on the geographical region and time interval assessed. For instance, incidence rates in the United States have been shown to vary as much as from 1/25,000 to 1/1,500 live births when covering the years 1997 to 2010 [86–89]. Importantly, despite these low rates of detectable HSV infection at birth (symptomatic infection), prevalence of HSV-2 in children has been reported to be as high as 3–5 % and reach up to 15 % for some populations, such as children in urban regions of Tanzania [90, 91]. The significant differences between neonatal infection rates and prevalence of HSV infection in children could be due to unnoticed intrauterine or neonatal infection with HSV-2 [92], or non-sexual transmission of HSV by contaminated fingers and hands [13]. Noteworthy, mothers that are infected with HSV-2 recur more frequently as they progress through pregnancy, thus increasing the chances of virus transmission to the infant at the moment of birth [93]. These recurrences not necessarily need to be symptomatic to transmit infection, which complicates prevention of neonate infection [94, 95]. Nevertheless, the highest risk of infection to neonates occurs when mothers are undergoing primary infection and present severe HSV clinical manifestations [94, 96, 97]. Importantly, diagnosis of primary HSV infection at the time of delivery will require the analysis of both blood and swab samples for determining the type of infection [97].
Factors that modulate infection by HSV-2
Some factors that increase or decrease the probability of healthy individuals to acquire HSV-2 have been identified. One of these factors is the length of partner relationship, which correlates with the number of sexual intercourses. Indeed, it is estimated that 3.5 months of partner relationship or approximately 40 sexual intercourses are needed for an individual to be infected with HSV-2 by his/her partner [98]. This type of studies is possible thanks to the follow-up of discordant couples for HSV-2, which allow to identify risk factors associated with infection, as well as the probability of acquiring this virus [99]. The use of antivirals, such as valacyclovir (prodrug of acyclovir), has been shown to help decrease the probability of infection in this type of couples [100, 101]. Another factor that increases the probability of acquiring HSV-2 is evidently the number of sexual partners, which increases significantly the chances of encountering an HSV-2 infectious individual [102, 103]. On the other hand, factors that decrease the probability of infection with HSV-2 include the presence of natural antiviral molecules in the genital tissues. Indeed, infection with HSV-2 varies significantly between women depending on the presence of antiviral soluble compounds in the vagina that fluctuate with the hormonal cycle. For instance, human neutrophil peptides 1–3 (HNP1-3), interleukin 8 (IL-8), lactoferrin, lysozyme, immunoglobulin A (IgA) and IgG have been correlated with positive vaginal antiviral activities [104]. It is important to point out that individuals at the follicular phase present higher concentrations of these compounds as compared to individuals that use oral contraceptives, suggesting that the latter would be more vulnerable to infection with HSV-2 [104]. Consistent with this notion, some studies have associated the use of contraceptives with an increased risk of infection with HSV-2 and increased virus dissemination from the infected tissue [105, 106]. On the other hand, host molecules called defensins secreted in the genital tissue possess the capacity to block the infectivity of HSV-2. In vitro and in vivo experiments have demonstrated that human α-defensins (HD) HNP1-4, HD5 and HD6, as well as β-defensin 3 inhibit infection by HSV though the blockade of virus entry in target cells and during later processes [107]. Indeed, cervicovaginal washes of healthy patients contain these defensins and are able to reduce HSV-2 infection in vitro [108]. These results suggest that defensins present in cervicovaginal secretions play relevant roles in innate resistance to infection with HSV-2.
Aiming to evaluate possible differences between the susceptibility to HSV-2 infection between sexually active adolescent and adult women, some studies have quantified bacteria, such as Lactobacillus jensenii and Lactobacillus crispatus, in the genital tissue as biomarkers for mucosal immunity and to determine the relationship between these bacteria with susceptibility to infection by HSV-2. One of these studies suggests that adolescents posses significantly less activity against E. coli as compared to adult women, as well as reduced concentrations of total IgG and IgA which could account for lesser antimicrobial immunity [109]. However, adolescents presented significantly higher activities against HSV-2 in their vaginal secretions, as compared to adults and were more resistant to infection by this virus [109].
Taken together, we are just commencing to identify molecular factors that modulate infection at the genital tract, such as antimicrobial molecules present in this tissue, as well as their relationship with the infective capacity of pathogens. It is also important to consider the differential expression of these molecules in individuals of distinct ages for the design of future treatment strategies against vaginal infections.
Relationship between HSV-2 and HIV
An important finding in the last decades is the fact that infection with HSV-2 increases 3–4 times the likelihood of acquiring HIV, thus significantly contributing to the epidemics of acquired immunodeficiency syndrome (AIDS) [110–112]. Moreover, it has been evidenced that HSV-2 significantly increases the dissemination of HIV in the genitalia of those co-infected with these viruses [113]. It is important to note that the relationship between HIV and HSV-2 is reciprocal, as infection with HIV also significantly increases the probability of acquiring HSV-2, and that reactivation with HSV-2 is more frequent in HIV-positive patients [114–116]. Noteworthy, the relationship between HSV-2 and HIV is synergic and goes beyond risk factors associated with the acquisition of these viruses.
The molecular mechanisms that relate HSV-2 and HIV are just beginning to be elucidated. One of the mechanisms by which infection with HSV-2 increases the probability of acquiring HIV is through the generation of microlesions by HSV-2 at the site of infection, which exposes immune cells in the epidermis and dermis to HIV (Fig. 2). Effectively, immune cells such as dendritic cells (DCs), Langerhans cells and macrophages reside these zones and patrol the tissues to detect microbes (Fig. 2) [117]. These cells are important targets for HSV-2 and HIV [118, 119]. On the other hand, a recent study found that the foreskin of individuals infected with HSV-2 contained significantly higher number of CD4+ T cells than foreskins from non-infected individuals, thus increasing the probability that HIV encounters these target cells at this site (Fig. 2) [120]. Another clinical study found that vaginal infection with HSV-2 significantly increased the number of DCs (up to tenfold) and CD4+ T cells (threefold) at the site of infection, and that these cells expressed DC-SIGN and CCR5, respectively, both receptors for HIV (Fig. 2) [121]. Furthermore, it has been found that macrophages infected with HSV-2 increased their expression of CCR5 at their surface, rendering them more susceptible to HIV infection [122]. Finally, infection of Langerhans cells with HSV-2 reduced the expression of langerin in these cells, a molecule that mediates the capture and degradation of HIV [123]. These effects promote DC infection with HIV and consequent transfer of this virus to T cells in the tissue and lymph nodes (Fig. 2). Moreover, infection of dendritic cells with HSV-2 significantly increases the expression of integrin α4β7 at the surface of CD4+ T cells, which makes these cells more susceptible to HIV infection (Fig. 2) [119]. Taken together, infection with HSV-2 promotes the recruitment of immune cells at the site of infection that are targets for HIV, thus increasing the chances that these cells be successfully infected by the latter virus (Fig. 2).

Relationship between HSV-2 and HIV at the site of infection. Infection with HSV-2 produces microlesions that expose the epidermis, dermis and subjacent immune cells at the site of infection to incoming viruses, such as HIV. 1. Infection of genital epithelia by HSV-2 produces the secretion of pro-inflammatory cytokines (e.g., IL-6, TNF-α, IL-1β, RANTES) in response to infection, which attracts immune cells, such as dendritic cells and T cells to the site of infection. 2. Residing and infiltrating immune cells (DCs and T cells) are infection targets for HSV-2 and HIV. 3. DC infection with HSV-2 induces the secretion of soluble molecules at the site of infection that promote the reactivation of HIV from CD4+ T cells in HIV+ individuals with subsequent replication and dissemination. 4. DCs infected with HSV-2 recruit T cells that express α4β7, a molecule that facilitates infection of these cells by HIV. 5. DCs accumulating at the site of infection can be used by HIV as Trojan horses to reach and infect T cells in lymph nodes
On the other hand, it has been recently demonstrated that human dendritic cells infected with HSV-2 secrete cytokines that promote the reactivation of HIV from cells latently infected with this retrovirus (Fig. 2) [118]. Although this last observation does not directly relate to the probability of acquiring HIV, these results suggest that HIV+/HSV-2− individuals would display increased progression to AIDS after infection with HSV-2, as well as they would manifest increased probabilities of infecting their sexual partners with HIV.
Thanks to the implementation of a murine model that recapitulates the higher susceptibility to acquire HIV after HSV-2 infection, new molecular mechanisms that relate both viruses will likely be identified in the near future [124].
Another aspect relating HIV infection with increased transmission of HSV-2 is combination antiretroviral therapy (cART). Some studies have shown that suppressive antiretroviral therapy in HIV-positive patients can significantly increase genital ulcer disease (GUD) because of HSV-2 reactivation [125], although some have not observed this phenomenon [126]. Nevertheless, patients receiving cART therapy together with the antiviral acyclovir generally display significantly less GUD and HSV-2 shedding than those receiving cART alone, and thus, HIV+/HSV+ individuals are frequently prescribed with acyclovir [125, 127–129]. However, the chronic use of acyclovir in these patients has been shown to promote the emergence of HSV isolates that are resistant to this drug. Indeed, acyclovir-resistant HSV can be isolated in 3.5–10 % of immunocompromised patients receiving this antiviral [127–129] compared to <1 % of immunocompetent individuals [127, 129, 130]. HSV-resistance to acyclovir mainly results as a consequence of point mutations in the viral thymidine kinase (TK, UL23) and the viral DNA polymerase (DNA pol, UL30) (Table 1) [131–136].
Importantly, viral strains with point mutations in the viral TK or DNA pol genes display reduced neurovirulence in animal models, and loss of viral TK activity dramatically impairs virus reactivation from latency [137–139]. Because of the reduced virulence of acyclovir-resistant isolates, these viruses are detected nearly ten times less in healthy individuals than those that are immunocompromised [138, 140]. Nevertheless, viral strains presenting impaired TK activity can produce significant disease in neonates, such as encephalitis [141, 142].
A drug frequently used to combat acyclovir-resistant HSV strains in AIDS patients and bone marrow transplanted individuals is foscarnet (phosphonoformic acid, PFA) [143, 144]. This compound is a pyrophosphate analogue that inhibits the viral DNA polymerase by mimicking the structure of pyrophosphate recruited during the elongation of HSV DNA [145]. Although foscarnet-resistant HSV strains are rarely isolated (Table 1) [135, 146–148], unfortunately this drug displays some renal toxicity and requires administration under medical supervision. Other alternatives against acyclovir-resistant HSV isolates include cidofovir (which also affects the kidneys) and imiquimod, which is emerging as an attractive alternative to common antiviral drugs [149, 150].
Therapeutic and prophylactic antivirals and microbicides
Therapeutic and prophylactic antivirals
Antivirals such as acyclovir have been used since long to treat infections with HSV and other herpesviruses. The efficacy of this drug is significant when applied early after infection; however, it does not clear viral latency or alter the frequency of recurrences after acute treatment. Acyclovir is a guanosine nucleoside analog that once internalized into infected cells is processed into a monophosphorylated form that is subsequently phosphorylated into acyclovir triphosphate by host kinases [151]. In this form, acyclovir triphosphate inhibits viral replication by interfering with the activity of the viral polymerase [151, 152]. Because of its antiviral properties, acyclovir and its prodrug valacyclovir have also been assessed as therapeutic and prophylactic drugs for decreasing the transmission of HSV-2 onto sexual partners. Although valacyclovir has been shown to help decrease the transmission of HSV-2 from HSV-2+/HIV− individuals onto their couples [100, 101], acyclovir has been shown ineffective to do so for partners of HSV-2+/HIV+ co-infected individuals (Table 2) [153].
On the other hand, an antiviral that has acquired important attention, mainly because of its anti-HIV activity is tenofovir (TFV) and the oral pro-drug tenofovir disoproxil fumarate (TDF) [154]. Clinical studies in humans orientated to determinate the efficacy of TFV against HIV have demonstrated that this drug also possesses anti-HSV-2 effects, reducing the transmission of this virus up to 51 % [155]. Consistent with this observation, in vitro and in vivo studies in the murine model and macaques have demonstrated that TFV and TDF both possess antiviral capacity against HSV-2 (Table 2) [156–158]. However, one study determined that TDF would not affect the dissemination of HIV and HSV-2 in individuals co-infected with both viruses [159].
One of the major challenges of using antivirals is adherence. It is estimated that negative results recently obtained in a clinical study with TFV would mainly be due to this problem [160]. Thus, novel strategies seek to deliver these dugs in a continuous way using synthetic intravaginal rings (IVRs) that could be introduced into the vaginal cavity to release TDF in a steady fashion. To date, IVRs have been miniaturized with the aim to assess their efficacy in assays with vaginal tissue explants and double-chamber models with polarized epithelial cells, in which it has been observed that this strategy significantly reduces the infective capacity of HIV and HSV-2 [161]. More recently, IVRs that release TDF have also been tested, with promising results in non-human primates to prevent infection with HIV [157]. Effectively, macaques that received IVRs with TDF and were challenged with virus continuously for 16 weeks remained seronegative for SIVH (an HIV chimera for simians) (Table 2) [157]. It is expected that similar experiments will be performed in the short term to assess the protective capacity of these IVRs against HSV-2.
Microbicides/antivirals assessed in clinical settings
Because of the important association between HSV-2 and HIV, as well as the recurrence of genital lesions produced by infection with HSV, the identification of novel prophylactic drugs against these viruses is of high interest for public health systems worldwide. Although vaccines usually present better cost-effectiveness ratios than drugs, the absence of prophylactic alternatives against HSV-2 has promoted the search for new microbicides/antivirals that could help combat infection or clinical manifestations produced by this virus. A compound with anti-HSV-2 activity in vitro is the spermicide Nonoxynol-9 (N-9), which has been used for more than 30 year as a contraceptive (Table 2) [162]. N-9 also possess anti-HIV activity in vitro [163]. However, when assessed in clinical assays, it was found that repetitive applications of N-9 significantly increased the probability of acquiring vaginal infection with HIV [164] and papilloma virus (VPH) [165]. Furthermore, the application of N-9 in animal models has shown to significantly increase their susceptibility to infection with HSV-2 [166, 167]. Thus, the recommendation to use N-9 as a microbicide is being discontinued in individuals at risk of contracting sexually transmitted infections. These negative effects of N-9 would be due in part to the toxicity of this compound to the genital tissue, as evidenced in animals [168]. Furthermore, N-9 would interrupt the junctions between epithelial cells, characterized by the loss of adhesion and tight junction proteins, thus allowing the entry of viral particles to a broader range of target cells in the genital tract [167].
Another compound with microbicide potential is SPL7013, better known as VivaGel® (Table 2) [169]. This compound has been demonstrated to significantly inhibit the entry of HIV and HSV into human cells obtained from biopsies [170]. A clinical study evaluated the security, adherence and effect of VivaGel® on the microbiome of the vagina of women sexually active and found that, although this compound modified the microbiome, this did not translate into serious complications for the individuals [170]. Because of these favorable results, it is expected that this microbicide will soon be evaluated in more advanced clinical trials in humans. Other microbicides with anti-HSV-2 and anti-HIV activity in vitro include the surfactants SDS and C31G (Savvy), which act by interrupting the membrane and proteins of these viruses (Table 2) [171, 172]. Nevertheless, as for N-9, these compounds have demonstrated some toxicity in the vaginal tissue in animal models, as well as increased susceptibility to infection with HSV-2 [168]. Furthermore, in a phase I clinical trial, C31G produced adverse effects, such as vulvovaginitis, and thus, its use as a microbicide has been discontinued [173].
Cellulose sulfate [174] and PRO 2000 [175] are two molecules that emulate sulfate heparan and display an important capacity to block the entry of HSV-2 into target cells (Table 2). Although in vitro studies suggest that these molecules bind to the viral surface glycoprotein gB, which is essential for virus infection [176], clinical assays with these drugs demonstrated that PRO 2000 and cellulose sulfate would not confer significant protection against HSV-2 or HIV, and even associated the use of cellulose sulfate with an increase in contracting HIV (Table 2) [177, 178].
Noteworthy, studies aiming at identifying effective compounds against HSV have found that the seminal plasma can significantly reduce the effectiveness of microbicides, such as PRO 2000 and cellulose sulfate [179]. To obtain activities similar to those observed in the absence of seminal plasma, in some cases, it was necessary to increase the concentration of the drugs tested up to 100-fold [179]. Importantly, it has been observed that the seminal plasma can inhibit the binding of microbicides to the envelope of HSV-2 [179]. The main proteins identified to mediate this inhibition were fibronectin-1 and lactoferrin [179]. Thus, when assessing the effectiveness of microbicides, it will be important to consider the negative activity of seminal plasma over the activity of these compounds.
Microbicides/antivirals currently being evaluated in vitro and in vivo
Because microbicides have emerged as an attractive strategy to combat HSV, numerous compounds are currently being evaluated in vitro and in vivo for determining their anti-HSV properties. Two of these compounds are labyrinthopeptin A1 and PD 404,182 (Table 2). Labyrinthopeptin A1 is a carbacyclic lantibiotic with potent activity against HIV and HSV in vitro, capable of acting synergistically with acyclovir and tenofovir, within others [180]. PD 404,182, on the other hand, is capable of affecting the integrity of HIV and HSV through its interaction with viral protein components [181]. This latter compound has been demonstrated to be innocuous for the vaginal tissue and its microflora, as well as preserve its pH in the presence of seminal plasma.
Another microbicide with potential is the combination of zinc acetate and carrageenin (AZ/CG) (Table 2) [182]. AZ/CG decreases up to 80 % infection with HSV-2 in macaques, and its activity would be mediated by the binding of Zn to viral membrane glycoproteins impairing the entry of the virus into target cells [183]. Nevertheless, upon intravaginal challenge with HIV and HSV-2, the effectiveness of AZ/CG against HSV-2 was dramatically reduced up to 20 %, while protection against HIV was maintained [184].
Finally, some microbicides/antivirals derived from natural compounds with anti-HSV-2 activity have been characterized and tested in animal models, such as Meliacine and MI-S (Table 2). Meliacine is a derivative extracted from Melia azedarach and has demonstrated favorable results in mice against HSV-2 after topic application immediately after challenge with this virus [185]. MI-S, on the other hand, is a sulfated compound of a polysaccharide of the fungus Agaricus brasilensis, which has been observed to act over the binding of the virus and its entry into host cells; this compound reduced the clinical severity of the disease produced by HSV, as well as mortality in the murine model after one single application 20 min before infection (Table 2) [186]. Finally, the homo-dimeric lectin griffithsin extracted from the red algae Griffithsia sp., binds manose N-glycosilations in gp120 from HIV to block its binding to its receptor (Table 2) [187]. Additionally, griffithsin has been shown in the murine model of HSV-2 infection to reduce infection with this virus, as well as pathology by blocking cell-to-cell spread in the infected tissue [188]. These compounds should now progress into macaques and then humans.
The assessment of microbicides as an alternative against HSV-2 has experimented a significant increase in the field of prophylactics against this virus with some promising results that should translate into clinical studies. Noteworthy, the majority of the microbicides against HSV-2 assessed until now also share activity against HIV and vice versa. This finding suggests similar properties between both viruses as well as a somewhat lack of specificity for these types of antimicrobial agents. Important challenges for future microbicides/antivirals include granting constant administration of their active principles, reducing their toxic effects over the genital tissues and identifying early on detrimental effects, such as increasing infection with other pathogens. It is also important to consider the effectiveness of these compounds under variations of the hormonal cycle of individuals, age and presence of seminal plasma.
Concluding remarks
Although herpes simplex viruses were identified and isolated early in human history, to date, there is no treatment available to cure infection or vaccine to prevent their acquisition. The lack of successful treatment alternatives and vaccines against herpes simplex viruses has not only been troubled by effective molecular mechanisms evolved by these pathogens, but also likely by their somewhat unrecognized impact in public health until recently. Indeed, the currently identified relationship between HSV-2 and HIV calls for significant attention at controlling HSV-2 infection, which is highly prevalent worldwide. In this regard and in spite of the lack any vaccine against HSV, microbicides have emerged as promising alternatives that could reach the population fast enough to slow down HSV-2 spread. Recent favorable results in animal models should translate shortly into new clinical trials in humans that will seek to limit the impact of HSV-2 infection on the HIV epidemics.
References
Kolb AW, Ane C, Brandt CR (2013) Using HSV-1 genome phylogenetics to track past human migrations. PLoS ONE 8(10):e76267
Roizman B, Whitley RJ (2001) The nine ages of herpes simplex virus. Herpes 8(1):23–27
Ebers P (1937) Greatest Egyptian medical document, 1552 BC. Translated by Ebbell B. Levin & Munksgaard, Copenhagen
Astruc J (1736) De morbis venereis libri sex. G Cavelier, Paris
Unna P (1883) On herpes progenitalis, especially in women. J Cutan Vener Dis 1:321–334
Diday P, Doyon A (1886) Les herpes genitaux. Therapeutique des maladies veneriennes et des maladies cutanees. Paris: V Mason, Paris
Vidal J (1873) Ann Dermatol Syphiligr 1:350
Gruter W (1924) Das herpesvirus, seine atiologische und klinische bedeutung. Munch Med Wschr 71:1058
Smith G (2012) Herpesvirus transport to the nervous system and back again. Annu Rev Microbiol 66:153–176
Gupta R, Warren T, Wald A (2007) Genital herpes. Lancet 370(9605):2127–2137
Anderson BJ (2003) The epidemiology and clinical analysis of several outbreaks of herpes gladiatorum. Med Sci Sports Exerc 35(11):1809–1814
Bernstein DI, Bellamy AR, Hook EW 3rd, Levin MJ, Wald A, Ewell MG, Wolff PA, Deal CD, Heineman TC, Dubin G, Belshe RB (2013) Epidemiology, clinical presentation, and antibody response to primary infection with herpes simplex virus type 1 and type 2 in young women. Clin Infect Dis 56(3):344–351
Lowhagen GB, Tunback P, Bergstrom T (2002) Proportion of herpes simplex virus (HSV) type 1 and type 2 among genital and extragenital HSV isolates. Acta Derm Venereol 82(2):118–120
Tang SJ, McCrady W, Rawls D (2011) Acute herpes simplex gingivostomatitis and esophagitis (with video). Gastrointest Endosc 74(1):195–206 (discussion 196)
Usatine RP, Tinitigan R (2010) Nongenital herpes simplex virus. Am Fam Phys 82(9):1075–1082
Rowe AM, St Leger AJ, Jeon S, Dhaliwal DK, Knickelbein JE, Hendricks RL (2013) Herpes keratitis. Prog Retin Eye Res 32:88–101
Mathias RA, Weinberg A, Boguniewicz M, Zaccaro DJ, Armstrong B, Schneider LC, Hata TR, Hanifin JM, Beck LA, Barnes KC, Leung DY (2013) Atopic dermatitis complicated by eczema herpeticum is associated with HLA B7 and reduced interferon-gamma-producing CD8+ T cells. Br J Dermatol 169(3):700–703
Leung DY (2013) Why is eczema herpeticum unexpectedly rare? Antivir Res 98(2):153–157
Hoff NP, Gerber PA (2012) Herpetic whitlow. CMAJ 184(17):E924
Wu IB, Schwartz RA (2007) Herpetic whitlow. Cutis 79(3):193–196
Anderson BJ (2008) Managing herpes gladiatorum outbreaks in competitive wrestling: the 2007 Minnesota experience. Curr Sports Med Rep 7(6):323–327
Farooq AV, Shukla D (2012) Herpes simplex epithelial and stromal keratitis: an epidemiologic update. Surv Ophthalmol 57(5):448–462
Kaye S, Choudhary A (2006) Herpes simplex keratitis. Prog Retin Eye Res 25(4):355–380
Tiwari V, Shukla SY, Yue BY, Shukla D (2007) Herpes simplex virus type 2 entry into cultured human corneal fibroblasts is mediated by herpesvirus entry mediator. J Gen Virol 88(Pt 8):2106–2110
Karaba AH, Kopp SJ, Longnecker R (2012) Herpesvirus entry mediator is a serotype specific determinant of pathogenesis in ocular herpes. Proc Natl Acad Sci USA 109(50):20649–20654
van Gelderen BE, Van der Lelij A, Treffers WF, van der Gaag R (2000) Detection of herpes simplex virus type 1, 2 and varicella zoster virus DNA in recipient corneal buttons. Br J Ophthalmol 84(11):1238–1243
Kaneko H, Kawana T, Ishioka K, Ohno S, Aoki K, Suzutani T (2008) Evaluation of mixed infection cases with both herpes simplex virus types 1 and 2. J Med Virol 80(5):883–887
Remeijer L, Duan R, van Dun JM, Wefers Bettink MA, Osterhaus AD, Verjans GM (2009) Prevalence and clinical consequences of herpes simplex virus type 1 DNA in human cornea tissues. J Infect Dis 200(1):11–19
Itoh N, Matsumura N, Ogi A, Nishide T, Imai Y, Kanai H, Ohno S (2000) High prevalence of herpes simplex virus type 2 in acute retinal necrosis syndrome associated with herpes simplex virus in Japan. Am J Ophthalmol 129(3):404–405
Grose C (2012) Acute retinal necrosis caused by herpes simplex virus type 2 in children: reactivation of an undiagnosed latent neonatal herpes infection. Semin Pediatr Neurol 19(3):115–118
Van Gelder RN, Willig JL, Holland GN, Kaplan HJ (2001) Herpes simplex virus type 2 as a cause of acute retinal necrosis syndrome in young patients. Ophthalmology 108(5):869–876
Ward KN, Ohrling A, Bryant NJ, Bowley JS, Ross EM, Verity CM (2012) Herpes simplex serious neurological disease in young children: incidence and long-term outcome. Arch Dis Child 97(2):162–165
Steiner I, Benninger F (2013) Update on herpes virus infections of the nervous system. Curr Neurol Neurosci Rep 13(12):414
Riancho J, Delgado-Alvarado M, Sedano MJ, Polo JM, Berciano J (2013) Herpes simplex encephalitis: clinical presentation, neurological sequelae and new prognostic factors. Ten years of experience. Neurol Sci 34(10):1879–1881
Miller S, Mateen FJ, Aksamit AJ Jr (2013) Herpes simplex virus 2 meningitis: a retrospective cohort study. J Neurovirol 19(2):166–171
Dennett C, Cleator GM, Klapper PE (1997) HSV-1 and HSV-2 in herpes simplex encephalitis: a study of sixty-four cases in the United Kingdom. J Med Virol 53(1):1–3
Fica A, Perez C, Reyes P, Gallardo S, Calvo X, Salinas AM (2005) Herpetic encephalitis: case series of 15 patients confirmed by polymerase chain reaction. Rev Chilena Infectol 22(1):38–46
Shalabi M, Whitley RJ (2006) Recurrent benign lymphocytic meningitis. Clin Infect Dis 43(9):1194–1197
Poulikakos PJ, Sergi EE, Margaritis AS, Kioumourtzis AG, Kanellopoulos GD, Mallios PK, Dimitrakis DJ, Poulikakos DJ, Aspiotis AA, Deliousis AD, Flevaris CP, Zacharof AK (2010) A case of recurrent benign lymphocytic (Mollaret’s) meningitis and review of the literature. J Infect Public Health 3(4):192–195
Sato R, Ayabe M, Shoji H, Ichiyama T, Saito Y, Hondo R, Eizuru Y (2005) Herpes simplex virus type 2 recurrent meningitis (Mollaret’s meningitis): a consideration for the recurrent pathogenesis. J Infect 51(4):e217–e220
Farazmand P, Woolley PD, Kinghorn GR (2011) Mollaret’s meningitis and herpes simplex virus type 2 infections. Int J STD AIDS 22(6):306–307
Lee HY, Byun JY, Park MS, Yeo SG (2013) Steroid-antiviral treatment improves the recovery rate in patients with severe Bell’s palsy. Am J Med 126(4):336–341
Hato N, Murakami S, Gyo K (2008) Steroid and antiviral treatment for Bell’s palsy. Lancet 371(9627):1818–1820
Turriziani O, Falasca F, Maida P, Gaeta A, De Vito C, Mancini P, De Seta D, Covelli E, Attanasio G, Antonelli G (2014) Early collection of saliva specimens from Bell’s palsy patients: quantitative analysis of HHV-6, HSV-1, and VZV. J Med Viro l86(10):1752–1758
Pitkaranta A, Piiparinen H, Mannonen L, Vesaluoma M, Vaheri A (2000) Detection of human herpesvirus 6 and varicella-zoster virus in tear fluid of patients with Bell’s palsy by PCR. J Clin Microbiol 38(7):2753–2755
Riediger C, Sauer P, Matevossian E, Muller MW, Buchler P, Friess H (2009) Herpes simplex virus sepsis and acute liver failure. Clin Transplant 23(Suppl 21):37–41
Capretti MG, Marsico C, Lazzarotto T, Gabrielli L, Bagni A, De Angelis M, Rossini R, Faldella G (2013) Herpes simplex virus 1 infection: misleading findings in an infant with disseminated disease. New Microbiol 36(3):307–313
Chong S, Kim TS, Cho EY (2010) Herpes simplex virus pneumonia: high-resolution CT findings. Br J Radiol 83(991):585–589
Short WR (2009) Fatal herpes simplex virus type 2 pneumonia in a person with AIDS. AIDS Read 19(2):51–52, 60–61
Tachikawa R, Tomii K, Seo R, Nagata K, Otsuka K, Nakagawa A, Otsuka K, Hashimoto H, Watanabe K, Shimizu N (2013) Detection of herpes viruses by multiplex and real-time polymerase chain reaction in bronchoalveolar lavage fluid of patients with acute lung injury or acute respiratory distress syndrome. Respiration 87(4):279–286
Meyers JD, Flournoy N, Thomas ED (1980) Infection with herpes simplex virus and cell-mediated immunity after marrow transplant. J Infect Dis 142(3):338–346
Graham BS, Snell JD Jr (1983) Herpes simplex virus infection of the adult lower respiratory tract. Medicine (Baltimore) 62(6):384–393
Kusne S, Schwartz M, Breinig MK, Dummer JS, Lee RE, Selby R, Starzl TE, Simmons RL, Ho M (1991) Herpes simplex virus hepatitis after solid organ transplantation in adults. J Infect Dis 163(5):1001–1007
Looker KJ, Garnett GP, Schmid GP (2008) An estimate of the global prevalence and incidence of herpes simplex virus type 2 infection. Bull World Health Organ 86(10):805–812
Paz-Bailey G, Ramaswamy M, Hawkes SJ, Geretti AM (2007) Herpes simplex virus type 2: epidemiology and management options in developing countries. Sex Transm Infect 83(1):16–22
Smith JS, Robinson NJ (2002) Age-specific prevalence of infection with herpes simplex virus types 2 and 1: a global review. J Infect Dis 186(Suppl 1):S3–S28
Centers for Disease, C and Prevention (2010) Seroprevalence of herpes simplex virus type 2 among persons aged 14–49 years—United States, 2005–2008. MMWR Morb Mortal Wkly Rep 59(15):456–459
Bradley H, Markowitz LE, Gibson T, McQuillan GM (2014) Seroprevalence of herpes simplex virus types 1 and 2—United States, 1999–2010. J Infect Dis 209(3):325–333
Kucera P, Gerber S, Marques-Vidal P, Meylan PR (2012) Seroepidemiology of herpes simplex virus type 1 and 2 in pregnant women in Switzerland: an obstetric clinic based study. Eur J Obstet Gynecol Reprod Biol 160(1):13–17
Doi Y, Ninomiya T, Hata J, Yonemoto K, Tanizaki Y, Arima H, Liu Y, Rahman M, Iida M, Kiyohara Y (2009) Seroprevalence of herpes simplex virus 1 and 2 in a population-based cohort in Japan. J Epidemiol 19(2):56–62
Carcamo CP, Campos PE, Garcia PJ, Hughes JP, Garnett GP, Holmes KK, Peru Pst (2012) Prevalences of sexually transmitted infections in young adults and female sex workers in Peru: a national population-based survey. Lancet Infect Dis 12(10):765–773
Clemens SA, Farhat CK (2010) Seroprevalence of herpes simplex 1-2 antibodies in Brazil. Rev Saude Publica 44(4):726–734
Cowan FM, French RS, Mayaud P, Gopal R, Robinson NJ, de Oliveira SA, Faillace T, Uuskula A, Nygard-Kibur M, Ramalingam S, Sridharan G, El Aouad R, Alami K, Rbai M, Sunil-Chandra NP, Brown DW (2003) Seroepidemiological study of herpes simplex virus types 1 and 2 in Brazil, Estonia, India, Morocco, and Sri Lanka. Sex Transm Infect 79(4):286–290
Schiffer JT, Corey L (2013) Rapid host immune response and viral dynamics in herpes simplex virus-2 infection. Nat Med 19(3):280–290
Dickson N, Righarts A, van Roode T, Paul C, Taylor J, Cunningham AL (2014) HSV-2 incidence by sex over four age periods to age 38 in a birth cohort. Sex Transm Infect 90(3):243–245
Roberts CM, Pfister JR, Spear SJ (2003) Increasing proportion of herpes simplex virus type 1 as a cause of genital herpes infection in college students. Sex Transm Dis 30(10):797–800
Buxbaum S, Geers M, Gross G, Schofer H, Rabenau HF, Doerr HW (2003) Epidemiology of herpes simplex virus types 1 and 2 in Germany: what has changed? Med Microbiol Immunol 192(3):177–181
Coyle PV, O’Neill HJ, Wyatt DE, McCaughey C, Quah S, McBride MO (2003) Emergence of herpes simplex type 1 as the main cause of recurrent genital ulcerative disease in women in Northern Ireland. J Clin Virol 27(1):22–29
Pereira VS, Moizeis RN, Fernandes TA, Araujo JM, Meissner RV, Fernandes JV (2012) Herpes simplex virus type 1 is the main cause of genital herpes in women of Natal, Brazil. Eur J Obstet Gynecol Reprod Biol 161(2):190–193
Xu F, Sternberg MR, Kottiri BJ, McQuillan GM, Lee FK, Nahmias AJ, Berman SM, Markowitz LE (2006) Trends in herpes simplex virus type 1 and type 2 seroprevalence in the United States. JAMA 296(8):964–973
Horowitz R, Aierstuck S, Williams EA, Melby B (2011) Herpes simplex virus infection in a university health population: clinical manifestations, epidemiology, and implications. J Am Coll Health 59(2):69–74
Samra Z, Scherf E, Dan M (2003) Herpes simplex virus type 1 is the prevailing cause of genital herpes in the Tel Aviv area, Israel. Sex Transm Dis 30(10):794–796
Manavi K, McMillan A, Ogilvie M (2004) Herpes simplex virus type 1 remains the principal cause of initial anogenital herpes in Edinburgh, Scotland. Sex Transm Dis 31(5):322–324
Kortekangas-Savolainen O, Orhanen E, Puodinketo T, Vuorinen T (2014) Epidemiology of genital herpes simplex virus type 1 and 2 infections in southwestern Finland during a 10-year period (2003–2012). Sex Transm Dis 41(4):268–271
Nieuwenhuis RF, van Doornum GJ, Mulder PG, Neumann HA, van der Meijden WI (2006) Importance of herpes simplex virus type-1 (HSV-1) in primary genital herpes. Acta Derm Venereol 86(2):129–134
Ryder N, Jin F, McNulty AM, Grulich AE, Donovan B (2009) Increasing role of herpes simplex virus type 1 in first-episode anogenital herpes in heterosexual women and younger men who have sex with men, 1992–2006. Sex Transm Infect 85(6):416–419
Halpern-Felsher BL, Cornell JL, Kropp RY, Tschann JM (2005) Oral versus vaginal sex among adolescents: perceptions, attitudes, and behavior. Pediatrics 115(4):845–851
Linehan MM, Richman S, Krummenacher C, Eisenberg RJ, Cohen GH, Iwasaki A (2004) In vivo role of nectin-1 in entry of herpes simplex virus type 1 (HSV-1) and HSV-2 through the vaginal mucosa. J Virol 78(5):2530–2536
Krummenacher C, Baribaud F, Ponce de Leon M, Baribaud I, Whitbeck JC, Xu R, Cohen GH, Eisenberg RJ (2004) Comparative usage of herpesvirus entry mediator A and nectin-1 by laboratory strains and clinical isolates of herpes simplex virus. Virology 322(2):286–299
Cocchi F, Menotti L, Mirandola P, Lopez M, Campadelli-Fiume G (1998) The ectodomain of a novel member of the immunoglobulin subfamily related to the poliovirus receptor has the attributes of a bona fide receptor for herpes simplex virus types 1 and 2 in human cells. J Virol 72(12):9992–10002
Lafferty WE, Coombs RW, Benedetti J, Critchlow C, Corey L (1987) Recurrences after oral and genital herpes simplex virus infection. Influence of site of infection and viral type. N Engl J Med 316(23):1444–1449
Solomon L, Cannon MJ, Reyes M, Graber JM, Wetherall NT, Reeves WC, R Task Force on Herpes Simplex Virus (2003) Epidemiology of recurrent genital herpes simplex virus types 1 and 2. Sex Transm Infect 79(6):456–459
Janier M, Scieux C, Meouchi R, Tournoux C, Porcher R, Maillard A, Fouere S, Taquin Y, Lassau F, Morel P (2006) Virological, serological and epidemiological study of 255 consecutive cases of genital herpes in a sexually transmitted disease clinic of Paris (France): a prospective study. Int J STD AIDS 17(1):44–49
Benedetti J, Corey L, Ashley R (1994) Recurrence rates in genital herpes after symptomatic first-episode infection. Ann Intern Med 121(11):847–854
Dickson N, Righarts A, van Roode T, Paul C, Taylor J, Cunningham AL (2013) HSV-2 incidence by sex over four age periods to age 38 in a birth cohort. Sex Transm Infect 90(3):243-245
Dinh TH, Dunne EF, Markowitz LE, Weinstock H, Berman S (2008) Assessing neonatal herpes reporting in the United States, 2000–2005. Sex Transm Dis 35(1):19–21
Xu F, Markowitz LE, Gottlieb SL, Berman SM (2007) Seroprevalence of herpes simplex virus types 1 and 2 in pregnant women in the United States. Am J Obstet Gynecol 196(1):43e1–43e6
Whitley R, Davis EA, Suppapanya N (2007) Incidence of neonatal herpes simplex virus infections in a managed-care population. Sex Transm Dis 34(9):704–708
Handel S, Klingler EJ, Washburn K, Blank S, Schillinger JA (2011) Population-based surveillance for neonatal herpes in New York City, April 2006–September 2010. Sex Transm Dis 38(8):705–711
Sauerbrei A, Schmitt S, Scheper T, Brandstadt A, Saschenbrecker S, Motz M, Soutschek E, Wutzler P (2011) Seroprevalence of herpes simplex virus type 1 and type 2 in Thuringia, Germany, 1999 to 2006. Euro Surveill 16:44
Kasubi MJ, Nilsen A, Marsden HS, Bergstrom T, Langeland N, Haarr L (2006) Prevalence of antibodies against herpes simplex virus types 1 and 2 in children and young people in an urban region in Tanzania. J Clin Microbiol 44(8):2801–2807
Marquez L, Levy ML, Munoz FM, Palazzi DL (2011) A report of three cases and review of intrauterine herpes simplex virus infection. Pediatr Infect Dis J 30(2):153–157
Brown ZA, Vontver LA, Benedetti J, Critchlow CW, Hickok DE, Sells CJ, Berry S, Corey L (1985) Genital herpes in pregnancy: risk factors associated with recurrences and asymptomatic viral shedding. Am J Obstet Gynecol 153(1):24–30
Brown ZA, Wald A, Morrow RA, Selke S, Zeh J, Corey L (2003) Effect of serologic status and cesarean delivery on transmission rates of herpes simplex virus from mother to infant. JAMA 289(2):203–209
Robinson JL, Vaudry WL, Forgie SE, Lee BE (2012) Prevention, recognition and management of neonatal HSV infections. Expert Rev Anti Infect Ther 10(6):675–685
Wald A, Zeh J, Selke S, Ashley RL, Corey L (1995) Virologic characteristics of subclinical and symptomatic genital herpes infections. N Engl J Med 333(12):770–775
Kimberlin DW, Baley J, D Committee on Infectious, F Committee on, and Newborn (2013) Guidance on management of asymptomatic neonates born to women with active genital herpes lesions. Pediatrics 131(2):383–386
Wald A, Krantz E, Selke S, Lairson E, Morrow RA, Zeh J (2006) Knowledge of partners’ genital herpes protects against herpes simplex virus type 2 acquisition. J Infect Dis 194(1):42–52
Gardella C, Brown Z, Wald A, Selke S, Zeh J, Morrow RA, Corey L (2005) Risk factors for herpes simplex virus transmission to pregnant women: a couples study. Am J Obstet Gynecol 193(6):1891–1899
Corey L, Wald A, Patel R, Sacks SL, Tyring SK, Warren T, Douglas JM Jr, Paavonen J, Morrow RA, Beutner KR, Stratchounsky LS, Mertz G, Keene ON, Watson HA, Tait D, Vargas-Cortes M, Valacyclovir HSVTSG (2004) Once-daily valacyclovir to reduce the risk of transmission of genital herpes. N Engl J Med 350(1):11–20
Patel R (2004) Antiviral agents for the prevention of the sexual transmission of herpes simplex in discordant couples. Curr Opin Infect Dis 17(1):45–48
Norris Turner A, Paul P, Norris AH (2013) Limited benefit of repeating a sensitive question in a cross-sectional sexual health study. BMC Med Res Methodol 13:34
Beydoun HA, Dail J, Ugwu B, Boueiz A, Beydoun MA (2010) Socio-demographic and behavioral correlates of herpes simplex virus type 1 and 2 infections and co-infections among adults in the USA. Int J Infect Dis 14(Suppl 3):e154–e160
Shust GF, Cho S, Kim M, Madan RP, Guzman EM, Pollack M, Epstein J, Cohen HW, Keller MJ, Herold BC (2010) Female genital tract secretions inhibit herpes simplex virus infection: correlation with soluble mucosal immune mediators and impact of hormonal contraception. Am J Reprod Immunol 63(2):110–119
Smith JS, Herrero R, Munoz N, Eluf-Neto J, Ngelangel C, Bosch FX, Ashley RL (2001) Prevalence and risk factors for herpes simplex virus type 2 infection among middle-age women in Brazil and the Philippines. Sex Transm Dis 28(4):187–194
Cherpes TL, Melan MA, Kant JA, Cosentino LA, Meyn LA, Hillier SL (2005) Genital tract shedding of herpes simplex virus type 2 in women: effects of hormonal contraception, bacterial vaginosis, and vaginal group B Streptococcus colonization. Clin Infect Dis 40(10):1422–1428
Hazrati E, Galen B, Lu W, Wang W, Ouyang Y, Keller MJ, Lehrer RI, Herold BC (2006) Human alpha- and beta-defensins block multiple steps in herpes simplex virus infection. J Immunol 177(12):8658–8666
John M, Keller MJ, Fam EH, Cheshenko N, Hogarty K, Kasowitz A, Wallenstein S, Carlucci MJ, Tuyama AC, Lu W, Klotman ME, Lehrer RI, Herold BC (2005) Cervicovaginal secretions contribute to innate resistance to herpes simplex virus infection. J Infect Dis 192(10):1731–1740
Madan RP, Carpenter C, Fiedler T, Kalyoussef S, McAndrew TC, Viswanathan S, Kim M, Keller MJ, Fredricks DN, Herold BC (2012) Altered biomarkers of mucosal immunity and reduced vaginal Lactobacillus concentrations in sexually active female adolescents. PLoS ONE 7(7):e40415
Wasserheit JN (1992) Epidemiological synergy. Interrelationships between human immunodeficiency virus infection and other sexually transmitted diseases. Sex Transm Dis 19(2):61–77
Barnabas RV, Wasserheit JN, Huang Y, Janes H, Morrow R, Fuchs J, Mark KE, Casapia M, Mehrotra DV, Buchbinder SP, Corey L, Network NHVT (2011) Impact of herpes simplex virus type 2 on HIV-1 acquisition and progression in an HIV vaccine trial (the Step study). J Acquir Immune Defic Syndr 57(3):238–244
Freeman EE, Weiss HA, Glynn JR, Cross PL, Whitworth JA, Hayes RJ (2006) Herpes simplex virus 2 infection increases HIV acquisition in men and women: systematic review and meta-analysis of longitudinal studies. AIDS 20(1):73–83
Corey L, Wald A, Celum CL, Quinn TC (2004) The effects of herpes simplex virus-2 on HIV-1 acquisition and transmission: a review of two overlapping epidemics. J Acquir Immune Defic Syndr 35(5):435–445
Bagdades EK, Pillay D, Squire SB, O’Neil C, Johnson MA, Griffiths PD (1992) Relationship between herpes simplex virus ulceration and CD4+ cell counts in patients with HIV infection. AIDS 6(11):1317–1320
Cowan FF, Pascoe SJ, Barlow KL, Langhaug LF, Jaffar S, Hargrove JW, Robinson NJ, Latif AS, Bassett MT, Wilson D, Brown DW, Hayes RJ (2006) Association of genital shedding of herpes simplex virus type 2 and HIV-1 among sex workers in rural Zimbabwe. AIDS 20(2):261–267
Mayaud, P, N Nagot, I Konate, A Ouedraogo, HA Weiss, V Foulongne, MC Defer, A Sawadogo, M Segondy, P Van de Perre, and AS Group (2008) Effect of HIV-1 and antiretroviral therapy on herpes simplex virus type 2: a prospective study in African women. Sex Transm Infect 84(5):332–337
Duluc D, Gannevat J, Anguiano E, Zurawski S, Carley M, Boreham M, Stecher J, Dullaers M, Banchereau J, Oh S (2013) Functional diversity of human vaginal APC subsets in directing T-cell responses. Mucosal Immunol 6(3):626–638
Stefanidou M, Ramos I, Mas Casullo V, Trepanier JB, Rosenbaum S, Fernandez-Sesma A, Herold BC (2013) Herpes simplex virus 2 (HSV-2) prevents dendritic cell maturation, induces apoptosis, and triggers release of proinflammatory cytokines: potential links to HSV-HIV synergy. J Virol 87(3):1443–1453
Martinelli E, Tharinger H, Frank I, Arthos J, Piatak M Jr, Lifson JD, Blanchard J, Gettie A, Robbiani M (2011) HSV-2 infection of dendritic cells amplifies a highly susceptible HIV-1 cell target. PLoS Pathog 7(6):e1002109
Johnson KE, Redd AD, Quinn TC, Collinson-Streng AN, Cornish T, Kong X, Sharma R, Tobian AA, Tsai B, Sherman ME, Kigozi G, Serwadda D, Wawer MJ, Gray RH (2011) Effects of HIV-1 and herpes simplex virus type 2 infection on lymphocyte and dendritic cell density in adult foreskins from Rakai. Uganda. J Infect Dis 203(5):602–609
Rebbapragada A, Wachihi C, Pettengell C, Sunderji S, Huibner S, Jaoko W, Ball B, Fowke K, Mazzulli T, Plummer FA, Kaul R (2007) Negative mucosal synergy between Herpes simplex type 2 and HIV in the female genital tract. AIDS 21(5):589–598
Sartori E, Calistri A, Salata C, Del Vecchio C, Palu G, Parolin C (2011) Herpes simplex virus type 2 infection increases human immunodeficiency virus type 1 entry into human primary macrophages. Virol J 8:166
de Jong MA, de Witte L, Taylor ME, Geijtenbeek TB (2010) Herpes simplex virus type 2 enhances HIV-1 susceptibility by affecting Langerhans cell function. J Immunol 185(3):1633–1641
Nixon B, Fakioglu E, Stefanidou M, Wang Y, Dutta M, Goldstein H, Herold BC (2013) Genital herpes simplex virus type 2 infection in humanized HIV-transgenic mice triggers HIV shedding and is associated with greater neurological disease. J Infect Dis 209(4):510–522
Tobian AA, Grabowski MK, Serwadda D, Newell K, Ssebbowa P, Franco V, Nalugoda F, Wawer MJ, Gray RH, Quinn TC, Reynolds SJ, Rakai Health Sciences P (2013) Reactivation of herpes simplex virus type 2 after initiation of antiretroviral therapy. J Infect Dis 208(5):839–846
Tan DH, Raboud JM, Kaul R, Walmsley SL (2014) Antiretroviral therapy is not associated with reduced herpes simplex virus shedding in HIV coinfected adults: an observational cohort study. BMJ Open 4(1):e004210
Ziyaeyan M, Alborzi A, Japoni A, Kadivar M, Davarpanah MA, Pourabbas B, Abassian A (2007) Frequency of acyclovir-resistant herpes simplex viruses isolated from the general immunocompetent population and patients with acquired immunodeficiency syndrome. Int J Dermatol 46(12):1263–1266
Lolis MS, Gonzalez L, Cohen PJ, Schwartz RA (2008) Drug-resistant herpes simplex virus in HIV infected patients. Acta Dermatovenerol Croat 16(4):204–208
Stranska R, Schuurman R, Nienhuis E, Goedegebuure IW, Polman M, Weel JF, Wertheim-Van Dillen PM, Berkhout RJ, van Loon AM (2005) Survey of acyclovir-resistant herpes simplex virus in the Netherlands: prevalence and characterization. J Clin Virol 32(1):7–18
Bacon TH, Boon RJ, Schultz M, Hodges-Savola C (2002) Surveillance for antiviral-agent-resistant herpes simplex virus in the general population with recurrent herpes labialis. Antimicrob Agents Chemother 46(9):3042–3044
Chibo D, Druce J, Sasadeusz J, Birch C (2004) Molecular analysis of clinical isolates of acyclovir resistant herpes simplex virus. Antivir Res 61(2):83–91
Sauerbrei A, Deinhardt S, Zell R, Wutzler P (2010) Phenotypic and genotypic characterization of acyclovir-resistant clinical isolates of herpes simplex virus. Antivir Res 86(3):246–252
Burrel S, Deback C, Agut H, Boutolleau D (2010) Genotypic characterization of UL23 thymidine kinase and UL30 DNA polymerase of clinical isolates of herpes simplex virus: natural polymorphism and mutations associated with resistance to antivirals. Antimicrob Agents Chemother 54(11):4833–4842
Burrel S, Aime C, Hermet L, Ait-Arkoub Z, Agut H, Boutolleau D (2013) Surveillance of herpes simplex virus resistance to antivirals: a 4-year survey. Antivir Res 100(2):365–372
Schmit I, Boivin G (1999) Characterization of the DNA polymerase and thymidine kinase genes of herpes simplex virus isolates from AIDS patients in whom acyclovirand foscarnet therapy sequentially failed. J Infect Dis 180(2):487–490
Burrel S, Bonnafous P, Hubacek P, Agut H, Boutolleau D (2012) Impact of novel mutations of herpes simplex virus 1 and 2 thymidine kinases on acyclovir phosphorylation activity. Antivir Res 96(3):386–390
Chen SH, Pearson A, Coen DM, Chen SH (2004) Failure of thymidine kinase-negative herpes simplex virus to reactivate from latency following efficient establishment. J Virol 78(1):520–523
Levin MJ, Bacon TH, Leary JJ (2004) Resistance of herpes simplex virus infections to nucleoside analogues in HIV-infected patients. Clin Infect Dis 39(Suppl 5):S248–S257
Harris W, Collins P, Fenton RJ, Snowden W, Sowa M, Darby G (2003) Phenotypic and genotypic characterization of clinical isolates of herpes simplex virus resistant to aciclovir. J Gen Virol 84(Pt 6):1393–1401
Hill EL, Hunter GA, Ellis MN (1991) In vitro and in vivo characterization of herpes simplex virus clinical isolates recovered from patients infected with human immunodeficiency virus. Antimicrob Agents Chemother 35(11):2322–2328
Kakiuchi S, Nonoyama S, Wakamatsu H, Kogawa K, Wang L, Kinoshita-Yamaguchi H, Takayama-Ito M, Lim CK, Inoue N, Mizuguchi M, Igarashi T, Saijo M (2013) Neonatal herpes encephalitis caused by a virologically confirmed acyclovir-resistant herpes simplex virus 1 strain. J Clin Microbiol 51(1):356–359
Oram RJ, Marcellino D, Strauss D, Gustafson E, Talarico CL, Root AK, Sharma PL, Thompson K, Fingeroth JD, Crumpacker C, Herold BC (2000) Characterization of an acyclovir-resistant herpes simplex virus type 2 strain isolated from a premature neonate. J Infect Dis 181(4):1458–1461
Reusser P, Cordonnier C, Einsele H, Engelhard D, Link D, Locasciulli A, Ljungman P (1996) European survey of herpesvirus resistance to antiviral drugs in bone marrow transplant recipients. Infectious Diseases Working Party of the European Group for Blood and Marrow Transplantation (EBMT). Bone Marrow Transplant 17(5):813–817
Safrin S, Assaykeen T, Follansbee S, Mills J (1990) Foscarnet therapy for acyclovir-resistant mucocutaneous herpes simplex virus infection in 26 AIDS patients: preliminary data. J Infect Dis 161(6):1078–1084
Oberg B (1989) Antiviral effects of phosphonoformate (PFA, foscarnet sodium). Pharmacol Ther 40(2):213–285
Danve-Szatanek C, Aymard M, Thouvenot D, Morfin F, Agius G, Bertin I, Billaudel S, Chanzy B, Coste-Burel M, Finkielsztejn L, Fleury H, Hadou T, Henquell C, Lafeuille H, Lafon ME, Le Faou A, Legrand MC, Maille L, Mengelle C, Morand P, Morinet F, Nicand E, Omar S, Picard B, Pozzetto B, Puel J, Raoult D, Scieux C, Segondy M, Seigneurin JM, Teyssou R, Zandotti C (2004) Surveillance network for herpes simplex virus resistance to antiviral drugs: 3-year follow-up. J Clin Microbiol 42(1):242–249
Safrin S, Kemmerly S, Plotkin B, Smith T, Weissbach N, De Veranez D, Phan LD, Cohn D (1994) Foscarnet-resistant herpes simplex virus infection in patients with AIDS. J Infect Dis 169(1):193–196
Langston AA, Redei I, Caliendo AM, Somani J, Hutcherson D, Lonial S, Bucur S, Cherry J, Allen A, Waller EK (2002) Development of drug-resistant herpes simplex virus infection after haploidentical hematopoietic progenitor cell transplantation. Blood 99(3):1085–1088
Muluneh B, Dean A, Armistead P, Khan T (2013) Successful clearance of cutaneous acyclovir-resistant, foscarnet-refractory herpes virus lesions with topical cidofovir in an allogeneic hematopoietic stem cell transplant patient. J Oncol Pharm Pract 19(2):181–185
Hirokawa D, Woldow A, Lee SN, Samie F (2011) Treatment of recalcitrant herpes simplex virus with topical imiquimod. Cutis 88(6):276–277
Elion GB (1982) Mechanism of action and selectivity of acyclovir. Am J Med 73(1A):7–13
Elion GB (1983) The biochemistry and mechanism of action of acyclovir. J Antimicrob Chemother 12(Suppl B):9–17
Mujugira A, Magaret AS, Celum C, Baeten JM, Lingappa JR, Morrow RA, Fife KH, Delany-Moretlwe S, de Bruyn G, Bukusi EA, Karita E, Kapiga S, Corey L, Wald A, HSVHIVTST Partners in Prevention (2013) Daily acyclovir to decrease herpes simplex virus type 2 (HSV-2) transmission from HSV-2/HIV-1 coinfected persons: a randomized controlled trial. J Infect Dis 208(9):1366–1374
Keller MJ, Madan RP, Torres NM, Fazzari MJ, Cho S, Kalyoussef S, Shust G, Mesquita PM, Louissaint N, Chen J, Cohen HW, Diament EC, Lee AC, Soto-Torres L, Hendrix CW, Herold BC (2011) A randomized trial to assess anti-HIV activity in female genital tract secretions and soluble mucosal immunity following application of 1% tenofovir gel. PLoS ONE 6(1):e16475
Tan D (2012) Potential role of tenofovir vaginal gel for reduction of risk of herpes simplex virus in females. Int J Womens Health 4:341–350
Vibholm L, Reinert LS, Sogaard OS, Paludan SR, Ostergaard L, Tolstrup M, Melchjorsen J (2012) Antiviral and immunological effects of tenofovir microbicide in vaginal herpes simplex virus 2 infection. AIDS Res Hum Retrovir 28(11):1404–1411
Smith JM, Rastogi R, Teller RS, Srinivasan P, Mesquita PM, Nagaraja U, McNicholl JM, Hendry RM, Dinh CT, Martin A, Herold BC, Kiser PF (2013) Intravaginal ring eluting tenofovir disoproxil fumarate completely protects macaques from multiple vaginal simian-HIV challenges. Proc Natl Acad Sci USA 110(40):16145–16150
Nixon B, Jandl T, Teller R, Taneva E, Wang Y, Nagaraja U, Kiser P, Herold B (2013) Vaginally Delivered Tenofovir Disoproxil Fumarate Provides Greater Protection than Tenofovir against Genital Herpes in a Murine Model of Efficacy and Safety. Antimicrobial Agents Chemother 58(2):1153–1160
Tan DH, Kaul R, Raboud JM, Walmsley SL (2011) No impact of oral tenofovir disoproxil fumarate on herpes simplex virus shedding in HIV-infected adults. AIDS 25(2):207–210
Marrazzo JM, Ramjee G, Nair G, Palanee T, Mkhize B, Nakabito C, Taljaajrd M, Piper J, Gomez Feliciano K, Chirenje M, Team aVS (2013) 20th Conference on retroviruses and opportunistic infections, Atlanta, GA
Mesquita PM, Rastogi R, Segarra TJ, Teller RS, Torres NM, Huber AM, Kiser PF, Herold BC (2012) Intravaginal ring delivery of tenofovir disoproxil fumarate for prevention of HIV and herpes simplex virus infection. J Antimicrob Chemother 67(7):1730–1738
Jennings R, Clegg A (1993) The inhibitory effect of spermicidal agents on replication of HSV-2 and HIV-1 in-vitro. J Antimicrob Chemother 32(1):71–82
Polsky B, Baron PA, Gold JW, Smith JL, Jensen RH, Armstrong D (1988) In vitro inactivation of HIV-1 by contraceptive sponge containing nonoxynol-9. Lancet 1(8600):1456
Van Damme, L, G Ramjee, M Alary, B Vuylsteke, V Chandeying, H Rees, P Sirivongrangson, L Mukenge-Tshibaka, V Ettiègne-Traoré, C Uaheowitchai, S Karim, B Mâsse, J Perriëns, M Laga, and COLS Group (2002) Effectiveness of COL-1492, a nonoxynol-9 vaginal gel, on HIV-1 transmission in female sex workers: a randomised controlled trial. Lancet 360(9338):971–977
Marais D, Carrara H, Kay P, Ramjee G, Allan B, Williamson AL (2006) The impact of the use of COL-1492, a nonoxynol-9 vaginal gel, on the presence of cervical human papillomavirus in female sex workers. Virus Res 121(2):220–222
Galen BT, Martin AP, Hazrati E, Garin A, Guzman E, Wilson SS, Porter DD, Lira SA, Keller MJ, Herold BC (2007) A comprehensive murine model to evaluate topical vaginal microbicides: mucosal inflammation and susceptibility to genital herpes as surrogate markers of safety. J Infect Dis 195(9):1332–1339
Wilson SS, Cheshenko N, Fakioglu E, Mesquita PM, Keller MJ, Herold BC (2009) Susceptibility to genital herpes as a biomarker predictive of increased HIV risk: expansion of a murine model of microbicide safety. Antivir Ther 14(8):1113–1124
Cone R, Hoen T, Wong X, Abusuwwa R, Anderson D, Moench T (2006) Vaginal microbicides: detecting toxicities in vivo that paradoxically increase pathogen transmission. BMC Infect Dis 6:90
Price C, Tyssen D, Sonza S, Davie A, Evans S, Lewis G, Xia S, Spelman T, Hodsman P, Moench T, Humberstone A, Paull J, Tachedjian G (2011) SPL7013 Gel (VivaGel®) retains potent HIV-1 and HSV-2 inhibitory activity following vaginal administration in humans. PLoS ONE 6(9):e24095
McGowan I, Gomez K, Bruder K, Febo I, Chen B, Richardson B, Husnik M, Livant E, Price C, Jacobson C, MTNP Team (2011) Phase 1 randomized trial of the vaginal safety and acceptability of SPL7013 gel (VivaGel) in sexually active young women (MTN-004). AIDS (London, England) 25(8):1057–1064
Krebs FC, Miller SR, Catalone BJ, Welsh PA, Malamud D, Howett MK, Wigdahl B (2000) Sodium dodecyl sulfate and C31G as microbicidal alternatives to nonoxynol 9: comparative sensitivity of primary human vaginal keratinocytes. Antimicrob Agents Chemother 44:1954–1960
Howett MK, Neely EB, Christensen ND, Wigdahl B, Krebs FC, Malamud D, Patrick SD, Pickel MD, Welsh PA, Reed CA, Ward MG, Budgeon LR, Kreider JW (1999) A broad-spectrum microbicide with virucidal activity against sexually transmitted viruses. Antimicrob Agents Chemother 43(2):314–321
Peterson L, Nanda K, Opoku B, Ampofo W, Owusu-Amoako M, Boakye A, Rountree W, Troxler A, Dominik R, Roddy R, Dorflinger L (2007) SAVVY (C31G) gel for prevention of HIV infection in women: a Phase 3, double-blind, randomized, placebo-controlled trial in Ghana. PLoS ONE 2(12):e1312
Anderson RA, Feathergill KA, Diao XH, Cooper MD, Kirkpatrick R, Herold BC, Doncel GF, Chany CJ, Waller DP, Rencher WF, Zaneveld LJ (2002) Preclinical evaluation of sodium cellulose sulfate (Ushercell) as a contraceptive antimicrobial agent. J Androl 23(3):426–438
Keller MJ, Zerhouni-Layachi B, Cheshenko N, John M, Hogarty K, Kasowitz A, Goldberg CL, Wallenstein S, Profy AT, Klotman ME, Herold BC (2006) PRO 2000 gel inhibits HIV and herpes simplex virus infection following vaginal application: a double-blind placebo-controlled trial. J Infect Dis 193(1):27–35
Cheshenko N, Keller MJ, MasCasullo V, Jarvis GA, Cheng H, John M, Li JH, Hogarty K, Anderson RA, Waller DP, Zaneveld LJ, Profy AT, Klotman ME, Herold BC (2004) Candidate topical microbicides bind herpes simplex virus glycoprotein B and prevent viral entry and cell-to-cell spread. Antimicrob Agents Chemother 48(6):2025–2036
Van Damme, L, R Govinden, F Mirembe, F Guédou, S Solomon, M Becker, B Pradeep, A Krishnan, M Alary, B Pande, G Ramjee, J Deese, T Crucitti, D Taylor, and CSS Group (2008) Lack of effectiveness of cellulose sulfate gel for the prevention of vaginal HIV transmission. N Engl J Med 359(5):463–472
Keller M, Mesquita PM, Torres N, Cho S, Shust G, Madan R, Cohen H, Petrie J, Ford T, Soto-Torres L, Profy A, Herold B (2010) Postcoital bioavailability and antiviral activity of 0.5% PRO 2000 gel: implications for future microbicide clinical trials. PLoS ONE 5(1):e8781
Patel S, Hazrati E, Cheshenko N, Galen B, Yang H, Guzman E, Wang R, Herold BC, Keller MJ (2007) Seminal plasma reduces the effectiveness of topical polyanionic microbicides. J Infect Dis 196(9):1394–1402
Ferir G, Petrova MI, Andrei G, Huskens D, Hoorelbeke B, Snoeck R, Vanderleyden J, Balzarini J, Bartoschek S, Bronstrup M, Sussmuth RD, Schols D (2013) The lantibiotic peptide labyrinthopeptin A1 demonstrates broad anti-HIV and anti-HSV activity with potential for microbicidal applications. PLoS ONE 8(5):e64010
Chamoun AM, Chockalingam K, Bobardt M, Simeon R, Chang J, Gallay P, Chen Z (2012) PD 404,182 is a virocidal small molecule that disrupts hepatitis C virus and human immunodeficiency virus. Antimicrob Agents Chemother 56(2):672–681
Kenney J, Rodríguez A, Kizima L, Seidor S, Menon R, Jean-Pierre N, Pugach P, Levendosky K, Derby N, Gettie A, Blanchard J, Piatak M, Lifson J, Paglini G, Zydowsky T, Robbiani M, Fernández Romero J (2013) A modified zinc acetate gel, a potential nonantiretroviral microbicide, is safe and effective against simian-human immunodeficiency virus and herpes simplex virus 2 infection in vivo. Antimicrob Agents Chemother 57(8):4001–4009
Kenney J, Aravantinou M, Singer R, Hsu M, Rodriguez A, Kizima L, Abraham CJ, Menon R, Seidor S, Chudolij A, Gettie A, Blanchard J, Lifson JD, Piatak M Jr, Fernandez-Romero JA, Zydowsky TM, Robbiani M (2011) An antiretroviral/zinc combination gel provides 24 hours of complete protection against vaginal SHIV infection in macaques. PLoS ONE 6(1):e15835
Hsu M, Aravantinou M, Menon R, Seidor S, Goldman D, Kenney J, Derby N, Gettie A, Blanchard J, Piatak M Jr, Lifson JD, Fernandez-Romero JA, Zydowsky TM, Robbiani M (2013) A combination microbicide gel protects macaques against vaginal simian human immunodeficiency virus-reverse transcriptase infection, but only partially reduces herpes simplex virus-2 infection after a single high-dose cochallenge. AIDS Res Hum Retroviruses 30(2):174–183
Petrera E, Coto CE (2009) Therapeutic effect of meliacine, an antiviral derived from Melia azedarach L., in mice genital herpetic infection. Phytother Res 23(12):1771–1777
Cardozo FT, Larsen IV, Carballo EV, Jose G, Stern RA, Brummel RC, Camelini CM, Rossi MJ, Simoes CM, Brandt CR (2013) In vivo anti-herpes simplex virus activity of a sulfated derivative of Agaricus brasiliensis mycelial polysaccharide. Antimicrob Agents Chemother 57(6):2541–2549
Alexandre KB, Gray ES, Lambson BE, Moore PL, Choge IA, Mlisana K, Karim SS, McMahon J, O’Keefe B, Chikwamba R, Morris L (2010) Mannose-rich glycosylation patterns on HIV-1 subtype C gp120 and sensitivity to the lectins, Griffithsin, Cyanovirin-N and Scytovirin. Virology 402(1):187–196
Nixon B, Stefanidou M, Mesquita PM, Fakioglu E, Segarra T, Rohan L, Halford W, Palmer KE, Herold BC (2013) Griffithsin protects mice from genital herpes by preventing cell-to-cell spread. J Virol 87(11):6257–6269
Acknowledgments
We thank Angello Retamal for critical revision of this manuscript. Authors are supported by grants FONDECYT 1140011 (CONICYT), FONDECYT 110397 (to AMK), FONDECYT 1100926 (to AMK), Biomedical Research Consortium (3CTI21526-P4 to AMK) and the Millennium Institute on Immunology and Immunotherapy (P09/016-F, Iniciativa Científico Milenio). AMK is a Chaire De La Région Pays De La Loire “Chercheur Étranger D'excellence” and a CDD-DR INSERM.
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Suazo, P.A., Tognarelli, E.I., Kalergis, A.M. et al. Herpes simplex virus 2 infection: molecular association with HIV and novel microbicides to prevent disease. Med Microbiol Immunol 204, 161–176 (2015). https://doi.org/10.1007/s00430-014-0358-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00430-014-0358-x