
Abstract
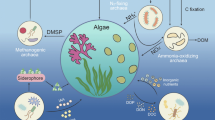
Antarctic “moss pillars” are lake-bottom biocenoses that are primarily comprised of aquatic mosses. The pillars consist of distinct redox-affected sections: oxidative exteriors and reductive interiors. Batteries of SSU rRNA genotypes of eukaryotes, eubacteria, and cyanobacteria, but no archaea, have been identified in these pillars. However, rRNA-based phylogenetic analysis provides limited information on metabolic capabilities. To investigate the microorganisms that have the potential for CO2 fixation in the pillars, we studied the genetic diversity of ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO, EC 4.1.1.39)—an enzyme involved in CO2 fixation. PCR clone libraries targeting all forms of the RuBisCO large subunit-encoding gene were constructed and 1,092 clones were randomly sequenced. Phylogenetic analysis indicated that proteobacterial form IA operational RuBisCO units (ORUs) were detected at the same frequency as the cyanobacterial form IB ORUs. Surprisingly, the form IA ORU, which was closely related to the sequences from deep-sea environments, was detected from all moss pillar sections. The form IB ORU related to Bryophyta, considered to be derived from moss, was identical to the sequence of Leptobryum sp. isolated from Lake Hotoke-Ike where the pillars were found. Moreover, certain cyanobacterial ORUs were found exclusively in the exterior of the pillar, whereas form II ORUs related to chemolithoautotrophic sulfur oxidizers and purple sulfur bacteria were found exclusively in the interior. No forms IC, ID, or III RuBisCO genes were detected. This is the first report demonstrating that bacteria with the potential for CO2 fixation and chemoautotrophy are present in the Antarctic moss pillar ecosystem.
Similar content being viewed by others


Introduction
Antarctic freshwater lake ecosystems are generally classified as oligotrophic or ultra-oligotrophic depending on the phytoplankton biomass in lake water (Padisák 2004; Lizotte 2008). These oligotrophic lakes are covered with ice for the majority of the year, leading to variation in the light environment depending on the ice thickness and snow cover (Vincent et al. 1998). Moreover, biologically harmful levels of solar radiation may penetrate through the ice cover and shallow water column (Vincent et al. 1998) because of low concentrations of UV-absorbing material (Morris et al. 1995). Benthic microbial communities in the lake produce carotenoids, xanthophyll, scytonemin, and mycosporine-like amino acids (Vincent et al. 1993a; Quesada et al. 1999; Hodgson et al. 2004; Tanabe et al. 2010), presumably to protect against intense light conditions (Kudoh et al. 2009). Despite this extreme environment, large pillar communities of aquatic mosses have been found in on the lake-bed near the Syowa Station located in East Antarctica (Imura et al. 1999). These pillars were distributed at depths greater than about 2 m (Imura et al. 1999). Analysis of lake environmental data obtained during a continuous year-long observation from January 2004 to February 2005 revealed that the chlorophyll a concentration, an indicator of primary production, was between 0.1 and 0.8 μg L−1 in the lake where moss pillars were observed (Tanabe et al. 2008), thereby classifying the environment as ultra-oligotrophic.
The exterior of the moss pillars is green with adherent oxygen bubbles resulting from photosynthesis, while the interior is packed with degraded moss tissues and gives off a strong odor of sulfur compounds (Imura et al. 2000). It is thought that an oxygen gradient exists between the exterior and interior (Kudoh et al. 2003a). Thus, the pillars have distinct redox-affected sections: oxidative exteriors and reductive interiors. Nakai et al. (2012a) reported high bacterial diversity and unique genotypes, or phylotypes, in moss pillars. Some phylotypes were distributed pillar-wide, while others were section-specific. Bacterial communities differ between the exterior and interior of the pillar and are hypothesized to participate in different biogeochemical processes. Moreover, 18S rRNA genetic diversity analysis revealed that a wide range of eukaryotic phylotypes, related to fungi, ciliates, tardigrades, rotifers, and nematodes, were present in the moss pillar ecosystem formed in ultra-oligotrophic lakes (Nakai et al. 2012b). However, SSU rRNA genes do not encode a physiological function that is important in biogeochemistry, and this sequence alone does not provide information on the metabolism of the cell. Thus, the biota involved in primary production and the formation and maintenance of this ecosystem remains unknown in ultra-oligotrophic lakes.
We investigated microorganisms with the potential for CO2 fixation in moss pillars by using genetic diversity analysis of ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO, E.C. 4.1.1.39)—an enzyme involved in CO2 fixation in the Calvin–Benson–Bassham (CBB) pathway. Since it constitutes approximately 30 % of the total protein in a plant leaf, RuBisCO is probably the most abundant protein (Chase 1993; Jensen 2000). The three known forms of RuBisCO consist of a hetero-16-unit polymer form I, a homodimer form II, and a homo-10-unit polymer form III (Badger and Bek 2008). Form I RuBisCO is generally divided into two types, green-like and red-like RuBisCOs. Green-like RuBisCO is classified into forms IA and IB, whereas red-like RuBisCO is classified into forms IC and ID based on homology of amino acid sequences, resulting in a total of four forms. Form IA RuBisCO is found primarily in proteobacteria, form IB in cyanobacteria and higher plants, form IC in proteobacteria, and form ID in heterokonts and haptophytes. Form II RuBisCO is found in proteobacteria and dinoflagellate algae. To date, form III RuBisCO has only been found in archaea (Berg et al. 2010).
The results of molecular phylogenetic analysis have been reported for a diverse range of RuBisCO genes in extreme environments, including the deep sea (Elsaied and Naganuma 2001; Elsaied et al. 2002, 2006, 2007), groundwater (Alfreider et al. 2009), caves (Chen et al. 2009), sediment from Lake Natron (Kovaleva et al. 2011), and salt lakes (Giri et al. 2004). Although the RuBisCO gene sequences from known isolated strains of Antarctic microorganisms have been reported (e.g., Liu et al. 2006), there is limited information on the genetic diversity in Antarctic moss pillars. Our data thus provide further insights into RuBisCO gene diversity among Antarctic microorganisms and the carbon cycle in Antarctic lakes.
Materials and methods
Study area
An Antarctic moss pillar specimen (diameter 22 cm; height 30 cm) was harvested on January 19, 2000, during the 42nd Japanese Antarctic Research Expedition at Lake Hotoke-Ike (69°28′S, 39°34′E), Skarvsnes, ~50 km south of Syowa Station in East Antarctica, as described in Nakai et al. (2012a). The water quality of the lake was described by Kudoh et al. (2003b).
DNA extraction
The moss pillar specimen was thawed partially and sectioned into 14 samples (seven exterior and seven interior samples) by separating the interior (blackish brown) from the exterior (green) layer and longitudinally dividing each specimen into seven horizontal sections. Exterior sections were labeled O1–O7 and interior sections were labeled I1–I7, as described previously (Nakai et al. 2012a). Sections were prepared by freeze-drying and then grinding with a sterilized mortar and pestle. Mixed microbial genomic DNA was extracted by applying a partially modified version of the bead-beating method reported by Miller et al. (1999) to 500 mg (dry weight) of each sample. Briefly, a sub-sample (100 mg dry weight) was placed in a 2-mL screw-cap microtube, with 1.2 g of sterilized 0.1-mm diameter zirconium/silica beads (BioSpec Products Inc., Bartlesville, OK, USA), 0.3 mL of phosphate buffer (100 mM NaH2PO4, pH 8.0), 0.3 mL of lysis buffer (10 % SDS, 100 mM NaCl, 500 mM Tris–HCl, pH 8.0), and 0.3 mL of chloroform-isoamyl alcohol (24:1). The microtube was set in a Mini Bead-Beater 8 (BioSpec Products) and shaken (3,200 rpm, 3 min). Beads and broken cell fragments in the tube were then removed by centrifugation (15,000 rpm, 5 min). The supernatant was purified using a Mag Extractor-Genome kit (Toyobo, Osaka, Japan) following the manufacturer’s instructions.
PCR amplification
A PCR clone library was generated from the RuBisCO gene in the purified genomic DNA. The cbbL gene of form I RuBisCO was amplified with primers IAB-595f and IAB-1385r, IC-537f and IC1212r, and ID-537f and ID-1212r (Table 1). The cbbM gene of form II RuBisCO was amplified with primers II-537f and II-1113r. The archaeal cbbL gene of form III RuBisCO was amplified with primers Arch-357f and Arch-891r. We used a previously reported primer specific to the form IB RuBisCO in Bryophyta (Tsubota et al. 1999) to obtain the near-full-length RuBisCO gene sequences derived from moss, the dominant component of moss pillars. The PCR conditions are described by Tsubota et al. (1999), Elsaied and Naganuma (2001), and Elsaid et al. (2007).
Construction of clone libraries and sequence analyses
The PCR-amplified products were cloned using a TOPO TA cloning kit (Invitrogen, Carlsbad, CA, USA) before transformation into Escherichia coli TOP10 (Invitrogen). The sequence of the inserted RuBisCO gene was determined using an ABI 3730XL automatic DNA sequencer (Applied Biosystems, Foster, USA). Similar sequences were grouped into operational RuBisCO units (ORUs) with >90 and 100 % identity for nucleotide and amino acid sequences, respectively, according to Elsaied and Naganuma (2001) and van der Wielen (2006). Inferred amino acid sequences for the ORUs were obtained using transeq (European Bioinformatics, http://www.ebi.ac.uk/emboss/transeq/). When the amino acid sequence contained the stop codon, it was excluded from further analysis. Nucleotide–nucleotide and protein–protein BLAST homology searches were conducted using the NCBI nt-database to compare the ORUs with known RuBisCOs (Altschul et al. 1997). Sequences that did not match with RuBisCO genes were excluded. We used CLUSTALX (Larkin et al. 2007) to perform alignments with ORU sequences and the closest sequences obtained from both nucleotide–nucleotide and protein–protein BLAST searches. The post-alignment sequences were used to create a phylogenetic tree by the neighbor-joining (NJ) method (Saitou and Nei 1987) using the MEGA 5.0 program (Tamura et al. 2011). The diversity of the ORUs was analyzed using the Rarefaction Calculator (http://www.biology.ualberta.ca/jbrzusto/rarefact.php). The ORU sequences were deposited in the DDBJ/EMBL/GenBank database under accession numbers AB695360 to AB695363 for bacterial form IA ORUs; AB695364 to AB695372 for bacterial form IB ORUs; AB695373 for a bryophytic form IB ORU; and AB695374 to AB695375 for bacterial form II ORUs.
Results and discussion
Distribution of RuBisCO genes obtained from the moss pillar
The form IA/IB RuBisCO gene was amplified using the primer set IAB-595f and IAB-1385r from all 14 sections (seven exterior and seven interior) of the studied moss pillar. The near-full-length form IB RuBisCO in Bryophyta was also amplified using the primer set rbcL7 and rbcL1346hR from all 14 sections. Conversely, form II was only amplified from I2 of the lower interior layer. No form IC, ID, and III RuBisCO genes were detected by PCR in any of the sections despite several PCR runs. In addition, the archaeal 16S rRNA gene was not detected in the microfloral analysis reported by Nakai et al. (2012a) for the moss pillar ecosystem. The number of archaea with the potential for CO2 fixation is low to non-existent, almost to the point of being undetectable by PCR.
A total of 29 PCR clone libraries were constructed for each section. We then randomly selected and sequenced 65 clones from each of the form IA/IB libraries, 9–10 clones from each of the bryophytic form IB libraries, and 45 clones from a form II library (total N = 1,092 clones). After determining the 1,092 clone sequences constructed from the PCR products, 884 clones were obtained for final analysis that excluded sequences containing the stop codon and sequences that did not hit with the RuBisCO gene. These 884 clones were grouped into 16 ORUs based on their ORU classification (matching >90 % of nucleotide sequences and 100 % of amino acid sequences) following to Elsaied and Naganuma (2001) and van der Wielen (2006). Four, ten, and two ORUs (out of 16) were assigned to form IA, IB, and II RuBisCO genes, respectively, by molecular phylogenetic analysis (Table 2; the distribution of each ORU in the 14 sections, results of the BLAST analysis for nucleotide identity, and amino acid identity are shown in Table S1).
There is a high probability that the 14 form I ORUs could function as CO2 fixation enzymes because they contained unique functional motifs (DFTKDDE) (Shinozaki et al. 1983; Newman and Gutteridge 1993) specific to form I RuBisCO. The previously reported functional motifs (GGDFIKNDE) (Schneider et al. 1990) were also found in the two form II ORUs obtained in this study. The rarefaction curves for bacterial form I RuBisCO gene obtained from sections O2, O3, I3, and I5 did not reach clear saturation, indicating that further sampling of these clone libraries would have revealed little additional diversity (Fig. 1).

Rarefaction curves for the expected number of ORUs for bacterial form I RuBisCO gene based on grouping of clones that have >90 and 100 % identity for nucleotide and amino acid sequences, respectively
Diversity of form IA RuBisCO genes from proteobacteria
We obtained 342 sequences belonging to the form IA RuBisCO gene from proteobacteria (approximately 49 % of the 702 total clones) among the clone libraries constructed with the primer set IAB-595f and IAB-1385r specific to the form IA/IB, and they were classified into four ORUs (Table 2). These were detected at roughly the same frequency as the form IB RuBisCO gene from cyanobacteria. ORU1 was most dominant (288/342 clones) of the four ORUs and was found in all 14 exterior and interior moss pillar sections (Table 2). ORU1 was most closely related to an environmental clone obtained from a tar oil-contaminated aquifer (EU926498), with 82 %ND (nucleotide identity) and 93 %AA (amino acid identity). The difference in identity between the nucleotide and amino acid sequence is influenced by the variability of the third base of the codon. A BLAST search for the RuBisCO genes of known isolates revealed that ORU1 was most closely related in nucleotide sequence to the RuBisCO gene of genus Thioalkalivibrio (CP001905) with 79 %ND, and in amino acid sequence to the RuBisCO gene of genus Bradyrhizobium (YP_001238690) with 89 %AA. ORU1 was also closely aligned on the phylogenetic tree with the genus Bradyrhizobium (Fig. 2). In addition, a previously reported bacterial 16S rRNA genetic diversity analysis (Nakai et al. 2012a) found phylotypes closely related to the genus Bradyrhizobium in all 14 exterior and interior sections. A member of the genus Bradyrhizobium can grow both heterotrophically, in the presence of oxygen, and anaerobically, as denitrifiers (van Berkum and Keyser 1985; Delgado et al. 2003). Moreover, some species fix carbon dioxide under microaerobic conditions using hydrogen as an electron donor (Hanus et al. 1979; Lepo et al. 1980), whereas others grow chemolithoautotrophically using thiosulfate as an electron donor (Masuda et al. 2010). The wide distribution of Bradyrhizobium-related sequences may reflected by the ability to grow under aerobic, microaerobic, and anaerobic conditions.

Phylogenetic tree of ORUs of the forms IA and IB RuBisCO amino acid sequences obtained from aquatic moss pillars. Evolutionary history was inferred by using the neighbor-joining method. The bootstrap consensus tree inferred from 1,000 replicates is taken to represent the evolutionary history of the taxa analyzed. Branches corresponding to partitions reproduced in <50 % bootstrap replicates were collapsed. The RuBisCO amino acid sequence of Olisthodiscus luteus (BAF80663) was used as outgroup. Accession numbers are showed in parentheses
Another surprising finding was that ORU2 (49/342 clones), found in all sections of the moss pillar, was closely related (81–85 %ND and 95 %AA) to the RuBisCO gene sequences from hydrothermal plumes and endosymbiotic bacteria from the deep-sea mussel Bathymodiolus azoricus. The phylogenetic tree indicated that it was closely aligned with a RuBisCO sequence found in endosymbiotic bacteria from deep-sea mussels (AAX48774) (Spiridonova et al. 2006) (Fig. 2). Deep-sea mussels are thought to rely on the primary production of chemoautotrophic bacteria that live symbiotically in the branchial cells of the mussels (McKiness and Cavanaugh 2005). There are three known types of symbiotic, chemoautotrophic bacteria: sulfur-oxidizing bacteria, methane-oxidizing bacteria, and both sulfur- and methane-oxidizing bacteria (McKiness and Cavanaugh 2005). A BLAST search for the RuBisCO genes of known isolates revealed that ORU2 found in this study was most closely related in nucleotide sequence to the sulfur-oxidizing bacteria genus Thiohalophilus RuBisCO gene (GQ888587) with 82 %ND, and in amino acid sequence to the sulfur-oxidizing bacteria genus Acidithiobacillus RuBisCO gene (P0C917) with 89 %AA. Hydrogen sulfide gas was undetected in the water column and at the surface of the phytobenthos in Lake Hotoke-Ike, but ≥1 μg g−1 (wet weight) of hydrogen sulfide gas was detected from deeper than 5 mm within the phytobenthos mat samples (Kudoh et al. 2009). The low concentration at the surface is assumed due to the rapid oxidation of hydrogen sulfide by sulfur-oxidizing bacteria. Thus, sulfur-oxidizing bacteria may contribute to primary production by synthesizing organic matter via the oxidation of hydrogen sulfide in the pillar.
The two remaining ORUs shared 83 %ND and 90 %AA, and 81 %ND and 89 %AA identity, respectively, with an environmental clone obtained from tropical muddy sediments (AY773062) and an environmental clone obtained from an aquifer (EU926498).
Our results suggest that proteobacteria are distributed throughout the moss pillar, where they may play a role in primary production via chemosynthesis in the moss pillar ecosystem by fixing carbon dioxide without relying on light energy. In addition, RuBisCO genes with extremely high identity were found at the deep-sea environments and in Antarctic lakes, two environments with widely differing conditions. Bacteria with this type of RuBisCO gene may be present in other Antarctic lakes since they are thought to be widely distributed.
Diversity in form IB RuBisCO genes from mosses and cyanobacteria
We obtained 137 sequences from the clone libraries that were constructed with the primer set rbcL7 and rbcL1346hR specific to the form IB RuBisCO gene in Bryophyta, and they were classified into a single ORU (Table 2). ORU14, considered to be the major component species of the pillar, shared 100 % nucleotide sequence identity with the RuBisCO gene sequence of Leptobryum sp. isolated from Lake Hotoke-Ike (AB690288) where the moss pillars were found. This agrees with the report by Imura et al. (1999) that Leptobryum sp. comprises the main structural component of moss pillars. However, the sequences related to Bryum, a minor structural species in moss pillars, were not obtained. Previous reports have shown that Leptobryum species are the primary mosses in moss pillars, whereas Bryum pseudotriquetrum is a minor or absent (Ohtani et al. 2001). Moreover, 18S rRNA gene-based diversity analysis (Nakai et al. 2012b) revealed clones closely related to Leptobryum 18S rRNA genes in all moss pillar sections, whereas Bryum-related clones were not found. Thus, Bryum sp. may have been absent in the moss pillar samples used in this study.
We obtained 360 sequences belonging to the form IB RuBisCO gene from cyanobacteria (approximately 51 % of total 702 clones) among the clone libraries constructed with the primer set IAB-595f and IAB-1385r specific to the form IA/IB, and they were classified into nine ORUs (Table 2). Excluding ORU10, these ORUs shared comparatively high identity in the range of 81–99 %ND and 91–98 %AA with the RuBisCO gene sequences of cyanobacteria such as Anabaena, Arthrospira, Fischerella, Leptolyngbya, Microcoleus, Nostoc, Raphidiopsis, Synechococcus, and Thermosynechococcus. Cyanobacteria form mats at the bottom of Antarctic lakes and they are thought to be the dominant primary producers on the lake-bottom (Vincent 1988, 2000; Quesada et al. 2008). ORU5 was the most dominant (209/360 clones) of these nine ORUs and was found in all 14 exterior and interior sections of the moss pillar (Table 2). ORU5 was highly homologous with the RuBisCO gene sequence of known species Leptolyngbya laminosa ETS-08, with 90 %ND and 97 %AA. ORU5 was closely aligned on the phylogenetic tree with an environmental clone obtained from an Antarctic lake, Lake Bonney, in the McMurdo Dry Valleys (ACY74728) (Fig. 2). A number of species of cyanobacteria, including Lyngbya, Oscillatoria, Nostoc, and Phormidium spp., were detected by using microscopy in samples of the lake-bed from Lake Bonney, the surface of which is covered with ice year-round (Vincent et al. 1993b; Wing and Priscu 1993; Paerl and Pinckney 1996). Our data demonstrate that RuBisCO genes from geographically distant Antarctic lakes, with Lake Bonney and Hotoke-Ike located approximately 3,000 km apart, are phylogenetically related. ORU13 was close on the phylogenetic tree to an environmental clone found in Lake Bonney (ACY74730), and it formed one independent cluster.
With 87 %ND and 96 %AA identity with the RuBisCO gene sequence for the genus Leptolyngbya, ORU11 was found exclusively in the exterior layer and it clustered with the widely distributed ORU5 on the phylogenetic tree. Thus, the distribution characteristics differed even among phylogenetically related ORUs. The phylotype related to the genus Leptolyngbya were also found exclusively in the exterior layer of moss pillars in a previous bacterial 16S rRNA genetic diversity analysis (Nakai et al. 2012a), indicating a high probability that some Leptolyngbya bacterial species are specific to the exterior layer of the pillar. ORU10 differed from the other eight ORUs as they shared homology with all RuBisCO gene sequences in the database. It was most closely related to the Nostoc RuBisCO gene with 84 %ND and 83 %AA. It was found only in the deepest part of the exterior layer (O1) of the moss pillar and formed an independent cluster on the phylogenetic tree (Fig. 2).
Diversity and phylogeny of form II RuBisCO genes from proteobacteria
We obtained 45 sequences from the clone libraries constructed with the primer set II-537f and II-1113r specific to the form II RuBisCO gene, and they were classified into two ORUs (Table 2). Form II was only detected from I2 of the lower interior layer. The first, ORU15 (35/45 clones), was most closely related in nucleotide sequence to an environmental clone from Movile Cave in Romania (FJ604806) with 92 %ND, and in amino acid sequence to an environmental clone from a tar oil-contaminated aquifer (ACH70392) with 93 %AA. A BLAST search of known RuBisCO gene isolates revealed that this ORU was most closely related to the RuBisCO gene sequence of the obligately chemolithoautotrophic, sulfur-oxidizing bacterium Halothiobacillus sp. RA13 (AY099399), with 90 %ND and 95 %AA. Salt is not essential for the growth of Halothiobacillus sp., although it is known to tolerate a high salt concentration (4 M NaCl and 0.25 M sodium thiosulfate) (Kelly and Wood 2000). The freshwater lakes inhabited by Leptobryum sp., the dominant component of moss pillars, have electronic conductivity in the range of 15.0–390.0 ms/mm (Imura et al. 2003). This highest value is close to one-tenth of the value in seawater.
The second, ORU16 (10/45 clones), was related to the RuBisCO gene sequence of the Chromatiales Thioalkalicoccus limnaeus B7-1 (AEN02453), with 86 %ND and 95 %AA. This result was supported by the phylogenetic tree (Fig. 3). T. limnaeus is an alkalophilic bacterium isolated from Lake Natron in southeast Siberia, Russia with a pH range of 9.5–10.1 (Bryantseva et al. 2000), which differs significantly from the 7–8-pH range of Lake Hotoke-Ike (Kimura et al. 2010). Purple sulfur bacteria, of the order Chromatiales, generally engage in photoautotrophic metabolism using reduced inorganic sulfur compounds (e.g., hydrogen sulfide) as electron donors. Thioalkalicoccus sp. contains bacteriochlorophyll b that functions intracellularly as a photosynthetic pigment (Bryantseva et al. 2000). Thus, this group can only grow in limited areas in anaerobic environments that receive light and contain a stable supply of hydrogen sulfide. The detection of the form II ORU related to Chromatiales may indicate that light is available to phototrophs even in the lower interior part of the pillar.

Phylogenetic tree of ORUs of the form II RuBisCO amino acid sequences obtained from aquatic moss pillars. Evolutionary history was inferred by using the neighbor-joining method. The bootstrap consensus tree inferred from 1,000 replicates is taken to represent the evolutionary history of the taxa analyzed. Branches corresponding to partitions reproduced in <50 % bootstrap replicates were collapsed. The RuBisCO amino acid sequence of Nostoc punctiforme PCC73102 (ACC82570) was used as outgroup. Accession numbers are showed in parentheses
Conclusion
We studied the distribution and diversity of all forms (forms IA, IB, IC, ID, II, and III) of the RuBisCO large subunit-encoding gene in a whole moss pillar. Proteobacterial form IA ORUs were present at the same frequency as cyanobacterial form IB ORUs. Surprisingly, the form IA ORU, which is closely related to the sequences from deep-sea environments such as hydrothermal plumes and endosymbiotic bacteria from deep-sea mussels, was detected in all moss pillar sections. The form IB ORU belonging to Bryophyta, considered to be derived from moss, was 100 % identical to the RuBisCO gene sequence from Leptobryum sp. In addition, some cyanobacterial ORUs were found exclusively in the exterior layer of the pillar, whereas form II ORUs related to obligately chemolithoautotrophic sulfur oxidizers, and purple sulfur bacteria were detected exclusively in the interior layer. This layer-specific distribution of ORUs reflects the double-layered structure of an oxidative exterior and a reductive interior layer in the pillars. This is the first report demonstrating that bacteria with the potential for CO2 fixation and chemoautotrophy are present in the Antarctic moss pillar ecosystem. CO2 fixation by chemoautotrophic bacteria is likely to be especially important during the polar night months in Antarctica. This could be verified by investigating the types and amounts of RuBisCO genes expressed in the field using a RNA-based PCR clone library analysis, or metatranscriptome and metaproteome analysis.
References
Alfreider A, Vogt C, Geiger-Kaiser M, Psenner R (2009) Distribution and diversity of autotrophic bacteria in groundwater systems based on the analysis of RuBisCO genotypes. Syst Appl Microbiol 32:140–150
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402
Badger MR, Bek EJ (2008) Multiple Rubisco forms in proteobacteria: their functional significance in relation to CO2 acquisition by the CBB cycle. J Exp Bot 59:1525–1541
Berg IA, Kockelkorn D, Ramos-Vera WH, Say RF, Zarzycki J, Hügler M, Alber BE, Fuchs G (2010) Autotrophic carbon fixation in archaea. Nat Rev Microbiol 8:447–460
Bryantseva IA, Gorlenko VM, Kompantseva EI, Imhoff JF (2000) Thioalkalicoccus limnaeus gen. nov., sp nov., a new alkaliphilic purple sulfur bacterium with bacteriochlorophyll b. Int J Syst Evol Microbiol 50:2157–2163
Chase MW (1993) Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rbcL. Ann Missouri Bot Gard 80:528–580
Chen Y, Wu LQ, Boden R, Hillebrand A, Kumaresan D et al (2009) Life without light: microbial diversity and evidence of sulfur- and ammonium-based chemolithotrophy in Movile Cave. ISME J 3:1093–1104
Delgado MJ, Bonnard N, Tresierra-Ayala A, Bedmar EJ, Muller P (2003) The Bradyrhizobium japonicum napEDABC genes encoding the periplasmic nitrate reductase are essential for nitrate respiration. Microbiology 149:3395–3403
Elsaied H, Naganuma T (2001) Phylogenetic diversity of ribulose-1,5-bisphosphate carboxylase/oxygenase large-subunit genes from deep-sea microorganisms. Appl Environ Microbiol 67:1751–1765
Elsaied H, Kimura H, Naganuma T (2002) Molecular characterization and endosymbiotic localization of the gene encoding ribulose 1,5-bisphosphate carboxylase-oxygenase (RuBisCO) form II in the deep-sea vestimentiferan trophosome. Microbiology 148:1947–1957
Elsaied HE, Kaneko R, Naganuma T (2006) Molecular characterization of a deep-sea methanotrophic mussel symbiont that carries a RuBisCO gene. Mar Biotech 8:511–520
Elsaied HE, Kimura H, Naganuma T (2007) Composition of archaeal, bacterial, and eukaryal RuBisCO genotypes in three Western Pacific arc hydrothermal vent systems. Extremophiles 11:191–202
Giri BJ, Bano N, Hollibaugh JT (2004) Distribution of RuBisCO genotypes along a redox gradient in Mono Lake, California. Appl Environ Microbiol 70:3443–3448
Hanus FJ, Maier RJ, Evans HJ (1979) Autotrophic growth of H2-uptake-positive strains of Rhizobium japonicum in an atmosphere supplied with hydrogen gas. Proc Natl Acad Sci USA 76:1788–1792
Hodgson DA, Vyverman W, Verleyen E, Sabbe K, Leavitt PR et al (2004) Environmental factors influencing the pigment composition of in situ benthic microbial communities in east Antarctic lakes. Aquat Microb Ecol 37:247–263
Imura S, Bando T, Saito S, Seto K, Kanda H (1999) Benthic moss pillars in Antarctic lakes. Polar Biol 22:137–140
Imura S, Takahashi H, Nakamura T (2000) Benthic moss pillars (Koke Bouzu) in Antarctic lakes—Analysis of colonization and growth by 14C dating. Sum Res using AMS, Nagoya Univ XI:176–183 (in Japanese with English Abstract)
Imura S, Bando S, Seto K, Ohtani S, Kudoh S, Kanda H (2003) Distribution of aquatic mosses in the Syowa Coast region, East Antarctica. Polar Biosci 16:1–10
Jensen RG (2000) Activation of Rubisco regulates photosynthesis at high temperature and CO2. Proc Natl Acad Sci USA 97:12937–12938
Kelly DP, Wood AP (2000) Reclassification of some species of Thiobacillus to the newly designated genera Acidithiobacillus gen. nov., Halothiobacillus gen. nov and Thermithiobacillus gen. nov. Int J Syst Evol Microbiol 50:511–516
Kimura S, Ban S, Imura S, Kudoh S, Matsuzaki M (2010) Limnological characteristics of vertical structure in the lakes of Syowa oasis, East Antarctica. Polar Sci 3:262–271
Kovaleva OL, Tourova TP, Muyzer G, Kolganova TV, Sorokin DY (2011) Diversity of RuBisCO and ATP citrate lyase genes in soda lake sediments. FEMS Microbiol Ecol 75:37–47
Kudoh S, Tuchiya Y, Ayukawa E, Imura S, Kanda H (2003a) Ecological studies of aquatic moss pillars in Antarctic lakes. 1: macro structure and carbon, nitrogen and chlorophyll a contents. Polar Biosci 16:11–22
Kudoh S, Watanabe K, Imura S (2003b) Ecological studies of aquatic moss pillars in Antarctic lakes. 2: temperature and light environment at the moss habitat. Polar Biosci 16:23–32
Kudoh S, Tanabe Y, Matsuzaki M, Imura S (2009) In situ photochemical activity of the phytobenthic communities in two Antarctic lakes. Polar Biol 32:1617–1627
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA et al (2007) Clustal W and clustal X version 2.0. Bioinformatics 23:2947–2948
Lepo JE, Hanus FJ, Evans HJ (1980) Chemoautotrophic growth of hydrogen-uptake-positive strains of Rhizobium japonicum. J Bacteriol 141:664–670
Liu CL, Huang XH, Wang XL, Zhang XC, Li GY (2006) Phylogenetic studies on two strains of Antarctic ice algae based on morphological and molecular characteristics. Phycologia 45:190–198
Lizotte MP (2008) Phytoplankton and primary production. In: Vincent WF, Laybourn-Parry J (eds) Polar lakes and rivers. Oxford University Press, Oxford, pp 157–178
Masuda S, Eda S, Ikeda S, Mitsui H, Minamisawa K (2010) Thiosulfate-dependent chemolithoautotrophic growth of Bradyrhizobium japonicum. Appl Environ Microbiol 76:2402–2409
McKiness ZP, Cavanaugh CM (2005) The ubiquitous mussel: Bathymodiolus aff. brevior symbiosis at the Central Indian Ridge hydrothermal vents. Mar Ecol Prog Ser 295:183–190
Miller DN, Bryant JE, Madsen EL, Ghiorse WC (1999) Evaluation and optimization of DNA extraction and purification procedures for soil and sediment samples. Appl Environ Microbiol 65:4715–4724
Morris DP, Zagarese H, Williamson CE, Balseiro EG, Hargreaves BR et al (1995) The attenuation of solar UV radiation in lakes and the role of dissolved organic carbon. Limnol Oceanogr 40:1381–1391
Nakai R, Abe T, Baba T, Imura S, Kagoshima H, Kanda H, Kanekiyo A, Kohara Y, Koi A, Nakamura K, Narita T, Niki H, Yanagihara K, Naganuma T (2012a) Microflorae of aquatic moss pillars in a freshwater lake, East Antarctica, based on fatty acid and 16S rRNA gene analyses. Polar Biol 35:425–433
Nakai R, Abe T, Baba T, Imura S, Kagoshima H, Kanda H, Kohara Y, Koi A, Niki H, Yanagihara K, Naganuma T (2012b) Eukaryotic phylotypes in aquatic moss pillars inhabiting a freshwater lake in East Antarctica, based on 18S rRNA gene analysis. Polar Biol (published online before print). doi:10.1007/s00300-012-1188-1
Newman J, Gutteridge S (1993) The X-ray structure of Synechococcus ribulose-bisphosphate carboxylase/oxygenase-activated quaternary complex at 2.2 Å resolution. J Biol Chem 268:25876–25886
Ohtani S, Kudoh S, Tsuchiya Y, Suyama K, Imura S (2001) Species composition of freshwater algae in benthic moss pillars in lakes, Skarvsnes, Antarctica. Abstracts for XXIV Symp Polar Biol Tokyo, Natl Inst Polar Res, p 77
Padisák J (2004) Phytoplankton. In: Sullivan PE, Reynolds CS (eds) The Lakes handbook. Limnology and limnetic ecology, vol 1. Blackwell, Oxford, pp 251–308
Paerl HW, Pinckney JL (1996) Ice aggregates as a microbial habitat in Lake Bonney, Dry Valley Lakes, Antarctica: nutrient-rich microzones in an oligotrophic ecosystem. Antarctic J USA 31:220–222
Quesada A, Vincent WF, Lean DRS (1999) Community and pigment structure of Arctic cyanobacterial assemblages: the occurrence and distribution of UV-absorbing compounds. FEMS Microbiol Ecol 28:315–323
Quesada A, Fernandez-Valiente E, Hawes I, Howard-Williams C (2008) Benthic primary production in polar lakes and rivers. In: Vincent WF, Laybourn-Parry J (eds) Polar lakes and rivers. Oxford University Press, Oxford, pp 179–196
Saitou N, Nei M (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Schneider G, Lindqvist Y, Lundqvist T (1990) Crystallographic refinement and structure of ribulose-1,5-bisphosphate carboxylase from Rhodospirillum Rubrum at 1.7 Å resolution. J Mol Biol 211:989–1008
Shinozaki K, Yamada C, Takahata N, Sugiura M (1983) Molecular cloning and sequence analysis of the cyanobacterial gene for the large subunit of ribulose-1,5-bisphosphate carboxylase/oxygenase. Proc Natl Acad Sci USA 80:4050–4054
Spiridonova EM, Kuznetsov BB, Pimenov NV, Turova TP (2006) Phylogenetic characterization of endosymbionts of the hydrothermal vent mussel Bathymodiolus azoricus by analysis of the 16S rRNA, cbbL, and pmoA genes. Mikrobiologiia 75:798–806
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28:2731–2739
Tanabe Y, Kudoh S, Imura S, Fukuchi M (2008) Phytoplankton blooms under dim and cold conditions in freshwater lakes of East Antarctica. Polar Biol 31:199–208
Tanabe Y, Ohtani S, Kasamatsu N, Fukuchi M, Kudoh S (2010) Photophysiological responses of phytobenthic communities to the strong light and UV in Antarctic shallow lakes. Polar Biol 33:85–100
Tsubota H, Nakao N, Arikawa T, Yamaguchi T, Higuchi M, Deguchi H, Seki T (1999) A preliminary phylogeny of Hypnales (Musci) as inferred from chloroplast rbcL sequence data. Bryol Res 7:233–248
van Berkum P, Keyser HH (1985) Anaerobic growth and denitrification among different serogroups of soybean rhizobia. Appl Environ Microbiol 49:772–777
van der Wielen PWJJ (2006) Diversity of ribulose-1,5-bisphosphate carboxylase/oxygenase large-subunit genes in the MgCl2-dominated deep hypersaline anoxic basin discovery. FEMS Microbiol Lett 259:326–331
Vincent WF (1988) Microbial ecosystems of Antarctica. Cambridge University Press, Cambridge
Vincent WF (2000) Cyanobacterial dominance in the polar regions. In: Whitton BA, Potts M (eds) The ecology of cyanobacteria: their diversity in time and space. Kluwer, Dordrecht, pp 321–340
Vincent WF, Downes MT, Castenholz RW, Howardwilliams C (1993a) Community structure and pigment organization of cyanobacteria-dominated microbial mats in Antarctica. Eur J Phycol 28:213–221
Vincent WF, Castenholz RW, Downes MT, Howardwilliams C (1993b) Antarctic cyanobacteria: light, nutrients and photosynthesis in the microbial mat environment. J Phycol 29:745–755
Vincent WF, Rae R, Laurion I, Howard-Williams C, Priscu JC (1998) Transparency of Antarctic ice-covered lakes to solar UV radiation. Limnol Oceanogr 43:618–624
Wing KT, Priscu JC (1993) Microbial communities in the permanent ice cap of Lake Bonney, Antarctica: relationships among chlorophyll-a, gravel and nutrients. Antarctic J USA 28:246–249
Acknowledgments
We gratefully acknowledge the members of the 42nd Japanese Antarctic Research Expedition (JARE). We thank Kazuko Ohishi and Tadasu Shin-i for their technical assistance. This study was supported by the National Polar Research Institute and the Transdisciplinary Research Integration Center, Research Organization of Information and Systems, Japan. This study also supported by JSPS Research Fellowships for Young Scientists (No. 11J30005). The study was conducted as part of the Microbiological and Ecological Responses to Global Environmental Changes in Polar Regions (MERGE) program during the International Polar Year of 2007–2008.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Nakai, R., Abe, T., Baba, T. et al. Diversity of RuBisCO gene responsible for CO2 fixation in an Antarctic moss pillar. Polar Biol 35, 1641–1650 (2012). https://doi.org/10.1007/s00300-012-1204-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-012-1204-5