
Abstract
An adaptive network model using SIS epidemic propagation with link-type-dependent link activation and deletion is considered. Bifurcation analysis of the pairwise ODE approximation and the network-based stochastic simulation is carried out, showing that three typical behaviours may occur; namely, oscillations can be observed besides disease-free or endemic steady states. The oscillatory behaviour in the stochastic simulations is studied using Fourier analysis, as well as through analysing the exact master equations of the stochastic model. By going beyond simply comparing simulation results to mean-field models, our approach yields deeper insights into the observed phenomena and help better understand and map out the limitations of mean-field models.









Similar content being viewed by others

References
Demirel G, Vazquez F, Böhme GA, Gross T (2014) Moment-closure approximations for discrete adaptive networks. Phys D Nonlinear Phen 267:68–80
Dmitriev N, Dynkin E (1945) On the characteristic roots of stochastic matrices. C R (Dokl) Acad Sci URSS 49:159–162
Eames KTD, Keeling MJ (2002) Modeling dynamic and network heterogeneities in the spread of sexually transmitted diseases. Proc Natl Acad Sci USA 99:13330–13335
Gillespie DT (1976a) A general method for numerically simulating the stochastic time evolution of coupled chemical reactions. J Comput Phys 22:403–434
Gillespie DT (1976b) Exact stochastic simulation of coupled chemical reactions. J Phys Chem 81:2340–2361
Gross T, D’Lima CJD, Blasius B (2006) Epidemic dynamics on an adaptive network. Phys Rev Lett 96:208701
Gross T, Kevrekidis IG (2008) Robust oscillations in SIS epidemics on adaptive networks: coarse graining by automated moment closure. Euro Phys Lett 82:38004
Gross T, Blasius B (2008) Adaptive coevolutionary networks: a review. J R Soc Interface 5:259–271
Halliday DM, Rosenberg JR, Amjad AM, Breeze P, Conway BA, Farmer SF (1995) A framework for the analysis of mixed time series/point process data-theory and application to the study of physiological tremor, single motor unit discharges and electromyograms. Prog Biophys Mol Biol 64:237–278
House T, Keeling MJ (2011) Insights from unifying modern approximations to infections on networks. J R Soc Interface 8:67–73
Juher D, Ripoll J, Saldaña J (2013) Outbreak analysis of an SIS epidemic model with rewiring. J Math Biol 67:411–432
Keeling MJ (1999) The effects of local spatial structure on epidemiological invasions. Proc R Soc Lond B 266:859–867
Kiss IZ, Berthouze L, Taylor TJ, Simon PL (2012) Modelling approaches for simple dynamic networks and applications to disease transmission models. Proc R Soc A 468(2141):1332–1355
Marceau V, Noël PA, Hébert-Dufresne L, Allard A, Dubé LJ (2010) Adaptive networks: coevolution of disease and topology. Phys Rev E 82:036116
Rogers T, Clifford-Brown W, Mills C, Galla T (2012) Stochastic oscillations of adaptive networks: application to epidemic modelling. J Stat Mech 2012:P08018
Sayama H, Pestov I, Schmidt J, Bush B, Wong C, Yamanoi J, Gross T (2013) Modeling complex systems with adaptive networks. Comput Math Appl 65:1645–1664
Shaw LB, Schwartz IB (2008) Fluctuating epidemics on adaptive networks. Phys Rev E 77:066101
Simon PL, Taylor M, Kiss IZ (2010) Exact epidemic models on graphs using graph automorphism driven lumping. J Math Biol 62:479–508
Szabó A, Simon PL, Kiss IZ (2012) Detailed study of bifurcations in an epidemic model on a dynamic network. Differ Equ Appl 4:277–296
Taylor M, Taylor TJ, Kiss IZ (2012) Epidemic threshold and control in a dynamic network. Phys Rev E 85:016103
Zhou J, Xiao G, Cheong SA, Fu X, Wong L, Ma S, Cheng TH (2012) Epidemic reemergence in adaptive complex networks. Phys Rev E 85:036107
Acknowledgments
Péter L. Simon acknowledges support from the Hungarian Scientific Research Fund (OTKA) (Grant No. 81403).
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix A: Bifurcation analysis of the mean-field model
In this appendix the steady states of system (1)–(4) and their stability is studied.
1.1 A1: Endemic steady state
The steady states are given by system (1)–(4) by putting zeros in the left hand sides (and omitting those parameters that are assumed to be zero), i.e. by system
where the closures
are used with
For simplicity, the steady state values of the four variables are denoted here by [I], [SI], [II] and [SS]. All of them will be expressed in terms of [S] and then a quartic equation for [S] will be derived. It is obvious that \([I]=N-[S]\), then from (8) we get \([SI]=\frac{\gamma }{\tau }[I]\). Multiplying (9) by 2 and adding it to (10) and to (11) one can express [SS] in terms of [I] and [S] as
From (10) we can express [II] in terms of [I] and [S] as
Multiplying (9) by 2 and adding it to (10) we get
Using the closure and dividing by 2[SI] yields
Now substituting \([SI]=\frac{\gamma }{\tau }(N-[S])\) and (13) into this equation one obtains the following quartic equation for \(x=[S]\)
with
where \(a=\frac{\omega _{SI}}{\alpha _{SS}}\), \(b=\frac{\gamma }{\tau }\) and \(c=\frac{\omega _{SI}}{\tau }.\)
Once (15) is solved for x, then the steady state ([I], [SI], [II], [SS]) is given by \([I]=N-x\), \([SI]=\frac{\gamma }{\tau }[I]\), (13) and (14). A solution x yields a biologically meaningful steady state if all of its coordinates are non-negative. An extensive numerical study showed that for any combination of the parameters there can be at most one endemic steady state. The endemic steady state exists if the disease-free equilibrium is unstable. The condition for that is determined in the next subsection. We note that the above calculation yields only the endemic steady states because during the derivation of the quartic equation there was a division by 2[SI], which is zero at the disease-free steady state.
1.2 A2: Stability of the disease-free steady state
The stability of the disease-free steady state is determined by the Jacobian matrix determined at \(([I], [SI], [II], [SS])=(0,0,0,N(N-1))\). In order to compute this matrix we determine the partial derivatives of the triples at this steady state, as they are given by the closures (12).
Using these partial derivatives the Jacobian matrix at the disease-free steady state can be given as
It can be easily seen that \(-\alpha _{SS}\) and \(-\gamma \) are eigenvalues of this matrix. The remaining two eigenvalues are the eigenvalues of the \(2\times 2\) matrix in the middle:
The disease-free steady state is asymptotically stable if and only if all the eigenvalues have negative real part. This has to be checked only for the above \(2\times 2\) matrix. For a \(2\times 2\) matrix the eigenvalues have negative real part if and only if its determinant is positive and its trace is negative. The determinant is positive if \(\gamma + \omega _{SI} -\tau (N-2)>0\). The trace is positive if \(3\gamma + \omega _{SI} -\tau (N-3)>0\). The first condition implies the second one, hence we proved Proposition 1.
1.3 A3: Stability of the endemic steady state
The stability of the endemic steady state can be determined only numerically. For a given set of the parameters the coordinates of the endemic steady state can be computed according to Appendix A1. The partial derivatives in the Jacobian J can be calculated analytically, then substituting the numerically obtained coordinates of the endemic steady state we get the entries of the Jacobian numerically. This enables us to calculate the coefficients of the characteristic polynomial
where \(b_3 = {{\mathrm{Tr}}}J\), \(b_0 = \det J\) and \(b_1,b_2\) can be given as the sum of some subdeterminants of the Jacobian, the concrete form of which is not important at this moment. To find the parameter values where Hopf bifurcation occurs we use the method introduced in Szabó et al. (2012). In the case of \(4\times 4\) matrices the necessary and sufficient condition for the existence of pure imaginary eigenvalues is
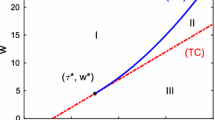
These relations will be considered here as necessary conditions for the Hopf-bifurcation. The transversality conditions will not be checked because of their complexity; instead, parameter values on the two sides of the bifurcation curve are chosen and the behaviour of the corresponding systems is checked numerically, to show that a Hopf-bifurcation really occurs along that curve. The Hopf-bifurcation set in the \((\tau ,\omega _{SI})\) parameter plane can be obtained as follows. For a given value of \(\tau \) we compute the value of \(b_0b^2_3= b_1(b_2b_3-b_1)\) numerically as \(\omega _{SI}\) is varied. It turns out that for a range of \(\tau \) values this expression changes sign twice as \(\omega _{SI}\) is varied. More precisely, for given values of the other parameters (\(N, \gamma , \alpha _{SS}\)) there exist \(\tau \) values \(\tau _1\) and \(\tau _2\), such that for \(\tau \in (\tau _1, \tau _2)\) we get \(\omega _1\) and \(\omega _2\) such that for \(\omega _{SI}=\omega _i\) (\(i=1,2\)) we have \(b_0b^2_3= b_1(b_2b_3-b_1)\), i.e. Hopf bifurcation occurs at \(\omega _{SI}=\omega _i\) (\(i=1,2\)). If \(\tau \) is not in the interval \((\tau _1, \tau _2)\), then there is no Hopf bifurcation, i.e., the relation \(b_0b^2_3= b_1(b_2b_3-b_1)\) cannot hold. For \(\tau \in (\tau _1, \tau _2)\) and \(\omega _{SI}\in (\omega _1,\omega _2)\) there is a stable periodic orbit. If \(\omega _{SI}\) is outside the interval \((\omega _1,\omega _2)\), then there is no periodic orbit and either the endemic or the disease-free steady state is stable. The final state of the system is shown in the bifurcation diagram in Fig. 1.
Appendix B: State space and transition matrix for adaptive graphs with \(N=2\) and \(N=3\)
In the case \(N=2\), i.e., for a graph with two nodes, the number of edges is at most \(E=1\). That is there are two graphs on two nodes, one is a single edge the other one consists of two disjoint nodes. We denote the states with \(\{ SS, SI, IS, II\}\) when the graph consists of two disjoint nodes. Here the state SI means that node 1 is S and node 2 is I. The states are denoted by \(\{ \overline{SS}, \overline{SI}, \overline{IS}, \overline{II} \}\) when the graph is a single edge. Thus the full state space for \(N=2\) contains the following 8 states: \(\{ SS, SI, IS, II, \overline{SS}, \overline{SI}, \overline{IS}, \overline{II}\}\). Consider now the transitions between these states. There are two types of transitions: epidemic transitions (infection and recovery) and network transitions (creating and deleting edges). Epidemic transitions may occur among states that belong to the same graph, that is within the subsets \(\{ SS, SI, IS, II\}\) and \(\{ \overline{SS}, \overline{SI}, \overline{IS}, \overline{II} \}\). Within the first subset only recovery may occur since these states belong to a graph that consists of two separate nodes. So the only possible transitions are \(SI\rightarrow SS\), \(IS\rightarrow SS\), \(II\rightarrow IS\) and \(II\rightarrow SI\), these may happen with rate \(\gamma \). Within the subset \(\{ \overline{SS}, \overline{SI}, \overline{IS}, \overline{II} \}\) infection may happen as well, hence the possible transitions are \(\overline{SI}\rightarrow \overline{II}\) and \(\overline{IS}\rightarrow \overline{II}\) with rate \(\tau \) and \(\overline{SI}\rightarrow \overline{SS}\), \(\overline{IS}\rightarrow \overline{SS}\), \(\overline{II}\rightarrow \overline{IS}\) and \(\overline{II}\rightarrow \overline{SI}\) with rate \(\gamma \). Network transitions occur between states in which the corresponding nodes are of the same type. For example, the transition \(SS\rightarrow \overline{SS}\) occurs at rate \(\alpha _{SS}\) since an SS type edge is created during this transition. Similarly, the transition \(\overline{SS}\rightarrow SS\) occurs at rate \(\omega _{SS}\) since an SS type edge is deleted during this transition. In general, the transition \(AB\rightarrow \overline{AB}\) happens at rate \(\alpha _{AB}\) and the transition \(\overline{AB}\rightarrow AB\) happens at rate \(\omega _{AB}\), where \(A,B\in \{S,I\}\). All the transitions are shown in Fig. 9. If the states are ordered as \(\{ SS, SI, IS, II, \overline{SS}, \overline{SI}, \overline{IS}, \overline{II}\}\), then the transition matrix of the corresponding Markov chain takes the form
where
The matrices \(M_{11}\) and \(M_{22}\) contain the transition rates corresponding to the epidemic transitions. These rates belong to the transitions within the subsets \(\{ SS, SI, IS, II\}\) and \(\{ \overline{SS}, \overline{SI}, \overline{IS}, \overline{II} \}\), respectively. The matrices A and \(\Omega \) contain the network transition rates that correspond to transitions between these two subsets. The master equation can be written as \(\dot{x}(t) = M x(t)\), where the coordinates of the eight dimensional vector x(t) are the probabilities of the states at time t.
Let us briefly consider the case \(N=3\). Then \(E=3\), hence there are \(2^3=8\) different possible graphs, one without any edges, three graphs with one edge, three line graphs with two edges and a triangle with three edges. Each graph can be in 8 possible states, namely \(\{ SSS, SSI, SIS, ISS, SII, ISI, IIS, III\}\). Hence there are \(2^3\cdot 2^3=64\) states altogether. Epidemic transitions may occur among states that belong to the same graph, for example, in the case of a triangle graph the transition \(IIS\rightarrow III\) happens at rate \(2\tau \), while its rate is \(\tau \) for a line graph where node 2 is connected to node 1 and node 3. The transition rate may be zero, e.g., in the case when there is only one edge in the graph connecting nodes 1 and 2, or in the case when there are no edges at all in the graph. Network transitions occur between states in which the corresponding nodes are of the same type. For example, denoting by SSS the state where all nodes are susceptible and the graph consist of three disjoint nodes, and by \(\overline{SS}S\) the state where all nodes are susceptible and the graph contains one edge that connects node 1 and 2, the rate of transition \(SSS\rightarrow \overline{SS}S\) is \(\alpha _{SS}\), while the rate of transition \(\overline{SS}S\rightarrow SSS\) is \(\omega _{SS}\). The master equations take again the form \(\dot{x}(t) = M x(t)\), where the coordinates of the 64-dimensional vector x(t) are the probabilities of the states at time t.
We note that the size of the state space can be reduced by lumping some states together, similarly to the case of static graphs (Simon et al. 2010). The lumping of the state space for dynamic network processes is beyond the scope of this paper, here we only mention a few simple cases where lumping can be carried out. In the case \(N=2\) it is easy to see that the states SI and IS can be lumped together, which means that their sum can be introduced as a new variable, and their differential equations can be added up. Similarly, the sum of \(\overline{SI}\) and \(\overline{IS}\) can be introduced as a new variable. (By adding their differential equations one can immediately see that the old variables will not appear in the remaining system of equations). Hence the eight-dimensional system \(\dot{x}(t) = M x(t)\) can be reduced to a six-dimensional system by lumping. In the case \(N=3\) we have even more chance for lumping if it is assumed that \(\alpha _{SI}=0\), \(\alpha _{II}=0\), \(\omega _{SS}=0\), \(\omega _{II}=0\). Without explaining the details we claim that in this case the 64-dimensional system of master equations can be reduced to a 20-dimensional one by lumping. In the case \(N=4\) the dimension of the state space can be reduced from 1024 to 89.
Rights and permissions
About this article

Cite this article
Szabó-Solticzky, A., Berthouze, L., Kiss, I.Z. et al. Oscillating epidemics in a dynamic network model: stochastic and mean-field analysis. J. Math. Biol. 72, 1153–1176 (2016). https://doi.org/10.1007/s00285-015-0902-3
Received:
Revised:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00285-015-0902-3