
Abstract
Bacillus lentus alkaline peptidase (BLAP) was used for casein (CN) hydrolysis in an enzyme membrane reactor (EMR) because it was found that BLAP was competitively inhibited by its products. The employed membranes had different molecular weight cut-offs (MWCO 1, 5 and 10 kDa). It was shown that the productivity of the EMR could be significantly improved (28 %) in comparison with batch hydrolysis under the same conditions after 20 h. All resulting EMR peptide mixtures showed a homogenous peptide pattern in HPLC–UV analysis. The obtained peptide mixtures exhibited Angiotensin-I-converting enzyme (ACE) inhibitory and antioxidative activity. The ACE inhibition of the peptide mixtures was dependent on the MWCO of the membranes. The resulting apparent IC50 values were 115, 131 and 420 μg ml−1 for the 1, 5 and 10 kDa MWCO membranes, respectively. In kinetic studies, a mixed-type inhibition was observed for the three peptide mixtures. The radical scavenging activity was determined with the ABTS assay, and IC50 values between 20 and 25 μg ml−1 were obtained for the generated peptide mixtures. In addition, the identified VYPFPGPIPN peptide exhibited ACE inhibition and antioxidant activity with IC50 values of 325 and 6.2 μM, respectively. The peptide YQEPVLGPVRGPFPIIV exhibited radical scavenging activity with an IC50 value of 5.2 μM.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Bacillus sp. are widely used for the production of several industrially utilised enzymes, such as amylases or peptidases [1]. The alkalophilic Bacillus lentus is one of these industrial organisms that produce a commercial peptidase preparation called ‘BLAP’ (Bacillus lentus alkaline peptidases) [2]. Today, one of the well-known applications of BLAP is in household detergents. Other applications should be sought, because a controlled and ‘tailor-made’ protein hydrolysis using affordable enzymes would also be of interest in food processing.
The enzymatic hydrolyses of proteins in batch processes have several disadvantages compared with hydrolyses in continuous systems, such as enzyme membrane reactors (EMRs). For example, the protein batch hydrolyses are more expensive because it is not possible to reuse the enzymes [3, 4]. Enzyme membrane reactor processes also have certain advantages due to the ease of immobilising the enzymes [5]. In such processes, peptidase activities can be exploited over a long reaction time, resulting in high productivity, continuous production of products of a consistent quality and permanent removal of the inhibiting peptide products [6–8]. An additional advantage for potential application in food processing is that enzyme inactivation is not required because this so-called ‘processing aid’ (enzyme) does not appear in the peptide mixtures (product) if the membranes are able to completely retain them.
Small peptides hydrolysed from milk and other protein sources have shown several bioactivities, such as inhibiting the Angiotensin-I-converting enzyme (ACE; EC 3.4.15.1) and lipoxygenase (LOX; EC 1.13.11.12) as well as antioxidative, antimicrobial and immunomodulatory properties [9–12]. Age-specific diseases have been related to oxidative stress [13], which might be reduced by antioxidants in food. The antioxidative properties of amino acids, peptides and proteins have been known for a long time [14–16]. In addition, the biofunctional and technofunctional properties of peptides are an interesting field of investigation. For example, lipid peroxidation causes deterioration in food quality, off-flavour, unacceptable taste and a shortened shelf life during storage.
The aim of our studies was to prepare consistent peptide mixtures by applying BLAP in an EMR using membranes with different molecular weight cut-offs (MWCOs). The assumed product inhibition that occurs frequently when working with BLAP should be prevented by use of the membranes. The standardised peptide mixtures were investigated for their ACE inhibitory activity and antioxidative capacity. To our knowledge, this is the first report of an investigation of BLAP in an EMR for this type of application.
Materials and methods
Reagents
Bacillus lentus alkaline peptidase (BLAP) was a kind gift from Biozym Scientific GmbH (Hessisch Oldendorf, Germany). Casein, hippuryl-l-histidyl-l-leucine (HHL) (≥98 %) and 2,2′azino-bis (3-ethylbenzothiazoline-6-sulphonic acid) diammonium salt (ABTS; 99 %) were obtained from Sigma-Aldrich (Schnelldorf, Germany). Peptide standards (95 % purity) were purchased from EMC (Tübingen, Germany). All other reagents used were of analytical grade.
Casein hydrolyses
Enzyme membrane reactor (EMR)
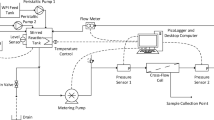
The EMR (Jülich Fine Chemicals, Jülich, Germany; Fig. 1) with a working volume of 10 ml was used with a 0.5 % (w/v) casein suspension at pH 8.0 in H2Odd, adjusted with NaOH (1 M). The EMR was operated at 40 °C to ensure a sufficient thermal stability of BLAP (unpublished data). Microbial growth was prevented by the addition of NaN3 [0.05 % (w/v)] to the ice-cooled feed tank at the beginning of the process, after 8 and 16 h. Transmembrane pressure was continuously monitored by a pressure gauge and did not exceed 2 bar. Casein hydrolysis was performed with BLAP (75 nkatcasein ml−1; spec. EA: 100 nkatcasein mg−1). During the hydrolysis process, casein (substrate) was retained by membranes with different molecular weight cut-offs [1, 5 and 10 kDa; polyethersulphone membranes, (PES)]. The substrate and permeate flows were kept constant at 10 ml h−1, so that a residence time of 1 h was achieved. The permeate was automatically fractionated (frac-100, GE, München, Germany) over 20 h. The productivity was calculated as gdry weight l−1 h−1, with the freeze-dried (CHRIST, ALPHA 1–2; Osterode am Harz, Germany) fractions corrected by salt using a freeze-dried blank without substrate (casein).

Design and flow chart of the enzyme membrane reactor used for production of biofunctional and technofunctional peptides
Batch hydrolyses
The batch casein hydrolysis was performed with a 10 % (w/v) casein suspension in H2Odd at pH 8.0 adjusted with NaOH (1 M) at 40 °C for 20 h. The hydrolysis was performed in a 10 ml scale using BLAP (75 nkatcasein ml−1; spec. EA: 100 nkatcasein mg−1). Microbial growth was prevented by the addition of NaN3 [0.05 % (w/v)] at the beginning of the process, after 8 and 16 h. After 20 h, the hydrolyses were stopped by ultrafiltration (10 kDa; Vivaspin 500, Sartorius, Göttingen, Germany). The permeate was freeze dried. The productivity was calculated as gdry weight l−1 h−1, with the freeze-dried fractions corrected by salt using a freeze-dried blank without substrate (casein).
Determination of proteolytic activity
The proteoloytic activity of BLAP was determined with a natural substrate and a chromogenic peptide.
Determination of proteolytic activity with the ninhydrin assay
Bacillus lentus alkaline peptidase activity was determined by a ninhydrin assay, which was performed according to the method of Reimerdes et al. [17] with minor modifications. Bovine casein [0.5 % (w/v)] dissolved in H2Odd at pH 8.0 (adjusted with 1 M NaOH) was used as the substrate. The substrate (450 μl) was preincubated with H2Odd (225 μl) at 40 °C for 5 min. Casein hydrolysis was initiated by the addition of the enzyme solution (225 μl, 40 °C). The enzymatic hydrolysis was terminated after 1–20 min in intervals of 1 min using 0.35 M TCA (1,075 μl). To precipitate the unhydrolysed casein, the assay mixture was centrifuged (13,000×g; 5 min). Afterwards, the resulting supernatant (250 μl) was mixed with acetate buffer (4 M, pH 5.5, 250 μl), NaOH (1 M, 75 μl) and ninhydrin reagent (225 μl, 1.74 g ninhydrin and 280 mg hydrindantin dihydrate dissolved in 100 ml methoxyethanol), incubated in a water bath (95 °C for 20 min) and then placed on ice for 5 min. Subsequently, the reaction mixture (300 μl) was mixed with 50 % (v/v) isopropanol/water (1,000 μl), and the absorbance was measured at 570 nm using a spectrophotometer (GE, München, Germany). The calibration curve was determined using tyrosine as a standard amino acid (45–450 μM). One kat was defined as the release of 1 mol tyrosine equivalent per second.
Determination of proteolytic activity with chromogenic substrate
Bacillus lentus alkaline peptidase activity was analysed with SUC-Ala-Ala-Pro-Phe-pNA (Bachem, Bubendorf, Switzerland) as substrate. To determine BLAP activity, 227.5 μl of the EMR permeate was used. The reaction mixture was preincubated at 40 °C for 10 min. Afterwards, the chromogenic peptide [12.5 μl of a 5 mg ml−1 solution, dissolved in dimethylformamide (DMF)] was added to start the reaction. The reaction was stopped by the addition of acetic acid [50 μl; 50 % (v/v)]. After centrifugation (8,000×g, 5 min), 240 μl of the solution was transferred to a microtiter plate, and the absorption was measured in an iEMS-Reader (Labsytems, Helsinki, Finland) at 405 nm. One katal of proteolytic activity was defined as the amount of enzyme that released 1 mol p-nitroaniline per second.
Determination of product inhibition of BLAP with azocasein assay
The azocasein assay was performed as described in [18] with minor modifications. Azocasein (5 mg ml−1) was dissolved in H2Odd, and the pH was adjusted to 8.0 (1 M NaOH). A 10 kDa ultrafiltration permeate from a 2 % (w/v) casein hydrolysis with 11 μkatcasein BLAP in H2Odd at pH 8.0 (10 h, 4.5 l scale; adjusted with 1 M NaOH) was used for BLAP inhibition studies. The casein hydrolysate was dissolved in each concentration of azocasein solution (0.25–5 mg ml−1). Azocasein inhibitor solution (250 μl) was preincubated at 40 °C for 5 min. The reaction was started by the addition of BLAP (25 μl; 1.5 mg ml−1 in H2O, 0.45 μm filtered). The reaction was terminated by TCA (2 M, 25 μl). Afterwards, the reaction mixture was centrifuged (13,000×g, 5 min). The resulting supernatant (187 μl) was transferred to a microtiter plate and NaOH (1 M, 62.5 μl) was added. The absorbance was measured at 450 nm in a microplate spectrophotometer (MULTISKAN FC, Thermo Scientific, Dreieich, Germany). One unit of enzyme activity was defined as the amount of enzyme which under the assay conditions described gives rise to an increase of one absorbancy at 450 nm min−1.
Angiotensin-I-converting enzyme (ACE) inhibition assay
The ACE inhibition assay was performed according to a previously published method with some modifications [19]. Hippuryl-l-histidine-l-leucine (HHL, 5 mM) was dissolved in borate buffer (0.1 M containing 0.3 M NaCl, pH 8.3) and used as the substrate. Porcine lung ACE (~37 U mg−1) was diluted in borate buffer (0.1 M containing 0.3 M NaCl, pH 8.3) to a final activity of 100 U l−1. The substrate (75 μl) and sample or buffer (10 μl) were preincubated at 37 °C for 5 min. The reaction was started by the addition of ACE (5 μl) and stopped by the addition of HCl (1 M, 20 μl) after 30 min. The products hippuric acid (HA) and l-histidine-l-leucine (HL), which were released during hydrolysis, were separated by RP-C18 HPLC (SpectraSYSTEMS™, Thermo Scientific™, Dreieich, Germany) and detected at 228 nm. Solvent A consisted of 0.1 % (v/v) formic acid (FA) in double-distilled water. Solvent B consisted of 0.1 % (v/v) FA in acetonitrile (ACN). The elution of HA and HL was performed with a gradient and a constant flow rate of 0.5 ml min−1. The conditions of the gradient elution were as follows: 0 min (A: 100 %; B: 0 %), 20 min (A: 50 %; B: 50 %), 22 min (A: 50 %; B: 50 %), 28 min (A: 100 %; B: 0 %), 35 min (A: 100 %; B: 0 %). A reversed phase C18 HPLC column (Aqua 5u 200 Å, 150 mm × 4.6 mm, 5 μM; Phenomenex, Germany) equipped with a GROM-SIL 120 ODS-3 CP; 3 μM (20 mm × 4.6 mm) precolumn was used. Usually, 10 μl was injected in the HPLC. The IC50 was defined as the amount of hydrolysate or peptide required to reduce the area of the HA peak to 50 %.
ABTS antioxidation assay
The ABTS [2,2′-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid) diammonium salt] antioxidation assay described in [20] was carried out with some modifications. The ABTS·+ stock solution was prepared with ABTS·+ (7 mM) and ammonium peroxodisulphate (2.45 mM) in double-distilled water. Both were allowed to react for 12–16 h in the dark. ABTS·+ stock solution was used for no more than 2 days. The ABTS·+ stock solution was diluted with phosphate-buffered saline (PBS; pH 7.4) to an absorption of 0.700 ± 0.05 at 734 nm. The antioxidative activity was measured at 734 nm in a spectrophotometer. The sample (10 μl) and diluted ABTS·+ reagent (1,000 μl) were mixed and incubated at 30 °C in the dark for 15 min. The IC50 was defined as the amount of peptide mixture, standard peptide or trolox (dissolved in ethanol) required to reduce the absorbance of ABTS·+ to 50 %. The antioxidant capacity of the two standard peptides was expressed as trolox equivalent antioxidant capacity (TEAC). TEAC was calculated as the quotient of the slope of the standard peptide divided by the slope of trolox.
HPLC–UV analysis of the EMR peptide mixtures
Enzyme membrane reactor hydrolysates were analysed by HPLC using a SpectraSYSTEMS™ chromatograph (Thermo Scientific, Dreieich, Germany). The peptides were eluted with two solvents. Solvent A consisted of 0.1 % (v/v) FA in H2Odd, and solvent B consisted of 0.1 % (v/v) FA in ACN. The conditions of the gradient were as follows: 0–5 min (A: 100 %; B: 0 %), 5–45 min (A: 40 %; B: 60 %), 45–55 min (A: 40 %; B: 60 %), 55–57 min (A: 100 %; B: 0 %), 57–65 min (A: 100 %; B: 0 %). The HPLC column was a reversed phase C18 (Aqua 5 u, 125 A, 250 mm × 4.6 mm, 5 μM; Phenomenex, Germany) equipped with a GROM-SIL 120 ODS-3 CP; 3 μM (20 mm × 4.6 mm) precolumn. Usually, 20 μl of the peptide mixture was injected. Separated peptides were eluted with a constant flow rate of 0.5 ml min−1 and detected at 220 nm.
Mass spectrometric detection of peptides
Nano-LC–ESI–MS/MS experiments were performed on an ACQUITY nano-UPLC system (Waters, USA) coupled to an LTQ-Orbitrap XLhybrid mass spectrometer (Thermo Fisher Scientific, Germany). The sample was concentrated and desalted on a precolumn (2 cm × 180 μm, Symmetry C18, 5 μm particle size, Waters, USA) and separated on a 20 cm × 75 μm BEH 130 C18 reversed phase column (1.7 μm particle size, Waters, USA). A gradient elution was performed from 1 to 50 % ACN in 0.1 % FA within 30 min. The LTQ-Orbitrap was operated under the control of XCalibur 2.0.7 software.
Results and discussion
Product inhibition of Bacillus lentus alkaline peptidase
Product inhibition occurs during batch food protein hydrolyses [21, 22], leading to decreased productivity. Therefore, BLAP was analysed for product inhibition using the azocasein assay (Fig. 2). This assay is based on the release of azo-derivatised casein peptides in the supernatant. Thus, non-derivatised peptides, which were generated by a prior BLAP casein hydrolysis, did not contribute to the absorbance measurement of the assay at 450 nm.

Product inhibition of BLAP analysed by a Lineweaver–Burk plot using the azocasein assay (450 nm, 40 °C, pH 8.0)
As shown in the Lineweaver–Burk plot, BLAP was inhibited by its products (Fig. 2). The apparent K m values for the azocasein substrate clearly increased with higher peptide concentrations. The intercept in Fig. 2 was minimally shifted from the y-axis; however, theoretically, various types of inhibitory interactions between the complex peptide mixture and BLAP were possible. Nevertheless, the inhibition type appeared to be competitive inhibition. In addition, in prior work, an apparent uncompetitive product inhibition of B. subtilis peptidases (Alcalase) was described using the laborious trinitrobenzene sulphonic acid (TNBS) assay [23], which is inappropriate at high blank values in combination with a low degree of hydrolysis (DH). Generally, the TNBS method should be used with caution at very low DH [24]. Therefore, we recommend using the azocasein assay in endopeptidase inhibition studies due to its feasibility. The azocasein assay was also used by Fahmy et al. [25] in inhibitory studies of a cysteine peptidase from wheat using specific cysteine peptidase inhibitors. The product inhibition of BLAP should be avoided by using an EMR because the generated peptides are continuously removed from the hydrolysis vessel. As a result, higher productivities should be achieved in protein hydrolysis.
Hydrolysis of casein in an enzyme membrane reactor (EMR)
The desired retention of BLAP through three different UF membranes (1, 5 and 10 kDa MWCOs) was checked in preliminary experiments (data not shown). In no case, BLAP activity could be detected in the permeates of the three MWCOs. The endopeptidases in BLAP did not pass through the membranes under the conditions used. Further investigations regarding the effect of the 1, 5 and 10 kDa membranes on the composition and bioactive properties of the resulting peptide mixtures were performed in a 10-ml-scale EMR (40 °C, 1 h residence time, 0.5 % casein feed, 75 nkatcasein ml−1 BLAP). First, the permeate samples containing the peptide mixtures were analysed by RP-C18 HPLC with UV detection. With each of the three membranes, the peptide patterns of the particular samples were consistent over the complete reaction time of 20 h. Similar results were obtained by Lin et al. [6], who mentioned that peptide mixtures produced in EMR have a higher homogeneity compared with peptide mixtures produced in batch hydrolyses. Comparing the particular HPLC–UV patterns of the three different peptide mixtures obtained with the three membranes, significant differences were observed between the peptide mixtures of the 1 and the 5 or 10 kDa MWCO membranes (Fig. 3). However, no significant differences in the HPLC–UV pattern of the peptides were observed between the 5 and 10 kDa MWCO membrane (data not shown).

Comparison of different HPLC–UV patterns of the resulting peptide mixtures in an EMR produced with 1 and 10 kDa MWCO membranes
The corresponding calculated space–time yields of the EMR decreased from 3.2 to 2.9 to 1.0 g l−1 h−1 for the 10, 5 and 1 kDa membranes, respectively (slopes in Fig. 4). Thus, feeding 5 g l−1 of casein over the reaction time of 20 h (100 g l−1 substrate in total), a final yield of 64 (10 kDa), 58 (5 kDa) and 20 g l−1 (1 kDa) was recovered as dried peptide mixture.

Productivity of casein hydrolysis using membranes with different molecular weight cut-offs (1, 5 and 10 kDa)
When using a 10 kDa membrane for peptide processing, the EMR reaction was compared with a batch reaction, applying the same total amounts of BLAP and substrate over 20 h. The total peptide yield was 28 % higher with the EMR (64 g l−1) than the batch approach (50 g l−1). We assume that both, the apparent prevention of product inhibition and the lower substrate concentration were responsible for the better performance of BLAP in the EMR.
Identification of generated peptides by nano-LC–ESI–MS/MS
The three different peptide mixtures were subjected to mass spectrometry to identify the generated peptide sequences. Peptide mixtures produced with 5 and 10 kDa MWCOs each contained 128 peptides that were mostly similar but not identical. Fifty-one peptides were distinguished. As expected, the peptide mixture produced with 1 kDa MWCO membrane contained considerably more peptides (221). For the penetration of the 1-kDa membrane, a higher degree of hydrolysis was necessary compared to the 5 and 10 kDa MWCOs, and thus, the total number of peptides was increased. All identified peptide sequences were aligned with a database (BIOPEP database) and the literature (Scopus). The seven bioactive peptides identified are listed in Table 1.
Bioactive potential of the peptide mixtures
The three different peptide mixtures were lyophilised and examined for their bioactive potential, that is, their ACE inhibitory activity and antioxidative effects. Two of the most abundant peptides (VYPFPGPIPN and YQEPVLGPVRGPFPIIV) from the RP-C18-MS/MS analysis were chemically synthesised and separately evaluated for their bioactive potential.
Antihypertensive effect and mechanism of ACE inhibition
The ACE inhibitory effect of the three different peptide mixtures was investigated. The apparent IC50 values were determined for all peptide mixtures with a minimum of seven concentrations and were calculated to be 115, 131 and 420 μg ml−1 for the 1, 5 and 10 kDa MWCO membranes, respectively. The apparent IC50 values of the 1 and 5 kDa peptide mixtures were lower than that reported for fermented milk, which was 266 μg ml−1 [26]. The concentration-dependent ACE inhibition for the 5-kDa peptide mixture is shown in Fig. 5a. The type of ACE inhibition was analysed for the continuously produced peptide mixtures by linearisation after Lineweaver–Burk analysis (Fig. 5b). An apparent mixed-type inhibition of the ACE was observed in all three cases. The apparent K i values were calculated with 31, 32 and 115 μg ml−1 for the 1, 5 and 10 kDa MWCO peptide mixtures, respectively. In comparison, the apparent K i -value for captopril, a competitive pharmaceutical inhibitor for the treatment of hypertension, is reported to be 6.7 ng ml−1 [27]. However, the presence of peptides with different inhibition mechanisms was still possible due to the heterogeneity of the generated peptide mixtures. Typically, the mechanism of ACE inhibition is competitive [28]; however, uncompetitive, non-competitive, competitive as well as mixed inhibition mechanisms have been reported in the literature [27–29].

ACE inhibition by the EMR 5 kDa peptide mixture (a) and Lineweaver–Burk plot of ACE inhibition in the presence of the BLAP 5 kDa MWCO peptide mixture (b)
Furthermore, the two peptides V-β-casomorphin-9 (VYPFPGPIPN) and casecidin-17 (YQEPVLGPVRGPFPIIV) were assayed separately for ACE inhibitory activity. Casecidin-17 showed no ACE inhibitory activity at all in our trials. The IC50 of V-β-casomorphin-9 against ACE was determined to be 325 μM. In the case of V-β-casomorphin-9, Saito et al. [30] previously published an IC50 value of 14.8 μM for β-casomorphin-9 (YPFPGPIPN), which exhibits a structural resemblance to V-β-casomorphin-9 (VYPFPGPIPN). Therefore, we conclude that additional Val at the N-terminal end of V-β-casomorphin-9 increased the IC50 value almost 22-fold from 14.8 to 325 μM.
Antioxidative effects of the peptide mixtures
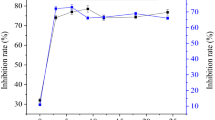
The antioxidative effects of the three peptide mixtures and the two peptides V-β-casomorphin-9 and casecidin-17 were determined with the ABTS decolorisation assay. The antioxidant activities of the peptide mixtures were expressed as IC50 values in μg ml−1 and compared with the standard trolox, a water-soluble vitamin E derivative. In our results, dose-dependent radical scavenging activity was observed.
The IC50 values were calculated to be 24, 25 and 20 μg ml−1 for the peptide mixtures produced with the 1, 5 and 10 kDa MWCO membranes, respectively. The substrate CN (IC50 = 36 μg ml−1) itself also showed radical scavenging activity. In comparison with the most effective peptide mixture obtained by the 10 kDa EMR (20 μg ml−1), 44 % more antioxidant capacity was generated due to the hydrolysis of CN.
Both peptides, casecidin-17 and V-β-casomorphin-9, showed a dose-dependent antioxidative effect [Fig. 6a, b]. Their IC50 values were 5.2 and 6.2 μM, respectively. Sadat et al. [31] also identified tyrosine-containing peptides from bovine α-lactalbumin hydrolysis. The calculated IC50 values were 6.2, 8.0 and 0.5 μM for the peptides YDTQA, INY and INYW, respectively. An antioxidative peptide with the sequence IVGGFPHYL was recently purified from sand eel Hypoptychus dybowskii by Lee et al. [32]. The purified peptide had a mass of 1,189 Da and an EC50 value of 22.75 μM determined with the DPPH assay.

Relationship between the concentration and antioxidant activity of casecidin-17 (a) and V-β-casomorphin-9 (b)
The resulting TEACs of the peptides were 1.95 (V-β-casomorphin-9) and 2.3 (casecidin-17), so both demonstrated a significantly higher antioxidant capacity than trolox.
Conclusions
Bacillus lentus alkaline peptidase was inhibited by its peptides produced during CN hydrolysis in a batch reactor. The product inhibition could be avoided by using an EMR. This strategy increased the productivity of the EMR (10 kDa MWCO membrane) up to 28 % compared with the batch hydrolysis when equal total amounts of CN were used. Different peptide mixtures were continuously generated by BLAP in the EMR using different MWCOs (1, 5 and 10 kDa). These peptide mixtures exhibited diverse ACE inhibitory and antioxidant activities. In kinetic studies, the peptide mixtures showed a mixed-type inhibition of ACE, and in addition, the apparent IC50 values were dependent on the employed MWCOs (1 < 5 < 10 kDa). The CN hydrolysate produced by the 10 kDa MWCO EMR showed the highest antioxidant activity of the three peptide mixtures. The peptide VYPFPGPIPN was identified as a strong antioxidative peptide and a weak ACE inhibitor, whereas the second verified peptide, YQEPVLGPVRGPFPIIV, showed no ACE inhibition but had strong antioxidant capacity.
References
De Boer AS, Priest F, Diderichsen B (1994) Appl Microbiol Biotechnol 40:595–598
Jørgensen PL, Tangney M, Pedersen PE, Hastrup S, Diderichsen B, Jørgensen ST (2000) Appl Environ Microbiol 66:825–827
Cheftel C, Ahern M, Wang DIC, Tannenbaum SR (1971) J Agric Food Chem 19:155–161
Guadix A, Camacho F, Guadix EM (2006) J Food Eng 72:398–405
Trusek-Holownia A, Noworyta A (2008) Desalination 221:543–551
Lin SB, Chiang WD, Cordle CT, Thomas RL (1997) J Food Sci 62:480–483
Wei JT, Chiang BH (2009) J Sci Food Agric 89:372–378
Cheison SC, Wang Z, Xu SY (2007) J Agric Food Chem 55:3896–3904
Jäkälä P, Vapaatalo H (2010) Pharmaceuticals 3:251–272
Jung WK, Mendis E, Je JY, Park PY, Byeng WS, Hyoung CK, Yang KC, Kim SK (2006) Food Chem 94:26–32
Hsu KC (2010) Food Chem 122:42–48
Schurink M, van Berkel WJH, Wichers HJ, Boeriu CG (2007) Peptides 28:2268–2275
Pihlanto A (2006) Int Dairy J 16:1306–1314
Marcuse R (1960) Nature 186:886–887
Bishov SJ, Henick AS, Koch RB (1960) Food Res 25:174–182
Bishov SJ, Henick AS (1972) J Food Sci 37:873–875
Reimerdes EH, Klostermeyer H (1976) Method Enzymol 45:26–28
Leighton TJ, Doi RH, Warren RAJ, Kelln RA (1973) J Mol Biol 76:103–122
Cushman DW, Cheung HS (1971) Biochem Pharmacol 20:1637–1648
Re R, Pellegrini N, Proteggente A, Pannala A, Yang M, Rice-Evans C (1999) Free Radic Bio Med 26:1231–1237
Sousa R Jr, Lopes GP, Tardioli PW, Giordano RLC, Almeida PIF, Giordano RC (2004) Brazil J Chem Eng 21:147–153
Tardioli PW, Sousa R Jr, Giordano RC, Giordano RLC (2005) Enzyme Microb Technol 36:555–564
Demirhan E, Apar DK, Özbek B (2011) J Food Sci 76:C64–C67
Nielsen PM, Petersen D, Dambmann C (2001) J Food Sci 66:642–646
Fahmy AS, Ali AA, Mohamed SA (2004) Bioresour Technol 91:297–304
Tsai JS, Chen TJ, Pan BS, Gong SD, Chung MY (2008) Food Chem 106:552–558
Tsai JS, Chen JL, Pan BS (2008) Process Biochem 43:743–747
Mäkinen S, Johannson T, Vegarud Gerd E, Pihlava JM, Pihlanto A (2012) J Funct Foods 4:575–583
Pedroche J, Yust MM, Girón-Calle J, Alaiz M, Millán F, Vioque J (2002) J Sci Food Agric 82:960–965
Saito T, Nakamura T, Kitazawa H, Kawai Y, Itoh T (2000) J Dairy Sci 83:1434–1440
Sadat L, Cakir-Kiefer C, N’Negue MA, Gaillard JL, Girardet JM, Miclo L (2011) Int Dairy J 21:214–221
Lee WS, Jeon JK, Byun HG (2011) Process Biochem 46:1207–1211
Hayes M, Stanton C, Slattery H, O’Sullivan O, Hill C, Fitzgerald GF, Ross RP (2007) Appl Environ Microbiol 73:4658–4667
Kudoh Y, Matsuda S, Igoshi K, Oki T (2001) J Jpn Soc Food Sci 48:44–50
Contreras MdM, Carrón R, MdM Contreras, Montero MJ, Ramos M, Recio I (2009) Int Dairy J 19:566–573
Yamamoto N, Akino A, Takano T (1994) J Dairy Sci 77:917–922
Birkemo GA, O’Sullivan O, Ross RP, Hill C (2009) J Appl Microbiol 106:233–240
Acknowledgments
The authors would like to thank Dr. Jens Pfannstiel and Iris Klaiber for LC–ESI–MS/MS measurements. We would like to express our gratitude to the German Federal Ministry of Economics and Technology (AIF/FEI Project No. 16541 N) for partial financial support of this research.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Eisele, T., Stressler, T., Kranz, B. et al. Bioactive peptides generated in an enzyme membrane reactor using Bacillus lentus alkaline peptidase. Eur Food Res Technol 236, 483–490 (2013). https://doi.org/10.1007/s00217-012-1894-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00217-012-1894-5