
Abstract
The B Lymphocyte Stimulator (BLyS) family of ligands and receptors regulates humoral immunity by controlling B lymphocyte survival and differentiation. Herein, we review the ligands and receptors of this family, their biological functions, and the biochemical processes through which they operate. Pre-immune B lymphocytes rely on BLyS signaling for their survival, whereas antigen experienced B lymphocytes generally interact more avidly with a homologous cytokine, A Proliferation Inducing Ligand (APRIL). The molecular basis for signaling via the three BLyS family receptors reveals complex interplay with other B lymphocyte signaling systems, affording the integration of selective and homeostatic processes. As our understanding of this system advances, molecular targets for manipulating humoral immunity in both health and disease should be revealed.
Similar content being viewed by others


Introduction
B lymphocytes are the effectors of humoral immunity, providing defense from pathogens through antibody production. The induction of humoral immune responses involves selective activation of B cells within pre-immune pools. Each cell in these quiescent populations expresses a B cell antigen receptor (BCR) with a unique specificity. When antigens avidly cross-link the BCR, intracellular signals are generated that lead to activation and differentiation, culminating in the formation of antibody-forming plasma cells and memory B cells. These antigen-experienced populations maintain protective antibody levels and contribute to anamnestic responses upon subsequent antigen challenges. Accordingly, an effective humoral immune system relies on the maintenance of pre-immune pools that are large enough to contain both a broad array of BCR specificities and provide adequate surveillance given the organism’s size; as well as upon the generation and accumulation of memory and plasma cell compartments to sustain protective immunity.
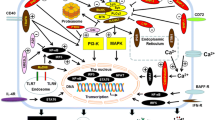
Our understanding of how the sizes, compositions, and behaviors of these related but independent pools are regulated has advanced substantially in the last decade, as appreciation for the molecular systems involved has grown. A central feature of current thought is that the establishment and maintenance of B cell pools relies on integrated signals from both the BCR and members of the B Lymphocyte Stimulator (BLyS, also termed BAFF) family of cytokines and receptors [1–3]. In these processes, the BCR provides specificity-based cues that determine the likelihood of successful maturation, activation, differentiation, and triage into various functional subsets [4]. In contrast, signals via BLyS family molecules directly mediate survival, determining the proportion of newly formed B cells that survive to enter mature pre-immune pools, the lifespan of these mature primary B cells, and the actions of antigen-experienced B cell subsets (Fig. 1). Since these survival processes impact most B cell activities, members of the BLyS family have emerged as key players in the etiology and treatment of autoimmunity and neoplasia; and as attractive targets for the manipulation of humoral immunity in vaccine development and transplant tolerance. Indeed, growing appreciation for the promise of these and other translational applications has prompted numerous commentaries and reviews [5–36].

Receptor Expression and Cytokine Dependence. B cells in pre-immune subsets express both BR3 and TACI, but only BLyS is required for their normal development. Antigenic stimuli result in changes in receptor expression. T cell dependent stimuli favor BR3 expression and lead to germinal center reactions, while T cell independent stimuli favor TACI expression on the resulting short-lived plasma cells. Although all recently activated B cells express one or more BLyS receptors, the exact roles played by BLyS and/or APRIL in their survival or differentiation has not been fully interrogated. Long-lived bone marrow plasma cells express BCMA, and require APRIL for normal survival. Memory cells express TACI, but are largely independent of either BLyS or APRIL
The BLyS family is a recent addition to the tumor necrosis factor (TNF) superfamily of biomolecules. It includes at least two ligands: BLyS itself and a “sister” cytokine named A Proliferation Inducing Ligand (APRIL); as well as three receptors, BLyS receptor 3 (BR3, also termed BAFFr), Transmembrane Activator and Calcium-signaling modulating and cyclophilin ligand (CAML) Interactor (TACI), and B Cell Maturation Antigen (BCMA). As with most TNF superfamily members, receptor ligation initiates interactions with one or more of the TNF receptor-associated factors (TRAFs) [37, 38], leading to downstream signaling cascades that can modulate survival and differentiation [39]. BLyS can interact with all three receptors; whereas APRIL binds BCMA and TACI, but not BR3. The distinct TRAF interactions of the three receptors, coupled with their dissimilar ligand-binding capacities and differential expression patterns, yields a broad range of combinations and thus affords independent regulation of various B cell subsets. For example, antigen-experienced B cells, such as memory B cells and long-lived bone marrow (BM) plasma cells, express different BLyS receptors than pre-immune B cells. Moreover, BLyS ligands may oligomerize or localize within specific anatomic compartments, fostering unique interactions or limiting trophic support to particular anatomic niches. Herein, we first review the nature, structure, and expression of BLyS family cytokines and receptors; followed by an in-depth discussion their actions and signaling characteristics in the context of B lymphocyte development, survival, and activation.
The BLyS Family of TNF Cytokines and Receptors
The TNF/TNF Receptor Family
Tumor necrosis factor family members impact many aspects of the immune system, ranging from organogenesis to lymphocyte activation, differentiation, and apoptosis (reviewed in [40]). In general, TNF ligands are type II transmembrane proteins whose active forms are homotrimers. These can act either at the cell surface in a membrane-bound form or, in some cases, as soluble cytokines after the post-translational cleavage of their extracellular regions. The TNF receptors are transmembrane proteins characterized by cysteine-rich domains (CRDs) in their ligand-binding regions. Some members of this receptor family also contain cytoplasmic death domains (DD) that interact with adaptor proteins known as TNF receptor-associated death domain proteins (TRADDs), and—as implied by their name—induce apoptosis upon ligation (e.g., Fas). Other TNF receptors lack DDs, and associate instead with TRAFs, which can mediate diverse downstream effects impacting survival, differentiation, or function. Finally, some TNF family members cross associate; yielding situations where more than one ligand can interact with a particular receptor, or where multiple receptors can bind the same ligand. This network of ligand-receptor pairs, as well as the cross talk and overlap in the downstream signaling cascades, yields the diversity needed to explain the wide variety of responses attributed to TNF family members.
Most members of the BLyS family were discovered through genomic homology searches [41–43]. Since some family members were discovered simultaneously by several laboratories, they possess multiple names and acronyms. A summary of the BLyS family ligands and receptors, their various synonyms and general properties is provided in Table 1. Throughout this review, we refer to the two ligands as APRIL and BLyS; and use the terms BCMA, TACI, and BR3 for the three receptors. The following two sections (BLyS family ligands and BLyS family receptors) detail the biochemical characteristics of BLyS subfamily ligands and receptors, respectively.
BLyS Family Ligands
There are two ligands in the BLyS family: BLyS and APRIL. Both cytokines are expressed by a variety of cell types, particularly among leukocytes. Both are initially synthesized as membrane-spanning monomers, but associate as trimers and are cleaved to soluble form by furin. Although they share homology with other TNF ligands, BLyS and APRIL are smaller than most—consisting of only transmembrane domains, a furin-cleavage site and a TNF-homology domain. In addition, receptor binding face structure and amino acid composition also distinguish these ligands from other superfamily members [44].
APRIL
APRIL was isolated based on mRNA expression in cell lines, and so-named because it could provoke proliferation in transfected fibroblasts [45]. The gene encoding APRIL is found on chromosome 17p13.3 in humans and chromosome 11 in the mouse [46]. Several features of this gene’s organization are noteworthy. First, it includes a furin-cleavage site exon, thus allowing the gene product to generate soluble forms. Second, the TRAF homology domain encoding region is interrupted by an intron, whereas most TNF family members use a single exon to encode this domain. Finally, the gene encoding APRIL is juxtaposed with another TNF family member named TNF Weak Inducer of Apoptosis (TWEAK) [47, 48]. This tandem organization of TWEAK and APRIL genes is conserved from mice to humans, suggesting a functional relationship; but again the biological significance is unclear. Interestingly, messenger RNA for a fusion product of the TWEAK and APRIL genes has been reported (termed TWE-PRIL), and the predicted structure for this fusion protein suggests TWE-PRIL could bind BCMA and TACI, but that processing to an active soluble form would follow pathways characteristic of TWEAK [48]. Equally intriguing, the TWEAK receptor, FN14, bears striking resemblance to BCMA both in terms of structure and TRAF binding capacity.
The 250 amino acid APRIL molecule shares only 20–30% homology with most other TNF family members; but displays nearly 50% homology with BLyS in the TNF-homology domain [49–52]. Of the three currently known BLyS family receptors, APRIL can interact with BCMA and TACI, but not with BR3. APRIL also bears an N-terminal region that interacts with surface proteoglycans through a short sequence of basic residues [27, 51, 53, 54]. The exact biological role(s) of this feature is not yet clear, but may be to facilitate binding of APRIL to TACI, BCMA or other receptors by generating higher order complexes [54].
Since APRIL knockouts lack a profound phenotype, the biological functions of APRIL are only now being explored in detail. Given their overlapping receptor binding specificities, many functions of APRIL may prove redundant with those of BLyS, at least to the extent that unique attributes may be difficult to discern unless studied in the absence of BLyS [9, 55–57]. Nevertheless, APRIL clearly plays a modulatory role in certain aspects of B cell activation and isotype switching [36, 58]; and likely interacts with bone marrow plasma cells via BCMA [59, 60]. Indeed, while expressed in a variety of tissues, APRIL message is seen at very high levels in both human [61] and mouse osteoclasts. The expression of this cytokine in bone may thus be vital to the support of long–lived plasma cells (LLPCs), as LLPCs reside in the BM and are reduced in mice lacking BCMA [62]. Dendritic cells were recently reported to secrete APRIL constitutively and this production is increased following stimulation of these cells with Toll Like Receptor (TLR) ligands [63]. Finally, APRIL fosters the viability or progression of some cancers [23, 26, 64, 65].
BLyS
Sequence searches also contributed to the discovery of BLyS, and the features of this cytokine were reported simultaneously by several groups [41–43, 66]. The gene encoding BLyS is located on human chromosome 13q34 and mouse chromosome 8. Although BLyS polymorphisms have been revealed in mice, these do not appear to yield functional differences. Whether this indicates redundancy with other survival mechanisms in humans, or instead indicates that null or non-functional variants are lethal or strongly selected against, is unclear. A dominant negative form of BLyS (delta BAFF) has been described resulting from a splice variant in which exon 4 is skipped [67, 68]. The resultant gene product reveals a cryptic N-glycosylation site that prevents the secretion of any trimer containing the mutant BAFF peptide(s) [69]. Consistent with a potential negative regulatory role, mice transgenic for delta BAFF expression exhibit reductions in peripheral B cell numbers and impaired humoral responses [67].
BLyS is a 285 amino acid transmembrane protein that is highly conserved, inasmuch as the chicken and duck homologs share more than 75% homology with murine and human BLyS [70–72], and recombinant human or murine BLyS have similar effects on both chicken and mouse B cells [70–73]. Several structural features distinguish BLyS from the other members in the TNF superfamily. First, it is shortened in the axis involved with trimerization, yielding a somewhat flatter molecule than other TNF ligands. In addition, BLyS can interact with Magnesium, and has an exceptionally deep and acidic cleft that in part explains its unique affinity for the comparatively basic BR3 receptor, as well as its lower affinity for TACI and BCMA when compared to APRIL.
BLyS is produced by a wide variety of cell types, including neutrophils, dendritic cells, monocytes, macrophages, and others [74]. BLyS message and furin-mediated release is upregulated by some cytokines, including interferon gamma and IL10 [75]. The furin-cleaved, soluble trimeric form is the primary effector of in vivo function, but BLyS can also exist as a 60-mers whose spectrum of receptor binding activities are expanded, especially with TACI [52]. Finally, heterotrimers of BLyS and APRIL exist, although their significance remains poorly explored [76].
BLyS Family Receptors
There are three receptors for BLyS and/or APRIL: BCMA [77], TACI [78], and BR3, the product of the previously defined bcmd locus (Bcmd/BR3) [1, 79–83]. In general, this family differs from other TNF receptors in their lack of intracellular death domains and small number (effectively one) of ligand–binding CRDs on the extracellular side. Further, the CRDs in BR3, TACI, and BCMA lack specific structural elements found in other TNF receptors that could contribute to ligand-binding specificity. These structural interactions are reviewed extensively by Bodmer et al. [44]. All are expressed among B lineage cells, and some are found on subsets of activated T cells and dendritic cells. It is thought that each receptor is expressed in the plasma membrane as a monomer, and that ligand binding induces receptor trimerization and TRAF-facilitated signal propagation. The basic features of these receptors are summarized here. More detailed considerations of the ligand-binding motifs, as well as descriptions of key contact residues and structural organization of each receptor, are reviewed elsewhere [51].
BCMA
B Cell Maturation Antigen is a 20 kDa Type III transmembrane protein that was originally identified in a human T cell lymphoma [77, 84]. It is encoded on chromosomes 16p13.1 and 16 in humans and mice, respectively. BCMA can bind both BLyS and APRIL, but the considerably higher affinity for APRIL suggests this may be the most relevant ligand in vivo. Structurally, BCMA has the least homology to other TNF receptor family members. Like BR3, BCMA contains a single characteristic CRD motif. Indeed, among the TNF family of receptors, only BCMA and BR3 exhibit this feature. BCMA can interact with TRAF 1, 2, and 3; and downstream effects include activation of NF-κB transcriptional mediators and JNK kinases [85].
The role played by BCMA in B cell biology remains somewhat enigmatic. BCMA is expressed in neither developing nor primary B cells, and BCMA knockout mice show no phenotypic abnormalities within the pre-immune B cell compartments. Further, the abnormalities seen in the primary B cell pools of other BLyS receptor knockouts are BCMA independent. More recently, BCMA expression has been confirmed on LLPC populations, and detailed studies of BCMA knockouts support a role for the generation and maintenance of these cells (see below).
TACI
TACI is a 293 amino acid Type III transmembrane protein that was initially identified based on its ability to interact with CAML [85]. The TACI gene is found on human chromosome 17p11.2 and on murine chromosome 11. Similar to BCMA, TACI can bind both BLyS and APRIL but, unlike BCMA, TACI binds each with similar affinities [86]. Structurally, TACI bears two CRDs, and has cytoplasmic domains capable of interacting with TRAF 2, 5, and 6. Receptor ligation activates a variety of downstream transcriptional regulatory pathways, including NFAT, AP-1 and the classical NF-κB pathway, presumably through a CAML-independent mechanism [78]. Signaling through TACI has been shown in numerous experimental and transformed cell types to result in the classical NF-κB mediated upregulation of the X-linked Inhibitor of Apoptosis Protein (XIAP), but not Bcl-2 family members [87].
TACI expression is restricted to B cells and a subset of activated T cells. Although TACI can bind both BLyS and APRIL with comparatively strong affinity, the biological effects of TACI ligation remain unclear and somewhat controversial. Early results from TACI knockout mice suggested a negative role for TACI [88], because these animals exhibited B cell hyperplasia and humoral autoimmune symptoms including glomerulonephritis, proteinuria, and anti-dsDNA antibodies. More recently, there has been accumulating evidence that TACI is a positive regulator of B cell survival. For example, APRIL stimulation of primary B cells ex vivo, which should engage only TACI, improves survival [89]. Further evidence for positive regulatory roles in survival or differentiation stems from findings that show TACI facilitates isotype switch recombination, particularly to gut-associated IgA isoforms [90, 91]. It is unclear whether this dichotomy reflects opposing roles for TACI in different cellular and anatomic contexts, or instead indicates that the B cell hyperplastic phenotype in knockouts is a secondary phenomenon.
BR3/BAFFr
BLyS receptor 3 is the most recent addition to the BLyS family receptors. Its discovery resulted from results suggesting that neither TACI nor BCMA dramatically influenced B cell activities, implying the existence of an additional BLyS-binding receptor. Similar to the discovery of BLyS, BR3 was reported simultaneously by several groups [85]. Importantly, it proved to be the gene product of a locus that had previously been shown to govern late primary B cell differentiation and mature B cell survival [81]. BR3-BLyS interactions have proven critical to the maintenance of primary B cells, as shown clearly in knockout and mutant mice [82, 92]. Structurally, BR3 contains one extracellular CRD that interacts with ligand and a single intracellular recognition site for TRAF3. Recent evidence suggests that BR3-mediated interactions with TRAF3 initiate and sustain downstream signaling [37]. These signaling targets include non-classical NF-κB pathway activation and subsequent upregulation of multiple Bcl-2 family members (detailed below). This strategy contrasts the survival genes regulated by TACI, in that Bcl-2 family members inhibit apoptosis upstream of the mitochondria.
BLyS Family Actions and Mechanisms in Pre-immune B Cell Pools
B Lymphocyte Stimulator family members play central roles in regulating the behavior of most, if not all mature B cell subsets. Accordingly, a brief overview of B cell subsets and their generation is provided, followed by in-depth considerations of how members of the BLyS and BLyS receptor family influence these processes and populations. The features of pre-immune pools and the role of BLyS family members are treated first; followed by a corresponding discussion of antigen-experienced pools. Table 2 lists pre-immune B cell subsets and their surface phenotypes, as well as their BLyS binding, BLyS receptor expression and BLyS response characteristics.
Primary B Cell Development and Subsets
B cells are generated throughout life from stem cells in fetal liver, neonatal spleen, or adult BM. Multi-potent cells arise from these progenitors and, upon initiation of lymphoid gene expression, yield B lineage precursors. Excellent, highly detailed reviews of early B cell development are available elsewhere [93–97]. Briefly, immunoglobulin heavy and light chain gene rearrangements occur as cells transit the BM pro- and pre-B stages, respectively. These genetic events culminate in the production of an immunoglobulin heavy/light chain pair that is assembled and, in conjunction with the Ig-alpha and Ig-beta signaling molecules, form a functional BCR that is transported to the cell surface. BCR surface expression marks the progress into the Immature (IMM) BM subset. These newly formed B cells continue their maturation after they migrate to the periphery, passing through several so-called transitional (TR) developmental stages before entrance into primary B cell pools. Cells completing these differentiative processes enter one of the major pre-immune subsets: the follicular (FO) and marginal zone (MZ) populations.
Primary B Cell Selection and Homeostasis Are Coupled Processes
While details of primary B cell differentiation are extensively reviewed elsewhere [98, 99], a feature critical to the concepts discussed here is that stringent selection, based on BCR specificity, occurs within the IMM and TR pools. This selection transpires at both the IMM and TR stages. Indeed, only about 10% of the IMM cells generated live to exit the BM, and fewer than half of the resulting TR cells survive to finally join mature pre-immune B cell pools [100, 101]. Both negative and positive selection events contribute to these cell losses.
Negative selection—the elimination of potentially autoreactive B cell clones—occurs at both the IMM and TR stages. Current thought holds that elimination at the IMM stage is engendered by high avidity BCR engagement, whereas cells undergoing less avid but sustained BCR interactions persist into the TR stages, where they then die before maturation [102]. Substantial evidence from transgenic mouse systems confirms that autoreactive clones are eliminated at both of these stages [103–105]. More recently this has been confirmed in humans by studies that revealed similar selection against autoreactive BCR specificities during TR maturation [106]. Moreover, these cell losses failed to occur in autoimmune patients [107].
Positive selection—the preferential survival of clones whose receptors meet minimum so-called “tonic” BCR signaling requirements—occurs at these stages as well, particularly among TR cells [108–111]. Moreover, mature primary B cells continue to require continuous sub-threshold BCR signals for survival, since conditional ablation of either the BCR itself or proximal BCR signaling components leads to the rapid death of most mature B cells [112]. The requisite for persistent, low-level BCR signaling is consistent with the observation that BCR engagement has a clear positive effect in cellular survival at the BM/periphery interface and beyond [113]. The consequence of this effect is best illustrated in mixed BM chimeras, where the mature B cell pool is derived from differing mixtures of BCR transgenic or other genetically manipulated B lineage progenitors [114–117]. These experiments show a distinct connection between BCR specificity and selection in conditions of interclonal competition.
An overarching conclusion arising from such experiments is that in the TR subsets and beyond, BCR specificity determines a cell’s relative fitness to capture or utilize limited trophic resources for survival [115, 116]. This in turn suggests that while a range of tonic BCR signaling can afford survival, those cells with optimal tonic BCR signaling levels will be more likely to survive to maturity and will persist longer in per-immune pools. Considered together, these concepts unify specificity-based selective processes with homeostatic control, because B cell production rate and lifespan are the determinants of the primary pool’s size. Further, the prediction that limiting trophic factor(s) would be the basis for homeostatic control foreshadowed the discovery of BLyS as the biological metric for “space” in primary B cell compartments.
BLyS Receptor Expression and Physiology in Developing and Primary B Cells
Extensive evidence links signals via BLyS receptors, particularly BR3, with the survival of TR, FO, and MZ B cells. Indeed, BR3 and TACI are first expressed among IMM B cells in the BM, and their expression continues to increase as cells transit through TR stages. Within pre-immune FO and MZ pools, BR3 and TACI reach relatively high and constant levels [118]. MZ B cells are highly sensitive to conditions when BLyS is limiting, likely related to the uniformly high levels of TACI and BR3 expressed [83, 119].
B Lymphocyte Stimulator signaling through BR3 is crucial for the survival of all pre-immune B cell subsets from the TR stage onward. This was initially evidenced by the profound mature B cell deficiency in the A/WySnJ mouse [79–82, 120, 121], a feature that segregated as a single autosomal trait reflecting a 400 bp insertion in the intracellular tail of BR3 [83]. This and subsequent studies have led to the notion that BLyS signaling via BR3 controls the size and composition of pre-immune B cell pools (reviewed in [122]). For example, either ectopic BLyS expression or exogenous BLyS administration yields B cell hyperplasia and humoral autoimmune manifestations [42, 118, 123]. Conversely, both BLyS and BR3 knockout mice display primary B cell deficiencies reminiscent of the A/WySnJ [82, 92].
Thus, the BLyS/BR3 axis is central to the homeostatic control of primary B cell numbers, with current thought favoring the notion that available BLyS levels determine both the proportion of TR cells that survive to enter the mature pools and the lifespan of mature B cells themselves (see [124] for reviews). By controlling the entry rate and lifespan of mature B cells, a set-point for steady-state numbers is thus imposed, whereby BLyS availability constitutes the “limiting trophic resource” for which cells in these primary pools compete. This prompts a conceptual model whereby TR, FO, and MZ B cells continuously compete for BLyS consumption (binding), such that when consumption and availability are balanced, the set-point for steady-state numbers is achieved [122].
The ability to vary the survival of TR B cells has obvious implications regarding negative selection, since it implies that the thresholds for negative and positive selection at the TR stage may vary, based on available BLyS and the tonic BCR signaling capabilities within the competing cohort of emerging cells. Recent studies in several transgenic systems verify this plasticity in TR selection stringency [125–127], showing that when BLyS is non-limiting, self-reactive clonotypes that normally die at the TR stage instead survive and mature. Importantly, BLyS over expression does not rescue cells deleted at the IMM stages, suggesting that a developmental switch affords the ability for BR3 and BCR to cross-regulate one another’s signaling outcomes. This may reflect the onset of BLyS receptor expression per se, but a growing literature suggests that differentiation-dependent changes in intracellular signaling systems that afford cross-talk between the BCR and BR3 may also play a role [128–132].
BLyS and BCR Signals Are Integrated in Primary B Cells
The BCR and BR3 receptors utilize different downstream mediators to generate and propagate their respective signaling cascades. The BCR relies on a combination of Src and tyrosine kinases activated upon receptor oligomerization to initiate primary and secondary message systems. Key among these processes is the recruitment of a primary signalasome that leads to Ca2+ sensitive responses, recruitment of the MALT/Bcl-10/CARMA complex, and classical NF-κB activation [133]. In contrast, BR3 signaling relies on modulation of TRAF 3, allowing NF-κB inducing kinase (NIK) to activate non-classical NF-κB pathway kinases resulting in p100 degradation to p52. At the most fundamental level, it appears that each receptor exerts differential effects through the independent engagement of different NF-κB systems. However, downstream integration seems to be a likely outcome given that p100, the major substrate for BR3 signaling, is a target of classical pathway [134]. Therefore, we review some of the important findings regarding BR3-mediated NF-κB activation.
The NF-κB family of transcription factors is activated downstream of multiple receptors expressed on or within B cells, including the BCR, TLRs and TNF receptors [135]. Currently, two NF-κB pathways have been described: the classical pathway and the non-classical pathway. While each pathway differs based on surface receptors utilized and kinetics of activation, they are similar with respect to their general mechanism of activation, which involves the removal of inhibitory domains that retain transcriptionally active dimers in the cytoplasm [136]. Typically, the genes targeted by NF-κB dimers play important roles in survival, and include Bcl-2 family members [83, 137]. Comparing the signaling properties of B cells from various BLyS receptor knockout mice reveals that TACI exclusively activates the classical pathway [138, 139]. Little debate exists regarding the role of BR3 in non-classical NF-κB activation [140], and striking phenotypic similarities exist between NF-κB2/p100 and BLyS/BR3 knock-out mice in regards to B cell development [141]. Similar to what is observed in A/WySnJ or BR3−/− mice, p100 deficient mice exhibit defects in germinal center (GC) formation, as well as humoral responses to T-independent (TI) and T-dependent (TD) antigens [142–144]. Consistent with these findings, expression of a non-processible form of p100 blocked B cell maturation beyond the TR stages [145]. Together, these results suggested that a functional BR3 receptor is critical for p100 processing and sustained B cell survival.
TRAF3 regulates BR3-initiated non-classical NF-κB activation. The link between TRAF3 and p100 processing consists of several kinases activated in a linear sequence of steps. The degradation of TRAF3 allows NIK stabilization, IKKα phosphorylation, and removal of the inhibitory domain on p100 by partial proteasomal degradation [146]. Conditional deletion of TRAF3 results in unrestricted p100 processing and enhancement of BLyS-mediated survival [37, 147]. This observation has led to a model whereby BR3 sequesters or directly causes degradation of TRAF3 to allow NIK stabilization and non-classical pathway initiation [148–150].
While NF-κB and associated downstream survival molecules are one result of BR3 signaling, other means of regulating cell fitness have been described for BLyS signaling as well. For instance, BLyS can influence survival through increased AKT, the mammalian target of rapamycin, thus varying Pim2 levels via phosphoinositide 3-kinase phosphorylation [151]. Ultimately, activation of this pathway leads to upregulation of the anti-apoptotic protein Myeloid Cell Leukemia 1, as well as an increase in overall metabolic function. BR3 signaling also alters the intracellular localization of Protein Kinase C-δ, blocking nuclear entry and cell death [152].
Accumulating evidence suggests that cross-talk between the downstream intracellular mediators of BCR and BLyS signaling are likely involved, with NF-κB transcriptional regulators playing a key role. First, the expression of a functional BCR is not solely sufficient to allow mature B cells to survive, as evidenced in BR3 deficient B cells. Likewise, the expression of BR3 alone does not allow for the survival of mature B cells in which the BCR has been conditionally deleted [112]. Coupled with the observations that BCR signaling activates the classical NF-κB pathway, NF-κB/p100 transcription is a target of classical pathway activation, and that BR3 uniquely activates p100 processing, the mechanisms underlying this dual receptor expression requirement may be resolved in the near future.
BLyS Family Actions in Antigen-Experienced B Cell Subsets
Although less extensively studied than pre-immune subsets, an increasing body of literature indicates that BLyS receptors and cytokines also play a role in the development and maintenance of antigen-experienced cells. Conceptually, the independence of antigen-experienced cells—in terms of homeostatic constraints governing primary B cells—is self-evident. First, activated B cell clones must expand substantially upon antigen challenge to mediate an effective humoral response—requiring freedom from the competitive constraints active among quiescent primary cells. In addition, both memory and plasma cells must persist indefinitely to afford protective immunity—implying lengthier cellular lifespan than seen in pre-immune pools. In accord with this general principle, the BLyS-BR3 axis plays a less critical role in most antigen-experienced pools. Instead, other receptor ligand pairs from within the BLyS family—and hence different downstream signaling systems—dominate. Overall, this leads to a model whereby various exogenous activation and differentiative cues, including those delivered through BCR, TLR, or CD40 engagement, engender characteristic BLyS receptor profiles. Due to the different ligand preferences for each BLyS family receptor, as well as their disparate signaling outcomes, acquiring novel BLyS receptor profiles will specify independent or overlapping homeostatic niches [25, 34]. Here we will first discuss the generation and characteristics of antigen-experienced B cell pools, followed by a discussion of how these cells interact with BLyS and APRIL.
The Generation and Identity of Antigen-Experienced B Cell Subsets
The clonal daughters of naïve B cells recruited into immune responses comprise antigen-experienced subsets. These are summarized in Table 3 and include the responding B cells at various stages of activation, as well as antibody-forming cells (AFCs) and memory cells. B cell responses are generally categorized as either TD or TI, depending on whether cognate T cell help is involved in initiating the response. TD responses arise from FO B cells and are elicited by protein antigens. Following receptor ligation and internalization, such antigens can be processed to short peptides that are complexed with and presented in the context of MHC class II molecules, facilitating T cell co-stimulation. A distinguishing feature of TD responses is the GC reaction, in which B cells undergo class switch recombination (CSR) and somatic hypermutation (SHM), as well as the specificity-based selection that underlies affinity maturation. Descendants of cells in the GC reaction include LLPCs that home to and reside in the BM; as well as memory B cells, whose exact homing and localization characteristics remain less clear. In contrast, TI responses do not involve T cell help and do not yield a GC reaction. Accordingly, they lack affinity maturation, do not yield long-lived plasma cells, and produce little if any humoral memory. Two types of TI antigens exist: TI-1 antigens induce proliferation and differentiation through the stimulation of pattern recognition receptors such as TLRs; whereas TI-2 antigens bear densely repeating epitopes, and are thought to yield activation via exceptionally strong and sustained BCR cross-linking. Both TI-1 and TI-2 responses preferentially arise from MZ and B1 B cells.
Reflecting these two major modes of activation, the participation and chronologic appearance of antigen-experienced subsets follow two general patterns. Both TD and TI responses yield an initial proliferative burst among the activated primary B cell clones. These rapidly expanding cells quickly give rise to short-lived plasma cells (SLPCs), which generate the initial IgM antibodies observed during early primary responses, and in some cases modest amounts of some IgG subclasses. Following these events, most TI responses dissipate within an 8–14 day course.
During TD responses, the early generation of SLPCs is paralleled by the initiation of a GC reaction. Within the GC, two populations emerge: one that is rapidly proliferating and one that is less mitotically active. These are polarized within the GC, and correspond to the historically named “dark zone” and “light zone”, respectively. CSR and SHM are initiated in the GC, following the upregulation of enzymatic machinery associated with these processes. In toto, these events result in the generation of B cells that have switched heavy chain Ig isotypes, and that have accumulated point mutations in their BCR combining site. Those daughter clones bearing novel specificities generated by SHM then undergo selection against either low antigen affinity or self-reactivity, and for higher affinity. The mechanisms underlying this selection remain debated, as do the compartmentalization and trafficking patterns of cells in the GC [153–155]. Regardless of exact mechanism, cells surviving GC selection give rise to LLPC that migrate to and reside in the BM, and to memory B cells. While the phenotypic characteristics of these cells remain debated, their respective roles in immunity are well documented [156–158]. The LLPCs provide ongoing protection from reinfection through continuous high affinity antibody secretion, and are the source of standing antibody titers associated with protective adaptive immunity. Memory B cells provide a long-lived population of expanded, antigen specific cells that are quickly activated upon subsequent antigen encounters, affording the rapid response time associated with anamnestic responses.
BLyS Receptor Profiles Change with TI Activation
Following encounter with either TI-1 or TI-2 antigens, B cells dramatically up-regulate TACI [89]. Moreover, TACI expression increases the number of AFCs generated in TI-2 responses [159]. The role played by TACI in these rapidly dividing but relatively short-lived cells remains puzzling, but recent findings have suggested it is involved in controlling entry and exit from cell cycle [159]. A potential role for BLyS in cell cycle entry and success has been posited, although the receptor involved was not clear from these studies [160]. TACI may be directly promoting survival of short-lived AFCs, particularly when bound by multimeric ligand arrays such as those afforded by oligomerized BLyS or surface bound APRIL [52]. SLPCs generated during either TI or TD responses also express high levels of TACI, in contrast to the phenotype of LLPCS (see below) which instead upregulate BCMA. The difference in which receptor is upregulated in each response may be key to the difference in lifespan.
Information is sparse regarding the signaling systems and downstream targets of TACI in B cells activated by TI antigens. Nonetheless, these likely involve the activation of classical NF-κB signals; and recent findings suggest that—in contrast to the pre-mitochondrial survival mechanisms afforded by BR3 signals, post-mitochondrial anti-apoptoic mechanisms, including stress response proteins such as XIAP, may be involved. Since such mechanisms are effective in preventing apoptosis engendered by cell cycle checkpoint failures or unfolded protein responses, these pro-survival mediators might be expected for B cells undergoing extensive division and differentiation to high levels of Ig protein production [161].
BLyS Family Signaling in the Germinal Center Reaction
The striking parallels between GC and primary B cell differentiation, in terms of negative and positive selection processes based on BCR specificity, suggest mechanistically similar processes. Substantial evidence suggests that members of the BLyS family may indeed play a role in the establishment and proper evolution of the GC reaction, but the members involved and mechanisms of action remain obscure. In general, BCR engagement per se, as well as concomitant CD40 ligation, leads to a BLyS receptor signature in which BR3 is upregulated [113]. Alternatively, Qian et al. showed that Act 1 functions as a negative regulator of both BR3 and CD40, suggesting potential cross talk and cross-modulation between different TNF family receptors expressed in the GC [162]. TACI is elevated following TD stimulation as well, although not to the extent seen with TI antigens. Interestingly, GC B cells retain this phenotype, suggesting that the BLyS-BR3 interactions may play a role in this aspect of TD responses. However, early studies in the BR3 mutant A/WySnJ mouse strain suggested GCs likely form, since TD memory could be generated—albeit with reduced efficacy and less extensive isotype switching [80]. In agreement with this, more recent work from the Manser laboratory showed that BR3 is not required for the generation of GCs [143], but plays a role in the appropriate evolution of the overall GC reaction. This finding was extended in studies that used soluble BCMA-Fc to block both APRIL and BLyS, yielding the same results [143, 163]. Thus, while not profoundly affected, GCs generated in the absence of appropriate BLyS or BR3 signaling are small, comparatively short-lived, and fewer in number. In addition to possible B cell intrinsic activities for BLyS in the GC reaction, follicular dendritic cells (FDC) networks do not mature in GCs when BLyS and APRIL are blocked [164]. This may reflect a loss of FDC supporting signals secondary to the decreased number of B cells in the GC, although the exact basis remains speculative.
The potential roles of BLyS family members in the selective processes that follow SHM within GCs have not been directly interrogated. This in part reflects the complexity associated with blocking BLyS-mediated signals necessary for primary cell survival, as any effects on GC selection might indirectly result from a reduction in primary B cells that seed the GC. In addition, analysis of negative and positive B cell selection in the GC requires single cell analyses, exacerbating the difficulty of this problem. Nonetheless, direct assessments of these possibilities are likely forthcoming.
BLyS Family Activities in LLPC and Memory B Cells
The BLyS receptor expression profiles and cytokine requirements for LLPC and memory B cells are unique. Recent evidence supports a role for BCMA in maintaining LLPC survival [62]. Inasmuch as this receptor binds both BLyS and APRIL, LLPC are sensitive to simultaneous withdrawal of both cytokines BLyS and APRIL [60]. The ability of at least some LLPCs to use APRIL as a survival factor is intriguing, since osteoclasts generate large amounts of this cytokine, and might thus provide localized high levels of APRIL in specialized BM niches. Further, APRIL exhibits the ability to bind proteoglycans [54], which are abundantly expressed on resident BM accessory cells where LLPCs co-exist. The downstream effects of BCMA signaling in LLPCs are not yet extensively characterized, as these are rare populations of B cells that are difficult to maintain in vitro. However, B cell and plasma cell lines reveal classical NF-κB activation, the upregulation of a number of proteins associated with antigen presentation and co-stimulation, and increased IL-2 production following BCMA ligation [139].
In contrast to LLPCs, memory B cells express elevated levels of TACI, at least early in their generation [60]. Despite this, most evidence points to a lack of reliance on either BLyS or APRIL [60]. Further supporting BLyS independence, anti-BLyS antibody failed to eliminate B cell memory in a variety of experimental systems [165].
BLyS Family Receptors and Cytokines in Human Health and Disease
Given the central roles of BLyS family members in primary and antigen-experienced B cell subsets, it is not surprising that strong relationships have been revealed for a panoply of disease states, particularly in autoimmunity and cancer. These associations may provide clues to fundamental disease mechanisms, as well as potential diagnostic and therapeutic targets. Further, as our understanding of the BLyS family’s actions expands, novel approaches to the manipulation of B cell immunity and tolerance may result, particularly in vaccine development and transplantation tolerance (Reviewed in [166]).
Elevated serum BLyS concentrations are associated with several autoimmune syndromes, particularly Systemic Lupus Erythematosis, Rheumatoid Arthritis, and Sjogrens syndrome [167–170]. Whether these are causal links is yet unclear, and their basis remains the target of intense investigation. The rescue of potentially autoreactive B cells during TR differentiation, a known outcome of elevated BLyS levels, may prove an underlying mechanistic connection [127].
In contrast to pathologic increases in BLyS receptor signaling, a TACI mutation that blocks ligand binding underlies the Ig secretion defect in patients with common variable immune disease (CVID) [171]. Given that cells expressing large amounts of Ig often express TACI, it is conceivable that the CVID phenotype results from an inability of these cells to capture BLyS or APRIL as a pro-survival function. BLyS receptor signaling proteins such as TRAF3 and other downstream modulators are becoming appreciated as tumor suppressors frequently inactivated in multiple myelomas. [172, 173].
Current therapy for lymphoma includes ablation of the B cell compartment with a B cell targeting antibody, Rituxan ®. BLyS-targeting therapeutics may achieve similar results alone or in combination with ablation. Further regulation of serum BLyS may be beneficial in assuring that reconstitution of the immune system following ablative therapy occurs in a regulated manner, thus avoiding pitfalls that might occur with excess BLyS.
Summary
The BLyS family of receptors and ligands forms a major axis in the homeostatic control of both pre-immune and antigen-experienced B cells. These interactions are most extensively studied in the pre-immune B cell subsets, where BLyS signals via the BR3 receptor modulate the survival of TR and mature B cells. These processes involve downstream cross talk between BR3 and the BCR, influencing the selection of autoreactive specificities. An understanding of BLyS family actions in antigen-experienced subsets, including activated, memory, and plasma cells, is only now beginning to emerge. These cells display shifts in BLyS family receptor expression, suggesting they display different BLyS family ligand requisites than their pre-immune progenitors. As our understanding of these relationships and their related intracellular signaling systems advances, novel molecular targets for manipulation of the humoral immune system should emerge.
References
Schiemann, B., Gommerman, J. L., Vora, K., Cachero, T. G., Shulga-Morskaya, S., Dobles, M., et al. (2001). An essential role for BAFF in the normal development of B cells through a BCMA-independent pathway. Science, 293, 2111–2114.
Schneider, P., Takatsuka, H., Wilson, A., Mackay, F., Tardivel, A., Lens, S., et al. (2001). Maturation of marginal zone and follicular B cells requires B cell activating factor of the tumor necrosis factor family and is independent of B cell maturation antigen. Journal of Experimental Medicine, 194, 1691–1697.
Thompson, J. S., Schneider, P., Kalled, S. L., Wang, L., Lefevre, E. A., Cachero, T. G., et al. (2000). BAFF binds to the tumor necrosis factor receptor-like molecule B cell maturation antigen and is important for maintaining the peripheral B cell population. Journal of Experimental Medicine, 192, 129–135.
Casola, S., Otipoby, K. L., Alimzhanov, M., Humme, S., Uyttersprot, N., Kutok, J. L., et al. (2004). B cell receptor signal strength determines B cell fate. Nature Immunology, 5, 317–327.
Do, R. K., & Chen-Kiang, S. (2002). Mechanism of BLyS action in B cell immunity. Cytokine and Growth Factor Reviews, 13, 19–25.
Ambrose, C. M. (2002). Baff-R. Journal of Biological Regulators and Homeostatic Agents, 16, 211–213.
Defrance, T., Casamayor-Palleja, M., & Krammer, P. H. (2002). The life and death of a B cell. Advances in Cancer Research, 86, 195–225.
Laabi, Y., Egle, A., & Strasser, A. (2001). TNF cytokine family: More BAFF-ling complexities. Current Biology, 11, R1013–R1016.
Mackay, F., & Ambrose, C. (2003). The TNF family members BAFF and APRIL: The growing complexity. Cytokine and Growth Factor Reviews, 14, 311–324.
Mackay, F., & Browning, J. L. (2002). BAFF: A fundamental survival factor for B cells. Nature Reviews Immunology, 2, 465–475.
Kalled, S. L. (2002). BAFF: A novel therapeutic target for autoimmunity. Current Opinion in Investigational Drugs, 3, 1005–1010.
Mak, T. W., & Yeh, W. C. (2002). Signaling for survival and apoptosis in the immune system. Arthritis Research, 3(4 Suppl), S243–S252.
Nardelli, B., Moore, P. A., Li, Y., & Hilbert, D. M. (2002). B lymphocyte stimulator (BLyS): A therapeutic trichotomy for the treatment of B lymphocyte diseases. Leukaemia & Lymphoma, 43, 1367–1373.
Stohl, W. (2002). B lymphocyte stimulator protein levels in systemic lupus erythematosus and other diseases. Current Rheumatology Reports, 4, 345–350.
Carter, R. H. (2003). A role for BLyS in tissue inflammation? Arthritis and Rheumatism, 48, 882–885.
Harless Smith, S., & Cancro, M. P. (2003). Integrating B cell homeostasis and selection with BLyS. Archivum Immunologiae et Therapiae Experimentalis (Warsz), 51, 209–218.
Harless Smith, S., & Cancro, M. P. (2003). BLyS: The pivotal determinant of peripheral B cell selection and lifespan. Current Pharmaceutical Design, 9, 1833–1847.
Mackay, F., Schneider, P., Rennert, P., & Browning, J. (2003). BAFF AND APRIL: A tutorial on B cell survival. Annual Review of Immunology, 21, 231–264.
Melchers, F. (2003). Actions of BAFF in B cell maturation and its effects on the development of autoimmune disease. Annals of the Rheumatic Diseases, 62(Suppl 2), ii25–ii27.
Schneider, P., & Tschopp, J. (2003). BAFF and the regulation of B cell survival. Immunology Letters, 88, 57–62.
Cancro, M. P. (2004). The BLyS family of ligands and receptors: An archetype for niche-specific homeostatic regulation. Immunological Reviews, 202, 237–249.
Gavin, A., Ait-Azzouzene, D., Martensson, A., Duong, B., Verkoczy, L., Skog, J. L., et al. (2004). Peripheral B lymphocyte tolerance. Keio Journal of Medicine, 53, 151–158.
Mackay, F., & Tangye, S. G. (2004). The role of the BAFF/APRIL system in B cell homeostasis and lymphoid cancers. Current Opinion in Pharmacology, 4, 347–354.
Anolik, J. H., & Aringer, M. (2005). New treatments for SLE: Cell-depleting and anti-cytokine therapies. Best Practice & Research. Clinical Rheumatology, 19, 859–878.
Crowley, J. E., Treml, L. S., Stadanlick, J. E., Carpenter, E., & Cancro, M. P. (2005). Homeostatic niche specification among naive and activated B cells: A growing role for the BLyS family of receptors and ligands. Seminars in Immunology, 17, 193–199.
Jelinek, D. F., & Darce, J. R. (2005). Human B lymphocyte malignancies: Exploitation of BLyS and APRIL and their receptors. Current Directions in Autoimmunity, 8, 266–288.
Kalled, S. L., Ambrose, C., & Hsu, Y. M. (2005). The biochemistry and biology of BAFF, APRIL and their receptors. Current Directions in Autoimmunity, 8, 206–242.
Mackay, F., Sierro, F., Grey, S. T., & Gordon, T. P. (2005). The BAFF/APRIL system: An important player in systemic rheumatic diseases. Current Directions in Autoimmunity, 8, 243–265.
Noelle, R. J., & Erickson, L. D. (2005). Determinations of B cell fate in immunity and autoimmunity. Current Directions in Autoimmunity, 8, 1–24.
Schneider, P. (2005). The role of APRIL and BAFF in lymphocyte activation. Current Opinion in Immunology, 17, 282–289.
Brink, R. (2006). Regulation of B cell self-tolerance by BAFF. Seminars in Immunology, 18, 276–283.
Mackay, F., & Leung, H. (2006). The role of the BAFF/APRIL system on T cell function. Seminars in Immunology, 18, 284–289.
Kalled, S. L. (2006). Impact of the BAFF/BR3 axis on B cell survival, germinal center maintenance and antibody production. Seminars in Immunology, 18, 290–296.
Treml, L. S., Crowley, J. E., & Cancro, M. P. (2006). BLyS receptor signatures resolve homeostatically independent compartments among naive and antigen-experienced B cells. Seminars in Immunology, 18, 297–304.
Mackay, F., Silveira, P. A., & Brink, R. (2007). B cells and the BAFF/APRIL axis: Fast-forward on autoimmunity and signaling. Current Opinion in Immunology, 19, 327–336.
Tangye, S. G., Bryant, V. L., Cuss, A. K., & Good, K. L. (2006). BAFF, APRIL and human B cell disorders. Seminars in Immunology, 18, 305–317.
Gardam, S., Sierro, F., Basten, A., Mackay, F., & Brink, R. (2008). TRAF2 and TRAF3 signal adapters act cooperatively to control the maturation and survival signals delivered to B cells by the BAFF receptor. Immunity, 28, 391–401.
Grech, A. P., Amesbury, M., Chan, T., Gardam, S., Basten, A., & Brink, R. (2004). TRAF2 differentially regulates the canonical and noncanonical pathways of NF-kappaB activation in mature B cells. Immunity, 21, 629–642.
Do, R. K., Hatada, E., Lee, H., Tourigny, M. R., Hilbert, D., & Chen-Kiang, S. (2000). Attenuation of apoptosis underlies B lymphocyte stimulator enhancement of humoral immune response. Journal of Experimental Medicine, 192, 953–964.
Locksley, R. M., Killeen, N., & Lenardo, M. J. (2001). The TNF and TNF receptor superfamilies: Integrating mammalian biology. Cell, 104, 487–501.
Mackay, F., Woodcock, S. A., Lawton, P., Ambrose, C., Baetscher, M., Schneider, P., et al. (1999). Mice transgenic for BAFF develop lymphocytic disorders along with autoimmune manifestations. Journal of Experimental Medicine, 190, 1697–1710.
Moore, P. A., Belvedere, O., Orr, A., Pieri, K., LaFleur, D. W., Feng, P., et al. (1999). BLyS: Member of the tumor necrosis factor family and B lymphocyte stimulator. Science, 285, 260–263.
Schneider, P., MacKay, F., Steiner, V., Hofmann, K., Bodmer, J. L., Holler, N., et al. (1999). BAFF, a novel ligand of the tumor necrosis factor family, stimulates B cell growth. Journal of Experimental Medicine, 189, 1747–1756.
Bodmer, J. L., Schneider, P., & Tschopp, J. (2002). The molecular architecture of the TNF superfamily. Trends in Biochemical Sciences, 27, 19–26.
Hahne, M., Kataoka, T., Schroter, M., Hofmann, K., Irmler, M., Bodmer, J. L., et al. (1998). APRIL, a new ligand of the tumor necrosis factor family, stimulates tumor cell growth. Journal of Experimental Medicine, 188, 1185–1190.
Kelly, K., Manos, E., Jensen, G., Nadauld, L., & Jones, D. A. (2000). APRIL/TRDL-1, a tumor necrosis factor-like ligand, stimulates cell death. Cancer Research, 60, 1021–1027.
Pradet-Balade, B., Medema, J. P., Lopez-Fraga, M., Lozano, J. C., Kolfschoten, G. M., Picard, A., et al. (2002). An endogenous hybrid mRNA encodes TWE-PRIL, a functional cell surface TWEAK-APRIL fusion protein. EMBO Journal, 21, 5711–5720.
Kolfschoten, G. M., Pradet-Balade, B., Hahne, M., & Medema, J. P. (2003). TWE-PRIL; a fusion protein of TWEAK and APRIL. Biochemical Pharmacology, 66, 1427–1432.
Wallweber, H. J., Compaan, D. M., Starovasnik, M. A., & Hymowitz, S. G. (2004). The crystal structure of a proliferation-inducing ligand, APRIL. Journal of Molecular Biology, 343, 283–290.
Hymowitz, S. G., Patel, D. R., Wallweber, H. J., Runyon, S., Yan, M., Yin, J., et al. (2005). Structures of APRIL-receptor complexes: Like BCMA, TACI employs only a single cysteine-rich domain for high affinity ligand binding. Journal of Biological Chemistry, 280, 7218–7227.
Bossen, C., & Schneider, P. (2006). BAFF, APRIL and their receptors: Structure, function and signaling. Seminars in Immunology, 18, 263–275.
Bossen, C., Cachero, T. G., Tardivel, A., Ingold, K., Willen, L., Dobles, M., et al. (2008). TACI, unlike BAFF-R, is solely activated by oligomeric BAFF and APRIL to support survival of activated B cells and plasmablasts. Blood, 111, 1004–1012.
Hendriks, J., Planelles, L., de Jong-Odding, J., Hardenberg, G., Pals, S. T., Hahne, M., et al. (2005). Heparan sulfate proteoglycan binding promotes APRIL-induced tumor cell proliferation. Cell Death and Differentiation, 12, 637–648.
Ingold, K., Zumsteg, A., Tardivel, A., Huard, B., Steiner, Q. G., Cachero, T. G., et al. (2005). Identification of proteoglycans as the APRIL-specific binding partners. Journal of Experimental Medicine, 201, 1375–1383.
Khare, S. D., & Hsu, H. (2001). The role of TALL-1 and APRIL in immune regulation. Trends in Immunology, 22, 61–63.
Stein, J. V., Lopez-Fraga, M., Elustondo, F. A., Carvalho-Pinto, C. E., Rodriguez, D., Gomez-Caro, R., et al. (2002). APRIL modulates B and T cell immunity. Journal of Clinical Investigation, 109, 1587–1598.
Medema, J. P., Planelles-Carazo, L., Hardenberg, G., & Hahne, M. (2003). The uncertain glory of APRIL. Cell Death and Differentiation, 10, 1121–1125.
Sakurai, D., Hase, H., Kanno, Y., Kojima, H., Okumura, K., & Kobata, T. (2007). TACI regulates IgA production by APRIL in collaboration with HSPG. Blood, 109, 2961–2967.
Belnoue, E., Pihlgren, M., McGaha, T. L., Tougne, C., Rochat, A. F., Bossen, C., et al. (2008). APRIL is critical for plasmablast survival in the bone marrow and poorly expressed by early-life bone marrow stromal cells. Blood, 111, 2755–2764.
Benson, M. J., Dillon, S. R., Castigli, E., Geha, R. S., Xu, S., Lam, K. P., et al. (2008). Cutting edge: The dependence of plasma cells and independence of memory B cells on BAFF and APRIL. Journal of Immunology, 180, 3655–3659.
Moreaux, J., Cremer, F. W., Reme, T., Raab, M., Mahtouk, K., Kaukel, P., et al. (2005). The level of TACI gene expression in myeloma cells is associated with a signature of microenvironment dependence versus a plasmablastic signature. Blood, 106, 1021–1030.
O’Connor, B. P., Raman, V. S., Erickson, L. D., Cook, W. J., Weaver, L. K., Ahonen, C., et al. (2004). BCMA is essential for the survival of long-lived bone marrow plasma cells. Journal of Experimental Medicine, 199, 91–98.
Hardenberg, G., Planelles, L., Schwarte, C. M., van Bostelen, L., Le Huong, T., Hahne, M., et al. (2007). Specific TLR ligands regulate APRIL secretion by dendritic cells in a PKR-dependent manner. European Journal of Immunology, 37, 2900–2911.
Ware, C. F. (2000). APRIL and BAFF connect autoimmunity and cancer. Journal of Experimental Medicine, 192, F35–F38.
Chiu, A., Xu, W., He, B., Dillon, S. R., Gross, J. A., Sievers, E., et al. (2007). Hodgkin lymphoma cells express TACI and BCMA receptors and generate survival and proliferation signals in response to BAFF and APRIL. Blood, 109, 729–739.
Batten, M., Groom, J., Cachero, T. G., Qian, F., Schneider, P., Tschopp, J., et al. (2000). BAFF mediates survival of peripheral immature B lymphocytes. Journal of Experimental Medicine, 192, 1453–1466.
Gavin, A. L., Ait-Azzouzene, D., Ware, C. F., & Nemazee, D. (2003). DeltaBAFF, an alternate splice isoform that regulates receptor binding and biopresentation of the B cell survival cytokine, BAFF. Journal of Biological Chemistry, 278, 38220–38228.
Gavin, A. L., Duong, B., Skog, P., Ait-Azzouzene, D., Greaves, D. R., Scott, M. L., et al. (2005). deltaBAFF, a splice isoform of BAFF, opposes full-length BAFF activity in vivo in transgenic mouse models. Journal of Immunology, 175, 319–328.
Vogt, G., Chapgier, A., Yang, K., Chuzhanova, N., Feinberg, J., Fieschi, C., et al. (2005). Gains of glycosylation comprise an unexpectedly large group of pathogenic mutations. Nature Genetics, 37, 692–700.
Koskela, K., Nieminen, P., Kohonen, P., Salminen, H., & Lassila, O. (2004). Chicken B-cell-activating factor: Regulator of B-cell survival in the bursa of fabricius. Scandinavian Journal of Immunology, 59, 449–457.
Guan, Z. B., Ye, J. L., Dan, W. B., Yao, W. J., & Zhang, S. Q. (2007). Cloning, expression and bioactivity of duck BAFF. Molecular Immunology, 44, 1471–1476.
Schneider, K., Kothlow, S., Schneider, P., Tardivel, A., Gobel, T., Kaspers, B., et al. (2004). Chicken BAFF—a highly conserved cytokine that mediates B cell survival. International Immunology, 16, 139–148.
Kothlow, S., Morgenroth, I., Graef, Y., Schneider, K., Riehl, I., Staeheli, P., et al. (2007). Unique and conserved functions of B cell-activating factor of the TNF family (BAFF) in the chicken. International Immunology, 19, 203–215.
Scapini, P., Bazzoni, F., & Cassatella, M. A. (2008). Regulation of B-cell-activating factor (BAFF)/B lymphocyte stimulator (BLyS) expression in human neutrophils. Immunology Letters, 116, 1–6.
Nardelli, B., Belvedere, O., Roschke, V., Moore, P. A., Olsen, H. S., Migone, T. S., et al. (2001). Synthesis and release of B-lymphocyte stimulator from myeloid cells. Blood, 97, 198–204.
Roschke, V., Sosnovtseva, S., Ward, C. D., Hong, J. S., Smith, R., Albert, V., et al. (2002). BLyS and APRIL form biologically active heterotrimers that are expressed in patients with systemic immune-based rheumatic diseases. Journal of Immunology, 169, 4314–4321.
Madry, C., Laabi, Y., Callebaut, I., Roussel, J., Hatzoglou, A., Le Coniat, M., et al. (1998). The characterization of murine BCMA gene defines it as a new member of the tumor necrosis factor receptor superfamily. International Immunology, 10, 1693–1702.
von Bulow, G. U., & Bram, R. J. (1997). NF-AT activation induced by a CAML-interacting member of the tumor necrosis factor receptor superfamily. Science, 278, 138–141.
Miller, D. J., & Hayes, C. E. (1991). Phenotypic and genetic characterization of a unique B lymphocyte deficiency in strain A/WySnJ mice. European Journal of Immunology, 21, 1123–1130.
Miller, D. J., Hanson, K. D., Carman, J. A., & Hayes, C. E. (1992). A single autosomal gene defect severely limits IgG but not IgM responses in B lymphocyte-deficient A/WySnJ mice. European Journal of Immunology, 22, 373–379.
Lentz, V. M., Cancro, M. P., Nashold, F. E., & Hayes, C. E. (1996). Bcmd governs recruitment of new B cells into the stable peripheral B cell pool in the A/WySnJ mouse. Journal of Immunology, 157, 598–606.
Harless, S. M., Lentz, V. M., Sah, A. P., Hsu, B. L., Clise-Dwyer, K., Hilbert, D. M., et al. (2001). Competition for BLyS-mediated signaling through Bcmd/BR3 regulates peripheral B lymphocyte numbers. Current Biology, 11, 1986–1989.
Yan, M., Brady, J. R., Chan, B., Lee, W. P., Hsu, B., Harless, S., et al. (2001). Identification of a novel receptor for B lymphocyte stimulator that is mutated in a mouse strain with severe B cell deficiency. Current Biology, 11, 1547–1552.
Laabi, Y., Gras, M. P., Brouet, J. C., Berger, R., Larsen, C. J., & Tsapis, A. (1994). The BCMA gene, preferentially expressed during B lymphoid maturation, is bidirectionally transcribed. Nucleic Acids Research, 22, 1147–1154.
Stockfleth, E., Trefzer, U., Garcia-Bartels, C., Wegner, T., Schmook, T., & Sterry, W. (2003). The use of Toll-like receptor-7 agonist in the treatment of basal cell carcinoma: An overview. The British Journal of Dermatology, 149(Suppl 66), 53–56.
Day, E. S., Cachero, T. G., Qian, F., Sun, Y., Wen, D., Pelletier, M., et al. (2005). Selectivity of BAFF/BLyS and APRIL for binding to the TNF family receptors BAFFR/BR3 and BCMA. Biochemistry, 44, 1919–1931.
Roth, W., Wagenknecht, B., Klumpp, A., Naumann, U., Hahne, M., Tschopp, J., et al. (2001). APRIL, a new member of the tumor necrosis factor family, modulates death ligand-induced apoptosis. Cell Death and Differentiation, 8, 403–410.
Seshasayee, D., Valdez, P., Yan, M., Dixit, V. M., Tumas, D., & Grewal, I. S. (2003). Loss of TACI causes fatal lymphoproliferation and autoimmunity, establishing TACI as an inhibitory BLyS receptor. Immunity, 18, 279–288.
Treml, L. S., Carlesso, G., Hoek, K. L., Stadanlick, J. E., Kambayashi, T., Bram, R. J., et al. (2007). TLR stimulation modifies BLyS receptor expression in follicular and marginal zone B cells. Journal of Immunology, 178, 7531–7539.
Castigli, E., Scott, S., Dedeoglu, F., Bryce, P., Jabara, H., Bhan, A. K., et al. (2004). Impaired IgA class switching in APRIL-deficient mice. Proceedings of the National Academy of Sciences of the United States of America, 101, 3903–3908.
Castigli, E., Wilson, S. A., Scott, S., Dedeoglu, F., Xu, S., Lam, K. P., et al. (2005). TACI and BAFF-R mediate isotype switching in B cells. Journal of Experimental Medicine, 201, 35–39.
Shulga-Morskaya, S., Dobles, M., Walsh, M. E., Ng, L. G., MacKay, F., Rao, S. P., et al. (2004). B cell-activating factor belonging to the TNF family acts through separate receptors to support B cell survival and T cell-independent antibody formation. Journal of Immunology, 173, 2331–2341.
Hardy, R. R., Kincade, P. W., & Dorshkind, K. (2007). The protean nature of cells in the B lymphocyte lineage. Immunity, 26, 703–714.
Osmond, D. G., Rolink, A., & Melchers, F. (1998). Murine B lymphopoiesis: Towards a unified model. Immunology Today, 19, 65–68.
Burrows, P. D., Kearney, J. F., Schroeder, H. W., Jr., & Cooper, M. D. (1993). Normal B lymphocyte differentiation. Baillieres Clinical Haematology, 6, 785–806.
Singh, H. (1996). Gene targeting reveals a hierarchy of transcription factors regulating specification of lymphoid cell fates. Current Opinion in Immunology, 8, 160–165.
Georgopoulos, K. (2002). Haematopoietic cell-fate decisions, chromatin regulation and ikaros. Nature Reviews Immunology, 2, 162–174.
Cancro, M. P. (2004). Peripheral B-cell maturation: The intersection of selection and homeostasis. Immunological Reviews, 197, 89–101.
Srivastava, B., Lindsley, R. C., Nikbakht, N., & Allman, D. (2005). Models for peripheral B cell development and homeostasis. Seminars in Immunology, 17, 175–182.
Allman, D. M., Ferguson, S. E., Lentz, V. M., & Cancro, M. P. (1993). Peripheral B cell maturation. II. Heat-stable antigen(hi) splenic B cells are an immature developmental intermediate in the production of long-lived marrow-derived B cells. Journal of Immunology, 151, 4431–4444.
Osmond, D. G. (1986). Population dynamics of bone marrow B lymphocytes. Immunological Reviews, 93, 103–124.
Goodnow, C. C., Sprent, J., Fazekas de St Groth, B., & Vinuesa, C. G. (2005). Cellular and genetic mechanisms of self tolerance and autoimmunity. Nature, 435, 590–597.
Nemazee, D., & Buerki, K. (1989). Clonal deletion of autoreactive B lymphocytes in bone marrow chimeras. Proceedings of the National Academy of Sciences of the United States of America, 86, 8039–8043.
Basten, A., Brink, R., Peake, P., Adams, E., Crosbie, J., Hartley, S., et al. (1991). Self tolerance in the B-cell repertoire. Immunological Reviews, 122, 5–19.
Hartley, S. B., Crosbie, J., Brink, R., Kantor, A. B., Basten, A., & Goodnow, C. C. (1991). Elimination from peripheral lymphoid tissues of self-reactive B lymphocytes recognizing membrane-bound antigens. Nature, 353, 765–769.
Yurasov, S., Wardemann, H., Hammersen, J., Tsuiji, M., Meffre, E., Pascual, V., et al. (2005). Defective B cell tolerance checkpoints in systemic lupus erythematosus. Journal of Experimental Medicine, 201, 703–711.
Yurasov, S., Hammersen, J., Tiller, T., Tsuiji, M., & Wardemann, H. (2005). B-cell tolerance checkpoints in healthy humans and patients with systemic lupus erythematosus. Annals of the New York Academy of Sciences, 1062, 165–174.
Wang, H., Ye, J., Arnold, L. W., McCray, S. K., & Clarke, S. H. (2001). A VH12 transgenic mouse exhibits defects in pre-B cell development and is unable to make IgM + B cells. Journal of Immunology, 167, 1254–1262.
Levine, M. H., Haberman, A. M., Sant’Angelo, D. B., Hannum, L. G., Cancro, M. P., Janeway, C. A., Jr., et al. (2000). A B-cell receptor-specific selection step governs immature to mature B cell differentiation. Proceedings of the National Academy of Sciences of the United States of America, 97, 2743–2748.
Clarke, S. H., & McCray, S. K. (1993). VH CDR3-dependent positive selection of murine VH12-expressing B cells in the neonate. European Journal of Immunology, 23, 3327–3334.
Gu, H., Tarlinton, D., Muller, W., Rajewsky, K., & Forster, I. (1991). Most peripheral B cells in mice are ligand selected. Journal of Experimental Medicine, 173, 1357–1371.
Lam, K. P., Kuhn, R., & Rajewsky, K. (1997). In vivo ablation of surface immunoglobulin on mature B cells by inducible gene targeting results in rapid cell death. Cell, 90, 1073–1083.
Smith, S. H., & Cancro, M. P. (2003). Cutting edge: B cell receptor signals regulate BLyS receptor levels in mature B cells and their immediate progenitors. Journal of Immunology, 170, 5820–5823.
Rosado, M. M., & Freitas, A. A. (1998). The role of the B cell receptor V region in peripheral B cell survival. European Journal of Immunology, 28, 2685–2693.
McLean, A. R., Rosado, M. M., Agenes, F., Vasconcellos, R., & Freitas, A. A. (1997). Resource competition as a mechanism for B cell homeostasis. Proceedings of the National Academy of Sciences of the United States of America, 94, 5792–5797.
Freitas, A. A., Rosado, M. M., Viale, A. C., & Grandien, A. (1995). The role of cellular competition in B cell survival and selection of B cell repertoires. European Journal of Immunology, 25, 1729–1738.
Sprent, J., & Basten, A. (1973). Circulating T and B lymphocytes of the mouse. II. Lifespan. Cellular Immunology, 7, 40–59.
Hsu, B. L., Harless, S. M., Lindsley, R. C., Hilbert, D. M., & Cancro, M. P. (2002). Cutting edge: BLyS enables survival of transitional and mature B cells through distinct mediators. Journal of Immunology, 168, 5993–5996.
von Bulow, G. U., van Deursen, J. M., & Bram, R. J. (2001). Regulation of the T-independent humoral response by TACI. Immunity, 14, 573–582.
Lentz, V. M., Hayes, C. E., & Cancro, M. P. (1998). Bcmd decreases the life span of B-2 but not B-1 cells in A/WySnJ mice. Journal of Immunology, 160, 3743–3747.
Hoag, K. A., Clise-Dwyer, K., Lim, Y. H., Nashold, F. E., Gestwicki, J., Cancro, M. P., et al. (2000). A quantitative-trait locus controlling peripheral B-cell deficiency maps to mouse Chromosome 15. Immunogenetics, 51, 924–929.
Miller, J. P., Stadanlick, J. E., & Cancro, M. P. (2006). Space, selection, and surveillance: Setting boundaries with BLyS. Journal of Immunology, 176, 6405–6410.
Stohl, W. (2005). BlySfulness does not equal blissfulness in systemic lupus erythematosus: A therapeutic role for BLyS antagonists. Current Directions in Autoimmunity, 8, 289–304.
Cancro, M. P., & Smith, S. H. (2003). Peripheral B cell selection and homeostasis. Immunologic Research, 27, 141–148.
Lesley, R., Xu, Y., Kalled, S. L., Hess, D. M., Schwab, S. R., Shu, H. B., et al. (2004). Reduced competitiveness of autoantigen-engaged B cells due to increased dependence on BAFF. Immunity, 20, 441–453.
Thien, M., Phan, T. G., Gardam, S., Amesbury, M., Basten, A., Mackay, F., et al. (2004). Excess BAFF rescues self-reactive B cells from peripheral deletion and allows them to enter forbidden follicular and marginal zone niches. Immunity, 20, 785–798.
Hondowicz, B. D., Alexander, S. T., Quinn, W. J., 3rd, Pagan, A. J., Metzgar, M. H., & Cancro, M. P. (2007). The role of BLyS/BLyS receptors in anti-chromatin B cell regulation. International Immunology, 19(4), 465–475.
Stadanlick, J. E., & Cancro, M. P. (2008). BAFF and the plasticity of peripheral B cell tolerance. Current Opinion in Immunology, 20, 158–161.
Sasaki, Y., Derudder, E., Hobeika, E., Pelanda, R., Reth, M., Rajewsky, K., et al. (2006). Canonical NF-kappaB activity, dispensable for B cell development, replaces BAFF-receptor signals and promotes B cell proliferation upon activation. Immunity, 24, 729–739.
Shinners, N. P., Carlesso, G., Castro, I., Hoek, K. L., Corn, R. A., Woodland, R. T., et al. (2007). Bruton’s tyrosine kinase mediates NF-kappa B activation and B cell survival by B cell-activating factor receptor of the TNF-R family. Journal of Immunology, 179, 3872–3880.
Siebenlist, U., Brown, K., & Claudio, E. (2005). Control of lymphocyte development by nuclear factor-kappaB. Nature Reviews Immunology, 5, 435–445.
Stadanlick, J. E., Kaileh, M., Karnell, F. G., Scholz, J. L., Miller, J. P., & Quinn, W. J., 3rd. (2008). Tonic B cell antigen receptor signals supply an NF-kB substrate for prosurvival signaling. Nature Immunology, 9, 1379–1387.
Shinohara, H., Yasuda, T., Aiba, Y., Sanjo, H., Hamadate, M., Watarai, H., et al. (2005). PKC{beta} regulates BCR-mediated IKK activation by facilitating the interaction between TAK1 and CARMA1. Journal of Experimental Medicine, 202, 1423–1431.
Weih, F., Durham, S. K., Barton, D. S., Sha, W. C., Baltimore, D., & Bravo, R. (1997). p50-NF-kappaB complexes partially compensate for the absence of RelB: Severely increased pathology in p50(-/-)relB(-/-) double-knockout mice. Journal of Experimental Medicine, 185, 1359–1370.
Hayden, M. S., & Ghosh, S. (2004). Signaling to NF-kappaB. Genes and Development, 18, 2195–2224.
Bonizzi, G., & Karin, M. (2004). The two NF-kappaB activation pathways and their role in innate and adaptive immunity. Trends in Immunology, 25, 280–288.
Sen, R. (2006). Control of B lymphocyte apoptosis by the transcription factor NF-kappaB. Immunity, 25, 871–883.
Enzler, T., Bonizzi, G., Silverman, G. J., Otero, D. C., Widhopf, G. F., Anzelon-Mills, A., et al. (2006). Alternative and classical NF-kappa B signaling retain autoreactive B cells in the splenic marginal zone and result in lupus-like disease. Immunity, 25, 403–415.
Yang, M., Hase, H., Legarda-Addison, D., Varughese, L., Seed, B., & Ting, A. T. (2005). B cell maturation antigen, the receptor for a proliferation-inducing ligand and B cell-activating factor of the TNF family, induces antigen presentation in B cells. Journal of Immunology, 175, 2814–2824.
Kayagaki, N., Yan, M., Seshasayee, D., Wang, H., Lee, W., French, D. M., et al. (2002). BAFF/BLyS receptor 3 binds the B cell survival factor BAFF ligand through a discrete surface loop and promotes processing of NF-kappaB2. Immunity, 17, 515–524.
Claudio, E., Brown, K., Park, S., Wang, H., & Siebenlist, U. (2002). BAFF-induced NEMO-independent processing of NF-kappa B2 in maturing B cells. Nature Immunology, 3, 958–965.
Sasaki, Y., Casola, S., Kutok, J. L., Rajewsky, K., & Schmidt-Supprian, M. (2004). TNF family member B cell-activating factor (BAFF) receptor-dependent and -independent roles for BAFF in B cell physiology. Journal of Immunology, 173, 2245–2252.
Rahman, Z. S., Rao, S. P., Kalled, S. L., & Manser, T. (2003). Normal induction but attenuated progression of germinal center responses in BAFF and BAFF-R signaling-deficient mice. Journal of Experimental Medicine, 198, 1157–1169.
Franzoso, G., Carlson, L., Poljak, L., Shores, E. W., Epstein, S., Leonardi, A., et al. (1998). Mice deficient in nuclear factor (NF)-kappa B/p52 present with defects in humoral responses, germinal center reactions, and splenic microarchitecture. Journal of Experimental Medicine, 187, 147–159.
Tucker, E., O’Donnell, K., Fuchsberger, M., Hilton, A. A., Metcalf, D., Greig, K., et al. (2007). A novel mutation in the Nfkb2 gene generates an NF-kappa B2 “super repressor”. Journal of Immunology, 179, 7514–7522.
Xiao, G., Harhaj, E. W., & Sun, S. C. (2001). NF-kappaB-inducing kinase regulates the processing of NF-kappaB2 p100. Molecular cell, 7, 401–409.
He, J. Q., Zarnegar, B., Oganesyan, G., Saha, S. K., Yamazaki, S., Doyle, S. E., et al. (2006). Rescue of TRAF3-null mice by p100 NF-kappa B deficiency. Journal of Experimental Medicine, 203, 2413–2418.
Liao, G., Zhang, M., Harhaj, E. W., & Sun, S. C. (2004). Regulation of the NF-kappaB-inducing kinase by tumor necrosis factor receptor-associated factor 3-induced degradation. Journal of Biological Chemistry, 279, 26243–26250.
Ni, C. Z., Oganesyan, G., Welsh, K., Zhu, X., Reed, J. C., Satterthwait, A. C., et al. (2004). Key molecular contacts promote recognition of the BAFF receptor by TNF receptor-associated factor 3: Implications for intracellular signaling regulation. Journal of Immunology, 173, 7394–7400.
Xu, L. G., & Shu, H. B. (2002). TNFR-associated factor-3 is associated with BAFF-R and negatively regulates BAFF-R-mediated NF-kappa B activation and IL-10 production. Journal of Immunology, 169, 6883–6889.
Patke, A., Mecklenbrauker, I., Erdjument-Bromage, H., Tempst, P., & Tarakhovsky, A. (2006). BAFF controls B cell metabolic fitness through a PKC beta- and Akt-dependent mechanism. Journal of Experimental Medicine, 203, 2551–2562.
Mecklenbrauker, I., Kalled, S. L., Leitges, M., Mackay, F., & Tarakhovsky, A. (2004). Regulation of B-cell survival by BAFF-dependent PKCdelta-mediated nuclear signalling. Nature, 431, 456–461.
Hauser, A. E., Junt, T., Mempel, T. R., Sneddon, M. W., Kleinstein, S. H., Henrickson, S. E., et al. (2007). Definition of germinal-center B cell migration in vivo reveals predominant intrazonal circulation patterns. Immunity, 26, 655–667.
Schwickert, T. A., Lindquist, R. L., Shakhar, G., Livshits, G., Skokos, D., Kosco-Vilbois, M. H., et al. (2007). In vivo imaging of germinal centres reveals a dynamic open structure. Nature, 446, 83–87.
Allen, C. D., Okada, T., & Cyster, J. G. (2007). Germinal-center organization and cellular dynamics. Immunity, 27, 190–202.
Anderson, S. M., Hannum, L. G., & Shlomchik, M. J. (2006). Memory B cell survival and function in the absence of secreted antibody and immune complexes on follicular dendritic cells. Journal of Immunology, 176, 4515–4519.
Ahmed, R., & Gray, D. (1996). Immunological memory and protective immunity: Understanding their relation. Science, 272, 54–60.
Gray, D., Bergthorsdottir, S., van Essen, D., Wykes, M., Poudrier, J., & Siepmann, K. (1997). Observations on memory B-cell development. Seminars in Immunology, 9, 249–254.
Mantchev, G. T., Cortesao, C. S., Rebrovich, M., Cascalho, M., & Bram, R. J. (2007). TACI is required for efficient plasma cell differentiation in response to T-independent type 2 antigens. Journal of Immunology, 179, 2282–2288.
Huang, X., Di Liberto, M., Cunningham, A. F., Kang, L., Cheng, S., Ely, S., et al. (2004). Homeostatic cell-cycle control by BLyS: Induction of cell-cycle entry but not G1/S transition in opposition to p18INK4c and p27Kip1. Proceedings of the National Academy of Sciences of the United States of America, 101, 17789–17794.
Holcik, M., & Korneluk, R. G. (2001). XIAP, the guardian angel. Nature Reviews. Molecular Cell Biology, 2, 550–556.
Qian, Y., Qin, J., Cui, G., Naramura, M., Snow, E. C., Ware, C. F., et al. (2004). Act1, a negative regulator in CD40- and BAFF-mediated B cell survival. Immunity, 21, 575–587.
Vora, K. A., Wang, L. C., Rao, S. P., Liu, Z. Y., Majeau, G. R., Cutler, A. H., et al. (2003). Cutting edge: Germinal centers formed in the absence of B cell-activating factor belonging to the TNF family exhibit impaired maturation and function. Journal of Immunology, 171, 547–551.
Hase, H., Kanno, Y., Kojima, M., Hasegawa, K., Sakurai, D., Kojima, H., et al. (2004). BAFF/BLyS can potentiate B-cell selection with the B-cell coreceptor complex. Blood, 103, 2257–2265.
Scholz, J. L., Crowley, J. E., Tomayko, M. M., Steinel, N., O’Neill, P. J., Quinn, W. J., 3rd, et al. (2008). BLyS inhibition eliminates primary B cells but leaves natural and acquired humoral immunity intact. Proceedings of the National Academy of Sciences of the United States of America, 105, 15517–15522.
Treml, L. S., Quinn, W. J., 3rd, Treml, J. F., Scholz, J. L., & Cancro, M. P. (2008). Manipulating B cell homeostasis: A key component in the advancement of targeted strategies. Archivum Immunologiae et Therapiae Experimentalis (Warsz), 56, 153–164.
Stohl, W., Xu, D., Kim, K. S., Koss, M. N., Jorgensen, T. N., Deocharan, B., et al. (2005). BAFF overexpression and accelerated glomerular disease in mice with an incomplete genetic predisposition to systemic lupus erythematosus. Arthritis and Rheumatism, 52, 2080–2091.
Cambridge, G., Stohl, W., Leandro, M. J., Migone, T. S., Hilbert, D. M., & Edwards, J. C. (2006). Circulating levels of B lymphocyte stimulator in patients with rheumatoid arthritis following rituximab treatment: Relationships with B cell depletion, circulating antibodies, and clinical relapse. Arthritis and Rheumatism, 54, 723–732.
Stohl, W. (2006). Therapeutic targeting of B lymphocyte stimulator (BLyS) in the rheumatic diseases. Endocrine, Metabolic & Immune Disorders Drug Targets, 6, 351–358.
Gottenberg, J. E., Busson, M., Cohen-Solal, J., Lavie, F., Abbed, K., Kimberly, R. P., et al. (2005). Correlation of serum B lymphocyte stimulator and beta2 microglobulin with autoantibody secretion and systemic involvement in primary Sjogren’s syndrome. Annals of the Rheumatic Diseases, 64, 1050–1055.
Garibyan, L., Lobito, A. A., Siegel, R. M., Call, M. E., Wucherpfennig, K. W., & Geha, R. S. (2007). Dominant-negative effect of the heterozygous C104R TACI mutation in common variable immunodeficiency (CVID). Journal of Clinical Investigation, 117, 1550–1557.
Keats, J. J., Fonseca, R., Chesi, M., Schop, R., Baker, A., Chng, W. J., et al. (2007). Promiscuous mutations activate the noncanonical NF-kappaB pathway in multiple myeloma. Cancer Cell, 12, 131–144.
Annunziata, C. M., Davis, R. E., Demchenko, Y., Bellamy, W., Gabrea, A., Zhan, F., et al. (2007). Frequent engagement of the classical and alternative NF-kappaB pathways by diverse genetic abnormalities in multiple myeloma. Cancer Cell, 12, 115–130.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Treml, J.F., Hao, Y., Stadanlick, J.E. et al. The BLyS Family: Toward a Molecular Understanding of B Cell Homeostasis. Cell Biochem Biophys 53, 1–16 (2009). https://doi.org/10.1007/s12013-008-9036-1
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12013-008-9036-1