
Abstract
Pyroptosis is a form of programmed cell death characterized by cell swelling, pore formation in the plasma membrane, lysis, and releases of cytoplasmic contents. To date, the molecular mechanism of human and murine Gasdermin D-mediated pyroptosis have been fully investigated. However, studies focusing on molecular mechanism of bovine Gasdermin D (bGSDMD)-mediated pyroptosis and its function against pathogenic infection were unclear. In the present study, we demonstrate that bovine caspase-1 (bCaspase-1) cleaves bGSDMD at amino acid residue D277 to produce an N-terminal fragment (bGSDMD-p30) which leads to pyroptosis. The amino acid residues T238 and F239 are critical for bGSDMD-p30-mediated pyroptosis. The loop aa 278-299, L293 and A380 are the key sites for autoinhibitory structure of the full length of bGSDMD. In addition, bCaspase-3 also cleaves bGSDMD at residue Asp86 without inducing cell death. Therefore, our study provides the first detailed elucidation of the mechanism of bovine GSDMD-mediated pyroptosis. The results will establish a significant foundation for future research on the role of pyroptosis in bovine infectious diseases.
Similar content being viewed by others
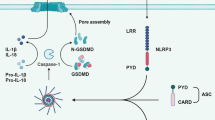


Introduction
Pyroptosis is a form of lytic programmed cell death dependend on inflammatory caspases such as Caspase-1, -4, -5, and -11, which are activated by inflammasome in response to infection or danger signals [1, 2]. Pyroptosis is characterized by extensive membrane blebbing followed by ballooning of the membrane and eventual loss of membrane integrity [3]. Pyroptosis can defend against intracellular infection by eliminating the compromised cell, thereby removing the protective niche of the pathogen, and releasing cellular contents to elicit an immune response [4]. However, excessive pyroptosis can release numerous intracellular molecules such as DAMP, together with a burst in pro-inflammatory cytokines, resulting in a highly inflammatory response and tissue damage [5]. Cytosolic LPS-mediated pyroptosis was found to be the main driver for sepsis and septic shock [6].
Gasdermin D (GSDMD), as an effector of inflammasome-triggered pyroptosis, is a member of the conserved gasdermin protein family including GSDMA, GSDMB, GSDMC, DFNAS/GSDME, and DFNB59 [7]. GSDMD is cleaved into GSDMD-N-terminal domain (GSDMD NT) and GSDMD-C-terminal domain (GSDMD CT) by inflammatory caspases [8,9,10]. GSDMD NT oligomerizes into ring-shaped structures to form a large 10–15 nm diameter pore in the plasma membrane, allowing for the passage of IL-1β (4.5 nm in diameter) and IL-18 (5 nm in diameter) [11,12,13,14]. GSDMD NT has an affinity for phosphatidylinositol phosphate species, such as PIP1, PIP2, and PIP3, and phosphatidylserine (PS) or cardiolipin, not binding with phosphatidylinositol (PI), phosphatidylcholine (PC), and phosphatidylethanolamine (PE) [12, 13]. Since PIP species and PS are restricted to the inner plasma membrane, GSDMD NT can only form pores from the cytosolic face and does not harm neighboring cells. Furthermore, cardiolipin is a component of bacterial membranes, thus GSDMD NT could kill bacteria, at least in vitro. GSDMD CT could repress pore-forming activity of GSDMD NT by interacting with GSDMD NT in a large surface area [15].
Although the detailed characterization of GSDMD-mediated pyroptosis and the interaction between GSDMD and pathogen in human and rodent systems, our understanding of GSDMD-mediated pyroptosis in bovine is limited. Therefore, this study aims to investigate the characterization of bovine GSDMD in the execution of pyroptosis, including the cleavage site by bovine Caspase-1, critical fragments, or amino acids for pyroptotic activity.
Materials and methods
Antibody and reagent
Mouse anti-Flag (AF519) was purchased from Beyotime Biotechnology. Mouse anti-Myc-Tag (#2276), Rabbit anti-GFP (#2956) and Rabbit anti-Gasdermin D (L60, #93,709) antibodies were obtained from Cell Signaling Technology. Mouse anti-GAPDH (AC033) was purchased from Abclonal Technology. Goat-anti-Mouse and Goat-anti-Rabbit antibodies were purchased from Jackson ImmunoResearch.
Cell cultures and transfection
HEK293T and MBDK cells were obtained from ATCC. HEK293T and MBDK cells were grown in Dulbecco’s modified Eagles’s medium (DMEM) basic supplemented with 10% fetal bovine serum (FBS). Cells were grown at 37 °C in a 5% CO2 incubator. Transient transfection of HEK293T cells was performed using JetPRIME (Polyplus Transfection).
Plasmid construction
Complementary DNA (cDNA) for bovine GSDMD (NM_001359976.1) was amplified from reverse-transcribed cDNA of bovine macrophages. The fragments bGSDMD with or without GFP tag were cloned into p3xFlag-CMV-7.1 or p3xFlag-CMV-14 vectors. The substitution mutants and truncated mutants of bGSDMD were cloned into p3xFlag-CMV-7.1 vector. cDNAs for bovine Caspase-1 and Caspase-3 were synthesized by Tsingke Biotechnolog and cloned into pCMV-Myc vector.
Cytotoxicity assay
A CytoTox 96 Non-Radioactive Cytotoxicity assay (G1780, Promega) was used to measure cell death. HEK 293 T cells were seeded at a density of 1 × 105 per well in 48-well plates overnight. The cells were then transfected plasmids as indicated for 24 h. To investigate the pyroptosis induced BVDV infection, MDBK cells were seeded at a density of 2.5 × 105 per well in 24-well plates overnight. Subsequently, the cells were infected with BVDV at MOI 1 for the indicated time. Following either transfection or infection, the supernatant of the cells was collected for detection of LDH release.
Microscopy imaging of cell death
To examine cell death morphology, HEK293T cells were seeded at 1 × 105 per well in 24-well plates overnight for static image capture or in coverslips for fluorescence imaging. Following transfection with the specified plasmids for 24 h, static bright field images of pyroptotic cells were captured using a Nikon ECLIPSE Ts2. To investigate the location of N-GSDMD, cells were incubated with Dil (a red fluorescent probe for cell membrane, Beyotime Biotechnology, C1991S) and DAPI (Beyotime Biotechnology, C1006) for 10 min. Subsequently, cells were rinsed twice with PBS to remove unbound probe. Images were acquired using a confocal laser-scanning microscope (Olympus). The acquired images were processed with Image J software. All image data shown are representative of at least three randomly selected fields.
Western blot and immunoprecipitation
HEK293T or MDBK cells were lysed using Cell lysis buffer supplemented with a protease inhibitor cocktail (Biomark). The lasytes were centrifuged at 12 000 × g for 10 min, and the supernatant was obtained and then denatured in 4 × SDS sample buffer. The protein samples were separated by 12% SDS-PAGE gels and transferred onto PVDF membrane. Membranes were incubated with antibodies and developed using ECL substrate (Bio-Rad). For immunoprecipitations, lysates were incubated overnight at 4 °C with anti-FLAG Beads (Sigma Aldrich). The beads were washed five times with lysis buffer and the immunoprecipitated were eluted with SDS loading buffer and subjected to Western blot assay.
Statistical analysis
All assays were performed at least three times independently. Data are presented as mean ± SD and analyzed using two-tailed Student’s t-test or one-way ANOVA by Prism software (GraphPad). The differences were considered significant when P < 0.05 (*), P < 0.01 (**), P < 0.001 (***).
Results
Bovine Caspase-1 cleaves bovine GSDMD to induce pyroptosis
We employed BVDV as a representative bovine pathogen to investigate pyroptosis and bovine GSDMD (bGSDMD) cleavage in MDBK cells. The results showed that BVDV induced pyroptosis at 24 h and 48 h post-infection (Additional file 1A), while bGSDMD cleavages were detected as early as 6 h post-infection in BDBK cells (Additional file 1B). To investigate the role of bGSDMD in the induction of pyroptosis, HEK293T cells were co-transfected with plasmids expressing bovine Caspase-1 (bCaspase-1) and bGSDMD, respectively. The results, as depicted in Figure 1A, revealed that HEK293T cells co-expressing bCaspase-1 and bGSDMD exhibited a higher release of LDH into the supernatant. Additionally, the co-expression of bCaspase-1 and bGSDMD in HEK293T cells led to extensive cell death characterized by the typical pyroptosis morphology, which includes cell swelling, membrane rupture, and cytoplasmic leakage (Figure 1B). To investigate the cleavage of bGSDMD by bCaspase-1, we constructed a plasmid expressing bGSDMD with a Flag tag at its N-terminus and a GFP tag at its C-terminus. As illustrated in Figure 1C, bCaspase-1 cleaved bGSDMD, resulting in the generation of an N-terminal fragment with a Flag tag (approximately 35 kDa, plus a 2.9 kDa Flag tag) and a C-terminal fragment with a GFP tag (approximately 50 kDa, plus a 26.9 kDa GFP tag). Additionally, we observed an interaction between bCaspase-1 and bGSDMD (Figure 1D). Futhermore, the microscopic examination revealed the presence of N-GSDMD with a GFP tag localized in the cellular membrane (Figure 1E). These findings provide evidence that bGSDMD is cleaved by bCaspase-1, leading to the induction of pyroptosis.

bCaspase-1 is responsible for bGSDMD cleavage. HEK293T cells were co-transfected with plasmids encoding bCaspase-1 and bGSDMD for 24 h. A The supernatants were collected and analyzed for LDH release levels. B Microscopic images of the indicated HEK293T cells. Scale bar, 50 μm. C Cell lysates were analyzed with anti-Flag, anti-GFP, anti-Myc, and anti-GAPDH by Western blot. D Cell lysates were immunoprecipitated with anti-Myc antibody and then analyzed with anti-Flag and anti-Myc antibodies by Western blot. FL: full length; NT: N-terminal fragment; CT: C-terminal fragment. E Cells were incubated with Dil (a red fluorescent probe for cell membrane) and DAPI for 10 min. The fluorescent signals were observed with confocal immunofluorescence microscopy. * P < 0.05, ** P < 0.01, *** P < 0.001
bCaspase-1 cleaves bGSDMD at amino acid residue D277
Based on the predicted cleavage site preference of Caspase-1 (XXXD motif) and the sizes of the cleaved fragments of bGSDMD, we performed site-directed mutagenesis to introduce alanine substitutions at the predicted cleavage sites D270 and D277 (Additional file 2), generating single- or double-site mutated forms of bGSDMD. In HEK293T cells, both the wild-type bGSDMD and the D270A mutant induced significant cell death with clear pyroptosis morphology, as depicted in Figures 2A and B. In contrast, the D277A mutant only caused a slight induction of cell death (Figures 2A and B). Furthermore, the D277A mutant resisted cleavage by bCaspase-1 (Figure 2C). The ratio of pyroptosis observed in cells transfected with D270A D277A double mutant was comparable to that of cells transfected with the D277A mutant. These findings indicate that the amino acid residue D277 is the only inflammatory-Caspase cleavage site present in bovine GSDMD.

Residue D277 of bGSDMD is the cleavage site for bCaspase-1. HEK293T cells were individually transfected with indicated plasmids for 24 h. A The supernatants were collected and analyzed for LDH release levels. B Microscopic images of the indicated HEK293T cells. Scale bar, 50 μm. C Cell lysates were analyzed with anti-Flag, anti-Myc, and anti-GAPDH by Western blot. * P < 0.05, ** P < 0.01, *** P < 0.001
Amino acid residues T238 and F239 are key sites for bGSDMD to pyroptosis
EV71 3C protein cleaved human GSDMD at residue Gln 193 to preclude GSDMD activation, whereas PEDV N protein cleaved porcine GSDMD at residue Gln 193 to inhibit GSDMD activation, indicating that the key fragment to maintain GSDMD NT function is between Gln193 and Asp 275 (Asp279 in porcine GSDMD) [16]. To investigate whether bGSDMD has the same characteristic as human GSDMD and porcine GSDMD, we constructed nine bGSDMD truncated mutants expressing amino acids 1-192, 1-202, 1-212, 1-222, 1-232, 1-242, 1-252, 1-262, and 1-277. These mutants were individually transfected into HEK 293 T. As shown in Figure 3A, HEK 293 T cells expressing amino acids 1-242,1-252,1-262, and 1-272 exhibited a high rate of cell death, whereas those expressing 1-192, 1-202, 1-212, 1-222, and 1-232 did not. The same results were observed in cell morphological features (Figure 3B) and PI staining (Additional file 3A), indicating that the key activity sites located between amino acids 232-242. Next, we constructed the single-site mutants of bGSDMD NT in which amino acid residues between 232 and 242 were replaced by Alanine or Aspartate. The results showed that the T238D and F239D mutants attenuate pyroptosis-inducing activity of bGSDMD NT in HEK293T cells (Figures 3C and D, Additional file 3B). Moreover, we observed T238D and F239D mutants still produced comparable interaction of bGSDMD NT with Flag tag or HA tag compared with wild type (Figures 3E and F), indicating that lower cell death caused by T238D and F239D mutants were not associated with bGSDMD NT self-oligomerization. Collectively, these data suggested that Amino acid residues T238 and F239 are essential sites for bGSDMD NT to induce pyroptosis.

T238 and F239 are the key sites for bGSDMD NT to induce pyroptosis. HEK293T cells were individually transfected with indicated plasmids for 24 h. A and C The supernatants were collected and analyzed for LDH release levels. B and D Microscopic images of the indicated HEK293T cells. Scale bar, 50 μm. E and F Cell lysates were immunoprecipitated with anti-Flag antibody and then analyzed with anti-Flag and anti-HA antibodies by Western blot. * P < 0.05, ** P < 0.01, *** P < 0.001
Functional analyses of bGSDMD autoinhibition
While GSDMD is intact, GSDMD C-terminal domain (bGSDMD CT) could autoinhibit pyroptosis-inducing activity of bGSDMD NT. The flexible loop (aa 276-296) in human GSDMD inserts into the GSDMD NT pocket, stabilizing the overall structure of GSDMD [15]. Based on the multiple-sequence alignment of GSDMD from human, mouse, porcine, and bovine, we hypothesize that the fragment between aa 278 and aa 299 in bGSDMD might have the same function. We constructed the mutant of bGSDMD in which the fragment (aa278-299) was substituted by a short linker sequence, GSGGGS, without changing the protein quaternary structure. Indeed, the mutant induced more cell death than bGSDMD FL in HEK293T cells (Figures 4A and B). Furthermore, we also investigate amino acids L293, Y376, and A380 in bGSDMD homologous to L290, Y373, and A377 in hGSDMD which have been reported as the key sites in bGSDMD CT autoinhibition. As shown in Figures 4C and D, the results indicated that bGSDMD mutants L293D and A380D rather than Y376 exhibited spontaneous pyroptosis-inducing activity. Collectively, the findings suggested that the loop aa 278-299, L293, and A380 are essential to maintain the stability and autoinhibition of GSDMD.

The loop aa 278–299, L293 and A380 are essential to maintain the autoinhibition of bGSDMD. HEK293T cells were individually transfected with indicated plasmids for 24 h. A and C The supernatants were collected and analyzed for LDH release levels. B and D Microscopic images of the indicated HEK293T cells. Scale bar, 50 μm. * P < 0.05, ** P < 0.01, *** P < 0.001
bCaspase-3 cleaves bGSDMD without inducing cell death
It has been reported that bCaspase-3 has capable of cleaving bGSDMD at residue Asp87 during apoptosis. To investigate whether bCaspase-3 has the same function as cleaving bGSDMD, we transfected the plasmid expressing bCaspase-3 together with that expressing bGSDMD with a GFP tag at its N-terminus and a Flag tag at its C-terminus into HEK293T cells. As shown in Figure 5A, bCaspase-3 indeed cleaved bGSDMD into two fragments: the smaller-size N-terminal domain (approximately 37 kDa, plus with 26.9 kDa GFP tag) and the bigger-size C-terminal domain (approximately 46 kDa, plus with 2.9 kDa Flag tag). Based on the Caspase-3 cleavage motif (DXXD), we substituted Asp20 and Asp86 with Alanine. D86A, rather than D20A, was resistant to the cleavage by bCaspase-3 (Figure 5B). However, the cleavage of bGSDMD by bCaspase-3 did not induce more cell death (Figures 5C and D). Thus, the findings indicated that bCaspase-3 cleaves bGSDMD at residue Asp86 without inducing cell death.

bCaspase-3 cleaved bGSDMD at residue D86. HEK293T cells were individually transfected with indicated plasmids for 24 h. A and B Cell lysates were analyzed with anti-Flag, anti-GFP, and anti-GAPDH by Western blot. C The supernatants were collected and analyzed for LDH release levels. D Microscopic images of the indicated HEK293T cells. Scale bar, 50 μm. * P < 0.05, ** P < 0.01, *** P < 0.001
Discussion
Various stimuli or pathogens induce pyroptosis in bovine cells [17,18,19], however, the precise mechanisms underlying bovine GSDMD-mediated pyroptosis remain unclear. In this study, we show that bovine GSDMD as a substrate for bovine Caspase-1 is essential for pyroptosis. We further elucidate the mechanism of the cleavage site, critical fragment, and key amino acids involved in bGSDMD-mediated pyroptosis. Thus, our research findings provide a detailed elucidation of the mechanism by which GSDMD is involved in pyroptosis, laying the foundation for future research for pyroptosis-related diseases in bovine species.
Prior to our work, human/murine/porcine GSDMD had been demonstrated as a direct substrate of Caspase-1/4/5/11 and serves as the key mediator for pyroptosis [8, 10, 20]. However, the specific amino acid sequence and molecular characterization of bovine GSDMD have remained unexplored. To investigate the molecular mechanism of bGSDMD-mediated pyroptosis, we first confirmed that bCaspase-1 cleaves bovine GSDMD at D277, generating the active peptide bGSDMD-p30, which induces pyroptosis. Furthermore, previous studies revealed that C38/C39 and C191/C192 (human/murine) mutations impaired hGSDMD-p30/mGSDMD-p30 oligomerization [14, 21]. Our results showed that mutation of pGSDMD-p30 T238D or F239D failed to induce LDH release indicating that these two sites are key sites for GSDMD to active pyroptosis. Further investigations are required to determine if GSDMD inhibitors (NSA and NSC) inhibit bGSDMD-p30 oligomerization directly or use alternative mechanisms to suppress bGSDMD-p30-induced pyroptosis. In addition, like other Caspases, Caspase-3 cleaves multiple substrates, including hGSDME [22] and hGSDMD [23]. It has been observed that active caspase-3 inhibits pyroptosis by cleaving hGSDMD at D87 [23]. Consistent with this finding, our study demonstrated that bCaspase-3 cleaves bGSDMD at D86 but does not induce cell death. Therefore, it is reasonable to hypothesize that bCaspase-3 may employ a similar mechanism to suppress pyroptosis in bovine cells.
In conclusion, our study demonstrates that bGSDMD is cleaved by bCaspase-1 at the D277 residue, leading to the initiation of pyroptosis. The specific amino acid residues T238 and F239 are crucial for the pyroptotic activity of bovine GSDMD. Additionally, our investigation into the autoinhibition mechanism of bovine GSDMD sheds light on the regulatory processes involved in pyroptosis in bovine systems.
Availability of data and materials
The data supporting the conclusions of this article are included within the article. Additional data used and/or analysed during the current study are available from the corresponding author upon reasonable request.
References
Brennan MA, Cookson BT (2000) Salmonella induces macrophage death by caspase-1-dependent necrosis. Mol Microbiol 38:31–40
Kayagaki N, Warming S, Lamkanfi M, Walle LV, Louie S, Dong J, Newton K, Qu Y, Liu J, Heldens S, Zhang J, Lee WP, Roose-Girma M, Dixit VM (2011) Non-canonical inflammasome activation targets caspase-11. Nature 479:117–121
Fink SL, Cookson BT (2006) Caspase-1-dependent pore formation during pyroptosis leads to osmotic lysis of infected host macrophages. Cell Microbiol 8:1812–1825
Fink SF, Bergsbaken T, Cookson BT (2008) Anthrax lethal toxin and Salmonella elicit the common cell death pathway of Caspase-1-dependent pyroptosis via distinct mechanisms. Proc Natl Acad Sci USA 105:4312–4317
Kang R, Zeng L, Zhu S, Xie Y, Liu J, Wen Q, Cao L, Xie M, Ran Q, Kroemer G, Wang H, Billiar TR, Jiang J, Tang D (2018) Lipid peroxidation drives gasdermin D-mediated pyroptosis in lethal polymicrobial sepsis. Cell Host Microbe 24:97-108.e4
Wang D, Yu S, Zhang Y, Huang L, Luo R, Tang Y, Zhao K, Lu B (2019) Caspse-11-GSDMD pathway is required for serum ferritin secretion in sepsis. Clin Immunol 205:148–152
Kovacs SB, Miao EA (2017) Gasdermins: effectors of pyroptosis. Trends Cell Biol 27:673–684
Shi J, Zhao Y, Wang K, Shi X, Huang H, Zhaung Y, Cai T, Wang F, Shao F (2015) Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 526:660–665
He W, Wan H, Hu L, Chen P, Wang X, Huang Z, Yang Z, Zhong C, Han J (2015) Gasdermin D is an executor of pyroptosis and required for interleukin-1 beta secretion. Cell Res 25:1285–1298
Kayagaki N, Stowe IB, Lee BL, Rourke KO, Anderson K, Warming S, Cuellar T, Haley B, Roose-Girma M, Phung QT, Liu PS, Lill JR, Li H, Wu J, Kummerfeld S, Zhang J, Lee WP, Snipas SJ, Morris LX, Fitzgerald L, Zhang Y, Bertram EM, Goodnow CC, Dixit VM (2015) Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 526:666–671
Aglietti RA, Estevez A, Gupta A, Ramirez MG, Liu PS, Kayagaki N, Ciferri C, Dixit VM, Dueber EC (2016) GsdmD p30 elicited by caspase-11 during pyroptosis forms pores in membranes. Proc Natl Acad Sci USA 113:7858–7863
Ding J, Wang K, Liu W, She Y, Sun Q, Shi J, Sun H, Wang D, Shao F (2016) Pore-forming activity and structural autoinhibition of the gasdermin family. Nature 535:111–116
Liu X, Zhang Z, Ruan J, Pan Y, Magupalli VG, Wu H, Lieberman J (2016) Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 535:153–158
Sborgi L, Rühl S, Mulvihill E, Pipercebic J, Heilig R, Stahlberg H, Farady CJ, Müller DJ, Broz P, Hiller S (2016) GSDMD membrane pore formation constitutes the mechanism of pyroptotic cell death. Embo J 35:1766–1778
Kuang S, Zheng J, Yang H, Li S, Duan S, Shen Y, Ji C, Gan J, Xu X, Li J (2017) Structure insight of GSDMD reveals the basis of GSDMD autoinhibition in cell pyroptosis. Proc Natl Acad Sci U S A 114:10642–10647
Lei X, Zhang Z, Xiao X, Qi J, He B, Wang J (2017) Enterovirus 71 inhibits pyroptosis through cleavage of gasdermin D. J Virol 91:e01069-17
Ma X, Li Y, Shen W, Oladejo AO, Yang J, Jiang W, Imam BH, Wu X, Ding X, Yang Y, Wang S, Yan Z (2021) LPS mediates bovine endometrial epithelial cell pyroptosis directly through both NLRP3 classical and non-classical inflammasome pathways. Front Immunol 12:676088
Shen W, Ma X, Shao D, Wu X, Wang S, Zheng J, Lv Y, Ding X, Ma B, Yan Z (2022) Neutrophil extracellular traps mediate bovine endometrial epithelial cell pyroptosis in dairy cows with endometritis. Int J Mol Sci 23:14013
Wang X, Liu M, Geng N, Du Y, Li Z, Gao X, Han B, Liu J, Liu Y (2022) Staphylococcus aureus mediates pyroptosis in bovine mammary epithelial cell via activation of NLRP3 inflammasome. Vet Res 53:10
Shi F, Lv Q, Wang T, Xu J, Xu W, Shi Y, Fu X, Yang T, Yang Y, Zhuang L, Fang W, Gu J, Li X (2022) Coronaviruses Nsp5 antagonizes porcine gasdermin D-mediated pyroptosis by cleaving pore-forming p30 fragment. mBio. https://doi.org/10.1128/mbio.02739-21
Rathkey JK, Zhao J, Liu Z, Chen Y, Yang J, Kondolf HC, Benson BL, Chirieleison SM, Huang AY, Dubyak GR, Xiao TS, Li X, Abbott DW (2018) Chemical disruption of the pyroptotic pore-forming protein gasdermin D inhibits inflammatory cell death and sepsis. Sci Immunol 3:e2738
Wang Y, Gao W, Shi X, Ding J, Liu W, He H, Wang K, Shao F (2017) Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 547:99–103
Taabazuing CY, Okondo MC, Bachovchin DA (2017) Pyroptosis and apoptosis pathways engage in bidirectional crosstalk in monocytes and macrophages. Cell Chem Biol 24:507-514.e4
Acknowledgements
This study was supported by Natural Science Foundation of China (No. 32272951), Zhejiang Provincal Natural Science Foudation (No. LY21C180001), the Fundamental Research Funds for the Provincial Universities of Zhejiang (No. 2022YQ008). The funding bodies had no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Funding
The National Natural Science Foundation of China, No. 32272951, Yang Yang, Zhejiang Provincal Natural Science Foudation, No. LY21C180001, Yang Yang, the Fundamental Research Funds for the Provincial Universities of Zhejiang, No. 2022YQ008, Yang Yang.
Author information
Authors and Affiliations
Contributions
Y. Yang designed research, ZG, JX and KY performed research, SC provided BVDV and MDBK cells, LW, BC, JT, JZ, AX and BH analyzed data; YY and HS wrote the paper. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Handling editor: Tom McNeilly
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: BVDV infection induces GSDMD-mediated pyroptosis.
MDBK cells were infected with BVDV at MOI 1 for 6 h, 24 h and 48 h. A The supernatants were collected and analyzed for LDH release levels. B Cell lysates were analyzed with anti-bGSDMD by Western blot.
Additional file 3: T238 and F239 are the key sites for bGSDMD NT to induce pyroptosis.
HEK293T cells were individually transfected with indicated plasmids for 24 h and subsequently dyed with PI for 15 min. The fluorescent signals were observed with confocal immunofluorescence microscopy.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ge, Z., Xu, J., Yang, K. et al. Molecular mechanism of bovine Gasdermin D-mediated pyroptosis. Vet Res 55, 26 (2024). https://doi.org/10.1186/s13567-024-01282-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-024-01282-1