
Abstract
Background
Increasing exposure to engineered inorganic nanoparticles takes actually place in both terrestric and aquatic ecosystems worldwide. Although we already know harmful effects of AgNP on the soil bacterial community, information about the impact of the factors functionalization, concentration, exposure time, and soil texture on the AgNP effect expression are still rare. Hence, in this study, three soils of different grain size were exposed for up to 90 days to bare and functionalized AgNP in concentrations ranging from 0.01 to 1.00 mg/kg soil dry weight. Effects on soil microbial community were quantified by various biological parameters, including 16S rRNA gene, photometric, and fluorescence analyses.
Results
Multivariate data analysis revealed significant effects of AgNP exposure for all factors and factor combinations investigated. Analysis of individual factors (silver species, concentration, exposure time, soil texture) in the unifactorial ANOVA explained the largest part of the variance compared to the error variance. In depth analysis of factor combinations revealed even better explanation of variance. For the biological parameters assessed in this study, the matching of soil texture and silver species, and the matching of soil texture and exposure time were the two most relevant factor combinations. The factor AgNP concentration contributed to a lower extent to the effect expression compared to silver species, exposure time and physico–chemical composition of soil.
Conclusions
The factors functionalization, concentration, exposure time, and soil texture significantly impacted the effect expression of AgNP on the soil microbial community. Especially long-term exposure scenarios are strongly needed for the reliable environmental impact assessment of AgNP exposure in various soil types.
Similar content being viewed by others

Background
The production volume and the application fields of silver nanoparticles (AgNP) increased continuously in the last decade given by their unique properties such as high surface-to-volume ratio, high chemical reactivity, and specific optical properties [1,2,3]. Apart from the initial medical utilization, AgNP are actually used in households, industry and agriculture such as for water purification, plant growth promotion and textiles cleaning [4, 5]. In consequence, their emission into the environment during all stages of the life cycle, including production, product use, disposal and weathering is unavoidable [6]. The exact extent of the release is unknown due to missing reliable and robust analytical methods for detecting trace concentrations of AgNP in complex matrices [7]. Thus, several scientists modeled the fate and concentrations of AgNP in the environment and named the soil compartment as one of the main sink of AgNP released into the environment [2, 6, 8,9,10,11,12,13,14]. For Europe, an annual AgNP increase of 0.6 t and 2.09 t was calculated for soils and sediments, respectively [8].
Today, information about the impact of AgNP on the soil microbiome are still rare, although microbial communities are important and sensitive targets for determining the environmental hazards of AgNP [15]. Recently, we registered significant negative effects on soil microbial biomass (− 38.0%), bacterial ammonia oxidizers (− 17.0%), and the beta-Proteobacteria population (− 14.2%) after 1-year exposure to 0.01 mg AgNP/kg in a loamy soil, while Acidobacteria (44.0%), Actinobacteria (21.1%) and Bacteroidetes (14.6%) were significantly stimulated [16, 17]. Therefore, a detrimental disturbance on soil ecosystem functions, such as nitrification, organic carbon transformation and chitin degradation could be assumed.
Numerous studies documented the differing physico–chemical and concomitant toxicological behavior of AgNP in dependence of the soil type. As a function of pH, ionic strength, temperature, amount of dissolved ions and of natural organic matter, oxygen concentration, grain size distribution and others [18,19,20,21], AgNP could undergo various physico–chemical transformations such as reduction, oxidation, aggregation, dissolution, complexation and further secondary reactions [21,22,23,24]. Consequently, these transformations in turn affect the toxicity mechanism of AgNP as well as their bioavailability. For example, in comparative studies with different soil types, Schlich and Hund-Rinke [19] as well as Rahmatpour et al. [25] showed that AgNP caused lower toxicity in soils with higher clay content due to the AgNP immobilization by heteroaggregation with clay particles [19, 23, 25].
In addition, the AgNP species itself may significantly impact on its environmental behavior. Their size, shape, surface-coating agent, charge and stability are only a few of the properties by which AgNP can differ [1]. Today, extensive functionalization strategies are available to modify the surface chemistry of a variety of engineered nanoparticles (NP) [26]. Those coatings are used to stabilize NP against aggregation when stable suspensions are required for product functionality or for improved delivery of the product. The coating may also provide other functionalities, such as biocompatibility or targeting of specific cells in biomedical applications [27]. For example, AgNP can be coated by citrate or polyvinylpyrrolidine to increase their stability [28], modified with ATP to act as a selective antibiotic [29] or equipped with COOH– and NH2-groups to affect their surface charge in terms of their function in imaging and drug delivery [30]. Once released into the environment, the surface functionalization of AgNP significantly determines its physico–chemical fate, its bioavailability and its toxicity [22, 26]. Exemplary, Wu et al. [31] observed in a nanocosm experiment that polyethylene glycol AgNP had the highest overall toxicity, followed by silica AgNP, and lastly aminated silica-coated AgNP due to there different dissolution rates und thus stability.
Further to the soil type and the AgNP functionalization, several studies documented a significant impact of the exposure time on the toxicity of AgNP in soils [16, 32,33,34]. By a statistically significant regression and correlation analysis between silver toxicity and exposure time we recently confirmed loamy soils as a sink for silver nanoparticles and their concomitant silver ions due to ageing processes of the silver species and their slow return to the biological soil system [17].
Considering the predicted increase of AgNP into the soil environment, the known toxicity of AgNP to the soil microbial community as well as the variable fate of AgNP in the soil compartment, the aim of this study was to give a more holistic view of the impact of AgNP exposure on the soil microbial community in dependence of the factors AgNP functionalization, AgNP concentration, exposure time, and soil texture. The study was conducted with a long-term incubation period of 90 days using three soil textures (loam, clay, sand) and two different charged AgNP at concentrations in a range of 0.01–1.00 mg AgNP/kg soil. We quantified the effects on several biological parameter: the microbial biomass, the abundance of bacteria, the enzymatic activity as well as marker genes for selected processes of the inorganic nitrogen cycle and for selected higher bacterial taxa. Furthermore, we used AgNO3 as control to determine the effect of Ag+ ions on the AgNP results. Based on our preceding observation that the nitrate content of 36.5% in the AgNO3 compound might also have effects on the microbial community [16], we used NO3 as a further control. This experiment was restricted to the loamy soil. Here, we analysed the impact charged and uncharged AgNP as well as the effects of AgNO3 and NO3 on the microbial community.
Materials and methods
Silver nanoparticles and controls
Two differently functionalized AgNP were used, Ag10-COOH functionalized with carboxy groups and Ag10-NH2 functionalized with amino groups. The AgNP were synthesized by a ligand exchange starting from hydrophobic silver particles (Ag-HPB) and the addition of toluene and mercaptopropionic acid or cysteamine hydrochloride in MeOH to receive the final Ag10-COOH or Ag10-NH2 colloidal solutions. The concentration of the stock solutions were 180 mg/L for Ag10-COOH and 21 mg/L for Ag10-NH2. Size, shape and nanoparticle surface charge (ζ-potential) of AgNP were analysed by transmission electron microscopy (Philips CM 12, Netherlands), dynamic light scattering (DLS, Zetasizer Nano S, Malvern Instruments Ltd., UK), asymmetrical flow field-flow fractionation (AF4, AF2000 MT, Postnova Analytics GmbH, Germany) and Laser-Doppler-microelectrophoresis (Malvern Zetasizer Nano-ZS, Malvern Instruments Ltd., UK).
Methods of synthesis and particle characterization can be found in detail in Additional file 1.
Additionally to the analysis of the stock solution, ζ-potential and hydrodynamic diameter of the AgNP were determined at different pH values (pH 4, pH 7.4 and pH 10). Prior to the measurements the stock solution was diluted in pure water (Millipore) with the pH-values 4, 7.4 or 10 to a concentration of 10 µg/mL, vortexed for 10 s, incubated for 1 h or 24 h under permanent rotation (100 rpm at 37 °C) and vortexed for 30 s prior to analysis via Zetasizer Nano-ZS.
Silver nitrate (AgNO3) was used as a positive control. Silver concentrations in the AgNO3 controls were the same as those in the AgNP treatments. As a further control, NO3− was used in form of KNO3. The nitrate concentration in the KNO3 controls were the same as those in the AgNO3 treatments.
Test soils
Three soil textures were selected: a silty sand, a loamy clay and a silty loam. Approximately 20 kg of each soil was sampled from the A-horizon (0–30 cm depth) in spring 2015 next to Trier, Germany. Land-use was forest for the sandy soil and arable field for the clayey and the loamy soil. The soil had a clay content of 0–5% for the sandy, 45–65% for the clayey, and 25–35% for the loamy soil, respectively. After sampling, the soils were thoroughly sieved to < 2 mm and stored at 6 °C until further use. Characteristic soil parameters are listed in Table 1.
Experimental design
Before starting the experiment, the soil was moistened and incubated at 18 °C for 7 days. The application of the test materials was performed in petri dishes, each filled with soil equivalent to 25 g dry weight.
AgNP test solutions were prepared immediately before use: AgNP stock solutions were sonicated at 42 W/L for 15 min and gradually diluted with UPW. Then, 1 mL of the agent species Ag10-COOH, Ag10-NH2, AgNO3 or KNO3 solutions, at different concentrations, was added in small drops onto the soil surface to obtain final concentrations of 0.01, 0.10 and 1.00 mg/kg dry weight. Negative controls only received an application of UPW. Soil water content after the addition of the test solutions were average 22.0% (sand), 19.4% (clay) and 18.5% (loam), which was equivalent to 42.6%, 41.4%, and 37.4% WHCmax, respectively. For each soil texture, agent species, concentration, and day, separate samples in different soil dishes were prepared as 4 replicates (e.g. 4 × 0.01 mg Ag10-COOH kg−1 sand for day 1). Subsequently, soils were extensively mixed by stirring with a spoon, and then they were transferred to plastic containers (Centrifuge Tubes, 50 mL, VWR, Darmstadt, Germany) and sealed by Parafilm®. They were incubated at 15.1 (± 1.8 °C) in the dark for 1, 14, 28, and 90 days. Water evaporation was determined gravimetrically and then compensated with the addition of UPW. Samples were finally stored at − 20 °C. For analyses, samples were defrosted by incubation overnight at 6 °C. Each replicate was analysed on the effect expressions of the target variables leucine aminopeptidase activity, microbial biomass as well as of functional and taxonomic genes.
Furthermore, results of our previous studies [16, 17] of the effect assessment of AgPure, with an average size of 20 nm and polyacrylate stabilization, were used to calculate the impact of the five agent species AgPure, Ag10-COOH, Ag10-NH2, AgNO3, and KNO3 at concentrations of 0.01; 0.1; and 1.0 mg Ag/kg soil after exposure of 1, 14, 28, and 90 days in the silty loam soil on the same target variables.
Analysis of biological parameters
Leucine aminopeptidase activity
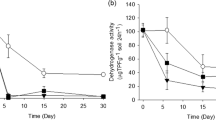
Leucine aminopeptidase (EC 3.4.1.1; LAP) was investigated according to Marx et al. [35], with modifications [36]. Briefly, 1 mol/L L-leucin-7-AMC was used as substrate for LAP, and 7-amino-4-methylcoumarin [37] was used as a standard. Incubation of the soil slurry with the substrate was performed at 30 °C. Measurements of fluorescence were performed after 0 and 2 h using a Victor Multilabel Plate Reader (Perkin Elmer, Germany; excitation wavelength: 355 nm, emission wavelength: 460 nm). LAP activity was calculated as substrate turnover per g dry soil h.
The potential contribution of AgNP to the total fluorescence signal was measured for each AgNP concentration. Then, AgNP was added to autoclaved soil, and the same procedure was conducted. The resulting fluorescence signal was compared to the fluorescence intensity of the pure autoclaved soil. At this, the used AgNP did not exhibit autofluorescence (data not shown).
DNA extraction and microbial biomass measurements
DNA extraction and purification were performed using the Genomic DNA from soil kit (Macherey–Nagel, Düren, Germany) according to the manufacturer’s instructions and stored at − 20 °C. For the measurement of microbial biomass, 10 µL of DNA was transferred into the well of a 96-well microplate and shaken for 5 s before absorbance was measured at 260 nm [38] using Victor Multilabel Plate Reader (Perkin Elmer, Germany).
Quantitative detection of functional and taxonomic genes
16S rRNA genes were used as a proxy to quantify the abundance of bacteria, as described by Bach et al. [39]. The abundance of bacteria harboring the nifH gene was used as a marker for the potential to fix nitrogen and measured according to Rösch et al. [40]. To analyze the effects of the different silver materials on the ammonia-oxidizing bacteria, the amoA primer system described by Rotthauwe et al. [41] was used. To quantify the abundance of taxon-specific 16S rRNA gene copy numbers, qPCR assays for Acidobacteria [42, 43], Actinobacteria [43, 44], alpha-Proteobacteria [45], Bacteroidetes [43, 46], and beta-Proteobacteria [47, 48] were performed. Detailed descriptions of the used assays are given by Grün and Emmerling [17] and Grün et al. [16].
All qPCR reactions were conducted on a thermal cycler equipped with an optical module (Analytik Jena, Jena, Germany). All samples were run in triplicate wells. Single qPCR reactions were prepared in a total volume of 20 µL. The InnuMix SYBR-Green qPCR Master-Mix was purchased from Analytik Jena (Jena, Germany). Primer concentrations were 10 pmol/µL, and amplification specificity was assessed by melting curve analysis and gel electrophoresis on a 1.5% agarose gel after qPCR. Standard curves were based on cloned PCR products from the respective genes [43].
Statistical analyses
All data were processed using IBM SPSS Statistics for Windows, Version 23.0 (IBM Corp., Armonk, USA). The obtained biological values of qPCR (copy gene number per kg dry soil), leucine aminopeptidase activity (substrate turnover per g dry soil h) and measurement of microbial biomass (ng DNA per g dry soil) of negative controls (0.00 mg Ag/kg) of the 4 sample replicates were averaged for each biological parameter and day. Subsequent, the relative variation of each silver treated sample of one concentration and one sampling date were calculated as follow:
In the following, the relative variation was set as target variables. Exposure time (1, 14, 28, 90 days), concentration (0.01; 0.1; 1.0 mg Ag/kg soil), soil texture (loam, sand, clay) and silver species (Ag10-COOH, Ag10-NH2, AgNO3) were set as factors with different factor levels (e.g. Ag10-COOH, Ag10-NH2, AgNO3).
For the effect assessment of a factor level on the target variable, the mean of a target variable (e.g. leucine aminopeptidase) at a distinct factor level (e.g. Ag10-COOH) was calculated. Within a factor, the target variable was also pre-evaluated for a normal distribution by the Shapiro–Wilk test and variance homogeneity by the Levene test.
To test the influence of the four factors as well as the influence of the factor combination on the target variable, multi-factorial ANOVA was performed. Here, tests of between-subject effects provided information about significant relationships. By means of the Bonferroni post hoc test the significance of group mean differences within a factor were calculated.
Finally, a multivariate analysis of variance was performed to simultaneously mitigate the influence of the four factors on the 10 dependent target variables. The factors were set as independent variables, whereas the relative variations of the biological parameters were set as dependent variables. The test statistic was computed by Pillai’s trace.
Results
Characterization of the AgNP suspensions prior to application into soils
Characteristics of the Ag10-COOH stock solution in water were as follows: hydrodynamic diameter (DLS, z-average): 85.3 ± 2.9 nm; polydispersity index (PDI) (DLS) = 0.234 ± 0.031; ζ-potential: − 41.4 ± 1.1 mV (in UPW). AF4-UV-DLS measurement revealed an average hydrodynamic diameter of 18.1 ± 0.5 nm at the UV peak maximum. Over the main UV peak the hydrodynamic diameter ranged from 18 nm to around 28 nm. Larger particle sizes with a hydrodynamic diameter up to around 118 nm were detected as well but larger particle fractions were low in concentration based on the corresponding UV signal (Fig. 1a).

AF4-UV-DLS measurements of Ag10-COOH (a) and Ag10-NH2 (b)
Characteristics of the Ag10-NH2 stock solution in water were as follows: hydrodynamic diameter (DLS, z-average): 62.1 ± 3.1 nm; polydispersity index (PDI) (DLS) = 0.379 ± 0.072; ζ-potential: + 39.2 ± 0.2 mV (in UPW). Average hydrodynamic diameter at the UV peak maximum obtained from AF4-UV-DLS measurement: 82.3 nm ± 3.1 nm with a size distribution from around 8 nm to 142 nm (Fig. 1b).
The analysis of the ζ-potential and hydrodynamic diameter of the AgNP under different pH values resulted in pH-dependent characteristics of the NP (Fig. 2). The hydrodynamic diameter was 197 ± 13.9 nm after 1 h incubation in a pH 4 solution and 169 ± 9.2 nm in a pH 7.4 solution for Ag10-NH2 (Fig. 2a). There was no difference in the results after 1 h and 24 h incubation. Under alkaline conditions (pH 10), the hydrodynamic diameter increased to 601 ± 85 nm (1 h) or 1911 ± 475 nm (24 h). Starting from the stock solution, with a positive surface charge of + 39.2 ± 0.2 mV, the surface charge decreased with increasing pH value (Fig. 2b). After 1 h incubation of Ag10-NH2 in aqueous pH 4 solution, the ζ-potential was + 36.9 ± 2.5 mV and decreased to − 7.4 ± 1.5 mV after 1 h incubation in a matrix with pH 10. But there was no significant difference between the results after 1 h and 24 h incubation.

PH-dependent hydrodynamic diameter and ζ-potential of Ag10-COOH (a, b) and Ag10-NH2 (c, d). N = 6
Ag10-COOH behaved contrarily. After 1 h incubation, the hydrodynamic diameter of Ag-COOH at pH 4, 7.4 and 10 was comparable. After 24 h the size significantly increased at pH 4, but not at pH 7.4 and 10 (Fig. 2c). The ζ-potential mainly did not vary significantly between the different pH values. After incubation for 24 h at pH 10 there was a significant increase in the surface charge to negative (− 54.1 ± 2.9 mV) (Fig. 2d).
Transmission electron microscopy revealed an averaged diameter of 5.7 ± 2.3 nm for Ag10-COOH (Fig. 3a). Their distribution among the grid was not homogeneous but rather arranged in clusters. Between the dark grey and black single particles there was a lighter colored layer with very small objects in it. This could be a residue from the production process. The single particles were shaped roundish to oval. The Ag10-NH2-NP showed an average diameter of 6.2 ± 3.4 nm (Fig. 3b). They were not distributed homogeneously among the grid and agglomerate to secondary particles with a diameter of over 100 nm. Beside these particles there were areas with single nanoparticles which had a roundish to oval shape and a plain surface.

Transmission electron microscope (TEM) image of Ag10-COOH (a) and Ag10-NH2 (b) in water
Impact of silver species, exposure time, concentration, soil texture and their combinations on biological parameters
Results of multi-factorial ANOVA revealed significant main effects of the factors silver species, concentration, exposure time and soil texture on the relative variation of the bacterial phyla Actinobacteria, Acidobacteria, alpha-Proteobacteria, Bacteroidetes and beta-Proteobacteria (Table 2). Comparable significant results were observed for the leucine aminopeptidase (LAP) activity, the microbial biomass, the abundance of all bacteria (16S rRNA), as well as for the ammonia-oxidizing bacteria (amoA) (Table 2). In case of the relative variation of the free-living nitrogen-fixing bacteria (nifH) only the factors and factor levels of exposure time and soil texture caused significant effects (Table 2). Silver species and concentration provoked no significant impacts on nifH (Table 2). The interaction effects of the factor combinations were predominantly significant for all variables (Table 2).
The considered main factors as well as their interactions could explain at least 81.5% (R2 = 0.815) of the variance of the respective target variables; in the case of beta-Proteobacteria even 93.9% (R2 = 0.939, Table 2).
Results of multivariate analysis revealed significant main effects for all factors and factor combinations (Table 2). MANOVA scored the factor combination soil texture × silver species as the strongest option (F = 53.2), followed by the combination of soil texture × exposure time (F = 46.8) (Table 2). Nevertheless, also the main factors silver species (F = 44.2), exposure time (F = 43.9) and soil texture (F = 46.5) could broadly explain the variance compared to the error variance itself (Table 2). The main factor concentration could elucidate the least variance (F = 12.7) (Table 2).
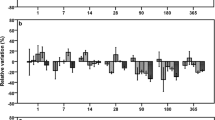
In Fig. 4 the influence of the highest scored factor combinations silver species × soil texture and exposure time × soil texture on the relative variation of the biological parameters are highlighted.

Impact of factor combinations on the entirety of the relative variation of the biological parameters. a Silver species × soil texture, b exposure time × soil texture. The error bars represent the 95% confidence interval. N = 4320. The relative variations (%) were set as dependent variable. The factors were set as independent variable. To calculate the impact of the factor combination silver species× soil texture on the relative variation of the biological parameters, the relative variations were unconnected to the factors concentration and exposure time (a). To calculate the impact of the factor combination exposure time × soil texture on the relative variation of the biological parameters, the relative variations were unconnected to the factors concentration and silver species (b)
The silver species Ag10-COOH caused similar effects in the loamy (101.2%) and the clayey (99.4%) soil, whereas Ag10-COOH diminished the relative variation of the biological parameters significantly in pairwise comparison to the sandy soil (93.3%; p = 0.000) (Fig. 4a). The effects of Ag10-NH2 were also very similar between the loamy (95.8%) and the clayey soil (98.0%), causing no significant differences. The pairwise comparison of clayey and sandy (94.0%) soil exhibited significant differences on the biological parameters (p = 0.003), while between sandy and loamy soil no differences could be detected. AgNO3 control slightly stimulated the entirety of the relative variation of the biological parameters in the loamy (101.2%) and sandy (103.6%) soil. In contrast AgNO3 caused a significant decrease of the relative variation (89.1%, p = 0.000) in the clayey soil. The order of increasing toxicity were for the loam Ag10-COOH = AgNO3 < Ag10-NH2, for the clayey soil Ag10-COOH = Ag10-NH2 < AgNO3 and for the sandy soil AgNO3 < Ag10-NH2 = Ag10-COOH.
As shown in Fig. 4b, a 1-day exposure to the silver species at different concentration led to a clear distinction between the effect in the sandy soil relative to the clayey (p = 0.029) as well as the loamy soil (p = 0.001). Here, the sandy soil exhibited the lowest toxicity. During the mid-term exposure of 14 and 28 days, this trend reversed and the sandy soil proved to be more toxic in response to treatment with silver compared to the loamy soil (p ≤ 0.05). However, after 90 days of silver exposure, an increase in the relative deviation of the biological variables from their untreated controls from the sandy to the clayey to the loamy soil could be observed (Fig. 4b). At this, there were only significant pairwise differences between the sandy and the loamy soil (p = 0.000), as well as between the sandy and the clayey soil (p = 0.001).
Impact of agent species, exposure time, concentration and their combinations on biological parameters in a loamy soil
Results of multi-factorial ANOVA revealed significant main effects of the factors agent species and exposure time on the relative variation of all biological target variables in the loamy soil (Table 3). In the case of the main factor concentration, only LAP activity, microbial biomass, amoA, Actinobacteria, Acidobacteria, and beta-Proteobacteria were significantly affected by the factor levels. The main factor exposure time was able to explain the largest part of the variance compared to the error variance of all target variable except for Acidobacteria and beta-Proteobacteria (Table 3). For these, the highest F-values were created by the main factor agent species. The main factor concentration could elucidate the least variance.
The majority of significant mean differences in a factor were found for Ag10-COOH and AgPure, 0.01 and 0.10 mg/kg silver as well as 0.01 and 1.00 mg/kg silver, and 1 day and 14 days, 14 days and 90 days as well as 28 days and 90 days (Table 3). No factor combination achieved higher variance explanations than the single main factors (Table 3).
Based on the results of the MANOVA, all factors and factor combinations revealed significant main effects (Table 3). The main factor exposure time clarified the largest amount of variance by a F-value of 56.0, followed by the main factor agent species (F = 18.8) (Table 3). The main factor concentration could elucidate the least variance again. The means of the highest scored individual factor levels of exposure time and agent species are visualized in Fig. 5.

Impact of the factors exposure time (a) and agent species (b) in a loamy soil. The error bars represent the 95% confidence interval. N = 2400. The relative variations (%) were set as dependent variable. The factors were set as independent variable. To calculate the effect of the exposure time on the relative variation of the biological parameters, the relative variations were calculated unconnected to the factors concentration and agent species (a). For the calculation of the effect expression of the factor agent species, the relative variations were unconnected to the factors concentration and exposure time (b)
The relative variation of the biological parameter in the presence of the different silver and nitrate concentrations changed significantly with the exposure time (p = 0.000) (Fig. 5a). After a decrease of the relative variation on day 1, the relative variation increased by 14.5% on day 14 to 109.4% (p = 0.000). Within the mid-term exposure, a similar, but negatively change of 10.8% could be observed at day 28 (p = 0.000). At the end of the experiment, a further, but not significant decline of the relative variation of the biological parameter could be observed (Fig. 5a).
By means of the entirety of the relative variation of the biological parameters the means of AgPure (96.4%) and Ag10-NH2 (95.8%) caused no significant pairwise disparity in their effect characteristic (Fig. 5b). This could also be stated for the means of Ag10-COOH (101.2%), AgNO3 (101.2%) and KNO3 (104.2%) (Fig. 5b). In contrast, AgPure and Ag10-NH2 each differed significantly from Ag10-COOH, AgNO3 and KNO3 in their pairwise comparisons (p ≤ 0.007).
The results of the factor combinations of the multivariate multi-factorial analysis revealed lower F-values than for the main factors agent species and exposure time individually. However, the factor combination agent species × exposure time was scored as the strongest option (F = 14.5) within the factor combination possibilities (Table 3). At this, 1 day exposure demonstrated Ag10-NH2 (89.8%) as the agent species with the highest influence on the relative variation of the biological parameter in the loamy soil, followed by Ag10-COOH (92.3%) (Fig. 6). AgPure and AgNO3 showed only small influences on the relative variation. Between AgNO3 and KNO3 no significant pairwise difference could be observed. At day 14, all agent species provoked an increase of the relative variation. The biological parameter were stimulated by 1.8% (AgPure) to 16.0% (AgNO3) (p = 0.000). Again, between AgNO3 and KNO3 no significant pairwise difference could be observed. After 4 weeks of exposure, the stimulating effect of all agent species weakened (Fig. 6). In the case of Ag10-COOH and KNO3 a decrease of the relative variation of 7.3% and 5.8%, respectively, could be observed. The strongest effects on the relative variation of the biological parameters due to the different agent species were created at day 90 (Fig. 6). The agent species Ag10-COOH and KNO3 provoked significant stimulations of the biological parameters of 12.5% and 15.1%, respectively (Table 4). In contrast, AgPure, AgNO3 and Ag10-NH2 significantly diminished the relative variation by 14.8, 16.3 and 17.8%, respectively (Table 4).

Impact of the factor combinations exposure time × agent species in a loamy soil. The error bars represent the 95% confidence interval. N = 240. The relative variations (%) were set as dependent variable. The factors were set as independent variable. To calculate the impact of the factor combination exposure time × agent species on the relative variation of the biological parameters, the relative variations were unconnected to the factor concentration
Discussion
The soil microbial community is responsible for several soil ecosystem services, like the recycling of organic matter, the soil fertility and structure, the biogeochemical nutrient cycles, the toxin degradation and the pathogen control [49,50,51,52,53]. Especially bacteria are the main performer of functional processes, which are integral for maintenance of healthy soil environments [54].
In this study, we used the DNA content of soil samples as a proxy for the microbial biomass and the abundance of 16S rRNA genes as an indicator of bacterial abundance. Furthermore, we measured LAP activity as well as the gene abundance of nifH and amoA as proxies for the nitrogen cycle, which drives many ecosystem activities in soils, including plant production [40, 41, 55]. Finally, we documented the response of Acidobacteria, Actinobacteria, alpha-Proteobacteria, Bacteroidetes and beta-Proteobacteria as representatives of the main soil bacterial phyla. Despite the phylogenetic diversity [56], not cultivability [57] and functional redundancy [58] still make it difficult to link members of bacterial phyla in soils with their function, some specific soil functions could be assigned to specific soil bacteria [41, 59,60,61,62,63].
The influence of freezing and defrosting is known to influence the structure and function of microbial communities [64, 65]. Nevertheless, this procedure was executed considering the high amount of samples to be processed. Since this method was applied to all samples, the results were comparable and give statistical inferences about the influence of the factors silver/agent species, concentration, exposure time and soil texture.
Silver/agent species
The main determining factor silver species caused almost significant effects on the investigated biological parameters of the three test soils (Table 2), whereby the main factor agent species caused significant effects on all investigated biological parameters in the loamy soil (Table 3).
In a simultaneous consideration of all biological parameters by the MANOVAs, an increase in the toxicity of KNO3 (104.2%) = Ag10-COOH (98.0%/101.2%) = AgNO3 (97.95%/101.2%) < AgPure (96.4%) = Ag10-NH2 (95.9%/95.8%) could be observed, whereby between Ag10-COOH and AgNO3 no significant pairwise difference could be calculated (Additional file 1: Fig. S1). Values of the main factor agent species are shown in italic.
The coating of NP is crucial for their reactivity and physico–chemical transformations, such as the dissolution rate, the availability of surface areas, the surface charge, the aggregation rate and the long-term stability [27, 66, 67]. Analysis of the ζ-potential of the AgNP under different pH values (Fig. 2b, d), indicated no significant impact of pH on ζ-potential of the Ag10-COOH particles. The high negative ζ-potentials of − 33.9 ± 1.8 mV at pH 4 and − 39.4 ± 1.5 mV at pH 7.4 of the Ag10-COOH indicate high stability of these nanoparticles. The COOH-coating could minimize Ag+ ion release as well as direct contact of the AgNP with microorganisms or soil compartments such as clay particles or natural organic matter due to their function as a physical barrier [27, 31, 66]. The investigation of Long et al. [68] confirmed this hypothesis. They examined the Ag+ release and toxicity of different coated AgNP which were very similar to our Ag10-COOH particles and observed only a slightly release of Ag+ and a lower associated toxicity to Escherichia coli. In addition, the negative surface charge could led to an attachment of soil cations such as Ca2+, Mg2+ or K+ on the Ag10-COOH particles and an increase of their physico–chemical barrier.
Furthermore, the coating of AgNP plays a crucial role in determining their cellular uptake mechanism [26], where the negative surface charge of Ag10-COOH indicates a low affinity to negatively charged membranes of microorganisms. Nevertheless, Ag10-COOH induced both adverse and advantageous effects in view of the investigated individual biological parameters. Here, individual defense strategies [28, 69] as well as species dependent toxicological susceptibility [17, 52, 70] of the biological parameters might be the underlying reasons.
In contrast, the ζ-potential of Ag10-NH2 decreased with increasing pH in our experiment (Fig. 2b) and consequently a decrease of stability could be deduced in the test soils. As a consequence of the missing physico–chemical barrier a high availability of AgNP surface area as well a high release rate of Ag+ ions is expectable. Both, the mean of the relative variation of all biological parameters (95.9%/95.8%) as well as the mean of LAP activity (96.5%/95.1%), 16S rRNA (95.7%/92.0%), amoA (92.6%/92.9%), Acidobacteria (89.9%/78.7%) and Bacteroidetes (93.5%/96.6%) (Table 2, 3) underlined the toxic effects of Ag10-NH2. Moreover, the positive surface charge of Ag10-NH2 could have led to a strong association with the negatively charged membranes of microorganisms [71, 72] and affect the surface tension of the membrane resulting in an increase of pore formations [26].
AgPure showed similar harmful effects such as Ag10-NH2, which could be also attributed to release of Ag+ ions. Their low particle concentration, their polyacrylate stabilization and the high pH value of soil could have prevented initial aggregation and agglomeration of AgPure [16, 17, 73]. In addition, the high concentration of divalent cations, such as Ca2+ und Mg2+ in the loamy soil (Table 1) could promote AgPure and Ag10-NH2 dissolution, resulting in the displacement of Ag+ ions from the nanoparticle surface and thus toxicity [74].
Surprisingly, the AgNO3 control documented low toxicity by average 97.95% and 101.2% of the relative variation. The use of silver nitrate is ubiquitously as control in toxicological studies with AgNP to estimate the influence of dissolved Ag+ ions from AgNP [1, 4, 75, 76]. Here, the toxicity order of the MANOVAs suggest a high release of Ag+ ions due to Ag10-COOH (98.0%/101.2%) relative to Ag10-NH2 (95.9%/95.8%) and AgPure (96.4%) and thus a high direct toxic impact of Ag10-NH2 and AgPure NP itself. However, an individual view on the biological parameters with regard to the ANOVAs resulted in a different manner. The bacterial abundance (S16 rRNA) and the abundance of Actinobacteria were stimulated by Ag10-COOH exposure and diminished by AgNO3, whereas LAP activity, microbial biomass and Acidobacteria were stimulated or not influenced by AgNO3 and diminished by Ag10-COOH (Table 2). These individual responses of the biological parameters explain the majority of significant mean differences found between Ag10-COOH and AgNO3 within the main factors silver species. By averaging MANOVA, these important observations are lost and could have led to deceptive conclusions. Based on the individual ANOVAs (Tables 2, 3), as well as the particle and soil characterizations, the hypothesis of a high release of Ag+ by Ag10-COOH should be excluded and could be related rather to Ag10-NH2 and AgPure.
In detail, the presented results in Tables 2 and 3 revealed no or stimulation effects due to AgNO3 in case of LAP, amoA, Acidobacteria and Bacteroidetes, whereas the used AgNP caused predominantly lower relative variations of these biological parameters. Similar observations were documented in short-term studies dealing with the effect of low concentrations of AgNP and AgNO3 on organisms related to nitrogen cycle, where AgNO3 might cause lower or even stimulating effects relative to the AgNP. Quite recently, we observed stimulating effects by AgNO3 to ammonia-oxidizing and nitrogen fixing bacteria after short-term exposure, whereby AgNP led to their decrease [16]. Schlich et al. [77] documented a significant stimulation of the nitrite production after 7 day exposure to 0.19 mg/kg AgNO3 up to 19.4%, whereby AgNP caused an inhibition. Yang et al. [78] found that 2.5 µg/L of Ag+ as AgNO3 upregulated AMOA genes amoA1 and amoC2 by 2.1 by 3.3-fold. Furthermore, Choi et al. [79], Masrahi et al. [24] and Liang et al. [80] observed lower effects on microbial nitrification due to AgNO3 relative to AgNP after short-term exposure. Here, in view of the very similar effect expressions of AgNO3 (101.2%) and KNO3 (104.2%) we suspect, that the nitrate contained in the AgNO3 might also impact the biological parameter such as the silver itself in the AgNO3 control. LAP activity and amoA are proxies for the nitrogen cycle. Because of the spatial structure imposed by soil particles resulting in local variations in oxygen availability over small distance [81], both aerobic and anaerobic conditions can be found in the same soil sample. Thus, AgNO3 control could act as substrate for nitrate reducing bacteria, such as gamma-Proteobacteria or Acidobacteria [63, 82, 83], which were less sensitive to AgNP or Ag+ in form of AgNO3 [78, 84, 85] and promoted their increase due to nitrate utilization via denitrification and/or dissimilatory nitrate reduction [81, 83] resulting in new nitrogen sources for the ammonia-oxidizing and nitrogen fixing bacteria.
The increase of the abundance of Bacteroidetes might be the result of harboring silver resistance genes [69] and simultaneously increased availability of carbon through decreased silver-sensitive microbes, which promoted the as r-strategist know Bacteroidetes [56]. Nevertheless, the average factor expression of AgNO3 in case of Actinobacteria (90.8%/88.7%) and beta-Proteobacteria (94.0%/94.3%) (Tables 2, 3) indicated still harmful effects. Consequently, there could be an interplay of stimulating and detrimental effects, which might underlay in the agent itself, but also in consequential shifts of the microbial community and nutrient availability.
The F-values of the biological parameters in both MANOVAs indicated the main factors silver species and agent species as strong options to explain large parts of the variances compared to the error variances (Tables 2, 3). With respect to the three different used AgNP, our results confirmed a distinctive impact of the AgNP functionalization on their fate and toxicity in the soil environment. The general applicability of AgNO3 as suitable positive control should be subject of further investigations.
Concentration
The main determining factor concentration caused as well almost significant effects on the investigated biological parameters in the three test soils (Table 2). The majority of significant mean differences in the factor were found for 0.01 and 1.00 mg/kg silver. Here, the toxicity of the silver species increased predominantly with increasing concentrations (Additional file 1: Fig. S2), which was already observed in a variety of recent soil studies [5, 24, 86, 87].
The concentration of AgNP strongly influences their physico–chemical transformation. Once released into the environment, AgNP concentration is crucial for dissolution and aggregation mechanisms [28]. Merrifield et al. [63] documented AgNP homoaggregation as insignificant at realistic environmental concentrations, whereby dissolution and also heteroaggregation processes were more likely. Based on the low test concentrations of 0.01–1.00 mg/kg silver, dissolution seemed to be the most probable transformation type in our study. Dissolution hypothesis was supported by the average high pH value of soils that could have prevented initial aggregation and agglomeration of AgNP [73] as well as the average high concentration of divalent cations, such as Ca2+ und Mg2+ (Table 1), which promoted AgNP dissolution and resulted in the displacement of Ag+ ions from the nanoparticle surface [74]. Furthermore, the high ζ-potentials of + 36.8 and + 39.2 mV for Ag10-NH2 as well as of − 33.9 and − 39.4 mV for Ag10-COOH at pH 4 and 7.4, respectively, indicated high interparticular repulsive forces, which diminished the aggregation probability due to high stability [88]. However, according to Klitzke et al. [14] a decrease of ζ-potential after AgNP exposure to soil solution has to be assumed.
A direct interaction of the microbes with the differently charged AgNP is also conceivable. However, the relative variation of the biological parameters differed only by 2.0% between 0.01 and 1.00 mg/kg silver. This low effect expression can be attributed to the low bioavailability of the silver species as well as to bacterial resistance mechanisms. The released silver ions could bind to clay particles [19, 33] or organic material [20, 89, 90] and thus be less bioavailable. In addition, the likelihood of a microbe to encounter a silver particle or ion is generally low concerning the applied low test concentrations. In the event of a coincidence, bacteria possess various commonly defense mechanisms to escape the toxic influence of silver, such as the thickness of their peptidoglycan-membrane as the first line of defense [91], efflux systems to extrude heavy metal ions [4, 92, 93] and the production of extracellular proteins and exopolysaccharide [94] to neutralize small amounts of toxic compounds [95, 96]. Furthermore, some bacteria obtain specific silver resistance genes, which encode periplasmic silver-specific binding protein (SilE), silver efflux pumps (P-type ATPase), and membrane potential-dependent polypeptide cations/proton antiporter (SilCBA) [4].
Nevertheless, compared to the F-values of the other main factors, the factor concentration could elucidate the least variance of the mean values considering all variables (Tables 2, 3). Based on our results, the main factor concentration at environmentally relevant levels has, therefore, only a very weak influence on the AgNP effect characteristics respecting the microbial community in soils.
Exposure time
The main determining factor exposure time caused consistently significant effects on the investigated biological parameters in the three test soils after silver exposure (Table 2), with the majority of significant mean differences between the factor expressions 1 day and 28 d. With the exception of the relative variation of the Bacteroidetes and the ammonia-oxidizing bacteria (amoA), all biological parameter were negatively affected at day 1 (Table 2). This observation was rather unusual, because several studies measured high and fast sensitivity of ammonia-oxidizing bacteria with AgNP and AgNO3 [24, 77, 78]. Apart from that, we recently recognized a tolerance of amoA harboring bacteria after short-term exposure to AgNP [16]. Furthermore, Schlich et al. [77] and Samarajeewa et al. [86] observed stimulatory effects with ionic and nanoparticulate silver to ammonia-oxidizing bacteria, which could be ascribed to hormone-like responses to low silver concentrations. By contrast, the stimulation of Bacteroidetes due to silver addition was in agreement with previous observations [52, 69, 70, 97, 98]. They exhibit silver resistance genes [78].
As the exposure time increased until day 28, the injury on the microbial community decreased (Additional file 1: Fig. S3). It is likely that the short-term effects after 1 day were observed as a result of the initial release of bioavailable Ag+ by AgNP and AgNO3, causing toxic effects on the microbial community. Dissolution experiments [99, 100] verified fast solubility of AgNP in soils. With increasing exposure time, the silver species became less bioavailable due to interactions with organic matter, clay minerals or pedogenic oxides [19, 33]. Furthermore, upcoming mechanisms such as self-protection [4, 91,92,93, 95, 96], resistance [4], resilience [58] and/or cryptic growth [101], might also be possible explanations for the limited effects on the microbial community in the soils.
Results of the ANOVA using data of the loamy soil only documented a similar trend (Table 3). Here, all biological parameters were also negatively affected at day 1, with the exception of the relative variation of the Bacteroidetes and the ammonia-oxidizing bacteria (amoA). In virtue of the high clay content (approximately 30%) of the loamy soil and its high content of organic carbon (2.9%) (Table 1), the retention of AgNP and Ag+ ions arose earlier and the toxicity of the agents decreased even at day 14 (Table 3) [14, 20, 102, 103].
On day 90, an impairment of the microbial community could be observed relative to day 28 for both ANOVAs (Tables 2, 3). Similar trends were observed in our previous studies [16, 17]. As AgNP and Ag+ ions gradually aged, they slowly returned to the biological soil system as a continuously sink of silver agents. Diez-Ortiz et al. [33] also documented a progressive increase in AgNP toxicity with time and assumed a time-dependent enlargement of silver in soil pore water due to slow dissolution.
The F-values of the biological parameters in both MANOVA indicated the main factor exposure time as a strong option to explain a large part of the variances compared to the error variances (Tables 2, 3). Especially in case of the exclusion of the factor soil texture from the MANOVA, the factor exposure time yielded the maximum F-value (Table 3). These results strongly underline the significance of the exposure time for AgNP ecotoxicity investigations, attributable to the changes of silver bioavailability and its specification status during long-term experiments in soils.
Soil texture
Also the main factor soil texture caused significant effects on all investigated biological parameters in the three test soils (Table 2). The majority of significant mean differences were found between the loamy soil and the clayey soil. This was noticeable, because their investigated soil characteristics such as pH, TOC, TN and CEC were very similar. The most distinct difference resulted from their grain size distributions of clay (45–65% vs. 25–35%) and sand (5–40% vs. 25–45%) (Table 1). Actually, recent studies have shown a positive correlation between the clay content of soils and the toxicity of AgNP: lower grain size of soils resulted in lower toxicity of AgNP [19, 23, 25]. Indeed, this was appropriate for the sandy soil with a mean of 97.0% and the loamy soil with a mean of 99.4% relative abundance of all biological parameters (Additional file 1: Fig. S4). However, the mean of the relative abundance of the clayey soil (95.5%) illustrated the opposed situation: lower grain size of the clayey soil resulted in higher toxicity of silver (Additional file 1: Fig. S4). In fact, not only the particle size distribution can be responsible for the silver toxicity. Schlich and Hund-Rinke [19] documented that the highest AgNP toxicity was associated with more acidic soils, whereas the lowest toxicity was associated with more alkaline soils. They supposed that the soil pH value influences AgNP dissolution and the release of ions [19]. Similarly, this could be observed for the loamy soil (pH = 7.2) and the sandy soil (pH = 3.2) in this study (Table 2). At the same time, the results of the clayey soil with a pH value of 7.4 seemed to disagree again with the hypothesis.
The results of the factor combination silver species × soil texture (Fig. 4a) might elucidate the crux of the observed mean silver toxicity order loamy soil < sandy soil < clayey soil. In case of Ag10-NH2 and Ag10-COOH, the loamy and the clayey soil exhibited lower toxicity compared to the sandy soil (Fig. 4a). This confirmed the positive correlation between the grain size distribution of soils and the toxicity of AgNP as well as the association of high AgNP toxicity by more acidic soils [19, 23, 25]. In contrast, AgNO3 caused a strong decrease of the entirety of the relative variation of the biological parameters in the clayey soil (Fig. 4a), which caused an overwhelmed influence on the average formation in the MANOVA of the silver species. This indicated a completely different fate of AgNO3 compared to AgNP in the test soils.
Soil texture× silver species
The factor combination soil texture × silver species yielded the highest F-value in the MANOVA considering the entirety of the relative variation of the biological parameters (Table 2).
With regard to the investigated AgNP it could be observed that their toxicity was lower in the loamy and clayey soil than in the sandy soil (Fig. 4a). In case of Ag10-NH2 the high availability of soil cations in the loamy and the clayey soil (Table 1) might have promoted the dissolution of the lower stable AgNP [14], resulting in the release of Ag+ ions. These were bound to clay particles and became less bioavailable [33]. The absent effect of the AgNO3 control as measure for Ag+ impact in the loamy soil might confirm this hypothesis. Furthermore, the high content of natural organic matter, derived by the land-uses of the loam and clay locations, could have been reduced the toxicological effects of the Ag10-NH2 due to inhibition of oxidation [104]. At this, positive charged Ag10-NH2 were adsorbed by negatively charged organic matter, which dominated now the surface properties leading to higher steric stability and thus also lower bioavailability [18].
In contrast, the sandy soil exhibited contrary soil properties and thus toxicity (Table 1). Although the low amount of soil cations reduced the dissolution affinity of AgNP, the low pH of 3.2 and the concomitant higher amount of OH− ions might decreased the Ag10-NH2 stability due to the deprotonation of the amino groups. Under aerobe conditions, AgNP oxidation followed and Ag+ ions were released [104]. The small clay content of the sandy soil (0.0–5.0%) did not provide enough capacity for Ag+ retention resulting in direct harmful interactions of Ag+ with the microorganisms, such as interactions with enzymes of the respiratory chain reaction, increase of DNA mutation frequencies or morphological changes of cell wall membrane [105]. However, the stimulating effects of the AgNO3 control on the biological parameters in the sandy soil disagreed with the Ag+ dissolution theory (Fig. 4a). Therefore, it might be that the Ag10-NH2 particle itself caused the negative effects in the sandy soil. Positively charged NP were observed to strongly associate with membranes, which leads to a higher cellular uptake [26].
The lower toxicity of the Ag10-COOH relative to the Ag10-NH2 particles based in general on their higher stability and their surface barrier, which reduced Ag+ dissolution. Furthermore, their negative surface charge promoted the attachment of soil cations of the loamy and the clayey soil and could have prevented direct interactions with negatively charged membranes of microorganisms [26, 27, 31, 66, 68]. Analogously to the Ag10-NH2, Ag10-COOH showed strongest toxicity in the sandy soil (Fig. 4a). Here, interactions with soil cations as well as natural organic material seemed to be less probable considering the soil and AgNP characteristics (Table 1). In addition to a low oxidation of Ag10-COOH and the concomitant lower Ag+ release, a direct interaction of these AgNP with microorganisms can be assumed. An internalization of negatively charged nanoparticles could be occured through nonspecific binding and clustering of the particles on cationic sites on the plasma membrane and subsequent endocytosis or by direct diffusion through the cell membrane [106]. Here, Ag10-COOH could act as a trojan horse: metabolization processes in food vacuoles and lysosomes allow the uncoating of AgNP and enable the direct release of Ag+ ions into the cytoplasm causing intracellular damages [32, 107].
However, there is always a challenge to accurately differentiate what proportion of the toxicity is from the ionic form and what proportion of the nanoform [76]. For resolving this question, AgNO3 was used as measure for Ag+ impact in our study. Results of silver nitrate exposure in the loamy soil confirmed their low contribution to the AgNP toxicity (Fig. 4a). Ag+ ions seemed to be bound to clay particles or other soil compartments and became less bioavailable. Conspicuously, AgNO3 caused the highest toxicity in the clayey soil and a low stimulation in the sandy soil. In fact, Schlich and Hund-Rinke [19] observed also a lower toxicity of AgNO3 compared to AgNP in a sandy soil (RefeSol 04A) investigating the potential ammonium oxidation, but there was no promotion. The high toxicity of AgNO3 in the clayey soil resulted by a decrease of the bacterial taxa, whereas LAP activity, nifH and amoA were hardly influenced or even stimulated (data not shown). In case of the proxies for nitrogen cycle, we suspect a significant portion of the nitrate compound of the AgNO3 control in the results. However, reasons for the detrimental effect on the bacterial structure were intricate to find. Probably, interactions with existing soil contaminants could be a reason for the high AgNO3 toxicity. The clay location was farmed and treated with herbicides and pesticides. With this, AgNO3 might have been bound to such contaminants, thus creating a new toxic agent. For example, Uwizeyimana et al. [108] indicated that pesticide and metal mixtures negatively affected earthworms. However, detailed studies on the impact of nanoparticle-pesticide combinations on microbial community in soils are currently lacking, but necessary to perform a comprehensive risk assessment of AgNP in soils.
Soil texture× exposure time
The factor combination soil texture × exposure time revealed the second highest F-value of 46.8 in the MANOVA (Table 2). With regard to the results in Fig. 4b, it could be clearly observed, that the spans of the effect characteristics of the individual soils differ significantly from each other during the four examination dates. While the effect levels in the sandy soil on days 1, 14, 28 and 90 extended over an amount of 6.3% (lowest value of 93.7% on day 14, highest value of 100.0% on day 90), the amounts for the loamy and the clayey soil were 21.5% and 20.3%, respectively. This indicated the loamy and the clayey soil as more complex soil systems considering the interplay of physico–chemical and biological interactions and transformations of the silver species contrary to the sandy soil. Whereas the effect expressions of the biological parameters in the sandy soil were indicative for consistent conditions, the effect expressions of the loamy and the clayey soil suggested a variety of temporal changes in the complex soil-time framework.
In case of the sandy soil it can be assumed, that there were continuously barely interactions of the silver species with the soil type characteristics, causing throughout similar effect expressions. In addition, the indigenous microbial community might show no observable adaption ability over time.
In contrast, the relative variations of the biological parameters in the loamy and the clayey soil differed not only in dependence on the exposure time, but also in their effect strength from each other, although their soil characteristics were very similar (Table 1). For example, on day 14, it could be assumed for the loamy soil, that the silver species were transformed, might be bound to the clay particles or capped by organic matter [33, 104], resulting in a decrease of toxicity and a stimulation of the biological parameters due to their defense arsenal [4, 91,92,93, 95, 96] (Fig. 4b). Based on the similar soil characteristics, this should be also noted for the clayey soil. However, the biological parameter was diminished by 14.0% in the clayey soil at day 14 suggesting divergent indigenous microbial communities with different capacities to resist and adapt on silver emission. Also interactions with herbicides and pesticides of the clayey soil might have led to the creation of a new toxicant, which provoked the harmful effect. At day 28, the effective expressions of the loamy and the clayey soil reproached and none or stimulating effects were documented (Fig. 4b). At this, ageing of the silver species and their return to the biological soil system could have reduced the stimulating effects on the biological parameters in the loamy soil. Contrary, the ageing in the clayey soil of the silver species or the new toxicants as well as the now developed defense mechanisms of the microbial community after 4 weeks exposure caused stimulation by 6.3% (Fig. 4b).
Based on our data, however, we can only speculate about these events. More detailed investigations are necessary to reveal the time-dependent fate of the silver bioavailability in different soils, as well as the time-dependent responses of the microbial community to these silver species. For this purpose, the development of reliable and robust analytical methods for detecting specific silver species at trace concentrations in complex matrices is a fundamental requirement. Next, batch experiments can reveal time-dependent silver transformations and the concomitant effects on the microbial community. Furthermore, it would be useful to further deepen the soil characterization to evaluate possible effects of anthropogenic residues from e.g. herbicides or pesticides on the ecotoxicological potential of silver nanoparticles.
Agent species× exposure time
Regarding the MANOVA with the main factors agent species, concentration and exposure time in the loamy soil, the factor exposure time was able to explain the largest part of variance. The factor combinations achieved lower F-values. Nevertheless, the factor combination agent species × exposure time still achieved the highest F-value of 14.5 for the factor combinations. Furthermore, the results of the pairwise comparisons of the post hoc test (Table 4) as well as the ambiguous role of AgNO3 control as a measure of Ag+ dissolution gave reason to pay attention to this interaction.
Short-term exposure led only to a small difference between the investigated agents AgPure, Ag10-COOH, Ag10-NH2, AgNO3 and KNO3 (Fig. 6). Ag10-COOH diminished the relative variations of the biological parameters by 7.7%. Based on their higher stability and the short exposure time, physico–chemical transformation could be excluded and led to the assumption of direct harmful interaction of these AgNP with microorganisms. In contrast, Ag10-NH2 was reported as strong instable AgNP due to their surface functionalization. At this, higher dissolution of Ag+ ions as toxicological agent might be a probable explanation for the decrease of the biological parameters by 10.2% (Fig. 6). However, relating to the low effects of the AgNO3 control (97.95%), the Ag+ ion dissolution theory seemed less likely. Therefore, a direct interaction could be assumed as well. AgPure caused on average no effects (100.7%) on day 1 and thus proved to be inert to physico–chemical transformations after short-term exposure. The KNO3 control diminished the biological parameters on average by 6.3%, but without significant difference to AgNO3 (Fig. 6, Table 4). This slightly adverse effect might be the result of NO2− accumulation, due to nitrate reduction [109, 110].
After 14 days exposure, the differences between the effect characteristics of the agent species changed. Ag10-NH2 (111.2%), AgNO3 (116.0%) as well as KNO3 (112.0%) stimulated the biological parameter significantly, whereas AgPure (101.8%) and Ag10-COOH (107.0%) caused only low stimulatory effects. In case of the silver species, their bioavailability as well as those of the released Ag+ ions could be reduced at this time point due to interactions with organic matter, clay minerals or pedogenic oxides [14, 20, 102, 103]. Furthermore, emerging self-protection mechanisms, such as the production of extracellular proteins or polysaccharides of the soil microbiome could neutralize toxic ions or cap AgNP [95, 96]. In addition, resilience mechanisms [58, 78, 101] might be possible explanations for the limited effects on the microbial community in the soil at day 14. The increase of the biological parameter due to AgNO3 and KNO3 exposure might result by an increase of nitrate reduction and the concomitant increase of nitrogen for the microbiome [81, 83]. Due to the synergy of microbial silver resistance mechanisms as well as the nitrogen substrate source, AgNO3 increased the relative variation of the biological parameters. As already mentioned, several authors documented stimulation or minor negative effects on microorganisms related to nitrogen cycle after exposure of low concentrations of AgNO3 in short-term exposure experiments [16, 24, 77,78,79,80].
Starting by day 28, ageing of the silver species and their slow return to the biological soil system presented a continuous sink of bioavailable silver, reducing the stimulating effects on the biological parameters (Fig. 6). An increase in AgNP toxicity with time can be linked to time-dependent enlargement of silver in soil pore water due to dissolution [33, 111]. However, the defense arsenal of bacteria was sufficient to resist silver toxicity at day 28. After 3 months exposure, there seemed to be a shock load of silver in case of AgPure (85.2%), Ag10-NH2 (82.4%) and AgNO3 (83.8%) to which the bacterial community was not immediately prepared. Small-scale bioavailability, chemical alterations and possible transformations (oxidation, reduction, dissolution, sulfidation) of AgNP and Ag+ [23, 24, 33, 90, 112, 113] in the loamy soil are possible physicochemical causes for the abrupt toxicity. Furthermore, it might be assumed that after short- and mid-term adaption to the silver contamination as well as the positioning of the silver species in the soil system, the bacterial population might have lost members with silver tolerance and were unanticipatedly shocked by the return of silver toxicant at day 90 resulting in strong reductions of the biological parameters [17].
In contrast, Ag10-COOH caused a significant stimulation of the investigated parameters by 12.5%, confirming their high stability. With prolonged exposure time, the likelihood of bounds between the negative charged Ag10-COOH and soil cations increased and with that their physico–chemical barrier, resulting in lower bioavailability. Together with the presumably low number of free Ag+ ions in case of Ag10-COOH, no toxicity could be observed.
Based on the missing significant differences of AgNO3 and KNO3 at days 1 and 14, it could not be determined, if the effects were caused only by the Ag+ of the AgNO3 control, but maybe due to the NO3− of the KNO3 control. First at the exposure times of 28 days and 90 days, significantly different effect characteristics of AgNO3 and KNO3 by 13.0% (p = 0.000) and 31.5% (p = 0.000) (Fig. 6, Table 4), respectively, could be observed. This caused us to suspect, that initial at this late time points the nitrate contained in the AgNO3 control did not further exceedingly influence the results of the AgNO3 control. In consequence, especially in case of biological parameters related to nitrogen cycle, such as LAP activity or the abundance von amoA harboring bacteria, the usage of AgNO3 as proxy for Ag+ release in AgNP short-term effect assessments could be deceptive. There is an urgent need for further research into the suitability of AgNO3 as a measure of Ag+ release. For example, batch experiments investigating all steps within the soil nitrogen cycle (in particular nitrogen fixation, nitrification, denitrification, dissimilatory nitrate reduction to ammonium) after short- and long-term exposure of AgNO3 and NO3− will be helpful to resolve in detail, at which step the nitrate released from the AgNO3 control presents an advantage for the microbial community and at which step the harmful influence of the silver predominates.
Conclusions
Impacts of the factors silver species, agent species, concentration, exposure time and soil texture on the relative variation of 10 biological parameters were analysed by 16S rRNA qPCR as well as fluorometric and photometric techniques. The used AgNP were characterized in detail by electron microscopy, dynamic light scattering and asymmetrical flow field-flow fractionation. Analyses of variance revealed the factors silver species, exposure time and soil texture as the most relevant determinants for the effect expressions of the biological parameters. Furthermore, the factor combinations soil texture × silver species as well as soil texture × exposure time facilitates even larger explanations of the variance of the biological parameter.
Overall, the presented results demonstrate the importance of considering several factors in the effect assessment of AgNP. Based on our study, the relationship of the soil texture × silver species is the most significant factor combination for the environmental fate and toxicities of AgNP in soils.
Abbreviations
- AgNP:
-
silver nanoparticle(s)
- AF4:
-
asymmetrical flow field-flow fractionation
- Ag-HPB:
-
hydrophobic silver particle(s)
- DLS:
-
dynamic light scattering
- MPA:
-
mercaptopropionic acid
- NP:
-
nanoparticle(s)
- PDI:
-
polydispersity index
- UPW:
-
ultrapure water
References
Reidy B, Haase A, Luch A, Dawson K, Lynch I (2013) Mechanisms of silver nanoparticle release, transformation and toxicity: a critical review of current knowledge and recommendations for future studies and applications. Materials 6:2295–2350
Sun TY, Mitrano DM, Bornhöft NA, Scheringer M, Hungerbühler K, Nowack B (2017) Envisioning Nano release dynamics in a changing world: using dynamic probabilistic modeling to assess future environmental emissions of engineered nanomaterials. Environ Sci Technol 51:2854–2863
Abbasi E, Milani M, Fekri Aval S, Kouhi M, Akbarzadeh A, Tayefi Nasrabadi H, Nikasa P, Joo SW, Hanifehpour Y, Nejati-Koshki K (2016) Silver nanoparticles: synthesis methods, bio-applications and properties. Crit Rev Microbiol 42:173–180
Pareek V, Gupta R, Panwar J (2018) Do physico–chemical properties of silver nanoparticles decide their interaction with biological media and bactericidal action? A review. Mater Sci Eng C 90:739–749
Hänsch M, Emmerling C (2010) Effects of silver nanoparticles on the microbiota and enzyme activity in soil. J Plant Nutr Soil Sci 173:554–558
Gottschalk F, Kost E, Nowack B (2013) Engineered nanomaterials in water and soils: a risk quantification based on probabilistic exposure and effect modeling. Environ Toxicol Chem 32:1278–1287
Nowack B, Baalousha M, Bornhöft N, Chaudhry Q, Cornelis G, Cotterill J, Gondikas A, Hassellöv M, Lead J, Mitrano DM (2015) Progress towards the validation of modeled environmental concentrations of engineered nanomaterials by analytical measurements. Environ Sci Nano 2:421–428
Sun TY, Gottschalk F, Hungerbühler K, Nowack B (2014) Comprehensive probabilistic modelling of environmental emissions of engineered nanomaterials. Environ Pollut 185:69–76
Gottschalk F, Sonderer T, Scholz RW, Nowack B (2009) Modeled environmental concentrations of engineered nanomaterials (TiO2, ZnO, Ag, CNT, fullerenes) for different regions. Environ Sci Technol 43:9216–9222
Gottschalk F, Nowack B (2011) The release of engineered nanomaterials to the environment. J Environ Monit 13:1145–1155
Benn TM, Westerhoff P (2008) Nanoparticle silver released into water from commercially available sock fabrics. Environ Sci Technol 42:4133–4139
Sun TY, Conroy G, Donner E, Hungerbühler K, Lombi E, Nowack B (2015) Probabilistic modelling of engineered nanomaterial emissions to the environment: a spatio-temporal approach. Environ Sci Nano 2:340–351
Dale A, Casman E, Lowry G, Lead J, Viparelli E, Baalousha M (2015) Modeling nanomaterial environmental fate in aquatic systems. Environ Sci Technol 49:2587
Klitzke S, Metreveli G, Peters A, Schaumann GE, Lang F (2014) The fate of silver nanoparticles in soil solution—sorption of solutes and aggregation. Sci Total Environ 535:54–60
Holden PA, Schimel JP, Godwin HA (2014) Five reasons to use bacteria when assessing manufactured nanomaterial environmental hazards and fates. Curr Opin Biotechnol 27:73–78
Grün A-L, Straskraba S, Schulz S, Schloter M, Emmerling C (2018) Long-term effects of environmentally relevant concentrations of silver nanoparticles on microbial biomass, enzyme activity, and functional genes involved in the nitrogen cycle of loamy soil. J Environ Sci 69:12–22
Grün A-L, Emmerling C (2018) Long-term effects of environmentally relevant concentrations of silver nanoparticles on major soil bacterial phyla of a loamy soil. Environ Sci Eur 30:1–13
Cornelis G, Hund-Rinke K, Kuhlbusch T, Van den Brink N, Nickel C (2014) Fate and bioavailability of engineered nanoparticles in soils: a review. Crit Rev Environ Sci Technol 44:2720–2764
Schlich K, Hund-Rinke K (2015) Influence of soil properties on the effect of silver nanomaterials on microbial activity in five soils. Environ Pollut 196:321–330
Settimio L, McLaughlin MJ, Kirby JK, Langdon KA, Janik L, Smith S (2015) Complexation of silver and dissolved organic matter in soil water extracts. Environ Pollut 199:174–184
Bundschuh M, Filser J, Lüderwald S, McKee MS, Metreveli G, Schaumann GE, Schulz R, Wagner S (2018) Nanoparticles in the environment: where do we come from, where do we go to? Environ Sci Eur 30:1–17
Lowry GV, Gregory KB, Apte SC, Lead JR (2012) Transformations of nanomaterials in the environment. Environ Sci Technol 46:6893–6899
Pachapur VL, Larios AD, Cledón M, Brar SK, Verma M, Surampalli R (2016) Behavior and characterization of titanium dioxide and silver nanoparticles in soils. Sci Total Environ 563:933–943
Masrahi A, VandeVoort AR, Arai Y (2014) Effects of silver nanoparticle on soil-nitrification processes. Arch Environ Con Tox 66:504–513
Rahmatpour S, Shirvani M, Mosaddeghi MR, Nourbakhsh F, Bazarganipour M (2017) Dose-response effects of silver nanoparticles and silver nitrate on microbial and enzyme activities in calcareous soils. Geoderma 285:313–322
Saei AA, Yazdani M, Lohse SE, Bakhtiary Z, Serpooshan V, Ghavami M, Asadian M, Mashaghi S, Dreaden EC, Mashaghi A (2017) Nanoparticle surface functionality dictates cellular and systemic toxicity. Chem Mat 29:6578–6595
Louie SM, Tilton RD, Lowry GV (2016) Critical review: impacts of macromolecular coatings on critical physicochemical processes controlling environmental fate of nanomaterials. Environ Sci Nano 3:283–310
Lead JR, Batley GE, Alvarez PJ, Croteau MN, Handy RD, McLaughlin MJ, Judy JD, Schirmer K (2018) Nanomaterials in the environment: behavior, fate, bioavailability, and effects—an updated review. Environ Toxicol Chem 27:2029–2063
Datta LP, Chatterjee A, Acharya K, De P, Das M (2017) Enzyme responsive nucleotide functionalized silver nanoparticles with effective antimicrobial and anticancer activity. New J Chem 41:1538–1548
Fröhlich E (2012) The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int J Nanomed 7:1–15
Wu F, Harper BJ, Harper SL (2017) Differential dissolution and toxicity of surface functionalized silver nanoparticles in small-scale microcosms: impacts of community complexity. Environ Sci Nano 4:359–372
Grün A-L, Scheid P, Hauröder B, Emmerling C, Manz W (2017) Assessment of the effect of silver nanoparticles on the relevant soil protozoan genus Acanthamoeba. J Plant Nutr Soil Sci 180:602–613
Diez-Ortiz M, Lahive E, George S, Ter Schure A, Van Gestel CA, Jurkschat K, Svendsen C, Spurgeon DJ (2015) Short-term soil bioassays may not reveal the full toxicity potential for nanomaterials; bioavailability and toxicity of silver ions (AgNO 3) and silver nanoparticles to earthworm Eisenia fetida in long-term aged soils. Environ Pollut 203:191–198
Zhai Y, Hunting ER, Wouterse M, Peijnenburg W, Vijver MG (2016) Silver nanoparticles, ions and shape governing soil microbial functional diversity: nano shapes micro. Front Microbiol 7:1–9
Marx MC, Wood M, Jarvis SC (2001) A microplate fluorimetric assay for the study of enzyme diversity in soils. Soil Biol Biochem 33:1633–1640
Ernst G, Henseler I, Felten D, Emmerling C (2009) Decomposition and mineralization of energy crop residues governed by earthworms. Soil Biol Biochem 41:1548–1554
Kreuzer K, Adamczyk J, Iijima M, Wagner M, Scheu S, Bonkowski M (2006) Grazing of a common species of soil protozoa (Acanthamoeba castellanii) affects rhizosphere bacterial community composition and root architecture of rice (Oryza sativa L.). Soil Biol Biochem 38:1665–1672
Töwe S, Kleineidam K, Schloter M (2010) Differences in amplification efficiency of standard curves in quantitative real-time PCR assays and consequences for gene quantification in environmental samples. J Microbiol Methods 82:338–341
Bach H-J, Tomanova J, Schloter M, Munch J (2002) Enumeration of total bacteria and bacteria with genes for proteolytic activity in pure cultures and in environmental samples by quantitative PCR mediated amplification. J Microbiol Methods 49:235–245
Rösch C, Mergel A, Bothe H (2002) Biodiversity of denitrifying and dinitrogen-fixing bacteria in an acid forest soil. Appl Environ Microbiol 68:3818–3829
Rotthauwe J-H, Witzel K-P, Liesack W (1997) The ammonia monooxygenase structural gene amoA as a functional marker: molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl Environ Microbiol 63:4704–4712
Barns SM, Takala SL, Kuske CR (1999) Wide distribution and diversity of members of the bacterial kingdom Acidobacterium in the environment. Appl Environ Microbiol 65:1731–1737
Muyzer G, De Waal EC, Uitterlinden AG (1993) Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl Environ Microbiol 59:695–700
Stach JE, Maldonado LA, Ward AC, Goodfellow M, Bull AT (2003) New primers for the class Actinobacteria: application to marine and terrestrial environments. Environ Microbiol 5:828–841
Bacchetti De Gregoris T, Aldred N, Clare AS, Burgess JG (2011) Improvement of phylum- and class-specific primers for real-time PCR quantification of bacterial taxa. J Microbiol Methods 86:351–356
Manz W, Amann R, Ludwig W, Vancanneyt M, Schleifer K-H (1996) Application of a suite of 16S rRNA-specific oligonucleotide probes designed to investigate bacteria of the phylum cytophaga-flavobacter-bacteroides in the natural environment. Microbiol 142:1097–1106
Overmann J, Coolen MJ, Tuschak C (1999) Specific detection of different phylogenetic groups of chemocline bacteria based on PCR and denaturing gradient gel electrophoresis of 16S rRNA gene fragments. Arch Microbiol 172:83–94
Lane D (1991) 16S/23S rRNA sequencing. In: Stackebrandt E, Goodfellow M (eds) Nucleic acid techniques in bacterial systematics. Wiley, England
Kuramae EE, Yergeau E, Wong LC, Pijl AS, van Veen JA, Kowalchuk GA (2012) Soil characteristics more strongly influence soil bacterial communities than land-use type. FEMS Microbiol Ecol 79:12–24
Kallenbach CM, Frey SD, Grandy AS (2016) Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat Commun 7:1–10
Ma W, Jiang S, Assemien F, Qin M, Ma B, Xie Z, Liu Y, Feng H, Du G, Ma X (2016) Response of microbial functional groups involved in soil N cycle to N, P and NP fertilization in Tibetan alpine meadows. Soil Biol Biochem 101:195–206
McGee CF, Storey S, Clipson N, Doyle E (2018) Concentration-dependent responses of soil bacterial, fungal and nitrifying communities to silver nano and micron particles. Environ Sci Pollut Res 25:18693–18704
Emmerling C, Schloter M, Hartmann A, Kandeler E (2002) Functional diversity of soil organisms-a review of recent research activities in Germany. J Plant Nutr Soil Sci 165:408–420
Rincon-Florez VA, Carvalhais LC, Schenk PM (2013) Culture-independent molecular tools for soil and rhizosphere microbiology. Diversity 5:581–612
Zhang X, Liu W, Schloter M, Zhang G, Chen Q, Huang J, Li L, Elser JJ, Han X (2013) Response of the abundance of key soil microbial nitrogen-cycling genes to multi-factorial global changes. PLoS ONE 8:e76500
Fierer N, Bradford MA, Jackson RB (2007) Toward an ecological classification of soil bacteria. Ecology 88:1354–1364
Torsvik V, Øvreås L (2002) Microbial diversity and function in soil: from genes to ecosystems. Curr Opin Microbiol 5:240–245
Allison SD, Martiny JB (2008) Resistance, resilience, and redundancy in microbial communities. PNAS 105:11512–11519
Ventura M, Canchaya C, Tauch A, Chandra G, Fitzgerald GF, Chater KF, van Sinderen D (2007) Genomics of Actinobacteria: tracing the evolutionary history of an ancient phylum. Microbiol Mol Biol Rev 71:495–548
Zhang Y, Cong J, Lu H, Li G, Qu Y, Su X, Zhou J, Li D (2014) Community structure and elevational diversity patterns of soil Acidobacteria. J Environ Sci 26:1717–1724
Li X, Rui J, Xiong J, Li J, He Z, Zhou J, Yannarell AC, Mackie RI (2014) Functional potential of soil microbial communities in the maize rhizosphere. PLoS ONE 9:e112609
Ward NL, Challacombe JF, Janssen PH, Henrissat B, Coutinho PM, Wu M, Xie G, Haft DH, Sait M, Badger J (2009) Three genomes from the phylum Acidobacteria provide insight into the lifestyles of these microorganisms in soils. Appl Environ Microbiol 75:2046–2056
Kielak AM, Barreto CC, Kowalchuk GA, van Veen JA, Kuramae EE (2016) The ecology of Acidobacteria: moving beyond genes and genomes. Front Microbiol 7:1–16
Sharma S, Szele Z, Schilling R, Munch JC, Schloter M (2006) Influence of freeze-thaw stress on the structure and function of microbial communities and denitrifying populations in soil. 72:2148–2154
Feng X, Nielsen LL, Simpson MJ (2007) Responses of soil organic matter and microorganisms to freeze–thaw cycles. Soil Biol Biochem 39:2027–2037
Liu C, Leng W, Vikesland PJ (2018) Controlled evaluation of the impacts of surface coatings on silver nanoparticle dissolution rates. Environ Sci Technol 52:2726–2734
Badawy AME, Luxton TP, Silva RG, Scheckel KG, Suidan MT, Tolaymat TM (2010) Impact of environmental conditions (pH, ionic strength, and electrolyte type) on the surface charge and aggregation of silver nanoparticles suspensions. Environ Sci Technol 44:1260–1266
Long Y-M, Hu L-G, Yan X-T, Zhao X-C, Zhou Q-F, Cai Y, Jiang G-B (2017) Surface ligand controls silver ion release of nanosilver and its antibacterial activity against Escherichia coli. Int J Nanomed 12:1–14
Yang Y, Quensen J, Mathieu J, Wang Q, Wang J, Li M, Tiedje JM, Alvarez PJ (2014) Pyrosequencing reveals higher impact of silver nanoparticles than Ag+ on the microbial community structure of activated sludge. Water Res 48:317–325
McGee C, Storey S, Clipson N, Doyle E (2017) Soil microbial community responses to contamination with silver, aluminium oxide and silicon dioxide nanoparticles. Ecotoxicology 26:449–458
Cho EC, Xie J, Wurm PA, Xia Y (2009) Understanding the role of surface charges in cellular adsorption versus internalization by selectively removing gold nanoparticles on the cell surface with a I2/KI etchant. Nano Lett 9:1080–1084
Villanueva A, Cañete M, Roca AG, Calero M, Veintemillas-Verdaguer S, Serna CJ, del Puerto MM, Miranda R (2009) The influence of surface functionalization on the enhanced internalization of magnetic nanoparticles in cancer cells. Nanotechnology 20:1–9
Wang D, Jaisi DP, Yan J, Jin Y, Zhou D (2015) Transport and retention of polyvinylpyrrolidone-coated silver nanoparticles in natural soils. Vadose Zone J 14:vzj2015.01.0007
Li X, Lenhart JJ, Walker HW (2010) Dissolution-accompanied aggregation kinetics of silver nanoparticles. Langmuir 26:16690–16698
Maillard J-Y, Hartemann P (2013) Silver as an antimicrobial: facts and gaps in knowledge. Crit Rev Microbiol 39:373–383
McShan D, Ray PC, Yu H (2014) Molecular toxicity mechanism of nanosilver. J Food Drug Anal 22:116–127
Schlich K, Klawonn T, Terytze K, Hund-Rinke K (2013) Hazard assessment of a silver nanoparticle in soil applied via sewage sludge. Environ Sci Eur 25:1–14
Yang Y, Wang J, Xiu Z, Alvarez PJ (2013) Impacts of silver nanoparticles on cellular and transcriptional activity of nitrogen-cycling bacteria. Environ Toxicol Chem 32:1488–1494
Choi O, Deng KK, Kim N-J, Ross L, Surampalli RY, Hu Z (2008) The inhibitory effects of silver nanoparticles, silver ions, and silver chloride colloids on microbial growth. Water Res 42:3066–3074
Liang Z, Das A, Hu Z (2010) Bacterial response to a shock load of nanosilver in an activated sludge treatment system. Water Res 44:5432–5438
Giles ME, Morley NJ, Baggs EM, Daniell TJ (2012) Soil nitrate reducing processes-drivers, mechanisms for spatial variation, and significance for nitrous oxide production. Front Microbiol 3:407–423
Ji B, Yang K, Zhu L, Jiang Y, Wang H, Zhou J, Zhang H (2015) Aerobic denitrification: A review of important advances of the last 30 years. Biotechnol Bioprocess Eng 20:643–651
Tiedje JM (1988) Ecology of denitrification and dissimilatory nitrate reduction to ammonium. Biol Anaerob Microor 717:179–244
Asadishad B, Chahal S, Akbari A, Cianciarelli V, Azodi M, Ghoshal S, Tufenkji N (2018) Amendment of agricultural soil with metal nanoparticles: effects on soil enzyme activity and microbial community composition. Environ Sci Technol 52:1908–1918
Panáček A, Kvítek L, Smékalová M, Večeřová R, Kolář M, Röderová M, Dyčka F, Šebela M, Prucek R, Tomanec O (2018) Bacterial resistance to silver nanoparticles and how to overcome it. Nat Nanotechnol 13:65
Samarajeewa AD, Velicogna JR, Princz JI, Subasinghe RM, Scroggins RP, Beaudette LA (2017) Effect of silver nano-particles on soil microbial growth, activity and community diversity in a sandy loam soil. Environ Pollut 220:504–513
He S, Feng Y, Ni J, Sun Y, Xue L, Feng Y, Yu Y, Lin X, Yang L (2016) Different responses of soil microbial metabolic activity to silver and iron oxide nanoparticles. Chemosphere 147:195–202
Metreveli G, Frombold B, Seitz F, Grün A, Philippe A, Rosenfeldt RR, Bundschuh M, Schulz R, Manz W, Schaumann GE (2016) Impact of chemical composition of ecotoxicological test media on the stability and aggregation status of silver nanoparticles. Environ Sci Nano 3:418–433
Jacobson AR, McBride MB, Baveye P, Steenhuis TS (2005) Environmental factors determining the trace-level sorption of silver and thallium to soils. Sci Total Environ 345:191–205
Levard C, Hotze EM, Lowry GV, Brown GE Jr (2012) Environmental transformations of silver nanoparticles: impact on stability and toxicity. Environ Sci Technol 46:6900–6914
Tripathi DK, Tripathi A, Singh S, Singh Y, Vishwakarma K, Yadav G, Sharma S, Singh VK, Mishra RK, Upadhyay R (2017) Uptake, accumulation and toxicity of silver nanoparticle in autotrophic plants, and heterotrophic microbes: a concentric review. Front Microbiol 8:1–16
Jung WK, Koo HC, Kim KW, Shin S, Kim SH, Park YH (2008) Antibacterial activity and mechanism of action of the silver ion in Staphylococcus aureus and Escherichia coli. Appl Environ Microbiol 74:2171–2178
Nies DH (1999) Microbial heavy-metal resistance. Appl Microbiol Biotechnol 51:730–750
Keesstra SD, Bouma J, Wallinga J, Tittonell P, Smith P, Cerdà A, Montanarella L, Quinton JN, Pachepsky Y, van der Putten WH (2016) The significance of soils and soil science towards realization of the United nations sustainable development goals. Soil 2:111–128
Wu B, Wang Y, Lee Y-H, Horst A, Wang Z, Chen D-R, Sureshkumar R, Tang YJ (2010) Comparative eco-toxicities of nano-ZnO particles under aquatic and aerosol exposure modes. Environ Sci Technol 44:1484–1489
Sudheer Khan S, Bharath Kumar E, Mukherjee A, Chandrasekaran N (2011) Bacterial tolerance to silver nanoparticles (SNPs): aeromonas punctata isolated from sewage environment. J Basic Microbiol 51:183–190
Juan W, Kunhui S, Zhang L, Youbin S (2017) Effects of silver nanoparticles on soil microbial communities and bacterial nitrification in suburban vegetable soils. Pedosphere 27:482–490
Ma Y, Metch JW, Vejerano EP, Miller IJ, Leon EC, Marr LC, Vikesland PJ, Pruden A (2015) Microbial community response of nitrifying sequencing batch reactors to silver, zero-valent iron, titanium dioxide and cerium dioxide nanomaterials. Water Res 68:87–97
Cornelis G, Kirby JK, Beak D, Chittleborough D, McLaughlin MJ (2010) A method for determination of retention of silver and cerium oxide manufactured nanoparticles in soils. Environ Chem 7:298–308
Shoults-Wilson WA, Reinsch BC, Tsyusko OV, Bertsch PM, Lowry GV, Unrine JM (2011) Role of particle size and soil type in toxicity of silver nanoparticles to earthworms. Soil Sci Soc Am J 75:365–377
Postgate JR (1967) Viability measurements and the survival of microbes under minimum stress. Adv Microb Physiol 1:1–23
Cornelis G, DooletteMadeleine Thomas C, McLaughlin MJ, Kirby JK, Beak DG, Chittleborough D (2012) Retention and dissolution of engineered silver nanoparticles in natural soils. Soil Sci Soc Am J 76:891–902
Sagee O, Dror I, Berkowitz B (2012) Transport of silver nanoparticles (AgNPs) in soil. Chemosphere 88:670–675
Liu J, Hurt RH (2010) Ion release kinetics and particle persistence in aqueous nano-silver colloids. Environ Sci Technol 44:2169–2175
Marambio-Jones C, Hoek EM (2010) A review of the antibacterial effects of silver nanomaterials and potential implications for human health and the environment. J Nanopart Res 12:1531–1551
Verma A, Stellacci F (2010) Effect of surface properties on nanoparticle–cell interactions. Small 6:12–21
Park E-J, Yi J, Kim Y, Choi K, Park K (2010) Silver nanoparticles induce cytotoxicity by a Trojan-horse type mechanism. Toxicol Vitro 24:872–878
Uwizeyimana H, Wang M, Chen W, Khan K (2017) The eco-toxic effects of pesticide and heavy metal mixtures towards earthworms in soil. 55:20–29
Stein LY, Arp DJ (1998) Loss of ammonia monooxygenase activity in Nitrosomonas europaea upon exposure to nitrite. Appl Environ Microbiol 64:4098–4102
Bollag J-M, Henninger NM (1978) Effects of nitrite toxicity on soil bacteria under aerobic and anaerobic conditions. Soil Biol Biochem 10:377–381
Das P, Barua S, Sarkar S, Chatterjee SK, Mukherjee S, Goswami L, Das S, Bhattacharya S, Karak N, Bhattacharya SS (2018) Mechanism of toxicity and transformation of silver nanoparticles: inclusive assessment in earthworm-microbe-soil-plant system. Geoderma 314:73–84
Cornelis G, Pang L, Doolette C, Kirby JK, McLaughlin MJ (2013) Transport of silver nanoparticles in saturated columns of natural soils. Sci Total Environ 463:120–130
Gunsolus IL, Mousavi MP, Hussein K, Bühlmann P, Haynes CL (2015) Effects of humic and fulvic acids on silver nanoparticle stability, dissolution, and toxicity. Environ Sci Technol 49:8078–8086
Authors’ contributions
JC designed and manufactured the Ag10-NH2 and Ag10-COOH nanoparticles. SS performed and evaluated the transmission electron microscopy of AgNP. KY, MF, and DR performed the ζ-potential and size measurements of AgNP as well as the asymmetrical flow field-flow fractionation (AF4) of AgNP. GAL and EC designed and performed the soil experiments. GAL, EC and MW analysed the data and wrote the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We appreciate Elvira Sieberger (University of Trier) for her excellent laboratory support and assistance.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
All datasets on which the conclusions of the paper rely are presented in the main manuscript (Additional files 1 and 2).
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
The study was financially supported by the BMBF (Grant No. 03X0150) and the German Research Foundation (DFG Research unit FOR 1536, MA 3273/3-2).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Additional files
Additional file 1.
Characterization of AgNP.
Additional file 2.
Raw data.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Grün, AL., Manz, W., Kohl, Y.L. et al. Impact of silver nanoparticles (AgNP) on soil microbial community depending on functionalization, concentration, exposure time, and soil texture. Environ Sci Eur 31, 15 (2019). https://doi.org/10.1186/s12302-019-0196-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12302-019-0196-y