
Abstract
• Key message
Canopy closure and stand age significantly affected the litter decomposition. Therefore, stand-specific decomposition constants (k) should be calculated in forest carbon models for more accurate carbon budget estimation. Furthermore, to reduce the carbon release from decomposing litter, regeneration cutting should be carried out at later ages, and heavy thinning should not be implemented in mature as well as overmature stands.
• Context
Decomposition of litter has an important role in primary production with its influence on nutrient release for plant uptake and carbon flux in forest ecosystems. Thus, understanding the effects of the intervention on litter decomposition is crucial for carbon management in forestry.
• Aims
The effects of stand structure and exposure on litter decomposition rate in Pinus sylvestris stands were investigated.
• Methods
Samples were taken from young to overmature stands with open to dense canopy. The litterbag method was used to measure the mass loss of the litter. The k values were calculated from the mass loss of decaying litter. Carbon and nitrogen contents of the litter were determined.
• Results
Cutting caused the decomposition to accelerate at a rate of up to 58% depending on its intensity. The k values were found to fluctuate over time from 0.189 in moderately dense-canopied stands to 0.317 in open-canopied overmature stands. Stand basal area, incubation time, and remaining carbon concentration of the litter accounted for 75% of the variation in the k value.
• Conclusion
Changes in the stand structure affect the litter decomposition rate in forest ecosystems. Heavy thinning can alter the litter decomposition process drastically, while moderate thinning may not have a clear effect in the long run.
Similar content being viewed by others


1 Introduction
Plant litter decomposition through microbes and fungi is the only process that enables recycling of chemical elements in forest ecosystems, and is thus crucial for sustaining life on earth (Berg and Laskowski 2005). The decomposition of litter is an important process which is critical to understand the nutrient dynamics as well as the development of nutrient deficiency in a forest ecosystem and responses of the ecosystems to environmental disturbances (Prescott 2010). The most important product of litter decomposition influences soil productivity and microbial biodiversity (Berg and McClaugherty 2014). In the sense of climate change, decomposition of litter has an important role in primary production with its influence on nutrient release for plant uptake and carbon flux (Rodríguez et al. 2009). Decaying organic matter is an important source of carbon dioxide returned to the atmosphere which is the main greenhouse gas with a role in potential global climate change (Berg and McClaugherty 2014).
Silvicultural treatments, which change the structure of a stand by reducing the canopy cover and basal area, have been applied in forest stands to maintain the growth or to establish a new generation by forest management. Seed cut, a silvicultural harvesting method, which takes all trees but the seed trees, has been widely used for natural regeneration of trees (Youngblood 1991; Schönenberger and Brang 2004), although much of the forest is regenerated by clear-cutting (Dougherty and Duryea 1991). Scots pine stands are commonly regenerated using the seed cut method in Turkey, changing the canopy cover from open to moderately dense. As another silvicultural method, thinning has been a common practice applied in the forest to maintain the growth rate of stands for decades in forestry (Pourmajidian et al. 2010; Tian et al. 2010). All kinds of cutting alter the balance of the ecosystem. Although the effects of thinning on the stand growth, litter production, soil properties, and carbon sequestration are well documented, very few studies have been conducted on the effects of thinning on litter decomposition rates in forests (Cortina and Vallejo 1994; Kunhamu et al. 2009; del Rio et al. 2017). However, contrasting results have been presented by the studies. For example, Blanco et al. (2011) carried out a study in which two degrees of thinning with a control were applied in young Scots pine stands at two different sites, namely the Mediterranean and the continental sites, suggesting that the decomposition rate decreased with increasing thinning intensity. In contrast, Kunhamu et al. (2009) suggested that high thinning intensities accelerated the decay rate of litter in an Acacia mangium Willd. stand. However, Will et al. (1983) reported a non-significant effect of thinning on the decomposition rate of Pinus radiata D. Don litter.
Exposure is a critical land property for forest ecosystems that affects various ecological processes, such as growth and litter decomposition. However, contrasting results have been published regarding the effect of aspect on ecological processes (Sariyildiz 2014, 2015; Singh 2018; Måren et al. 2015). Thus, more local-level knowledge is needed for better understanding of the effect of aspect on various ecological processes.
The effects of climatic factors such as precipitation, temperature, and evapotranspiration, as well as litter chemistry on litter decomposition have been investigated by many researchers (Johansson 1994; Sariyildiz 2003; Prescott 2010; Kim 2012). These studies were mainly focused on the decomposition rate of the foliar litterfall because of the assumption that most of the litterfall consisted of needles or leaves. On the other hand, litterfall components, including needles, cones, and branches, differ in chemical composition and thus have different decomposition patterns (Hristovski et al. 2001; Berg 2014). Therefore, there is a gap of knowledge on the decomposition of total litterfall.
The objective of this study was to evaluate the effects of stand structure and facing slope on litter decomposition as well as carbon and nitrogen release via litter decomposition. We hypothesized that mass loss of decaying litter and nutrient release from litter decomposition would be higher (i) in sparse-canopied stands than in the dense-canopied stands, (ii) in young stands than in older stands, and (iii) on the southern slope than on the northern slope.
2 Methods
2.1 Study site
The study was conducted at pure Scots pine stands spreading over the Sundiken Mountain range in the inner Anatolia region of Turkey (39º57ʹN, 31°09ʹE) on a well-drained luvisol (IUSS Working Group WRB 2015) developed from mica schist, with a sandy loam texture and pH of 5.6–6.0. Sampling was performed in naturally regenerated stands. The study area has a continental climate and low rainfall, with mean temperature and rainfall of 9.5 °C and 486 mm, respectively. A detailed description of the study area was given by Çömez et al. (2019).
2.2 Sampling procedure
To cover a wide range of the forest life cycle and to understand the stand structure effect on litter decomposition, sample plots were chosen from three developmental stages, i.e. young, mature, and overmature stages; and three classes of canopy cover, i.e., open forest (10–40%), moderately dense canopy (40–70%), and very dense canopy (> 70%), except young stands because of the lack of open and moderately dense canopied stands, with four replications, 28 in total (Appendix, Table 5). To understand the effect of slope aspect on the decomposition rate, two of the replications were established on the northern slope, and the others on the southern slope. Mature and overmature stands studied were exposed silvicultural treatments 5 to 8 years before the measurements by the local authority. Litterfall samples were collected by 0.5 × 0.5 m traps for 2 years to attain enough amounts of litterfall samples to be placed in litter bags. After having collected the samples in each sample plot, litter samples with an air-dried weight of approximately 10 g including needles, bark, branches, and cones, taking into account their share in the total litterfall, were filled into the litter bags to determine the overall litter decomposition rate. The shares of litterfall components are given in Table 5. To determine the litter decomposition rate, 60 litter bags, made from nylon nets with 1 × 1 mm mesh and 20 × 15 cm in size, were filled with litter and placed on the ground of each sample plot. Four litter bag samples were collected from each plot every 3 to 5 months for 4 years. The mean mass loss rate was calculated from the four litter bags of each plot. To determine the structural properties of the stands, the diameter at breast height (dbh) and height of all trees were measured with a caliper and clinometer, respectively. Tree age was determined by counting tree rings on increment core samples extracted from the base of the trees using an increment borer. As a common parameter for forest stands, basal area was calculated as the cross-sectional area at breast height of the trees in the sample plots.
2.3 Laboratory methods
Litter bag samples were oven-dried at 70 ºC to constant weight before the weighing. Mass loss was calculated by Eq. (1):
where MR is remaining mass (%), M0 is the initial mass of litter (g), and Mt is the litter mass t time later (g).
Accumulated carbon (C) and nitrogen (N) release were calculated by Eq. (2):
where C, Nr is accumulated carbon or nitrogen release (%), MR is the remaining litter mass (%), and C, NConc. is the carbon or nitrogen concentration of the decomposing litter (%), respectively.
The decomposition constant was calculated according to Olson (1963), Eq. (3):
where k is the decomposition constant, M0 is the initial mass of litter (g), and Mt is the litter mass t time later (g).
The samples were analyzed for carbon and nitrogen by the Dumas method using a LECO CNH analyzer (Leco Corporation, St. Joseph, MI, USA). Nitrogen and carbon concentrations were standardized to initial concentrations by multiplying their concentrations by remaining mass percent.
2.4 Statistical analysis
Data were controlled by the Shapiro–Wilk test for normal distribution, and by Levene’s test for the homogeneity of variance. When data were normally distributed, variations in the mass loss over time and stand type were evaluated by repeated-measures analysis of variance (ANOVA), taking the maturity, canopy, and facing slopes as between-subject factors. All data showed normal distribution and homogeneous variance. One-way ANOVA was used to compare the remaining litter mass, k value, and carbon and nitrogen content of the litter of the stands at the end of the incubation period. The Scheffe test was used to determine the homogenous groups (Ozdamar 2002).
Decomposition constant was estimated by multiple regression analysis, using the basal area of the stands m2 ha−1), stand age (year), the length of the incubation period of the litter (days), and accumulated C and N concentration (%) as independent variables. Equations with the highest R2, the lowest standard error, and constants, with a significance level at P < 0.05, are given in this study. Statistical analyses were conducted using the package of IBM SPSS Statistics for Windows, Version 21.0 (IBM Corp., Armonk, NY, USA).
3 Results
3.1 Litter mass loss
The remaining mass of the litter and k constant varied significantly with time and stand canopy, as well as facing slope. Interactions between the time and stand canopy did not affect the k constant. Canopy closure and facing slope also significantly affected the decomposition rate. Both the remaining mass and the k value were influenced by the interaction among the maturity, stand canopy, and facing slope (Table 1).
At the end of the incubation time of 1367 days, 30% of the initial litter mass remained undecomposed in young stands, while almost half remained in moderately dense and dense stands. Although the remaining mass was higher in the moderately dense forest in mature stands, it was low in dense overmature stands. Open-canopied stands had a lower remaining mass in both mature and overmature ones. As the canopy became more open, the remaining mass of the litter decreased, except for moderately dense mature stands. An ascending trend was observed in stands from young to overmature stage in remaining litter mass, from 30% in young stands to 49% in overmature stands. The decomposition constant was found to be highest in young stands, while the lowest was found in moderately dense mature stands, with an increasing tendency from the dense canopy to open and young forest, with the exception of moderately dense mature stands (Fig. 1).

Mean remaining mass (a) and k constant (b) (± standard error of the mean) at the end of the study period. Comparison among stands was analyzed by one-way ANOVA with the Scheffe test. Different letters indicate significant differences (P < 0.05) among the stands
The remaining mass of litter was found to be very similar between northern and southern slopes in young stands, while that of northern slopes was lower than that of southern slopes in both mature and overmature stands. Litter was decomposed more in open-canopied stands than in both moderately dense- and dense-canopied ones, except in mature stands on the northern slope. The decomposition process occurred more rapidly on the northern slopes (Fig. 2).

Temporal changes in remaining mass of the litter with facing slopes, stand development, and canopy cover
The decomposition constant showed an increase towards the end of the first year in all stands, with the highest in young and the lowest in overmature dense-canopied stands. After the third year of the incubation, k constants of the moderately dense and very dense stands remained steady, while those of young and overmature open forests continued to increase slightly. Differences in the k constant between open- and moderately dense-canopied forests became pronounced after 687 days of incubation (Fig. 3).

The course of k values according to sampling periods in young, mature, and overmature stands with different canopy cover
Stand basal area alone accounted for 47% of the variation in the k constant calculated for 1367 days, but for 75% of that along with incubation days and remaining C. Basal area, incubation days, and remaining C in the litter were negatively related to the k constant (Table 2).
3.2 Carbon and nitrogen mineralization
Carbon concentrations of the litter fluctuated from 50 to 55% throughout the study period, with a significant difference (P < 0.01) according to repeated-measures ANOVA. Initial carbon concentrations among the stands were not significantly different, ranging from 51 to 54%. Cumulative carbon concentrations showed a steady decrease from approximately 53 to 21%. At the end of 1367 days of incubation, 24% of the initial carbon remained undecomposed in the dense stands, while 15% remained in young stands (Fig. 4 a–b).

Changes in (a) C concentration (%), (b) accumulated C concentration (%), (c) N concentration (%), (d) accumulated N concentration (%) of decomposing litter over time in young stands, open-canopied, moderately dense, and dense stands
The initial nitrogen concentration of the litter ranged from 0.92% in very dense-canopied stands to 1.24% in young stands, with a significant difference (P < 0.01) between young stands and the other stands. However, there was no significant difference at the end of 155 days, with a decline of 0.42 to 0.49% of N concentration. An increasing trend in N concentrations of the litter was observed after 155 days of incubation, with the effect of treatment and time. Accumulated N decreased considerably from 1.24–0.92% to 0.37–0.44% over time and stabilized around 0.45% between incubation days 524 and 1137. Subsequently, a slight decrease trend was observed after 1137 days (Fig. 4 c–d).
Another increasing trend was observed from very dense stands to open-canopied and young stands for the initial N concentrations in the litter. However, there were no significant differences in initial carbon concentrations among the stands, although litter from very dense stands had the highest carbon concentration, with a mean of 54%, and the moderately dense stands had the lowest with 51% (Table 3).
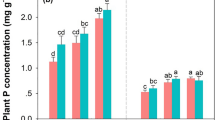
Remaining litter mass was correlated with the initial N concentration at a significance level of P < 0.01. The remaining mass decreased with increasing N concentration of the litterfall (Fig. 5).

The relationship between initial N concentration and the remaining mass of the litter at the end of the study period (n = 28, P < 0.05)
4 Discussion
4.1 Litter decomposition
The decomposition of forest litter is a process mainly driven by environmental conditions and the quality of the substrate (Sariyildiz and Kucuk 2008; Berg and McClaugherty 2014; Karishna and Mohan 2017). Silvicultural treatments, such as thinning and seed cutting, can alter environmental conditions as well as production (Prescott et al. 2004; Tian et al. 2010) and composition of litterfall. Besides, the quantity and quality of the forest floor can be changed by residues left in the stand after the cutting treatments. In our study, lower remaining litter mass in young and open-canopied forests compared to dense ones indicated that stand structure influenced the decomposition rate of the forest floor by changing the environmental conditions, namely temperature and moisture in the stands. Besides, thinning operations can affect litterfall composition, as suggested by Blanco et al. (2006). Specifically, needle litter was more dominant in young stands, leading to a high decomposition rate due to easily decomposable chemical composition. Cutting in forests results in a decrease in canopy closure and the basal area left, leading to more rainfall and light reaching the soil surface (Prescott et al. 2004; Tian et al. 2010). Thus, the activity of decomposers accelerates, and consequently the decomposition rate of the litter increases. Despite the low needle litterfall, an easily decomposable fraction due to low lignin and high nitrogen content, but a higher cones fraction, decomposition rates were higher in the open-canopied stands, likely due to the prevailing effect of microclimatic condition on soil biota. Unlike the needles, cones have a woody structure, indicating that they include more lignin than needles do (Taylor et al. 1991; Font et al. 2009). High lignin content causes a slow breakdown in the forest floor (Sariyildiz and Kucuk 2008; Wang et al. 2018). Our results clearly showed that cuttings accelerated the decomposition of litter in a mature stand, unlike the results reported by some researchers (Pausas 1997; Blanco et al. 2011). On the contrary, Chen et al. (2014) reported that thinning increased the decomposition rate in Pinus tabulaeformis Carrière plantations. In our study, cutting possibly resulted in an increase in moisture and temperature in the forest floor and topsoil, which created favorable conditions for the decomposers. Although the canopy of the young stands covered most of the ground, surprisingly, litter decomposition occurred faster than in the mature and overmature stands. Although the fast litter decomposition in the young stands may be attributable to the high share of needles in the total litter, the effect of microenvironmental conditions remains unclear in the present study. Therefore, further research is needed to understand the change in environmental conditions under young and mature stands.
The remaining mass of litter in young stands, which had a needle fraction of 87% of the total litterfall, was similar to some studies regarding needle litter decomposition (Berg and Staaf 1980; Laiho et al. 2004), likely due to similar climatic conditions and the high needles fraction in our study. On the other hand, Sariyildiz and Kucuk (2008) reported a higher decomposition rate compared to our results, likely because their results were for needle litterfall. Pausas (1997) reported that the k values varied between 0.221 and 0.283 for Scots pine stands, which were close to our results for moderately dense overmature and open-canopied mature forests (0.235 and 0.256, respectively) but lower for young (0.349) and higher for dense ones (0.189–0.201). An additional explanation may be the suitable climate of their study area with more moist conditions. On the other hand, Sariyildiz (2008) observed a decomposition rate closer to our results for young stands, with k values from 0.3 to 0.4. This may be attributed to the fact that most of the litter consisted of needles in young stands in our study, which was similar to the study of Sariyildiz (2008). Janušauskaité et al. (2013) reported a lower decomposition rate for northern sites, where the temperature is comparably low. This may be related to the retarding effect of low temperature on decomposition (Berg 2014; Krishna and Mohan 2017). Enez et al. (2015) found a lower decomposition rate of needle litter on scalped mineral soil than on non-harvesting activity areas under a Scots pine stand due to the lower microbial activity. Thus, litter decomposition dynamics of young stands might differ from those of mature ones due to the differences in composition of the litterfall and the changes in environmental conditions of the stands.
Many researchers have reported that the facing slope affects the decomposition rate, being faster on the northern slope than on the southern slope (Sariyildiz and Kucuk 2008; Jasińska et al. 2019). On the other hand, some researchers have reported a faster decomposition of litterfall on the southern aspects than on the northern (Mudrick et al. 1994; Qualls 2016), which is in line with our results. Furthermore, in open-canopied mature stands, the decomposition rate was found to be faster on the southern slopes in comparison to the north in contrast to the case of the dense canopy, likely due to age- and light-related changes in litterfall composition or balance between temperature and moisture on the forest floor. Details of the remaining litter mass by stand and facing slope are provided in Table 4.
We did not determine the decomposition rates of the litter fractions separately in our study, since all litterfall fractions make up the forest floor and decompose together. Considering only a certain fraction of litterfall such as needles or leaves could result in an overestimation of the litter decomposition rate.
4.2 Temporal changes in k constant and estimation models
Decomposition constant trends showed different patterns due to changes in the chemical composition of litter and microbial diversity during the decomposition process (Yue et al. 2018), indicating that the k constant would be stabilized after approximately 1000 days in dense and moderately dense forest, but not in open or young stands. In addition, the k constant might continue to increase slightly until 1400 days in open-canopied and young stands. Therefore, k values might be underestimated in both young and heavily thinned stands in cases of incubation time shorter than 1400 days. Our results partly confirmed those of Berg et al. (2010), who proposed that 1100 to 2000 days of the period was sufficient for limit values of litter decomposition. Still, there is a need for further studies on the decomposition rate in young and mature stands which are heavily treated.
Basal area is an important parameter that can be used for evaluating the stand structure. It is decreased by silvicultural interventions as well as natural or human-induced disturbances. Pausas (1997) reported that the decomposition rate of the litter in an open-canopied Scots pine stand was lower than that of the closed-canopy stand, which was studied in a Mediterranean site. The slower decomposition rate was attributed to low moisture, high solar radiation, steep slope, and high altitude, as well as open canopy cover by Pausas (1997). Similarly, Blanco et al. (2011) found that the decomposition rate decreased after thinning. A decline in the basal area, meaning also a decrease in canopy cover, leads to more sunlight and rainfall reaching the forest floor, favoring the environmental conditions for microbial activity. Therefore, the basal area gave a robust fit with the k constant.
4.3 Nitrogen and carbon release
Our results showed that the initial N concentration of litter was related to the decomposition rate. Higher N concentrations in young stands also supported this relationship. An increase in the N concentration causes a decrease in the C:N ratio, which is an index for litter decomposition. Sun et al. (2016) reported a decreasing trend in N concentration of senesced pine needles from young to older stands. The decreasing trend in N with stand age might be related to changes in soil nutrient availability and forest growth rates during stand development, as pointed out by Sun et al. (2016). Besides, the concentrations tended to increase slightly as the canopy structure of the stands became more open, indicating that cuttings might result in changes in nutrient withdrawal before needle shed in the pines, in contrast to Blanco et al. (2009). But our results confirmed the findings of Berg et al. (1995), who reported that N concentration in the needle litter of Scots pine was mainly related to climatic conditions. On the other hand, the initial N concentration of the litter in the dense forests of our study was similar to the results of Blanco et al. (2006) for needle litter of Scots pine stands in sites with a cold wet Mediterranean climate. However, they reported lower values for sites with a cold wet continental climate than our results. They also suggested that thinning affected N concentration of needle litter.
Initial N concentrations found in the present study were similar to those reported by Blanco et al. (2006), who found the N concentration of litterfall in the range of 0.79–1.10% depending on the climatic conditions and litterfall fractions for Scots pine. The nitrogen content of the needles may be influenced by many factors, such as agricultural and poultry operations, fertilization, and atmospheric N deposition (Ghaly and Ramakrishnan 2015). Atmospheric deposition can provide a significant amount of nutrients in forest ecosystems (Lovblad 2000). However, there were no dense settlements or agricultural, poultry, or livestock activities, which could be a source of atmospheric deposition, near the study area. Even if N deposition occurred in the study area, it was expected that all the plots would be affected similarly because the sample plots chosen were in the same district. Although an increasing trend was observed in the initial N concentration of the litter as the canopy cover decreased, this was not found significant. In contrast, Bueis et al. (2018) reported that tree removal might diminish nutrient uptake by trees because of the competition among the nutrients in the soil. Similarly, Blanco et al. (2006) showed that thinning decreased the N concentration of the litterfall in a Scots pine stand in the Mediterranean region. But they reported that the N concentrations were not affected by thinning in the continental climate. On the other hand, Chen et al. (2014) reported that thinning could result in a decrease or increase in the nutrient concentration of the litter. It seems that the effect of thinning on initial nutrient concentrations of the litterfall may change depending on environmental conditions, and further research is needed to clarify the thinning effects on nutrient concentration of the litterfall. In the present study, higher initial N concentration of litter resulted in a higher decomposition rate as revealed by many researchers (Gao et al. 2019; Sariyildiz 2003; Zhang et al. 2008) in contrast to Berg (2000). The enhancing effects of N on decomposition rate may be related to rich microbial diversity supported by the high nutritious value of the litter (Gao et al. 2015), which has a high proportion of needles that contain higher nutrients compared to other litter fractions in young stands. On the other hand, despite the lower nutrient content likely due to the inclusion of a higher portion of fine woody litter, the decomposition rate in open-canopied stands also was higher than in the moderately thinned and un-thinned stands, likely because of the prominent effects of temperature and humidity on the decomposition process.
Lower accumulated carbon concentration of decomposing litter in young stands than in mature stands indicated higher carbon emission arising from young stands until the formation of humus, which is an important stable carbon pool. Thus, our results support those of Jandl et al. (2007), who suggested the aging of the forest as a management tool to increase the soil carbon stock. On the other hand, similar to the results of Blanco et al. (2011), who reported that an increase in thinning intensity caused slowing of decomposition, more carbon was released from open-canopied overmature and mature stands in the present study. However, leaching of carbon as dissolved organic carbon may be possible due to higher throughfall as a result of the reduced canopy cover. However, this remained unclear in the present study. On the other hand, Kunhamu et al. (2009) reported a higher decomposition rate, thus high carbon emission, in thinned Acacia mangium forest compared to un-thinned one, which was in line with the results of the present study. It seems that the effect of thinning on decomposition process depends upon the climatic conditions to some extent.
5 Conclusions
In this study, it was concluded that cutting-induced changes in stand structure affected the litter decomposition process in forest ecosystems due to the micro-environmental change as well as the changes in litterfall composition and chemistry. Although moderate thinning was not able to change the litter decomposition process drastically, heavy treatments could do so in the long run. Thus, regeneration cuttings should be postponed to later ages and heavy thinning avoided in mature and overmature stands to reduce forestry-related carbon release. Heavy treatments should be avoided by forest management to keep the litter decomposition slower where the carbon-focused forestry will be implemented. Besides, the elongation of the rotation period of the forest should be used as a tool for carbon management. Relatively higher precipitation reaching the forest floor together with higher temperature due to the reduced canopy might cause rapid N mineralization, thus leading to a nitrogen-rich needle fall. However, there is a need for further researches on the effect of thinning on the nutrient re-translocation process. On the other hand, it is important to use the decomposition constant which relies on long-term incubation, with at least 1050 days. Besides, due to the significant effect of canopy closure on the decomposition rate, stand-specific or recalculated k constants according to stand basal area should be recommended for use in forest carbon models for more accurate carbon budget estimation.
Data availability
The data that support the findings of this study are available from the Turkish General Directorate of Forestry, but restrictions apply to the availability of these data, which were used under license for the current study, and so are not publicly available. Data are however available from the authors upon reasonable request and with permission of the Turkish General Directorate of Forestry.
References
Berg B (2000) Litter decomposition and organic matter turnover in northern forest soils. For Ecol Manag 133:13–22. https://doi.org/10.1016/S0378-1127(99)00294-7
Berg B (2014) Foliar Litter Decomposition: A Conceptual Model with Focus on Pine (Pinus) Litter—A Genus with Global Distribution. ISRN Forestry Volume 2014 Article ID 838169, 22 pages. https://doi.org/10.1155/2014/838169
Berg B, Staaf H (1980) Decomposition rate and chemical changes of scots pine needle litter. I-Influence of stand age Ecoll Bull 32:363–372
Berg B, Calvo de Anta R, Escudero A, Gärdenäs A, Johansson MB, Laskowski R, Madeira M, Mälkönen E, McClaugherty C, Meentemeyer M, Virzo De Santo A (1995) The chemical composition of newly shed needle litter of Scots pine and some other pine species in a climatic transect. X Long-term decomposition in a Scots pine forest. Can J Bot 73:1423–1435
Berg B, Laskowski R (2005) Litter Decomposition: A Guide to carbon and Nutrient Turnover. Academic Press, USA
Berg B, Davey MP, De Marco A, Emmett B, Faituri M, Hobbie SE, Johansson MB, Liu C, McClaugherty C, Norell L, Rutigliano FA, Vesterdal L, Virzo De Santo A (2010) Factors influencing limit values for pine needle litter decomposition: a synthesis for boreal and temperate pine forest systems. Biogeochemistry 100:57–73. https://doi.org/10.1007/s10533-009-9404-y
Berg B, McClaugherty C (2014) Plant Litter-Decomposition, Humus Formation. Springer-Verlag, Berlin, Carbon Sequestration
Blanco JA, Imbert JB, Castillo FJ (2006) Nutrient return via litterfall in two contrasting Pinus sylvestris forests in the Pyrenees under different thinning intensities. For Ecol Manage 256:1840–1852. https://doi.org/10.1016/j.foreco.2008.07.011
Blanco JA, Imbert JB, Castillo FJ (2009) Thinning affects nutrient resorption and nutrient-use efficiency in two Pinus sylvestris stands in the Pyrenees. Ecol Appl 19(3):682–698. https://doi.org/10.1890/1051-0761-19.3.682
Blanco JA, Imbert JB, Castillo FJ (2011) Thinning affects Pinus sylvestris needle decomposition rates and chemistry differently depending on site conditions. Biogeochemistry 106:397–414. https://doi.org/10.1007/s10533-010-9518-2
Bueis T, Bravo F, Pando V, Turrión MB (2018) Local basal area affects needle litterfall, nutrient concentration, and nutrient release during decomposition in Pinus halepensis Mill. plantations in Spain. Ann For Sci 75(1)
Chen X, Page-Dumroese D, Lv R, Wang W, Li G, Liu Y (2014) Interaction of initial litter quality and thinning intensity on litter decomposition rate, nitrogen accumulation and release in a pine plantation. Silva Fennica 48(4) article id 1211. 13 p
Cortina J, Vallejo VR (1994) Effects of clearfelling on forest floor accumulation and litter decomposition in a radiate pine plantation. For Ecol Manag 70:299–310. https://doi.org/10.1016/0378-1127(94)90095-7
Çömez A, Tolunay D, Güner ŞT (2019) Litterfall and the effects of thinning and seed cutting on carbon input into the soil in Scots pine stands in Turkey. Eur J Forest Res 138:1–14. https://doi.org/10.1007/s10342-018-1148-6
del Río M, Bravo-Oviedo A, Pretzsch H, Löf M, Ruiz-Peinado R (2017) A review of thinning effects on Scots pine stands: From growth and yield to new challenges under global change. Forest Syst 26 (2), eR03S, 19 pages. https://doi.org/10.5424/fs/2017262-11325
Dougherty PM, Duryea ML (1991) Regeneration: An Overview of Past Trends and Basic Steps Needed to Ensure Future Success. In: Duryea M, Daugherty PM (eds) Forest Regeneration Manual. Springer Science+Business Media, B.V., USA
Enez K, Aricak B, Sariyildiz T (2015) Effects of harvesting activities on litter decomposition rates of Scots pine, Trojan fir, and sweet chestnut. Šumarski List 139:361–368
Font R, Conesa JA, Moltó J, Muñoz M (2009) Kinetics of pyrolysis and combustion of pine needles and cones. J Anal Appl Pyrolysis 85:276–286. https://doi.org/10.1016/j.jaap.2008.11.015
Gao J, Kang F, Li T, Song X, Zhao W, Yu X, Han H (2015) Assessing the Effect of Leaf Litter Diversity on the Decomposition and Associated Diversity of Fungal Assemblages. Forests 6:2371–2386. https://doi.org/10.3390/f6072371
Gao J, Han H, Kang F (2019) Factors controlling decomposition rates of needle litter across a chronosequence of Chinese pine (Pinus tabuliformis Carr.) forests. Pol J Environ Stud 28(1):91–102. https://doi.org/10.15244/pjoes/84770
Ghaly AE, Ramakrishnan VV (2015) Nitrogen sources and cycling in the ecosystem and its role in air, water and soil pollution: A critical review. J Pollut Eff Cont 03(02)
Hristovski S, Melovski LJ, Grupche LJ, Šušlevska M (2001) Litter decomposition rate of leaves, branches and beech acorns in the beech ecosystem calamintho grandiflorae-fagetum in ‘Mavrovo’ national park. In: Radoglou K (ed) Proceedings of the International Conference: Forest research: a challenge for an integrated European approach, Thessaloniki, 1999
IUSS Working Group WRB, 2015. World reference base for soil resources 2014, update 2015. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports No. 106. FAO, Rome
Jandl R, Lindner M, Vesterdal L, Bauwens B, Baritz R, Hagedorn F, Johnson DW, Minkkinen K, Byrne KA (2007) How strongly can forest management influence soil carbon sequestration? Geoderma 137:253–268
Janušauskaitė D, Baliuckas V, Dabkevičius Z (2013) Needle litter decomposition of native Pinus sylvestris L. and alien Pinus mugo at different ages affecting enzyme activities and soil properties on dune sands. Baltic For 19(1):50–60
Jasińska J, Sewerniak P, Markiewicz M (2019) Links between slope aspect and rate of litter decomposition on inland dunes. CATENA 172:501–508. https://doi.org/10.1016/j.catena.2018.09.025
Johansson MB (1994) Decomposition rates of Scots pine needle litter related to site properties, litter quality, and climate. Can J For Res 24:1771–1781
Karishna MP, Mohan M (2017) Litter decomposition in forest ecosystems: a review. Energ Ecol Environ 2(4):236–249. https://doi.org/10.1007/s40974-017-0064-9
Kim JG (2012) Accumulated organic matter, litterfall production, and decomposition tell us the status of litter dynamics in forests. J Ecol Field Biol 35(2):99–109
Kunhamu TK, Kumar BM, Viswanath S (2009) Does thinning affect litterfall, litter decomposition, and associated nutrient release in Acacia mangium stands of Kerala in Peninsular India? Can J For Res 37:792–801. https://doi.org/10.1139/X09-008
Laiho R, Laine J, Trettin CC, Finér L (2004) Scots pine litter decomposition along drainage succession and soil nutrient gradients in peatland forests, and the effects of inter-annual weather variation. Soil Biol Biochem 36(1095–1):109. https://doi.org/10.1016/j.soilbio.2004.02.020
Lövblad G (2000) Nitrogen deposition now and in the future. In: Bertills U, Näsholm T (eds) Effects of nitrogen deposition on forest ecosystems. Berlings Skogs, Trelleborg, pp 21–28
Måren IE, Karki S, Prajapati C, Yadav RK, Shrestha BB (2015) Facing north or south: Does slope aspect impact forest stand characteristics and soil properties in a semiarid trans-Himalayan valley? J Arid Environ 121:112–123
Mudrick DA, Hoosein M, Hicks RR Jr, Townsend EC (1994) Decomposition of leaf litter in an Appalachian forest: effects of leaf species, aspect, slope position and time. For Ecol Manag 68:231–250. https://doi.org/10.1016/0378-1127(94)90048-5
Olson JS (1963) Energy storage and balance of producers and decomposers in ecological systems. Ecology 44:1–36. https://doi.org/10.2307/1932179
Ozdamar K (2002) Statistical data analysis by using package software. Kaan Press Company, Eskişehir (in Turkish)
Pausas JG (1997) Litter fall and litter decomposition in Pinus sylvestris forests of the eastern Pyrenees. J Veget Sci 8:643–650. https://doi.org/10.2307/3237368
Pourmajidian MR, Jalilvand H, Fallah A, Hosseini SA, Parsakhoo A, Vosoughian A, Rahmani A (2010) Effect of shelterwood cutting method on forest regeneration and stand structure in a Hyrcanian forest ecosystem. J Forest Res 21(3):265–272. https://doi.org/10.1007/s11676-010-0070-7
Prescott CE (2010) Litter decomposition: what controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 101:133–149. https://doi.org/10.1007/s10533-010-9439-0
Prescott CE, Blevins LL, Staley C (2004) Litter decomposition in British Columbia forests: Controlling factors and influences of forestry activities. BC J Ecosyst Manag 5(2):44–57
Qualls RG (2016) Long-Term (13 Years) Decomposition rates of forest floor organic matter on paired coniferous and deciduous watersheds with contrasting temperature regimes. Forests 7(231):1–17. https://doi.org/10.3390/f7100231
Rodríguez PCR, Durán ZVH, Muriel FJL, Martín PFJ, Franco TD (2009) Litter decomposition and nitrogen release in a sloping Mediterranean subtropical agroecosystem on the coast of Granada (SE, Spain): Effects of floristic and topographic alteration on the slope. Agr Ecosyst Environ 134:79–88. https://doi.org/10.1016/j.agee.2009.05.019
Sariyildiz T (2003) Litter decomposition of Picea orientalis, Pinus sylvestris and Castanea sativa trees grown in Artvin in relation to their initial litter quality variables. Turk J Agric For 27:237–243
Sariyildiz T (2008) Effects of tree canopy on litter decomposition rates of Abies nordmanniana, Picea orientalis and Pinus sylvestris. Scand J For Res 23:330–338. https://doi.org/10.1080/02827580802275816
Sariyildiz T, Kucuk M (2008) Litter Mass Loss Rates in Deciduous and Coniferous Trees in Artvin, Northeast Turkey: Relationships with Litter Quality, Microclimate, and Soil Characteristics. Turk J Agric For 32:547–559
Sariyildiz T (2014) Biochemical and Environmental Controls of Litter Decomposition. Forest Res 3:e109. https://doi.org/10.4172/2168-9776.1000e109
Sariyildiz T (2015) Effects of tree species and topography on fine and small root decomposition rates of three common tree species (Alnus glutinosa, Picea orientalis and Pinus sylvestris) in Turkey. For Ecol Manag 335:71–86
Schönenberger W, Brang P (2004) Silviculture in Mountain Forests. In: Burley J, Evans J, Youngquist JA. (eds) Encyclopedia of Forest Science. Academic Press, London
Singh S (2018) Understanding the role of slope aspect in shaping the vegetation attributes and soil properties in Montane ecosystems. Trop Ecol 59(3):417–430
Sun Z, Liu L, Peng S, Peñuelas J, Zeng H, Piao S (2016) Age-Related Modulation of the Nitrogen Resorption Efficiency Response to Growth Requirements and Soil Nitrogen Availability in a Temperate Pine Plantation. Ecosystems 19(4). https://doi.org/10.1007/s10021-016-9962-5
Taylor BR, Prescott CE, Parsons PWJ, Parkinson D (1991) Substrate control of litter decomposition in four Rocky Mountain coniferous forests. Can J Bot 69:2242–2250. https://doi.org/10.1139/b91-281
Tian DL, Peng YY, Yan WD, Fang X, Kang WX, Wang GJ, Chen XY (2010) Effects of thinning and litter fall removal on fine root production and soil organic carbon content in Masson pine plantations. Pedosphere 20(4):486–493. https://doi.org/10.1016/S1002-0160(10)60038-0
Wang W, Page-Dumroese D, Jurgensen M, Tirocke J, Liu Y (2018) Effect of forest thinning and wood quality on the short-term wood decomposition rate in a Pinus tabuliformis plantation. J Plant Res. https://doi.org/10.1007/s10265-018-1069-y
Will GM, Hodgkiss PD, Madgwick AI (1983) Nutrient losses from litterbags containing Pinus radiata litter: Influences of thinning, clearfelling, and urea fertilizer. NZ J Forestry Sci 13(3):291–304
Youngblood AP (1991) Radial growth after a shelterwood seed cut in a mature stand of white spruce in interior Alaska. Can J For Res 21:410–413. https://doi.org/10.1139/x91-052
Yue K, García-Palacios P, Parsons SA, Yang W, Peng Y, Tan B, Huang C, Wu F (2018) Assessing the temporal dynamics of aquatic and terrestrial litter decomposition in an alpine forest. Funct Ecol, 1-12. https://doi.org/10.1111/1365-2435.13143
Zhang D, Hui D, Luo Y, Zhou G (2008) Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors. J Plant Ecol 1(2):85–93. https://doi.org/10.1093/jpe/rtn002
Acknowledgements
We thank our colleague Rıza Karataş (MSc) for his help during the fieldwork and data collection.
Funding
The study was funded by Turkish General Directorate of Forestry in a framework of a research project (project no: ESK-09 (6303)/2010–2016).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Handling Editor: Andreas Bolte
Contribution of the co-authors Conceptualization: Aydın ÇÖMEZ, Şükrü Teoman GÜNER, Doğanay TOLUNAY; Methodology: Aydın ÇÖMEZ, Şükrü Teoman GÜNER, Doğanay TOLUNAY; Validation: Aydın ÇÖMEZ, Şükrü Teoman GÜNER; Formal Analysis: Aydın ÇÖMEZ; Investigation: Aydın ÇÖMEZ; Resources: Aydın ÇÖMEZ, Şükrü Teoman GÜNER; Data curation: Aydın ÇÖMEZ, Şükrü Teoman GÜNER; Writing – original draft: Aydın ÇÖMEZ; Writing – review & editing: Şükrü Teoman GÜNER, Doğanay TOLUNAY; Visualisation: Aydın COMEZ, Şükrü Teoman GÜNER, Doğanay TOLUNAY; Project Administration: Aydın ÇÖMEZ; Funding acquisition: Turkish General Directorate of Forestry
Appendix
Appendix
Rights and permissions
About this article

Cite this article
Çömez, A., Güner, Ş.T. & Tolunay, D. The effect of stand structure on litter decomposition in Pinus sylvestris L. stands in Turkey. Annals of Forest Science 78, 19 (2021). https://doi.org/10.1007/s13595-020-01023-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13595-020-01023-2