
Abstract
Converting road verges and Public Open Space (POS) to floral-rich resources is an important strategy for supporting populations of urban pollinators. One of the ways this can be achieved is through adjusting mowing schedules, either with a “delayed start” after the end of winter, or a reduction in mowing frequency. In Mediterranean climate ecosystems with mild, wet winters, plant growth continues through the winter season and the definition of a “delayed start” is unclear. This study sought to identify a strategic period for the suspension of mowing activities in a city in South Africa with a Mediterranean climate. It estimated the duration of the flowering to seed-broadcast season of 20 species of native geophytes. Observations were made of phenological status from bud to seed broadcast in ten city parks in Cape Town during the austral spring flowering seasons of 2019 and 2020 (August – December). The Underhill and Zucchini (1988) Moult Model was employed to estimate the duration of the flowering season. Model results showed that mowing should stop in the second week of August and should not resume until the beginning of November in the study context. The results are discussed against the relative biodiversity contribution that different parks and verges make and the potential conflicts with the utility objectives of urban society.
Similar content being viewed by others


Introduction
Parks and road verges form an important component of natural and green infrastructure in cities. They can attract a richness of fauna, particularly insects and birds, when the quality of the landscaping provides suitable habitats (Shwartz et al. 2013; Dylewski et al. 2020; Samways et al. 2020). Community parks and greenways of all sizes therefore present an opportunity for biodiversity “stepping stones” between larger nature parks and conserved natural areas (Goodness 2018), especially for insects and other pollinators (Threlfall et al. 2015; Hall et al. 2017; Banaszak-Cibicka et al. 2018; Davis et al. 2017).
One of the ways this potential can be managed to provide favourable landscapes for pollinators is through the provision of floral-rich lawns (Davis et al. 2017). Maintaining lawns with moderate to low frequency mowing events increases biodiversity and foraging resources for pollinators and represents a practical, timesaving intervention to support urban pollinator and plant populations (O’Sullivan et al. 2017; Lerman et al. 2018; Yang et al. 2019; Watson et al. 2020). In Europe, delaying the first post-winter mowing event from spring to summer has produced positive outcomes for local plant species richness (Humbert et al. 2012). Studies advocating for a “delayed start” to the mowing season have been predominantly carried out in European and North American cities that experience freezing temperatures and snowfall in winter (Watson et al. 2020). In contrast, cities in Mediterranean Climatic Ecosystems (MCE) experience winter rainfall, with snow limited to high-ground, and mowing continues throughout winter (Mooney et al. 2001). Thus, in these contexts, the notion of a “delayed start” to the grass mowing season is a misnomer. There is therefore a need to identify alternative strategies for biodiversity-considered mowing schedules. Some precedent from North America suggests that when moderate to low frequency mowing events are strategically aligned to the life-cycles of specific pollinators and plants, the population abundance of both is improved in the following year (Knight et al. 2019).
The city of Cape Town is located in an MCE in South Africa (Rundel et al. 2016). The city’s municipal horticultural landscaping management minimum standards prescribes norms for municipal contractors who are employed to maintain road verges and POS. Accordingly, these areas are currently mowed at least 10 times a year, resulting in a grass-cutting schedule with intervals of four to six weeks. This schedule means that it is likely that mowing occurs at least once in spring, cutting plants before fruits have matured and seeds have been broadcast, and thereby interrupting the floral reproductive cycle for spring flowers (Manning and Goldblatt 2012). Consequently, this study aims to optimize the duration of a grass-cutting suspension during the austral spring season in Cape Town. Specifically, it provides an indication of the number of weeks for which regular grass-cutting should be suspended during spring. To date, studies in Europe and North America have taken the approach of experimental patch, followed by subsequent biodiversity measurements (Wastian et al. 2016; Sehrt et al. 2020; Watson et al. 2020). In contrast, this study adopted a phenological approach to understanding the ecological processes which support floral diversity. By quantifying the duration of the floral reproductive season, an optimal mowing suspension window is identified.
This study developed methods to generate mowing policy recommendations. The first step was to develop an inventory of the species of flowering plants in each park and classify parks according to functional richness. For each native geophyte species, the phenophases (observable stages in the annual life-cycle of a plant) from budding to seed set were investigated. The Underhill-Zucchini Moult Model (Underhill and Zucchini 1988), an algorithm which was developed to quantify the duration of bird moult, was used to quantify flower phenology (duration of flower reproduction) and based on these results, recommendations were made for a period for each park (or group of parks) during which mowing should not take place. The approach is broadly replicable and can be deployed in other MCEs.
Methods
Phenophase status as an indicator of phenology
Traditionally, phenology was tracked using records of first event dates (e.g. the date on which the first bud of the season was detected) (Miller-Rushing et al. 2008). First-event phenology datasets typically quantify the phenological status of the most extreme individuals within a population of unknown size. Using this method, variations can be erroneously reported when there are differing population sizes or less frequent monitoring and it loses the detail of information about the shifts across the population (Elmendorf et al. 2016). Capturing only the most extreme cases cannot produce recommendations for strategic management outcomes nor provide enough information to balance competing interests and park facility usage trade-offs. Status monitoring, therefore provides more detail about the progress of the life-cycle and can capture repeat events, such as a second flowering flush after a cold snap (Denny et al. 2014; Elmendorf et al. 2016). Furthermore, the adoption of phenophase status and intensity instead of traditional first-event monitoring protocols has advantages (Denny et al. 2014; Elmendorf et al. 2016; Elzinga et al. 2007) Firstly, events that sometimes occur more than once in a year can be monitored. Secondly, variations in transition dates can be measured. Thirdly, the duration of phenophase can be quantified, and fourthly, monitoring small patches and marked individuals ensures that recorded dates are decoupled from population size. This overcomes the weaknesses of first-event monitoring and when used in conjunction with regular sampling enables phenophase change estimates (Elmendorf et al. 2016). It is therefore preferable to track either peak events (Elzinga et al. 2007) or status monitoring (Denny et al. 2014). In this study, phenophase status monitoring was adopted and the peak events and duration of the season were calculated using a statistical model that was originally developed to estimate the starting date and duration of primary moult in birds (Underhill and Zucchini 1988).
The field observation terminology and protocols adopted in this study are based on the USA National Phenology Network (Table 1). This protocol describes a phenophase as “an observable stage or phase in the annual life-cycle of a plant or animal that can be defined by a start and end point”. Phenophases generally have a duration of a few days or weeks. Examples include the period during which newly emerging leaves are visible, or the period in which open flowers are present on a plant“ (USA National Phenology Network, n.d) (Table 1).
Observation season and frequency
Globally, no standardised frequency of phenology monitoring exists. Instead, observation frequency protocols are devised on a case-by-case basis, and depend on the species being studied, the study objectives, and budgetary and logistical constraints (Elmendorf et al. 2016). Mazer et al. (2015) studied four plant species in California across broad environmental conditions, and considered that twice-weekly sampling was sufficient to detect onset dates of vegetation growth, flowering, and fruiting. Miller-Rushing et al. (2008) recommended sampling every second day to ensure a 97% probability of detecting significant change in date of flowering; they noted however, that for many studies less frequent sampling may be adequate to determine simple trend detection. At the opposite end of the scale, sampling every two weeks was recommended to monitor tropical tree phenology (Harrison et al. 2019). Because the purpose of this study is to give an indication of the number of weeks for which regular grass-cutting should be suspended during spring, a weekly fieldwork protocol was used. The intention of this study was to make recommendations relating to park and road verge management practices in an urban context, it was consequently not concerned with the most extreme early flowering individuals but rather the peak events. Peak events are defined as the point in time when the majority of a population are in a particular phenophase (observable stage in the life-cycle of an organism). The Underhill-Zucchini Moult Model (Underhill and Zucchini 1988) which we used, can predict the start dates of the season, even if they have already started when observations begin.
Study area and site selection
The study area, Cape Town, falls within the Cape Floristic Region and contains Renosterveld, Strandveld and Fynbos vegetation types (Mucina & Rutherford 2006; Rebelo et al. 2011). Renosterveld and Fynbos are particularly rich in geophytes (Rebelo et al. 2006; Manning and Goldblatt 2012), the majority of which emerge and flower during the austral spring season (August–November) (Manning and Goldblatt 2012). Cape Town is a major metropolitan and has small (0.1 ha) to medium (10 ha) parks throughout its urban fabric (City of Cape Town 2012). Many of these retain populations of native geophytes and annuals that emerge in the lawns and flower during spring (Manning and Goldblatt 2012;).
Temperature, and to a lesser extent, rainfall, vary across the city (Cowling et al. 1996; Wilkinson 2000) and, in order to capture and control for this variability, we first consulted data from the CHIRPS satellite, and selected areas that were similar in rainfall (de Sousa et al. 2020). Mean annual summer daytime surface temperatures were extracted from the Yale Urban Heat Index (2018) and demonstrated that the temperature variations were more likely to have an impact than the rainfall variations for most parts of the city. Two groups of parks were selected from contrasting parts of the city. Park area managers were approached to identify suitable parks under their jurisdiction in which there were a known abundance of spring flowers and geophytes. The parks were then visited to establish and confirm suitability.
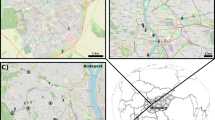
Neighbours and representatives from community stewardship groups were approached to obtain permission and co-operation in the project. Several of the larger park communities have formed “Friends of” groups or “Associations” which volunteer to undertake, and advocate for park upgrades, or coordinate volunteer activities. These groups were approached to notify them of the research activities and request co-operation; in total ten parks were chosen (Fig. 1). Mean summer temperatures varied by up to four degrees across the city. Four parks were identified as suitable parks in the “cool” precinct of the city and five parks were identified as suitable in the northern “warm” (< 2 degrees warmer) precinct of the city.

Location of survey parks in Cape Town, South Africa, where experimental no-mow plots were established in the austral spring of 2019 and 2020. Observations were made of the phenophases of flower bud emergence to the onset of seed broadcasting. A = Warm precinct, B = Cool precinct
Field observation methods
Phenological observations were made on 20 focal species in ten parks during the austral spring seasons of 2019 and 2020 (Tables 2 and 3). At two large parks with an abundant and diverse population of flowers, and already benefitting from a mowing suspension during spring, a series of transects were marked out along which flowers were counted in a 300 mm wide swathes. At each of the rest of the parks two plots of 5 × 5 m were staked out and “Do not mow” signs were placed in the middle of the plots. A general information sign, with contact details about the project, was placed in or near to the plots. Plots were positioned to cover stands of lawn flowers. Plots and transects were visited weekly during spring. Observations began in the second week of August 2019 and 2020, allowing for mowing to occur in the first week of August. Observations continued until more than 50% of the fruit population had cracked open. Counts were recorded of the heads of buds, open flowers, senescent flowers, fruits and open seed capsules for each species in the plots. When they were at their most abundant, and counting individual flower-heads became challenging, 1 × 1 m quadrats were used to keep track of counts and make estimates for the study area. They were recorded on the field observation sheet (Supplementary material 1) and tabulated in a spreadsheet. At the end of the observation season, mowing resumed. The plots were re-established in August of 2020 for a second observation season.
Analysis
An adaptation of the Moult Model of Underhill and Zucchini (1988) was used to describe flowering phenology. The original model was developed to estimate the timing of primary moult in birds. In that application, birds are observed in one of three categories; each observation has a date attached to it: birds with old feathers, which will commence moult on some unknown future date; birds in which the stage of development of the feathers can be quantified through a moult index which measures the stage of progress; birds with new feathers, which completed moult at some unknown past date. The Moult Model uses the method of maximum likelihood to estimate three parameters: the average start date of moult, the variability in the average start date and the duration of moult. The statistical problem of bird moulting is unusual because no birds are observed starting to moult, and the duration is therefore not observable either. The model undertakes the statistics required to estimate these parameters which are not directly observable. Erni et al. (2013) developed software in R (R Core Team 2021) which provides an algorithm which estimates the parameters of moult, and which enables model selection to be undertaken within a standard statistical framework. Here we used the Moult Model to describe flowering phenology. The model is suitable for this task in that with weekly monitoring one does not necessarily observe the transitions between the phenophases as described in Table 1. What is observable is the phenophase of a particular flower. Thus, a sample of plants at a site on each visit can be examined and the number of flowers in each phenophase counted.
The key parameters in this case are the mean start date of budding and the variability of this start date, which measures the extent of synchronicity of the phenomenon. The flower data are analogous to moult data with the following phases: (1) observations of plants which are not yet in bud and which will start some unknown time in the future (analogous to birds that have not yet started to moult); (2) observations of plants which have completed seed set at some unknown time in the past (analogous to birds which have completed moulting); and (3) observations of plants in the phenophases of Table 1 (analogous to actively moulting birds). In applications of the Moult Model to the primary moult of birds, the time spent in each phase has generally been regarded as equal, and this has simplified the calculation of the moult index. However, Redfern (1998) proposed a more nuanced approach for birds, in which he estimated the time in each stage of moult, and Beltran et al. (2019) pioneered a similar approach to quantifying the moult of Weddell seals Leptonychotes weddelli, by estimating the time spent in each moult stage. Our phenology index similarly estimated the number of days which each species spent in each phenophase and converting these to proportions (Table 2). As done for the moult indices, the phenology index for a particular stage was calculated by adding the proportions of time spent in preceding stages, and then adding half the proportion of the current stage. The value for flowers that had achieved seed broadcast was set at 1, analogous to the concept of “completed moult” in the moult model.
Twenty species of geophytes were selected for analysis based on the size of the populations (minimum 30 individual plants, and presence in more than one observation park) and an upright growth form which would be negatively impacted by grass-cutting. The R package Moult (Erni et al. 2013) was then used to compute the total flower reproductive duration from bud appearance to 50% fruit-crack/seed-broadcast. The Moult Model was run for the entire population first, and then separately on all species individually allowing both the start date and duration of the season to vary for each species. The R-script instructed the algorithm that the data were “type = 4”: A sample that identifies [flowers] which are in flowering phases and have completed broadcasting. No counts were made of numbers of plants in vegetative state prior to producing buds, thus this was the appropriate type to specify.
To determine if there was a meaningful difference between the phenophases of the warmer northern district population and the cooler south-western district, the model was run a further three more times on the total sample. It was first run holding the start date and the standard deviation of the start date constant whilst comparing the duration, then holding constant the duration and the standard deviation of the start date, and lastly, holding the duration and start date constant whilst comparing the standard deviations of the start date. A log-likelihood test was run on the results of the Moult Model to determine the model fit.
Results
Each observation patch contained between three and 12 focal species. Six species occurred at only one park. The most widespread species were Romulea rosea, Moraea miniata and Baeometra uniflora which each occurred at eight of the 10 observed parks. The number of observations made per species across both seasons varied between 183 for the elusive Moraea tripetala which had a small population that was difficult to track in Keurboom Park, to 12,005 observations for Sparaxis bulbifera which had strong populations at five of the parks.
Using the adapted Moult Model, the earliest estimated mean start date was 31 July for Wachendorfia multiflora (SE 1.4) and the latest mean start date was 22 September for Ornithogalum thyrsoides (SE 1.1) (Table 3). Half of the focal species (10), started budding in the fourth week of August. The standard deviation of the start dates was smallest for populations of Gladiolus carinatus (7), Ornithogalum thyrsoides (9) and Pterygodium catholicum (8). In contrast, the greatest standard deviation of the start date was recorded for Pauridia capensis (34), followed by 19 days for each Wachendorfia multiflora, Pauridia serrata and Moraea neglecta. The latest estimated mean seed broadcast date (more than 50% of fruiting bodies releasing seeds) was 22 November for Pauridia capensis.
The mean duration of the total measured phenophases from bud emergence to seed broadcast, “mean season duration”,across all species occurred between 14 August and 30 October with a standard deviation of 19 days. Gladiolus carinatus had the shortest mean season duration (28 days) and Pauridia capensis, Moraea tripetala, Wachendorfia multiflora and Moraea neglecta had the longest seasons at 85–98 days. With the exception of Wachendorfia multiflora, this group was relatively uncommon in city parks and tended to occur in sites with biodiversity stewardship plans in place, or low-intensity management practices (e.g. in vacant lots and undeveloped land). Parks with biodiversity stewardship plans in place had engagement from civil society for the protection of designated biodiversity areas during the flowering season. Of the common species (occurring at three or more parks), B. uniflora had the longest seasonal duration at 84 days and was trailed by a cluster of seven species with durations of 60–70 days. Although the mean duration of the flowering season was 77 days across all species, more than half of the species had durations between 50 and 70 days.
Observations of the impact of grass-cutting in late August and September on the remainder of the park (outside the no-mow plots) demonstrated that 30% of growing tips of Sparaxis bulbifera, B. uniflora and Moraea vegeta were lost in August and a further 30% of the tops of plants were cut without removing the growing tip, the remainder were undamaged. In contrast, 100% of flowering heads were lost by the September cut and did not regrow new buds, consequently losing the seed set for the reproductive season.
By the beginning of November, most species had begun broadcasting seeds and had some opportunity for dispersion (See mean end date in Table 3). Those species which had not started broadcasting by the second week of November occurred at sites of greater diversity with existing mowing suspensions in place (three of the ten parks, and where more than 10 species of angiosperm were present) and may be indicators for special management intervention.
The difference between the timing of the season duration (measured phenophases) in the warm district and the cooler areas was less than a week. This was not deemed to be biologically meaningful for the purposes of informing city wide strategic mowing practices.
Discussion
The purpose of city parks is to provide a leisure resource for the relaxation and recreation of citizens, but for many urban-dwellers, parks are the most accessible opportunity for experiencing and connecting to nature (Hand et al. 2017). A sense of connection to nature fosters cues to care and fuels a biophilic value set which can lead to pro-environmental behavioural outcomes (Martin and Czellar 2017). Time spent connecting with nature has psychological and physiological benefits. It decreases hypertension, increases cognitive function, and can mitigate obesity in communities with good access to recreational activities in natural settings (Hartig et al. 2014; Restall and Conrad 2015). What constitutes “good access” is not limited to proximity and convenience but also a function of the quality of the park. Aesthetic quality and biodiversity both contribute to positive nature experiences (Taylor and Hochuli 2015), providing reasons for implementing strategies aimed at improving biodiversity in urban and community parks, but safety is also of importance to determining use and access.
Safety within an urban setting is strongly related to crime prevention which is predicated on the opportunity for legitimate passive surveillance from individuals who feel a sense of ownership and territoriality about the park: neighbours, parents, child-minders and concerned citizens. Passive surveillance is achieved by maintaining sightlines and ensuring optimum usage (Zavadskas et al. 2019). Sightlines can become obscured by long grass (and overgrown shrubs) and hence, regular mowing is required. In addition to obscuring sightlines (thereby obstructing passive surveillance), long grass fuels the perception of degradation and lack of care (Türkseven Doğrusoy and Zengel 2017; Zavadskas et al. 2019). Parks should therefore aspire to a balance between utility, aesthetics, conservation and cues to care to ensure desired usage volumes (enough for passive surveillance, but not crowding), quality and accessibility (Li and Nassauer 2020; van den Berg and van Winsum-Westra 2010; Marshall et al. 2020).
Venn and Kotze (2014), in considering no-mow areas as a biodiversity-supporting solution, acknowledged the potential loss of utility to parks and proposed a strategic mowing schedule which first involved identifying lawns required for sitting or play and distinguishing them from lawns which can be landscaped as biodiversity meadows. The latter group can then be maintained with “benign neglect” to encourage biodiversity (Venn and Kotze 2014; Wastian et al. 2016). They proposed a two-tier strategy where leisure lawns are intensively mowed (7–10 times per year) and biodiversity lawns are extensively mowed (bi-annually) (Venn and Kotze 2014).
The proposed strategic halt for the duration of the flower to seed broadcast cycle, would be in-line with a “delayed start” mowing philosophy (Humbert et al. 2012). Therefore, this study set out to establish the length of time it takes for seed broadcast to begin in spring geophytes in order to inform better mowing practices for supporting urban populations of spring-flowering geophytes and their pollinators. The study adopted a model originally designed to estimate the duration of primary moult season in birds (Underhill and Zucchini 1988). Unlike passerine birds, the growth period for each phenophase of feather growth is non-linear. Therefore, the Moult Model introduces an index of assumed proportion of time that each phase of feather growth occupies within the greater progress of moult (Underhill and Zucchini 1988). This offers a level of flexibility for calculating phenological duration which, through the use of the estimated index, is adapted to different species. Instead of stages in feather development, stages in seed development were identified making it possible to transfer the algorithm and apply it to plants. This was a novel approach to the study of plant phenology and provides a valuable backbone for being able to estimate seasonal duration (van der Valk 2011).
Developing a strategic approach
This study proposes that an assessment of the relative quality and usage of each park is carried out beginning with a survey of the presence of spring-flowering geophytes and annuals. Citizen science can be mobilized as a powerful resource for determining the floral richness of each park and developing an inventory of native bulbs and annuals. The availability of crowd-sourced, geo-spatially available data makes the logistics of urban plant inventories accessible for this kind of biodiversity classification (Barve et al. 2020). Data were collected on floral richness in 142 randomly selected parks in 2017 and 2018 (Brom 2022). From those data, parks can be classified into four categories: Type (1) Parks which contained three or fewer species of flowers from the most common group of exotic weeds and annuals; Type (2) Parks which contained between three and 10 species of native angiosperms, the majority of which are geophytes; Type (3) Parks which contained more than 10 species of native flowers; and Type (4) Parks which contained summer species and/or perennials. In this study, most of the observation parks were type 2. Keurboom Park, was a type 3 park, and Jack Muller and St John’s POS were type 4 parks.
Type 1 parks require no grass-cutting suspension unless undergoing specific landscaping or rehabilitation treatments. This is because the common plants in this group have recovery strategies which make them well adapted to surviving frequent disturbance, over-grazing, and mowing events (Brom 2022). Type 2 parks benefit from the grass-cutting suspension window identified within the results of this study, namely August to mid-November. Type 3 parks would benefit from conservation or management interventions, rehabilitation, and invasive species management. Type 4 parks require further research and assessment for either a second mowing suspension window or alternative management strategies.
Four species within the study broadcast seed in late spring and were outliers to the median season of the focal species. Most notable was Moraea neglecta, a charismatic species which was enthusiastically pointed out by citizens at the parks where it occurred. The species is both relatively uncommon in the city and tends to occur on Type 3 parks justifying citizen stewardship. A charismatic species, it can be managed with engagement from the local community and volunteer stewards with ease; however, seed broadcast had already begun by the end of the recommended mowing suspension window and so even without special management, populations would benefit from the proposed suspension.
For less charismatic species, a more creative intervention may be required. Visibility of plants from the tractor seat is a likely impediment to the success of a limited area mowing suspension and a clear landscape architectural language needs to be developed to signal different mowing responses. For example, the early onset of the Dimorphotheca pluvialis (Fig. 2) season at one park provided a visible marker for communicating to the contractor the area where the community regularly encountered a flush of spring flowers. Boosting lawns with bold floral markers, easily recognizable in the field could provide an affordable and simple way of signalling no-mow areas if patches were systematically sewn with species that flowered for the full duration of the season. Unfortunately, no single flower is in bloom for the entire season and so using floral markers would require a mix of species flowering at different times in order to support seed set success. Ursinia nana follows Dimorphotheca pluvialis and the parasol seeds have a structure which resembles flowers. A mix of these two abundant, rapid-reseeders, may provide the markers needed to protect geopyte-rich lawns through the spring season in Cape Town. Further research is needed to establish appropriate native seed mixes in other regions.
A common charismatic indicator species
The combination of Baeometra uniflora’s commonality, charismatic growth form (Fig. 2B) and long fruiting phase (mean end date 5 November), make this a suitable plant to use as an indicator species for capturing the local variation in phenological occurrences. The thick fruiting bodies are easy to identify and take longer to mature than most of the other focal species. These qualities mean that attentive contractors can wait for B. uniflora to open as an indicator for when the common bulbs and annuals have already begun seed broadcasting.

A. A field of the annual Dimorphotheca pluvialis was used by community members in some parks to communicate to the tractor driver where the boundaries of a spring no-mow test site should start. B. The geophyte, Baeometra uniflora in bud, flower and fruit phenophases which has the longest fruiting phase of the observed flowers and, when the fruiting bodies crack, can be used as an indicator for the end of the spring flower season
Conclusions and recommendations
Maintaining lawns in public space and road verges for their biodiversity and pollinator habitats as floral-rich resources, is an affordable and accessible way of boosting urban insect and angiosperm biomass. Other studies have found that improved pollinator and insect populations can be achieved through benign neglect, and low-intensity mowing schedules (Davis et al. 2017; O’Sullivan 2017; Venn and Kotze 2014), but a total moratorium on grass-cutting in road verges and public space is undesirable due to safety and weed-control requirements (Türkseven Doğrusoy and Zengel 2017; Zavadskas et al. 2019). In parts of the world that experience frost or snow in winter, which prevents mowing, a delayed start to spring mowing has demonstrated biodiversity benefits in the following year (Humbert et al. 2012), however this is not practical in regions that do not experience winter freezing.
In this study, we observed the phenology of flower phases and employed the Moult Model of Underhill and Zucchini (1988) to establish the duration of the period from bud to seed broadcast of austral spring flowers in a city in a Mediterranean Climate Ecosystem. In the case of the commonly-occurring native spring-flowering species (in this case austral), the flowering season lasted from August to the second week of November. A classification system is proposed based on structural diversity. We propose that in the study city, for parks containing three to ten species of spring-blooming flowers, mowing is suspended from the second week of August to the beginning of November. Beaometra uniflora emerged as a good indicator species for when the season is over. It is easily identifiable, relatively common, and has the longest fruiting period of the spring-flowering geophytes. For parks containing more than ten species of indigenous plants, the feasibility of setting aside permanent no-mow biodiversity areas within the park should be explored.
Implementing these strategies, would require minor revisions to the city’s minimum standards to accommodate mowing suspensions without increasing frequency of mowing at other times of the year. Instead, resources should be deployed to managing invasive weeds during the spring season. Engagement with community organisations such as park associations can boost a city’s capacity for fine-tuning mowing and maintenance practices to balance potentially conflicting community needs for utility and environmental stewardship. Lastly, it is recommended that a system of landscape markers be developed to aid tractor-drivers in recognizing areas which have been earmarked for a mowing suspension.
The methods recorded in this study for using phenology to define a strategic mowing suspension are applicable to Mediterranean Climatic Ecosystems that experience winter rainfall and a distinct spring flowering season.
Data Availability
Data is available on request from the corresponding author.
Code Availability
Code was constructed in RStudio and is available on request from the corresponding author.
References
Banaszak-Cibicka W, Twerd L, Fliszkiewicz M, Giejdasz K, Langowska A (2018) City parks vs. natural areas - is it possible to preserve a natural level of bee richness and abundance in a city park? Urban Ecosyst 21(4):599–613. https://doi.org/10.1007/s11252-018-0756-8
Barve VV, Brenskelle L, Li D, Stucky BJ, Barve NV, Hantak MM, McLean BS, Paluh DJ et al (2020) Methods for broad-scale plant phenology assessments using citizen scientists’ photographs. Appl Plant Sci 8(1):1–10. https://doi.org/10.1002/aps3.11315
Beltran R, Kirkham A, Breed G, Testa J (2019) Reproductive success delays moult phenology in a polar mammal. Sci Rep 9(1):5221. https://doi.org/10.1038/s41598-019-41635-x
Brom P (2022) Monkey Beetles on the Beat: Urban monkey beetles reveal opportunities for pollinator habitat management. PhD Thesis. OpenUCTLibrary
City of Cape Town (2012) Zoning Scheme Regulations – a component of the policy-driven land use management system. City of Cape Town, Cape Town
Cowling RM, MacDonald IAW, Simmons MT (1996) The Cape Peninsula, South Africa: physiographical, biological and historical background to an extraordinary hot-spot of biodiversity. Biodivers Conserv 5(5):527–550. https://doi.org/10.1007/BF00137608
Davis AY, Lonsdorf EV, Shierk CR, Matteson KC, Taylor JR, Lovell ST, Minor ES (2017) Enhancing pollination supply in an urban ecosystem through landscape modifications. Landsc Urban Plann 162:157–166. https://doi.org/10.1016/j.landurbplan.2017.02.011
de Sousa K, Sparks A, Ashmall W, van Etten J, Solberg S (2020) Chirps: API client for the CHIRPS Precipitation Data in R. J Open Source Softw 5(51):2419. https://doi.org/10.21105/joss
Denny EG, Gerst KL, Miller-Rushing AJ, Tierney GL, Crimmins TM, Enquist CAF, Guertin P, Rosemartin AH et al (2014) Standardized phenology monitoring methods to track plant and animal activity for science and resource management applications. Int J Biometeorol 58(4):591–601. https://doi.org/10.1007/s00484-014-0789-5
Dylewski Ł, Maćkowiak Ł, Banaszak-Cibicka W (2020) Linking pollinators and city flora: how vegetation composition and environmental features shapes pollinators composition in urban environment. Urban Forestry & Urban Greening 56:126795. https://doi.org/10.1016/j.ufug.2020.126795
Elmendorf SC, Jones KD, Cook BI, Diez JM, Enquist CAF, Hufft RA, Jones MO, Mazer SJ et al (2016) The plant phenology monitoring design for the National Ecological Observatory Network. Ecosphere 7(4):e01303. https://doi.org/10.1002/ecs2.1303
Elzinga JA, Atlan A, Biere A, Gigord L, Weis AE, Bernasconi G (2007) Time after time: flowering phenology and biotic interactions. Trends Ecol Evol 22(8):432–439. https://doi.org/10.1016/j.tree.2007.05.006
Erni B, Bonnevie BT, Oschadleus HD, Altwegg R, Underhill LG (2013) Moult: an R package to analyze moult in birds. J Stat Softw 52(8):1–23. https://doi.org/10.18637/jss.v052.i08
Goodness J (2018) Urban landscaping choices and people’s selection of plant traits in Cape Town, South Africa. Environ Sci Policy 85:182–192. https://doi.org/10.1016/j.envsci.2018.02.010
Hall DM, Camilo GR, Tonietto RK, Ollerton J, Ahrné K, Arduser M, Ascher JS, Baldock KCR et al (2017) The city as a refuge for insect pollinators. Conserv Biol 31(1):24–29. https://doi.org/10.1111/cobi.12840
Hand KL, Freeman C, Seddon PJ, Recio MR, Stein A, van Heezik Y (2017) The importance of urban gardens in supporting children’s biophilia. Proc Natl Acad Sci 114(2):274–279. https://doi.org/10.1073/pnas.1609588114
Harrison T, Gibbs J, Winfree R (2019) Anthropogenic landscapes support fewer rare bee species. Landsc Ecol 34(5):967–978. https://doi.org/10.1007/s10980-017-0592-x
Hartig T, Mitchell R, de Vries S, Frumkin H (2014) Nature and health. Annu Rev Public Health 35(1):207–228. https://doi.org/10.1146/annurev-publhealth-032013-182443
Humbert J-Y, Pellet J, Buri P, Arlettaz R (2012) Does delaying the first mowing date benefit biodiversity in meadowland? Environ Evid 1(1):9. https://doi.org/10.1186/2047-2382-1-9
Knight SM, Norris DR, Derbyshire R, Flockhart DTT (2019) Strategic mowing of roadside milkweeds increases monarch butterfly oviposition. Global Ecol Conserv 19:e00678. https://doi.org/10.1016/j.gecco.2019.e00678
Lerman SB, Contosta AR, Milam J, Bang C (2018) To mow or to mow less: lawn mowing frequency affects bee abundance and diversity in suburban yards. Biol Conserv 221:160–174. https://doi.org/10.1016/j.biocon.2018.01.025
Li J, Nassauer J (2020) Cues to care: a systematic analytical review. Landsc Urban Plann 103821. https://doi.org/10.1016/j.landurbplan.2020.103821
Manning J, Goldblatt P (2012) Plants of the Greater Cape Floristic Region 1: The Core Cape flora. In Strelitzia Pretoria: South African National Biodiversity Institute. ISBN: 9781919976747
Marshall AJ, Grose MJ, Williams NSG (2020) Of mowers and growers: perceived social norms strongly influence verge gardening, a distinctive civic greening practice. Landsc Urban Plann 198:103795. https://doi.org/10.1016/j.landurbplan.2020.103795
Martin C, Czellar S (2017) Where do biospheric values come from? A connectedness to nature perspective. J Environ Psychol 52:56–68. https://doi.org/10.1016/j.jenvp.2017.04.009
Miller-Rushing AJ, Inouye DW, Primack RB (2008) How well do first flowering dates measure plant responses to climate change? The effects of population size and sampling frequency. J Ecol 96(6):1289–1296. https://doi.org/10.1111/j.1365-2745.2008.01436.x
Mazer SJ, Gerst KL, Matthews ER, Evenden A (2015) Species-specific phenological responses to winter temperature and precipitation in a water-limited ecosystem. Ecosphere 6(6):98.https://doi.org/10.1890/ES14-00433.1
Mooney HA et al (2001) Mediterranean-Climate Ecosystems. In: Chapin, F.S., Sala, O.E., Huber-Sannwald, E. (eds) Global Biodiversity in a Changing environment. Ecoloical Studies. 152. Springer, New York, NY. https://doi.org/10.1007/978-1-4613-0157-8_9
Mucina L, Rutherford M, Eds (2006) The vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19. South African National Biodiversity Institute, Pretoria
O’Sullivan OS, Holt AR, Warren PH, Evans KL (2017) Optimising UK urban road verge contributions to biodiversity and ecosystem services with cost-effective management. J Environ Manage 191:162–171. https://doi.org/10.1016/j.jenvman.2016.12.062
R Core Team (2021) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rebelo AG, Boucher C, Helme N, Mucina L, Rutherford MC (2006) Fynbos Biome. In: Mucina MC, Rutherford L (eds) The vegetation of South Africa, Lesotho and Swaziland. South African National Biodiversity Institute, Eds. Pretoria, pp 53–220
Rebelo AG, Holmes PM, Dorse C, Wood J (2011) Impacts of urbanization in a biodiversity hotspot: Conservation challenges in metropolitan Cape Town. S Afr J Bot 77(1):20–35. https://doi.org/10.1016/j.sajb.2010.04.006
Redfern C (1998) The analysis of primary moult using feather mass. Ringing Migr 19(1):39–40. https://doi.org/10.1080/03078698.1998.9674161
Restall B, Conrad E (2015) A literature review of connectedness to nature and its potential for environmental management. J Environ Manage 159:264–278. https://doi.org/10.1016/j.jenvman.2015.05.022
Rundel PW, Arroyo MTK, Cowling RM, Keeley JE, Lamont BB, Vargas P (2016) Mediterranean biomes: evolution of their vegetation, floras, and climate. Annu Rev Ecol Evol Syst 47:383–407. https://doi.org/10.1146/annurev-ecolsys-121415-032330
Samways MJ, Barton PS, Birkhofer K, Chichorro F, Deacon C, Fartmann T, Fukushima CS, Gaigher R et al (2020) Solutions for humanity on how to conserve insects. Biol Conserv 242:108427. https://doi.org/10.1016/j.biocon.2020.108427
Sehrt M, Bossdorf O, Freitag M, Bucharova A (2020) Less is more! Rapid increase in plant species richness after reduced mowing in urban grasslands. Basic Appl Ecol 42:47–53. https://doi.org/10.1016/j.baae.2019.10.008
Shwartz A, Cheval H, Simon L, Julliard R (2013) Virtual garden computer program for use in exploring the elements of biodiversity people want in cities. Conserv Biol 27(4):876–886. https://doi.org/10.1111/cobi.12057
Taylor L, Hochuli DF (2015) Creating better cities: how biodiversity and ecosystem functioning enhance urban residents’ wellbeing. Urban Ecosyst 18(3):747–762. https://doi.org/10.1007/s11252-014-0427-3
Threlfall CG, Walker K, Williams NSG, Hahs AK, Mata L, Stork N, Livesley SJ (2015) The conservation value of urban green space habitats for australian native bee communities. Biol Conserv 187:240–248. https://doi.org/10.1016/j.biocon.2015.05.003
Türkseven Doğrusoy I, Zengel R (2017) Analysis of perceived safety in urban parks: a field study in Büyükpark and Hasanaga Park. Metu J Fac Archit 34(1):63–84. https://doi.org/10.4305/METU.JFA.2017.1.7
Underhill LG, Zucchini W (1988) A model for avian primary moult. Ibis 130(4):358–372. https://doi.org/10.1111/j.1474-919X.1988.tb00993.x
USA National Phenology Network. n.d. Phenophase Available : https://usanpn.org/taxonomy/term/16 [Accessed: 20 April 2021]
van den Berg AE, van Winsum-Westra M (2010) Manicured, romantic, or wild? The relation between need for structure and preferences for garden styles. Urban Forestry & Urban Greening 9(3):179–186. https://doi.org/10.1016/j.ufug.2010.01.006
Van der Valk R (2011) On estimating the duration of phenological stages in bryophytes. Lindbergia 34:44–50 ISSN 0105–0761
Venn S, Kotze D (2014) Benign neglect enhances urban habitat heterogeneity: responses of vegetation and carabid beetles (Coleoptera: Carabidae) to the cessation of mowing of park lawns. Eur J Entomol 111(5):703–714. https://doi.org/10.14411/eje.2014.089
Wastian L, Unterweger PA, Betz O (2016a) Influence of the reduction of urban lawn mowing on wild bee diversity (Hymenoptera, Apoidea). J Hymenoptera Res 49(7):51–63. https://doi.org/10.3897/JHR.49.7929
Watson CJ, Carignan-Guillemette L, Turcotte C, Maire V, Proulx R (2020) Ecological and economic benefits of low‐intensity urban lawn management. J Appl Ecol 57(2):436–446. https://doi.org/10.1111/1365-2664.13542
Wilkinson P (2000) City profile: Cape town. Cities 17(3):195–205. https://doi.org/10.1016/S0264-2751(99)00059-1
Yang F, Ignatieva M, Wissman J, Ahrné K, Zhang S, Zhu S (2019) Relationships between multi-scale factors, plant and pollinator diversity, and composition of park lawns and other herbaceous vegetation in a fast growing megacity of China. Landsc Urban Plann 185:117–126. https://doi.org/10.1016/j.landurbplan.2019.02.003
Zavadskas EK, Bausys R, Mazonaviciute I (2019) Safety evaluation methodology of urban public parks by multi-criteria decision making. Landscape and Urban Planning 189(April 2018):372–381. DOI: https://doi.org/10.1016/j.landurbplan.2019.05.014
Funding
This work was supported by the SASAC/NRF scholarship grant number SASAC 170914262644 awarded to Peta Brom.
Open access funding provided by University of Cape Town.
Author information
Authors and Affiliations
Contributions
P.B. wrote the main manuscript text, carried out the fieldwork and conducted the analysis. L.U. provided statistical expertise and oversight. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Conflicts of interest/Competing interests
None.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brom, P.D., Underhill, L.G., Winter, K. et al. A mowing strategy for urban parks to support spring flowers in a mediterranean climate city in South Africa. Urban Ecosyst 26, 1143–1153 (2023). https://doi.org/10.1007/s11252-023-01369-1
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-023-01369-1