
Abstract
Trade-offs between high stand productivity and good wood quality exist for chestnut coppices and related wood-based products. The main objective of this study was to determine the most suitable duration (in years) of cutting cycles that maximizes stand productivity and preserve wood quality of chestnut coppices in a Mediterranean setting. To this aim, a stand-level growth model was developed to verify if wood quality of chestnut coppices at different stand ages varies when the rotation period is modified. Wood quality and stand productivity were analysed, using a chronosequence approach, in coppice stands in Southern Italy characterized by four cutting cycles (15, 25, 30, and 50 years). Results implied that the culmination of the mean annual increment occurs at 28 years, while the current annual increment culminates 10 years earlier. The MOEd values revealed a negative correlation with shoot age; however, a cutting cycle between 25 and 30 years might represent the best compromise for balancing stand productivity and wood quality. Results are discussed in the context of adaptive forest management.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Forests in the Mediterranean Europe are mainly located in mountain and inner areas and provide a range of ecosystem services with important social–ecological functions (Scarascia-Mugnozza et al. 2000; Pastorella et al. 2016). Coppice forests represent a relevant part of these landscapes (Cutini et al. 2021). The coppice system is extremely efficient, since it offers the benefits of simplified and flexible silvicultural approaches, fast tree regeneration, prompt biomass production (and carbon sequestration), and short cutting cycles and high stress tolerance (Moscatelli et al. 2007). In the second half of the twentieth century, coppice products (firewood, charcoal, paling) have suffered from the competition of industrial products, which has resulted in a decreasing interest on this traditional management system (Hédl et al. 2010). However, growing interests for renewable energy sources and wood products currently make coppices a promising multipurpose silvicultural system for addressing risks associated with climate change and related disturbances (e.g., drought spells, forest fires) in agricultural and forest landscapes of Mediterranean mountains and inner areas, while contributing to community resilience and halting land degradation (Kelly et al. 2015).
Coppice forests have gained a renewed interest in many European countries (Rydberg 2000). The main reason for this are as follows: (1) the increasing importance of renewable energy sources as a substitute for fossil fuels, (2) the preservation of coppice forests as an historical landscape element often characterized by a high nature conservation value (Buckley 1992; Lanuv 2007; Scherzinger 1996; Parisi et al. 2020), (3) the high ecological value of these ecosystems, useful to support a wide diversity of fauna and flora (Zlatanov et al. 2013). These reasons (economic, social, and ecological) may open new perspectives for managing the abandoned coppice stands in many areas of Southern Europe and beyond.
In this context, chestnut (Castanea sativa Mill.), as one of the most important tree species of Southern Europe (Fabbio 2016), has the potential to deliver the multifunctional services of climate smart forestry in mountain areas (Bowditch et al. 2020). Chestnut forests cover more than 2.5 million hectares (Conedera et al. 2016), and their expansion through European centuries over time has been favoured by the multiple uses of chestnut-derived wood and non-wood forest products. Chestnut forests are usually managed as coppice stands, with or without reserve trees, but also as traditional fruit orchards, often in the framework of agroforestry systems. In Italy, chestnut agroforestry systems cover an area of about 800,000 ha (Giannini et al. 2014), 70% of which are managed as coppices, while the remaining 30% are managed as orchards for fruit production (Greco et al. 2018), supporting agricultural systems and rural communities in mountain areas.
Traditionally, in the Mediterranean Europe, chestnut has been successfully managed as coppice forest, which has led to the development of sustainable silvicultural modules, reflecting the capacity of the coppice system to adapt to the market needs and environmental challenges of mountain areas (Ciancio et al. 2004; Patricio et al. 2005). Indeed, the flexibility and reversibility of this silvicultural system meet the market requirements, without compromising the resprouting ability and the provisioning of services (Manetti et al. 2006), e.g., slope stabilization. Depending on the social–ecological conditions, chestnut coppices can be either managed with short rotation periods (12–15–20 years) or with medium-to-long cutting cycles (25–30–50 years) to process a variety of wood-based products, including wood for pole, timber for sawing, firewood, tool handles, and fencing material (Manetti et al. 2009).
Recently, several studies have focused on the effect of coppicing on the overall productivity (and, therefore, carbon sequestration and stock capacity) of chestnut forests (Kneifl et al. 2015; Manetti et al. 2016; Esteban et al. 2018; Marcolin et al. 2020). However, in Europe, there has been less emphasis on investigating growth and yield models in chestnut coppices and wood quality of coppiced trees, with few exceptions based on non-destructive technologies (NDTs) (e.g., Russo et al. 2019, 2020). Referring to the first point, in chestnut coppices in northwest Spain, Menéndez-Miguélez et al. (2014) have developed an interesting and useful model system made by a taper function, a total volume equation, and a merchantable volume equation. On the other hand, the ecological behaviour and productive capacity of chestnut coppices are key factors in predicting tree growth and selecting management approach, especially in areas where timber production is the primary objective (Menéndez-Miguélez et al. (2015). With reference to the second point, stress wave-based non-destructive acoustic techniques have resulted in very useful methods for predicting the mechanical properties of woody materials (Guntekin et al. 2013). Among the parameters measurable by acoustic methods, the most important are the modulus of elasticity of wood (MOE) and the dynamic modulus of elasticity (MOEd), being related to wood anatomy and tree physiology. These parameters are fundamental for the evaluation of wood quality, providing information on the resistance to deflection and the stiffness of material (Teder et al. 2011; Wessels et al. 2011).
To answer questions on the effect of different management approaches and environmental factors on wood quality for a specific forest type in homogeneous environmental conditions, the history of tree growth can be reconstructed along a chronosequence. Chronosequences, assuming space-for-time substitution, aim to infer temporal dynamics from measurements at sites of different ages but similar in land-use histories (Aide et al. 2000; Hedde et al. 2008; Pawson et al. 2009). In forestry, a chronosequence is a set of forest stands that share similar attributes but are of different ages (Johnson and Miyanishi 2008), reflecting a time sequence (Salisbury 1952; Pickett 1989). The key assumption of chronosequences is that each of the sites represents different developmental stages, with the same initial conditions. When there are demonstrable linkages between stages (i.e., the successional trajectory is predictable), chronosequences provide a useful approach to study temporal changes in a long-term perspective (Walker et al. 2010).
In this study, a stand-level growth and yield model for chestnut coppices was developed using a chronosequence approach. Chestnut coppices of different ages, growing in homogeneous environmental (biotic and abiotic) conditions, were selected in the “Aspromonte” National Park (Calabria, Southern Italy). The model, replicable in other geographical contexts, was implemented to verify if the quality and quantity of wood-based products vary with changing rotation periods. More specifically, variation in wood quality and stand productivity of coppiced chestnut was analysed for four different cutting cycles, referring to 15, 25, 30, and 50 years (hereafter C15, C25, C30, and C50, respectively). These four cutting cycles were considered because they are the most used in the chestnut coppices of Calabria. Our study was aimed to answer the following main question: Does the extension of rotation periods induce a variation of wood quality and tree growth of coppiced chestnut? We hypothesized that wood production increased along with the elongation of rotation timespan in chestnut coppice stands in a typical Mediterranean mountainous region, without impairing wood quality.
Materials and methods
Study area
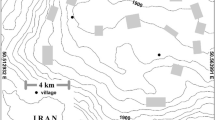
The study area is located in Southern Italy (Calabria), within of the “Aspromonte” National Park, (570,304.25 E; 4,226,245.48 N) at an altitude ranging between 800 and 1100 m a.s.l. (Fig. 1). Soils developed from igneous and metamorphic rocks and are classified as Umbrisols, Cambisols, and Leptosols (FAO 2014), with an udic soil regime moisture. The average annual rainfall is 1605 mm, and the mean annual temperature is 10.6 °C.

Location of the study area in Southern Italy (Calabria) and the applied experimental design
The studied forest stands were characterized by monospecific coppices dominated by chestnut, with an age varying from 6 to 50 years. Stands over 30 years of age were subjected to at least one thinning carried out between the 25th and 30th year of age. Altogether, the analysed plots were considered as chronosequences, based on the assumption that all coppice stands share similar biotic and abiotic conditions and disturbance legacies. These stands can be considered representative of most chestnut coppices, as widespread forest systems in the Mediterranean context.
Data collection and analysis
Data collection was realized in 44 plots located in 16 stands with tree age ranging from 6 to 50 years, arranged through a systematic sampling design. Table 1 shows the number of plots and their extent per each age class, with an extension ranging between 530 and 1200 m2. All plots were chestnut coppices, placed along with chronosequences, following a systematic sample grid, with a total area of 120 ha. On average, each chronosequence was about 7 ha.
In each plot, the following parameters were recorded: (1) number of stools, (2) number of shoots per stump, (3) the diameter at breast height (DBH) of all the shoots, and (4) the total height of 20% of the shoots, homogeneously distributed across the different DBH classes. Table 2 shows the descriptive statistics (mean and standard deviation) of the variables measured in the chestnut coppice stands aged 15, 20, 30, and 50 years.
Furthermore, the MOEd was measured on each shoot with a height greater than 1.5 m in 7 plots for the age class of 15 years, in 4 plots for the age class of 25 years, in 5 plots for the age class of 30 years, and in 5 plots for the age class of 50 years, for a total of 1952 shoots.
To measure the acoustic velocity, the TreeSonic™ (Fakopp Enterprise, Agfalva, Hungary) was used inserting two sensor probes (a transmitting probe and a receiving probe) into the sapwood, then introducing the acoustic energy into the tree through a hammer impact (for further details, Vanninen et al. 1996; Divos 2010; Russo et al. 2019). Three measurements were carried out for each selected shoot and the average of the three recordings was used as the final transit time. The acoustic velocity was then calculated considering the distance between the two sensor probes and the time-of-flight (TOF) data using the following equation:
where CT = tree acoustic velocity (m/s), S = distance between the two probes (sensors) (m), TOF = time-of-flight (s).
Afterwards, the MOEd was calculated, according to the following equation (Eq. 2):
where WDij = wood density (kg m−3), shared by DBH class (i) and age (j), and CT = velocity (m s−1).
Wood density was determined on a subsample of shoots in the 15-, 25-, 30-, and 50-year-old stands; more in detail, we measured wood density on approximately 20% (390) of the total shoots (1952), considering all the diameter classes. In detail, woody cores were extracted at breast height (1.30 m) with a Pressler borer from stems, referring to each of the DBH classes at different ages. The fresh weight and wood volume were measured in the laboratory. The wood volume was calculated with the cylinder formula, measuring the diameter of woody cores with a small electronic precision calliper, and their length with a centimetre to the nearest millimetre. Samples were then weighed to the nearest 0.01 g with an electronic balance. Oven drying of all samples was done at 105 °C to constant weight. Wood density (kg m−3) was calculated by dividing the dry mass by the sample volume.
Statistical analysis
Stand-level growth and yield models were fitted based on (1) the average number of shoots per ha, (2) the diameter-height-age relationship, and (3) the average values of basal area. All models were fitted by the ordinary least square’s method using ‘lm’ function for the R programming language (R Core Team 2019). The criteria used for comparing the models were based on the analysis of goodness-of-fit statistics. The adjusted coefficient of determination (R2 adj), root mean square error (RMSE), and Akaike's information criterion were used to select the best candidate models. In all models, the absence of multicollinearity among the predictors was tested by posing a variance inflation factor (VIF = 3) (Zuur et al. 2010).
In detail, starting from the number of shoots per ha (NS) recorded in each plot and considering Age as independent variable, the best model obtained through a stepwise procedure was the following equation:
The equation coefficients were determined analytically after a logarithmic linearization:
where NS is the number of shoots per ha and AGE represents the stand age (years).
The diameter-height-age model, according to the approach used by Clutter et al. (1983) and Marziliano et al. (2013), was derived using the following function (Eq. (4)):
where Ht = total shoot height (m), NS = number of shoots per ha, dbh = diameter at breast height (cm), AGE = stand age (years).
To estimate the development in basal area, we used the average tree basal area (g) as dependent variable, since the NS (density expression) is structurally included in the stand basal area (G); therefore, in this model, the independent variables were Age and NS per ha. Considering several combinations of these two variables, tested through stepwise procedures, the best model resulted the following equation:
where g = average tree basal area (m2), Age = Age (years), NS = number of shoots per ha.
The stand basal area (G) per ha was obtained multiplying the average basal area (g) estimated with Eq. (6), by the number of shoots (NS), occurring at each specific age, estimated with Eq. (4).
In the simulation of growth and yield of the coppice forests, once identified the initial conditions of the stand (number of shoots per hectare, age, stand mean diameter and height, basal area per hectare), stand volume and aboveground biomass were estimated using the equations reported for chestnut coppices in the National Forest Inventory protocols (INFC 2005; Tabacchi et al. 2011). More in detail, for each age class, the tree volume and aboveground biomass of the average tree (considering diameter and height of the shoots) were first estimated and then multiplied by the number of living shoots. Finally, the mean annual increment (MAI), the current annual increment (CAI), and the percentage of the current annual increment (PCAI) were calculated based on the estimates of tree volumes at the different ages.
Moreover, to estimate the assortments obtainable at different ages, a taper function developed for the chestnut coppices occurring in the region was used (Ciancio et al. 2004). For each diameter class i, using the taper function, we estimated the diameters along the stems at any height from the forest floor to the tree top. For each age, the assortments obtained for the diameter class i were multiplied by the number of trees belonging to the diameter class i. The assortments considered in this study were grouped in four categories (Table 3).
The analysis of variance (ANOVA) was carried out to test the differences in MOEd values obtained for the different ages and in relation to the DBH classes. The significance level of the differences was tested using Tukey’s test. When the significance level (p value) was ≤ 0.05, the null hypothesis was rejected and significant differences in the means were accepted.
Results
Growth and yield model
Values of NS in relation to the stand age are reported in Fig. 2. Table 4 shows the regression statistics, in which all the significant parameters are detailed. Values of NS decreased as tree age increased. More in detail, NS values were very high in the first years after coppicing, then significantly decreased due to competition between shoots occurring on the same stumps. On average, from the occurrence of about 10,000 shoots per ha in the first years after coppicing (5 years), a significant reduction in NS values was observed, halving ten years after cutting. This trend was also confirmed in the following years, reaching about 2000 and 1000 shoots per ha at 25 and 50 years after cutting, respectively. Shoot mortality was very intense up to 20–25 years, significantly reducing afterwards (Fig. 2).

Number of shoots per hectare, observed and estimated, in relation to the stand age
In Fig. 3, the tree height–diameter curve (i.e., the hypsometric relationship) is reported, showing the impacts of age factor. The regression coefficients of Eq. (5) and their standard errors show good accuracy of fitted curves. As expected, different tree heights were observed as ages varied: the height–diameter curve for young shoots (5 years old) was steeper than the other curves (e.g., from 20 to 50 years old).

Tree height–diameter relationship at different ages, estimated through Eq. 5
Table 5 shows results of the growth and yield model obtained for stand age ranging from 5 to 50 years. In Fig. 4, the trend of stand volume over time is reported, as well as MAI and CAI. Tree volumes ranged from about 160 to 535 m3 ha−1 for C15 and C50, respectively. The culmination of MAI occurred at 26 years (14 m3 ha−1 year−1), while CAI culminated about 10 years earlier (22 m3 ha−1 year−1). The percentage increment ranged from 26% at 6 years to 0.4% at 50 years.

Trends of the mean volumes, CAI, and MAI with increasing stand ages
Once the productivity and yield values were obtained, for each rotation cycle examined (C15, C25, C30 and C50), the obtainable wood assortments were estimated, through the taper function developed for the chestnut coppices in the Calabria Region (Ciancio et al. 2004). For C15 (the rotation period usually adopted), about 57% of the wood production consisted of small assortments (diameter ≤ 5 cm), while the small and large poles represented, respectively, 16 and 26% of the obtainable wood products. The yield of larger assortments was negligible (Fig. 4).
For C25 and particularly C30, a significant increase in large poles (respectively, of 42 and 54%) and a significant decrease in small assortments (respectively, of 31 and 28%) were observed. In both C25 and C30, higher quality assortments (beams and boards) were also observed, although to a lesser extent (about 8%). However, for C25 and C30, the prevailing assortment referred to small and large poles, equal to 61 and 63%, respectively (Fig. 4).
For C50, the percentage of beams and boards increased (of about 34%) representing, together with the large poles (35%), most of the obtainable assortments (Fig. 5).

Assortments for the four rotation periods (in percentage)
Wood quality
Results obtained for the MOEd along the chronosequences are shown in Fig. 6. The ANOVA highlighted a significant effect of the length of rotation periods on MOEd values (F3;1929 = 11.769; p ≤ 0.001). The highest values were obtained for C25 (on average 10,422 MPa), which significantly differed from C15 (on average 9536 MPa) and C50 (on average 8274 MPa). For cutting cycles ranging between 25 and 50 years, MOEd values decreased as the rotation period increased (Fig. 6).

Variation of the MOEd values at different stand age. The horizontal line indicates the median values; in the box, the lower limit corresponds to the value of the first quartile (Q1) of the distribution and the upper limit to the third quartile (Q3); the vertical lines (whiskers) delimit the intervals in which the lower values of Q1 (in the lower part) and the greater values of Q2 (in the upper part) are positioned
Figure 7 reports MOEd values recorded for all samples across the different DBH classes. The ANOVA revealed that MOEd values decreased as the DBH increased (F19;1929 = 1.995; p= 0.006); more specifically, with DBH over 30–33 cm, significantly lower MOEd values were observed in comparison with smaller DBH classes.

MOEd values at different DBH, regardless of age. The horizontal line indicates the median values; in the box, the lower limit corresponds to the value of the first quartile (Q1) of the distribution and the upper limit to the third quartile (Q3); the vertical lines (whiskers) delimit the intervals in which the lower values of Q1 (in the lower part) and the greater values of Q2 (in the upper part) are positioned
In Fig. 8, MOEd variation across the different DBH classes and in relation to the stand age is shown. Generally, lower values of MOEd always occurred for larger DBH, while higher values characterized smaller DBH. The ANOVA showed a significant effect of the DBH classes on MOEd values for C15 and C50, while minor effects were observed for C25 and C30.

Variation of the MOEd values as DBH increase for the stands at 15, 25, 30, and 50 years. In the boxplots, the horizontal lines indicate the median values; the lower limit corresponds to the value of the first quartile (Q1) of the distribution and the upper limit to the third quartile (Q3); the vertical lines (whiskers) delimit the intervals in which the lower values of Q1 (in the lower part) and the greater values of Q2 (in the upper part) are positioned. In each graph, the horizontal lines represent the minimum MOEd value recommended (7200 MPa; see Detter et al. 2008)
The MOEd values decreased as the DBH increased for C15, with significant differences observed between the smallest and largest DBH. Particularly, for DBH higher than 12 cm, faster growth trends induced a significant decrease in MOEd values.
On the contrary, for C25 and C30, the MOEd values did not vary significantly when the DBH increased (Fig. 8). Results demonstrated that in C25, but also in C30, though to a lesser extent, the MOEd values aligned, making the wood quality uniform across the different DBH classes.
Finally, in C50, the MOEd values were not only significantly lower than in C25 and C30, but decreased significantly as the DBH increased, especially starting from the 30–33 cm DBH class.
Discussion
We developed and tested a stand-level growth and yield model for simulating the temporal development of the main structural traits for chestnut coppices in the Mediterranean climate change hotspot. Although this study was focused on chestnut coppices at the southernmost distribution limit of the species, predictions revealed high yield potential of these forest stands, confirmed by the considerable wood volumes and growth trends (MAI and CAI). Aspects of coppice productivity have been often ignored in common growth modelling approaches (Vanclay 1994; Pretzsch 2009; Weiskittel et al. 2011). Therefore, this empirical growth and yield model might provide useful insights in production modelling and forest planning applied to chestnut coppices and aimed at balancing stand productivity and wood quality. Nevertheless, the specific geographical context and the complexity of environmental setting in which data were collected may limit comparisons with other modelling exercises (Bernetti 1980; von Gadow and Hui 1999).
Angelini et al. (2013) estimated, on average, MAI ranging from about 7.2 to 13.0 m3 ha−1 year−1 for 18–22-year-old chestnut coppices in Central Italy. Ciancio et al. (2004), for 15–45-year-old chestnut coppices in Calabria, found MAI ranging from 12 to 16 m3 ha−1 year−1. In Tuscany, Cutini (2001) recorded MAI of 17.6 and 12.8 m3 ha−1 year−1, for 15- and 38-year-old chestnut coppices, respectively. In an interesting study in chestnut coppices in North-Western Spain, Menéndez-Miguélez et al. (2016) showed, for high density coppices (7183 stems per hectare at 10 years and 880 stems per hectare at 60 years), MAI values from 8.1 m3 ha−1 at the age of 35 for low site index to MAI values of 38.8 m3 ha−1 at the age of 25 for high site index. Instead, for low density coppices (3366 stems per hectare at 10 years and 484 stems per hectare at 60 years), the MAI values ranged from 5.2 m3 ha−1 at the age of 35–24.3 m3 ha−1 at the age of 25. In both situations (low and high density), the culmination of MAI occurred at a younger age as the site index increased. It is worth noting that our sampling coppice stands, though differing in age, have similar disturbance legacies and occur on similar soil types and environmental conditions within the same climatic zone. Therefore, the commonly used site index (estimated based on stand height) was not considered suitable to characterize site productivity for these homogeneous forest stands (Skovsgaard and Vanclay 2013).
In our study, the optimal rotation length that produces the maximum sustainable yield was to 25 years (about 18 m3 ha−1 year−1). The same rotation length was considered optimal in chestnut coppices in North-Western Spain (Menéndez-Miguélez et al. 2016). This rotation length (25 years) is lower than that reported by Cabrera and Ochoa (1997) (31 years) and by Elorrieta (1949) (30 years) in Spain, and even than those proposed by Bourgeois et al. (2004) and Lemaire (2008) for high quality timber in France (40–45 years). In addition, the MAI estimated in our study at the culmination age (about 18 m3 ha−1 year−1 at 25 years) is higher than that reported in other studies done in different environmental conditions: 11 m3 ha−1 year−1 at 40 years in the Dean Forest in Southern England for the best qualities (Everard and Christie 1995) and 16 m3 ha−1 year−1 at 30 years in France (Bourgeois et al. 2004).
The present study reports model results aligned with previous observations on chestnut coppices. With reference to the basal area, values of 25 m2 ha−1 were observed for 11-year-old chestnut coppices in Tuscany (Manetti et al. 2016), while values ranging from 18 to 42 m2 ha−1 were recorded for 6–22-year-old chestnut coppices in Lazio (Mattioli et al. 2016), both considering Central Italy. Again, results of the present study are consistent with previous research on chestnut coppices and, thus, the model exercise may help implementation processes and scaling procedures in different environmental setting.
The variability in local conditions across different geographical contexts, due to specific environmental factors and different management options, may affect model results. However, the homogeneous environmental setting and land-use history across plots throughout the chronosequence clearly indicated high reliability of the model predictions and simulation of the coppice productivity. Indeed, shoot heights reached 22 m in 50-year-old coppice stands, thus potentially ensuring wood-based products of high economic value. Particularly, the height–diameter curves revealed that, during the earlier stages after coppicing, shoots might show a considerable height increment. In fact, a relevant height–diameter curve differentiation was observed at these growth stages, probably due to the strong competition for light between shoots occurring on each single stool. When the coppice reached an age of 15–20 years and beyond, the height–diameter curve flattened, indicating a high level of spatial competition between shoots (Marziliano et al. 2013, 2019). At these ages, the competition is mainly affected by diametrical differentiation rather than by hypsometric variation, shoots growing more in diameter than in height.
Chestnut is considered the tree species with the highest capacity to provide multiple goods and services among Mediterranean forest species (Giannini et al. 2014), producing a variety of assortments other than firewood, even when it is managed as coppice stand. In this study, we highlighted the great potential of chestnut for producing different wood-based products when the rotation period of coppice stands was extended. In this context, although the organic layer and the mineral soil, as a large carbon stock in forest ecosystems, were not accounted for in this study, implications for carbon sinks and mitigation purposes are also important.
On the other hand, for the same assortments, wood quality traits might significantly vary, depending on the length of rotation periods. In fact, MOEd values revealed a negative trend as stand age increased (from to 25 to 50 years) while such a trend was positive at stand ages growing from 15 to 25 years. Moreover, a negative trend of MOEd values was observed as DBH increased, both in C15 and C50. According to Detter et al. (2008), chestnut wood-based products can be considered of good quality when the MOEd is not lower than 7200 MPa. However, in C15 and C50, we recorded MOEd values lower than this threshold value, for DBH higher than 18 cm (7000–7100 MPa) and 45 cm (5587–7100 MPa), respectively. Therefore, almost all large poles (assortments of the greatest size) attainable in C15 and most of beams and boards (assortments with considerable size) obtainable in C50 did not have the minimum quality requirements for being classified as adequate. Quality wood could, however, be produced at relatively high stand age and DBH, when chestnut coppices grow in good site conditions and with appropriate silvicultural treatments (Manetti et al. 2020).
These results demonstrate that the advanced shoot ages, but also the high growth rates of chestnut coppices, negatively affected the wood quality. For this reason, the dynamics of stem radial increments might induce the production of a less-stiff mature wood, resulting in a significant loss of wood quality. Romagnoli et al. (2014) obtained similar results and observed, in coppice stands with age higher than 25–30 years, a decrease in the mechanical performance of chestnut wood near the threshold DBH of 35 cm. Therefore, to maximize and balance wood quality and stand productivity, coppicing in this Mediterranean context should be rethought in terms of strengths and weaknesses of the system, considering not only the shoot age, but also the shoot DBH at harvest (Genet et al. 2013), as well as shoot height and basal area (Marini et al. 2021).
We observed that, in the present conditions, when the rotation cycle ranged between 25 and 30 years, wood-based products of high quality could be obtained, as well as a variety of assortments. Nevertheless, 20–30-year-old chestnut stands, growing on favourable sites and properly managed, could be considered young and still have high growth rates (Cutini 2001; Conedera et al. 2004; Manetti et al. 2009). Similarly, we observed MAI equal to 13.3 m3 ha−1 year−1 at 26 years (year of culmination). Amorini and Manetti (1997) found that wood of good quality could be produced with rotation periods ranging between 30 and 50 years and with 2–4 medium-intensity thinning operations. It must be pointed out that, in many areas of Italy, common rotation periods range between 10 and 15 years (Ciancio et al. 2004). Such short rotation periods appear to be inadequate to ensure chestnut wood of good quality (Manetti et al. 2006). Indeed, only assortments of small size and poor quality were obtained in the present study, when short rotation periods were considered. Nowadays, commercial operators and the timber market in general often require assortments of good wood quality, obtainable from the chestnut coppices by lengthening to some extent the rotation period currently adopted in most chestnut coppices (Angelini et al. 2013; Mattioli et al. 2016; Manetti et al. 2017). Extended rotation periods (in the range of 30–50 years) would also positively affect the provisioning of ecosystem services related to environmental issues (Gondard and Romane 2005; Gondard et al. 2006). However, the lengthening of rotation periods without thinning might cause considerable competition stress, irregular radial growth and, consequently, ring shake risk.
Thinning would allow a greater average DBH of shoots to be obtained at earlier growth stages, with consequent differentiation of assortments (Mattioli et al. 2016). However, many studies have shown that marked increments in stem diameter after intensive thinning, especially if carried out late, might induce less-stiff mature wood, resulting in a significant loss of wood quality at high DBH (Zhang 1995; Ikeda and Kino 2000; Wang et al. 2003; Štefancík et al. 2018). Marini et al. (2021) found a negative correlation between wood density and dominant height and diametric growth of chestnut coppice stands, and this was associated to tree ring width. Should stand density, i.e., the number of shoots per ha, be a factor affecting positively wood density and the related mechanical properties, the application of low to moderate thinning might favour the formation of wood-based products of good quality in these chestnut coppices. However, caution is needed to avoid overextrapolation of these results. Indeed, stand can be of such poor quality that the obtained wood products have no commercial value. Therefore, particular care should be considered when thinning is planned and executed, avoiding both strong intensity and late thinning, and monitoring wood quality. By modifying competition (the number of shoots per ha) and, thus, shoot diameter growth and stand basal area, thinning might increase the risk of ring shake (Fonti et al. 2002; Becagli et al. 2006; Romagnoli et al. 2014). All these negative effects would limit the use of chestnut wood for the most valuable assortments, i.e., those usable as structural elements (Fonti et al. 2002).
Conclusions
The chronosequence approach was proved useful to show the effect of varying rotation periods on wood quality of homogeneous (except for age) chestnut coppice stands. A moderately negative correlation between shoot age and wood quality was observed. We also documented that innovative and non-destructive methods might produce wood technological indicators, i.e., MOEd, which relate shoot age and DBH to wood quality. These indicators may serve as tools for monitoring wood quality over time and provide valuable information that can aid decision-making in forest management.
Effective predictors for assessing wood quality may support stakeholders to develop management strategies that require control over stand characteristics. Nowadays, in many regions of Italy, the rotation period for chestnut coppices lasts 15–18 years. As such, harvest may take place too early compared to the economically and ecologically optimum rotation length for balancing forest profitability and carbon sequestration. Therefore, lengthening the rotation period to 25–30 years would benefit both productivity and quality of wood as well as landscape conservation and climate matter in Mediterranean mountain systems.
References
Aide TM, Zimmerman JK, Pascarella JB, Rivera L, Marcano Vega H (2000) Forest regeneration in a chronosequence of tropical abandoned pastures: implications for restoration ecology. Restor Ecol 8:328–338
Amorini E, Manetti MC (1997) Le ‘fustaie da legno’ di castagno del Monte Amiata. Ann Ist Sper Selv 28:53–61
Angelini A, Mattioli W, Merlini P, Corona P, Portoghesi L (2013) Empirical modelling of chestnut coppice yield for Cimini and Vicani mountains (Central Italy). Ann Silvic Res 37: 7–12. https://doi.org/10.12899/ASR-749
Becagli C, Amorini E, Manetti MC (2006) Incidenza della cipollatura in popolamenti cedui di castagno da legno del Monte Amiata. Annali Dell’istituto Sperimentale per La Selvicoltura 33:245–256
Bernetti G (1980) Auxometria dei boschi cedui italiani. L’italia Forestale e Montana 1:1–24
Bourgeois C, Sevrin E, Lemaire J (2004) Le châtaignier: un arbre, un bois, 2nd, revised. Institut pour le dèveloppement forestier, Paris, p 347
Bowditch E, Santopuoli G, Binder F, del Río M, La Porta N, Kluvankova T, Lesinski J, Motta R, Maciej P, Panzacchi P, Pretzsch H, Temperli C, Tonon G, Smith M, Velikova V, Weatherall A, Tognetti R (2020) What is Climate-Smart Forestry? A definition from a multinational collaborative process focused on mountain regions of Europe. Ecosyst Serv 43:101113
Buckley GP (1992) Ecology and management of coppice woodlands. Chapman & Hall, London
Cabrera BM, Ochoa (1997) Tablas de producción de castaño (Castanea sativa Mill.) tratado en monte bajo, en Asturias. In Puertas F, M Rivas M eds. II Congreso Forestal Español-Irati 97. Pamplona, Spain, pp 131–136
Ciancio O, Garfì V, Iovino F, Menguzzato G, Nicolaci A (2004) I cedui di castagno in Calabria: caratteristiche colturali, produttività e assortimenti ritraibili. L’italia Forestale e Montana 59:1–14
Clutter JL, Fortson JC, Pienaar LV, Brister GH, Bailey RL (1983) Timber management: a quantitative approach. John Wiley and Sons, Inc., New York, p 333
Conedera M, Manetti MC, Giudici F, Amorini E (2004) Distribution and economic potential of the Sweet Chestnut (Castanea sativa Mill.) in Europe. Ecol Mediterr 30:179–193
Conedera M, Tinner W, Krebs P, de Rigo D, Caudullo G (2016) Castanea sativa in Europe: distribution, habitat, usage and threats. In: San-Miguel-Ayanz J, de Rigo D, Caudullo G, Houston Durrant T, Mauri A (Eds.), European Atlas of Forest Tree Species. Publ. Off. EU, Luxembourg, pp. e0125e0+
Cutini A (2001) New management options in chestnut coppices: an evaluation on ecological basis. For Ecol Manag 141:165–174
Cutini A, Ferretti M, Bertini G, Brunialti G, Bagella S, Chianucci F, Fabbio G, Fratini R, Riccioli F, Caddeo C, Calderisi M, Ciucchi B, Corradini S, Cristofolini F, Cristofori A, Di Salvatore U, Ferrara C, Landi FL, , Marchino L, Patteri G, Piovosi M, Roggero PP, Seddaiu G, Gottardini E (2021) Testing an expanded set of sustainable forest management indicators in Mediterranean coppice area. Ecol Indic 130:108040. https://doi.org/10.1016/j.ecolind.2021.108040
Detter A, Cowell C, McKeown L, Howard P (2008) Evaluation of current rigging and dismantling practices used in arboriculture. Research Report RR668. Health and Safety Executive (HSE) and Forestry Commission (FC), UK, 370.
Divos F (2010) Acoustic tools for seedling, tree and log selection. In: Proceedings of the final conference of COST Action E53 the future of quality control for wood and wood products, Edinburgh, UK, 4–7 May 2010; p. 5.
Elorrieta J (1949) El castaño en España. Instituto Forestal de Investigaciones Agrarias (I.F.I.E.). Número 48. Madrid, España. Ministerio de Agricultura. 303 p.
Esteban E, Spinelli R, Aminti G, Laina RL, Vicens I (2018) Productivity, efficiency and environmental effects of whole-tree harvesting in spanish coppice stands using a drive-to-tree disc Saw Feller-Buncher. Croat J for Eng 39:163–172
Everard J, Christie JM (1995) Sweet chestnut: silviculture, timber quality and yield in the Forest of Dean. Forestry 68: 133–144.Fabbio G (2016) Coppice forests, or the changeable aspect of things, a review. Ann Silvicult Res 40:108–132
Fabbio G (2016) Coppice forests, or the changeable aspect of things, a review. Ann Silvicult Res 40(29): 108–132
FAO (2014) World reference base for soil resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps. World Soil Resources Reports No. 106; Roma, Italy.
Fonti P, Macchioni N, Thibaut B (2002) Ring shake in chestnut (Castanea sativa Mill.): state of the art. Ann For Sci 59:129–140. https://doi.org/10.1051/forest:2002007
Genet A, Auty D, Achim A, Bernier M, Pothier D, Cogliastro A (2013) Consequences of faster growth for wood density in northern red oak (Quercus rubra Liebl.). Forestry 86:99–110
Giannini R, Maltoni A, Mariotti B, Paffetti D, Tani A, Travaglini D (2014) Valorizzazione della produzione legnosa dei boschi di castagno. L’italia Forestale e Montana 69:307–317
Gondard H, Romane F (2005) Long-term evolution of understorey plant species composition after logging in chestnut coppice stands (Cevennes Mountain, Southern France). Ann Sci For 62:333–342
Gondard H, Romane F, Santa Regina I, Leonardi S (2006) Forest management and plant species diversity in chestnut stands of three Mediterranean areas. Biodivers Conserv 15:1129–1142
Greco S, Infusino M, Ienco A, Scalercio S (2018) How different management regimes of chestnut forests affect diversity and abundance of moth communities? Ann Silvic Res 42:59–67. https://doi.org/10.12899/asr-1503
Guntekin G, Emiroglu ZG, Yolmaz T (2013) Prediction of bending properties for Turkish red pine (Pinus brutia Ten.) Lumber using stress wave method. BioResources 8:231–237
Hedde M, Aubert M, Decaëns T, Bureau F (2008) Dynamics of soil carbon in a beechwood chronosequence forest. For Ecol Manag 255:193–202
Hédl R, Kopecky M, Komàrek J (2010) Half a century of succession in a temperate oakwood: from speciesrich community to mesic forest. Divers Distrib 16:267–276
Ikeda K, Kino N (2000) Quality evaluation of standing trees by a stress-wave propagation method and its application I. Seasonal changes of moisture contents of sugi standing trees and evaluation with stress-wave propagation velocity. Mokuzai Gakkaishi 46:181–188
INFC (2005) Le stime di superficie 2005. Inventario Nazionale delle Foreste e dei Serbatoi Forestali di Carbonio. MiPAF, Corpo Forestale dello Stato, CRA-ISAFA, Trento.
Johnson EA, Miyanishi K (2008) Testing the assumptions of chronosequences in succession. Ecol Lett 11:419–431
Kelly C, Ferrara A, Wilson GA, Ripullone F, Nolè A, Harmer N, Salvatic L (2015) Community resilience and land degradation in forest and shrubland socio-ecological systems: evidence from Gorgoglione, Basilicata, Italy. Land Use Policy 46:11–20
Kneifl MM, Kadavý J, Knott R, Adamec Z, Drápela K (2015) An inventory of tree and stand growth empirical modelling approaches with potential application in coppice forestry (A review). Acta Universitatis Agriculturae Et Silviculturae Mendelianae Brunensis 63:1789–1801
Lanuv (Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen), (2007) Niederwälder in Nordrhein-Westfalen. Galunder-Verlag, Nümbrecht-Elsenroth
Lemaire J (2008) Estimer la potentialité de son taillis de châtaignier et adapter les éclaircies. Forêt Enterprise 179:14–17
Manetti MC, Amorini E, Becagli C (2006) New silvicultural models to improve functionality of chestnut stands. Adv Hortic Sci 1:65–69
Manetti MC, Amorini E, Becagli C, Pelleri F, Pividori M, Schleppi P, Zingg A, Conedera M (2009) Quality wood production from chestnut (Castanea sativa Mill.) coppice forests - Comparison between different silvicultural approaches. In: Proceeding of the “1st European Congress on Chestnut” (Bounous G, Becarro G eds). Acta Horticulturae 866: 683–692.
Manetti MC, Becagli C, Sansone D, Pelleri F (2016) Tree-oriented silviculture: a new approach for coppice stands. iForest 9: 791–800
Manetti MC, Becagli C, Carbone F, Corona P, Giannini T, Pelleri F, Romano R (2017) Linee guida per la selvicoltura dei cedui di castagno. Rete Rurale Nazionale, Consiglio per la ricerca in agricoltura e l’analisi dell’economia agraria, Roma, ISBN: 9788899595579. 50 p.
Manetti MC, Marcolin E, Pividori M, Zanuttini R, Conedera M (2020) Coppice woodlands and chestnut wood technology. In: Beccaro G, Alma A, Bounous G, Gomes-Laranjo J (eds), The chestnut handbook. Crop and forest management, pp. 275–295. https://doi.org/10.1201/9780429445606-10.
Marcolin E, Manetti MC, Pelleri F, Conedera M, Pezzatti GB, Lingua E, Pividori M (2020) Seed regeneration of sweet chestnut (Castanea sativa Miller) under different coppicing approaches. For Ecol Manag 472:118273
Marini F, Manetti MC, Corona P, Portoghesi L, Viniguerra V, Tamantini S, Kuzminsky E, Zikeli F, Romagnoli M (2021) Influence of forest stand characteristics on physical, mechanical properties and chemistry of chestnut wood. Sci Rep 11:1549. https://doi.org/10.1038/s41598-020-80558-w
Marziliano PA, Lafortezza R, Colangelo G, Davies C, Sanesi G (2013) Structural diversity and height growth models in urban forest plantations: a case-study in northern Italy. Urban for Urban Green 12:246–254
Marziliano PA, Tognetti R, Lombardi F (2019) Is tree age or tree size reducing height increment in Abies alba Mill. at its southernmost distribution limit? Ann For Sci 76.
Mattioli W, Mancini LD, Portoghesi L, Corona P (2016) Biodiversity conservation and forest management: the case of the sweet chestnut coppice stands in Central Italy. Plant Biosyst 150:592–600. https://doi.org/10.1080/11263504.2015.1054448
Menéndez-Miguélez M, Canga E, Álvarez-Álvarez P, Majada J (2014) Stem taper functions for sweet chestnut (Castanea sativa Mill.) coppice stands in northwest Spain. Ann for Sci 71:761–770
Menéndez-Miguélez M, Álvarez-Álvarez P, Majada J, Canga E (2015) Effects of soil nutrients and environmental factors on site productivity in Castanea sativa Mill. coppice stands in NW Spain. New Forest 46:217–233
Menéndez-Miguélez M, Álvarez-Álvarez P, Majada J, Canga E (2016) Management tools for Castanea sativa coppice stands in northwestern Spain. Bosque 37:119–133
Moscatelli M, Pettenella D, Spinelli R (2007) Produttività e costi della lavorazione meccanizzata dei cedui di Castagno in ambiente appenninico. Forest@ 4, 51–62.
Parisi F, Lombardi F, Marziliano PA, Russo D, De Cristofaro A, Marchetti M, Tognetti R (2020) Diversity of saproxylic beetle communities in chestnut agroforestry systems. iForest 13: 456–465. https://doi.org/10.3832/ifor3478-013 [online 2020–10–07]
Pastorella F, Giacovelli G, Maesano M, Paletto A, Vivona S, Veltri A, Pellicone G, Scarascia Mugnozza G (2016) Social perception of forest multifunctionality in southern Italy: the case of Calabria Region. J For Sci 62:366–379
Patricio MS, Monteiro ML, Nunes LF, Mesquita S, Beito S, Casado J, Guerra H (2005) Management models evaluation of a Castanea sativa coppice in the northeast of Portugal. In: Proceedings of the Third International Chestnut (Ed.: Abreu CG, Rosa E, Monteiro AA). Acta Horticulturae 693: 721–725.
Pawson SM, Brockerhoff EG, Didham RK (2009) Native forest generalists dominate carabid assemblages along a stand-age chronosequence in anexotic Pinus radiata plantation. For Ecol Manag 258S:S108–S116
Pickett STA (1989) Space-for-time substitutions as an alternative to longterm studies. In: Likens GE (ed) Longterm studies in ecology. Springer, New York.
Pretzsch H (2009) Forest dynamics, growth and yield. Springer Verlag, Berlin
R Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. [online] URL: http://www.r-project.org
Romagnoli M, Cavalli D, Spina S (2014) Wood quality of chestnut: relationships between ring width, specific gravity and physical and mechanical properties. Bioresources 9:1132–1147. https://doi.org/10.15376/biores.9.1.1132-1147
Russo D, Marziliano PA, Macri G, Proto AR, Zimbalatti G, Lombardi F (2019) Does thinning intensity affect wood quality? An analysis of Calabrian Pine in Southern Italy using a non-destructive acoustic method. Forests 30:1–16
Russo D, Marziliano PA, Macrì G, Zimbalatti G, Tognetti R, Lombardi F (2020) Tree growth and wood quality in pure vs. mixed-species stands of European beech and Calabrian pine in Mediterranean mountain forests. Forests 11: 6. https://doi.org/10.3390/f11010006.
Rydberg D (2000) Initial sprouting, growth and mortality of European aspen and birch after selective coppicing in central Sweden. For Ecol Manag 130:27–35
Salisbury E (1952) Downs and Dunes: Their Plant Life and Environment. G. Bell & Sons Ltd., London, UK, p 1952
Scarascia-Mugnozza G, Oswald H, Piussi P, Radoglou K (2000) Forests of the Mediterranean region: gaps in knowledge and research needs. For Ecol Manag 132:97–109
Scherzinger W (1996) Naturschutz im Wald: Qualitätsziele einer dynamischen Waldentwicklung. Praktischer Naturschutz. — Stuttgart (Verlag Eugen Ulmer). 447 S., 51 Farbabb., 119 s/w-Abb., 36 Tab. ISBN 3-8001-3356-3.
Skovsgaard JP, Vanclay JK (2013) Forest site productivity: a review of spatial and temporal variability in natural site conditions. Forestry 86:305–315. https://doi.org/10.1093/forestry/cpt010
Štefancík I, Vacek Z, Sharma RP, Vacek S, Rösslová M (2018) Effect of thinning regimes on growth and development of crop trees in Fagus sylvatica stands of Central Europe over fifty years. Dendrobiology 79:141–155
Tabacchi G, Di Cosmo L, Gasparini P (2011) Aboveground tree volume and phytomass prediction equations for forest species in Italy. Eur J For Res 130:911–934
Teder M, Pilt K, Miljan M, Lainurm M, Kruuda R (2011) Overview of some non-destructive methods for in-situ assessment of structural timber. In Proceedings of the 3rd International Conference Civil Engineering, Jelgava, Latvia, 12–13 May 2011.
Vanclay JK (1994) Modelling forest growth and yield. CAB International, Wallingford
Vanninen P, Ylitalo H, Sievanen R, Makela A (1996) Effect of age and site quality on the distribution of biomass in Scots pine (Pinus sylvestris L.). Trees 10:231–238
Von Gadow K, Hui G (1999) Modelling forest development. Kluwer, Dordrecht
Walker et al (2010) The use of chronosequences in studies of ecological succession and soil development. J Ecol 98:725–736
Wang X, Ross RJ, Punches J, Barbour RJ, Forsman JW, Erickson JR (2003) Evaluation of small-diameter timber for valued-added manufacturing—a stress wave approach. In Proceedings of the II International Precision Forestry Symposium; Institute of Forest Resources, University of Washington: Seattle, WA, USA, pp. 91–96.
Weiskittel AR, Hann DW, Kershaw JA, Vanclay JK (2011) Forest growth and yield modeling. Wiley, Chichester
Wessels CB, Malan FS, Rypstra T (2011) A review of measurement methods used on standing trees for the prediction of some mechanical properties of timber. Eur J For Res 130:881–893
Zhang SY (1995) Effect of growth rate on wood specific gravity and selected mechanical properties in individual species from distinct wood categories. Wood Sci Technol 29:451–465
Zlatanov T, Schleppi P, Velichkov I, Hinkov G, Georgieva M, Eggertsson O, Zlatanova M, Harald V (2013) Structural diversity of abandoned chestnut (Castanea sativa Mill.) dominated forests: implications for forest management. For Ecol Manag 291:326–335. https://doi.org/10.1016/j.foreco.2012.11.015
Zuur AF, Ieno EN, Elphick CS (2010) A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol 1:3–14
Funding
Open access funding provided by Università degli Studi Mediterranea di Reggio Calabria within the CRUI-CARE Agreement. This work was supported by the Mediterranean University of Reggio Calabria.
Author information
Authors and Affiliations
Contributions
PAM and FL designed the study and analysed the data. MM and AL collected the data in the field. PAM, RT, and FL wrote the manuscript. All authors discussed the results and reviewed the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Competing interests
The authors declare no competing interests.
Additional information
Communicated by Miren del Río.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Marziliano, P.A., Tognetti, R., Mercuri, M. et al. Balancing stand productivity and wood quality in chestnut coppices using chronosequence approach and productivity model. Eur J Forest Res 141, 1059–1072 (2022). https://doi.org/10.1007/s10342-022-01488-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-022-01488-y