
Abstract
Background
Cytarabine (Ara-C) is an anticancer drug, which is considered as the mainstay in the treatment of hematological malignancies, known to cause various teratogenic effects. Alpha-lipoic acid (ALA) is a natural antioxidant and its supplementation proved to improve pregnancy outcomes in several pathological conditions. We aimed at exploring the benefits of maternal supplementation of ALA against in-utero Ara-C exposure-induced testicular toxicity in rat fetuses.
Methods
Pregnant rats (dams) received normal saline (control group), ALA 200 mg/kg (ALA group), Ara-C 12.5 mg/kg (Ara-C 12.5 group), Ara-C 25 mg/kg (Ara-C 25 group), and Ara-C 25 mg/kg + ALA 200 mg/kg (protection group) from gestational day (GD)8 to GD21. Ara-C and ALA were administered via the intraperitoneal and oral routes, respectively. The day of parturition was considered as postnatal day (PND)1. On PND1, all the live male pups were collected. The maternal parameters evaluated include (a) food intake, (b) bodyweight, and (c) oxidative stress (OS) markers. The fetal parameters evaluated include (a) bodyweight, (b) anogenital distances (AGD), (c) testicular weight (d) testicular testosterone levels (e) testicular histopathology, and (f) morphometrical parameters.
Results
A significant and dose-dependent decrease in maternal food intake, weight gain, and an increase in oxidative stress (OS) were observed in the pregnant rats of the Ara-C groups as compared to pregnant rats of the control group. Further, a significant and dose-dependent (a) reduction in bodyweight, AGD, testicular weight, and testosterone levels, (b) increase in OS, and (c) structural and morphometrical anomalies in fetal testes were observed in fetuses of Ara-C groups as compared to fetuses of the control rats. These deleterious effects observed in the Ara-C groups were found to be diminished in the pregnant rats and fetuses of the Protection group as compared to the pregnant rats and fetuses of the Ara-C 25 group.
Conclusions
From the results of this study, we conclude that the maternal supplementation of ALA may ameliorate the Ara-C exposure-induced impairment in prenatal development and function of the testes in the rat fetuses. However, future experimental and clinical studies are warranted to explore the possible mechanisms involved in the protection offered by maternal supplementation of ALA against Ara-C induced testicular toxicity.
Graphical Abstract

Similar content being viewed by others


1 Background
Cancer in pregnancy was considered infrequent as it was affecting 1 in 1000 pregnancies. However, several recent epidemiological studies emphasize the fact that the incidence of cancer in pregnancy is raising its bars in the recent years [1]. Breast, cervical and hematological malignancies are reported as the most frequent cancers during pregnancy [2]. Furthermore, among all the hematological malignancies, acute myeloid leukemia (AML) is considered as the most prevailing [3]. Treating malignancies during the period of pregnancy with chemotherapy is very challenging, as the treating physicians must consider the risk to both mother and fetuses [4]. Prenatal exposure to most of the chemotherapeutic agents leads to impaired prenatal and postnatal development, and function of various systems resulting in teratogenicity [5]. Anomalies in the development of the fetal reproductive system are reported to cause impairment in fertility and the quality of life during adult age [6].
Cytosine arabinoside (Ara-C, Cytarabine, 1-b-D-arabinofuranosylcytosine) is an anti-metabolite (deoxycytidine analog) anticancer drug serving as cornerstone therapy in AML treatment for the last four decades [7, 8]. Ara-C has been clinically used as an anticancer and antiviral drug. Ara-C has a similar structure with deoxycytidine but the presence of arabinose sugar instead of ribose sugar makes Ara-C different from deoxycytidine [9]. During the S-phase of cell cycle, Ara-C triphosphate metabolite (Ara-CTP) competitively inhibits the incorporation of deoxycytidine triphosphate by DNA polymerase into the growing DNA chain and thereby curtailing DNA synthesis during replication and repair [10,11,12]. Ara-C administration is found to elevate the oxidative stress (OS), DNA damage, restrict of cell proliferation, and promotes the apoptosis which contributes to its cytotoxic action [12,13,14]. Despite its therapeutic avenues, Ara-C was reported to cause various side effects like genotoxicity, bone marrow toxicity, neurotoxicity, gastrointestinal toxicity, testicular toxicity, and teratogenicity [15, 16]. The men treated for leukemia in childhood reported having impaired gonadal function [17]. Pre-pubertal Ara-C administration resulted in impaired postnatal testicular development, and spermatogenesis [12]. However, there were no studies that explored and characterized the outcomes of prenatal Ara-C exposure on fetal steroidogenesis, testicular development during embryonic period. Despite its various side effects, Ara-C is being used as a cornerstone therapy in the management of several hematological malignancies. Hence, there is a great need for developing strategies that offer protection against developmental and functional aberrations caused by Ara-C in the male reproductive system of rat fetuses.
Alpha-lipoic acid (1,2-dithiolane-3-pentanoic acid, ALA), a tissue component, acts as a biological antioxidant. ALA is a natural antioxidant which plays an imperative role in several biological pathways in plants, animals, and humans [18]. Furthermore, ALA is well known as a universal antioxidant as it offers antioxidant effects in both lipophilic and lipophobic biological environments [19, 20]. ALA is also known as ‘double antioxidant’ as both ALA and its metabolite, dihydro alpha-lipoic acid, holds excellent antioxidant activity. ALA plays an imperative role in several metabolic pathways by acting as a modulator, co-factor for various enzymes, a metal chelator, antioxidant, rejuvenator of other antioxidants [11]. Further, copious experimental and clinical data suggest ALA’s therapeutic potential in offering protection against several toxic pathological conditions induced by maternal diseases, chemical or toxicant exposures [18, 21,22,23]. In our previous study, we explored ALA’s protective effect against Ara-C-induced teratogenic effects and impaired skeletal system development in rat fetuses [11]. The maternal supplementation of ALA was found to offer protection against maternal nicotine- [24], dioxin- [25, 26], and carbimazole- [27] induced testicular toxicity in rat offspring. Further, the therapeutic potential of ALA in ameliorating testicular toxicity induced by chemical-, toxicant- or drug- exposure was proved in several studies.
In recent years, the use of the natural products as dietary components for the treatment of several diseases and for improving wellbeing is receiving an increasing attention globally [28]. Therefore, in view of the above considerations, we carried out this research work to explore the effects of maternal Ara-C exposure on the development of the fetal rat testes and the possible protective effects of ALA against Ara-C induced developmental anomalies in fetal rat testes.
2 Methods
2.1 Study design
The Institutional Animal Ethics Committee has approved the protocol (Approval Number: MLRIP/IAEC/2018/02) and all procedures were conducted as per the guidelines of the committee for the purpose of control and supervision of experimentation on animals. The animals used in this study were kept in an animal house that is maintained in a controlled environmental condition (22 ± 2 °C temperature, 50 ± 10% humidity, and 12 h light/12 h dark cycle). During the study period, all the animals were provided with standard laboratory animal feed and water. These animals were allowed to acclimatize to the experimental conditions for a weekdays prior to starting the experiments.
The female rats in estrous were allowed to mate with male rats in a ratio of 2 females and 1 male. On the next day morning, the female rats with a vaginal plug and/or traces of sperm in vaginal smears were considered pregnant [gestation day (GD) 0]. On GD7, the non-pregnant rats were separated from the dams. On GD8, 30 dams were randomly assigned to five groups and received normal saline (control group), ALA 200 mg/kg (ALA group), Ara-C 12.5 mg/kg (Ara-C 12.5 group), Ara-C 25 mg/kg (Ara-C 25 group), and Ara-C 25 mg/kg + ALA 200 mg/kg (Protection group) from GD8 to GD21 [11]. Ara-C and ALA were administered via the intraperitoneal and oral routes, respectively. During the study period, the food intake and bodyweights of each dam were recorded daily and a thorough examination for any clinical abnormalities, mortality, and morbidity was performed at least once a day.
2.2 Sampling
Blood was collected from dams on GD21 using the retro-orbital bleeding method and the same was used for the estimation of OS markers. The day of parturition was considered as postnatal day (PND)1. On PND1, all the live male pups were collected, weighed, and anogenital distance (AGD) (the distance from the midpoint of the anus to the genitalia) [29] were recorded. Further, fetal testes were carefully collected and used for histopathological, hormonal, and morphometrical evaluation.
2.3 Estimation of oxidative stress (OS) markers
One testis from each fetus were pooled up, homogenized, centrifuged (10 min at 700 g) and the resultant supernatant solution was isolated. The supernatant solution of fetal testes and plasma samples of dams were used for the evaluation of OS markers. The OS parameters evaluated in this study include malondialdehyde (MDA), catalase (CAT), superoxide dismutase (SOD), glutathione (GSH), glutathione peroxidase (GSH-Px), and protein content that were measured as per the methods described by Ohkawa et al. [30], Sinha [31], Kono [32], Ellman [33], Rotruck et al. [34], and Lowry et al. [35], respectively.
2.4 Estimation of testosterone levels
The fetal testosterone levels were determined using the supernatant obtained as per the instructions provided in the commercially available kits (Testosterone ELISA CE) obtained from Biochrome Scientific, Hyderabad. [36]
2.5 Histopathology and morphometry
The second testes of each fetus were fixed in 10% neutral buffered formalin (NBF). These formalin-fixed testes have undergone the below steps in a sequence: (1) soaking in phosphate buffer saline, (2) dehydration using a series of increasing concentrations (70%, 80%, 90%, and 100%) of ethanol solutions, (3) soaking in the xylene (to clear the alcohol), (4) embedding in paraffin, (5) sectioning (5 μm), (6) mounting on glass slides, (7) drying overnight, (8) deparaffinization, (9) rehydration, and (10) staining with Hematoxylin & Eosin (ES). The stained tissues on the slides were examined for any histological and morphometrical alterations using a microscope that was connected to a CCD camera and imaging software. The mean diameter (mean of vertical and horizontal diameters) and area of seminiferous tubules (STs) were calculated using a total of 90 STs. For tubular density, a total of 20 random focuses (60 mm square area) per animal were used. Further, a total of 30 STs from each animal were analyzed to calculate number of multinucleated germ (MNG) cells [37].
2.6 Statistical analysis
The results were expressed as mean ± SEM for each group. Statistical differences between the groups were determined by one-way ANOVA followed by multiple comparisons with Tukey’s test using Sigma Stat 2.03 statistical software. The level of statistical significance was set at P < 0.05.
3 Results
3.1 Maternal toxicity
No external signs of toxicity, morbidity, and mortality were observed in dams of all the study groups. A statistically significant and dose-dependent decrease in weight gain (Fig. 1A–C) and food intake (Fig. 1E) was observed in dams of Ara-C groups in a dose-dependent manner as compared to the dams of the control group. Interestingly, the dams of the protection group showed a significant increase in food intake and weight gains as compared to the dams of the Ara-C 25 group. Further, similar trends were observed in all the groups when percentage weight gain was analyzed (Fig. 1D). A dose-dependent elevation in MDA levels and reduction in non-enzymatic (GSH) and enzymatic (CAT, SOD, and GPx) antioxidant levels were observed in the dams of Ara-C groups as compared to the dams of the control group (Fig. 2A–E). Interestingly, a dose-dependent reduction in MDA levels and elevation in non-enzymatic (GSH) and enzymatic (CAT, SOD, and GPx) antioxidant levels were observed in the dams of the protection group as compared to the dams of the Ara-C 25 group.

Effect of Ara-C and ALA on maternal A bodyweight, B weight gain, C weekly bodyweight gain, D %weight gain, and E food intake. All the values are expressed as mean ± SEM, (n = 6). ***P < 0.001 versus control, **P < 0.01 versus control, *P < 0.05 versus control; ###P < 0.001 versus Ara-C 25 mg/kg, ###P < 0.01 versus Ara-C 25 mg/kg, #P < 0.05 versus Ara-C 25 mg/kg; For food intake, β, P < 0.001 versus control, @P < 0.001 versus control); §P < 0.001 versus Ara-C 25 mg/kg. ALA alpha-lipoic acid, Ara-C cytarabine, GD gestation day, g grams

Effect of Ara-C and ALA on maternal oxidative stress markers, A MDA, B SOD, C GSH, D GSH-Px, and E Catalase. All the values are expressed as mean ± SEM, (n = 6). ***P < 0.001 versus control, **P < 0.01 versus control, *P < 0.05 versus control; ###P < 0.001 versus Ara-C 25 mg/kg. ALA alpha-lipoic acid, Ara-C cytarabine, GD gestation day, GSH glutathione, GSH-Px glutathione peroxidase, g grams, MDA malondialdehyde, SOD superoxide dismutase
3.2 Fetal toxicity
3.2.1 Effect on bodyweight, organ weights and anogenital distance
The fetuses of Ara-C groups demonstrated a significant reduction in the bodyweight (Fig. 3A), testicular weight (Fig. 3B), and AGD (Fig. 3C) in a dose-dependent manner as compared to the fetuses of the control group. Interestingly, the fetuses of the protection group demonstrated a clear increase in the bodyweight and testicular weight as compared to the fetuses of the Ara-C 25 group.

Effect of Ara-C and ALA on fetal A bodyweight, B testicular weight and C anogenital distance. All the values are expressed as mean ± SEM, (n = 10). ***P < 0.001 versus control, **P < 0.01 versus control, *P < 0.05 versus control; ### P < 0.001 versus Ara-C 25 mg/kg. ALA alpha-lipoic acid, Ara-C cytarabine
3.2.2 Oxidative stress parameters
The fetuses of the Ara-C groups demonstrated a significant and dose-dependent elevation in MDA levels as compared to the fetuses of the control group. The fetuses of protection group demonstrated a clear reduction in MDA levels as compared to the fetuses of the Ara-C 25 group (Fig. 4A–E). Further, a significant and dose-dependent reduction in GSH, SOD, GPx, and CAT levels was observed as compared to the fetuses of the control group. Interestingly, the fetuses of the protection group demonstrated a clear increase in GSH, SOD, GPx, and CAT levels as compared to the fetuses of the Ara-C 25 group (Fig. 4A–E).

Effect of Ara-C and ALA on fetal oxidative stress markers A MDA, B SOD, C GSH, D GSH-Px, and E Catalase. All the values are expressed as mean ± SEM, (n = 10). ***P < 0.001 versus control, **P < 0.01 versus control; ###P < 0.001 versus Ara-C 25 mg/kg. ALA, Alpha-lipoic acid, Ara-C cytarabine, GD gestation day, GSH glutathione, GSH-Px glutathione peroxidase, g grams, MDA malondialdehyde, SOD superoxide dismutase
3.2.3 Testosterone levels
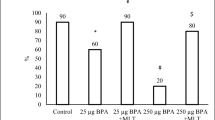
The fetuses of the Ara-C groups demonstrated a significant reduction in the testosterone levels in a dose-dependent manner as compared to the fetuses of the control group. Interestingly, the fetuses of the protection group demonstrated a clear increase in the testosterone levels as compared to the fetuses of the Ara-C 25 group (Fig. 5A).

Effect of Ara-C and ALA on A testosterone level, B density, C diameter, D multinucleated gonocytes, E vacuolization, and F number of cells of STs. All the values are expressed as mean ± SEM (n = 10). ***P < 0.001 versus control, ** P < 0.01 versus control, *P < 0.05 versus control; ###P < 0.001 versus Ara-C 25 mg/kg. ALA alpha-lipoic acid 200 mg/kg, Ara-C cytarabine, ST seminiferous tubule
3.3 Histopathology and morphometric analysis
The fetuses of the control group and ALA group were found to have a normal testicular architecture. However, the fetuses of Ara-C groups were found to have a distorted testicular architecture (Fig. 6). The histological anomalies observed in the testes of the fetuses of Ara-C groups include a reduction in the size of Leydig cells, number of supporting cells and an increase in vacuolization in STs and in the interstitial area (Fig. 5B–E). Interestingly, a significant increase in the size of Leydig cells and the number of supporting cells and a decrease in vacuolization in STs and interstitial area were witnessed in the fetuses of protection group as compared to the fetuses of the Ara-C 25 group. Further, a clear reduction in diameter of the STs and number of cells in STs, increase in density of STs, and degeneration of the seminiferous epithelium (DSE) were observed in the testes of the fetuses of Ara-C groups as compared to the fetuses of the control group (Fig. 5B–E). Interestingly, a clear increase in diameter of the STs and number of cells in STs, decrease in density of STs, and DSE were observed in the testes of the fetuses of the protection group as compared to the fetuses of Ara-C 25 group (Fig. 6A–E). No difference was observed in the incidence of MNGs among all study groups (Fig. 5D).

Effect of Ara-C and ALA on fetal testicular histology. Representative photomicrographs of testes of rat fetuses. White arrows indicate the testes of the rat fetus. Black arrows indicate the areas of vacuolization. ALA alpha-lipoic acid 200 mg/kg, Ara-C cytarabine
4 Discussion
The results of the study are clearly indicating the potential of ALA in alleviating pre-natal Ara-C exposure–induced impairment in the fetal testicular development and steroidogenesis.
In the present study, the administration of Ara-C during gestational period (GD8 to GD21) resulted in reduction of food intake and bodyweight gain and increase in OS. These results are in line with the results of our previous study [11] in which the exposure to Ara-C during the gestation period impaired the placental development as revealed by an increase in oxidative damage and reduction in placental weight. The OS and DNA damage were reported as key pathological damages implicated in Ara-C-induced cytotoxicity [12, 15, 38, 39]. Further, the prenatal exposure to Ara-C reported to impair cell proliferation and provoke apoptosis in the placenta (especially in the labyrinth zone) which eventually leads to impaired placental development [14]. Accumulating research evidence suggests that the oxidative damage impairs the maternal pathways that are crucial for the maintenance and success of the pregnancy [40, 41]. It is well known that the structural, nutritional, and biochemical support from the placenta is crucial for the development of organ systems during embryonic life and plays a critical role for postnatal development in the offspring. Further, the perturbations in the structural and functional capacity of the placenta reported to cause to developmental toxicity [37, 42, 43]. In line with the previous studies, a clear maternal toxicity was observed in this study which is clearly indicating the potential of Ara-C in causing developmental toxicity. Interestingly, ALA co-administration was found to reverse Ara-C-induced toxic effects in maternal rats. From these results, it is evident that ALA has the potential to ameliorate the maternal toxicity by Ara-C. Furthermore, several clinical and non-clinical studies have shown the ALA’s potential in offering protection against various pregnancy-associated ailments and chemical-induced developmental toxicity [25, 44, 45].
In this study, exposure to Ara-C in embryonic period during which male reproductive system development occurs resulted in clear oxidative imbalance in the testes of the fetuses. This claim was based on a clear imbalance between oxidative free radicals and antioxidant levels as evidenced by a significant elevation in MDA, and reduction in enzymatic (SOD, GSH-Px, and CAT levels) and non-enzymatic (GSH) antioxidants in fetal testes prenatally exposed to Ara-C. In our previous study, a clear oxidative damage in rat fetuses was observed after having Ara-C exposure during the gestation period as supported by elevation in oxidative stress and a decline in the antioxidant defenses [11]. These effects were reversed with co-treatment of ALA indicating its capability to safeguard the testes from Ara-C-induced OS warranting a clear beneficial effect against Ara-C induced toxicity in rat fetal testes. Further, a previous study reported that postnatal exposure to Ara-C during pre-pubertal age can cause impairment in the postnatal testicular development, spermatogenesis, and its function at puberty in rats [12]. Ara-C exposure during pre-pubertal age resulted in a massive elevation in DNA damage, apoptosis, and extreme loss of spermatogenic cells. A clear testicular atrophy and impairment in testicular function in terms of oligospermia was observed [12]. In our current study, Ara-C exposure during the embryonic period resulted in a distorted architecture and impaired prenatal development in testes of the fetus as evidenced by the reduced size of Leydig cells, clustering of Leydig cells, reduced number of cells, and vacuolization in STs and interstitial area. These histological and developmental anomalies are clearly indicating the Ara-C’s potential in impairing prenatal testicular development in fetuses. These negative claims were further supported by results of the morphometric analysis which revealed a clear reduction in diameter and number of cells in ST and increased tubular density in testes of fetuses. Numerous literature is emphasizing that fetuses are more vulnerable to OS [40]. Further, by virtue of its structural and functional uniqueness, testicular tissue is more prone to oxidative damage [46]. Therefore, the negative effects observed in testes of the fetuses of rats that had Ara-C exposure may be ascribed to Ara-C’s potential to induce OS, DNA damage, and cytotoxicity and inhibit cell proliferation [12]. It is well reported that the testicular development during fetal life is considered to have a decisive role in the post-natal development and function of the male reproductive system (van den [47]. Therefore, Ara-C exposure during embryonic period may impair the post-natal development and function of the testes.
We also found that prenatal Ara-C exposure causes deleterious effects on fetal Leydig cells and Sertoli cells that work together in a highly coordinated manner to produce testosterone. These deleterious effects of Ara-C are further supported by a significant reduction in fetal bodyweight, testosterone levels and testicular weight. The reduction in the bodyweight and testes is considered as a manifestation of reduction in androgen levels as testosterone was found to play a central role in the development of male reproductive organs and masculinization thereby maintaining the bodyweight [48, 49]. The dose-dependent decline in testosterone levels could be a reason at least in part for reduced bodyweight and testicular weight in rat fetuses that had Ara-C in-utero exposure.
Interestingly, the maternal supplementation of ALA showed to improve all the anomalies in testes of the rat fetuses induced by Ara-C’s in-utero exposure including the bodyweight and testicular weight. This is clearly indicating the beneficial effects of maternal supplementation of ALA while taking Ara-C during pregnancy. It is reported that due to its unique capability to serve as an antioxidant in both hydrophilic and hydrophobic environments (universal antioxidant) and serve as a redox couple, ALA exhibits pleiotropic effects thereby offering beneficial effects against anomalies developed due to pathological conditions and drug-exposure. [21, 50]. Several reports suggested that ALA’s beneficial effects are by virtue of its capability to regenerate endogenous antioxidants, serve as a co-factor for several enzymes and curb a wide variety of reactive oxygen species [50, 51]. Further, copious literature is quoting that ALA with a remarkable safety profile has great potential to improve the pregnancy outcomes in conditions like subchorionic hematomas, miscarriage, and preterm delivery [18, 22, 23, 52,53,54] and chemical- or disease-induced developmental toxicity [24, 45, 55]. In corroboration to the previous studies, in our current study, ALA demonstrated protective effects against the developmental anomalies in fetal testes induced by Ara-C exposure during the embryonic period. ALA might have inhibited or at least in part countered all the mechanisms through which Ara-C caused its deleterious effects in fetal testes like OS, curtailing cell proliferation, induction of DNA damage, and cellular apoptosis. In recent years, supplementation of natural products or antioxidants during the gestational period is being appraised as it is improving the pregnancy disorders and curtailing chemical- or stress-induced maternal and fetal toxicity [56]. Therefore, it can be postulated that maternal administration of ALA could prevent the Ara-C-induced anomalies in prenatal developmental and functional aspects of testes and thereby postnatal development and function of the male reproductive system in rats. Though this study proved ALA’s potential in alleviating Ara-C-induced testicular toxicity in the rat fetuses, future studies must aim at exploring molecular mechanisms implicated in ALA’s protective effects.
5 Conclusions
In conclusion, the findings of the current study show that in-utero exposure to Ara-C impairs antioxidant reserve, testosterone production, and development of testes in rat fetuses in a dose-dependent manner. The maternal supplementation of ALA found to protection against the Ara-C-induced anomalies in the development and function of testes in rat fetuses.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- AGD:
-
Anogenital distances
- ALA:
-
Alpha-lipoic acid
- AML:
-
Acute myeloid leukemia
- ANOVA:
-
Analysis of variance
- Ara-C:
-
Cytarabine, 1-b-D-arabinofuranosylcytosine
- CAT:
-
Catalase
- DSE:
-
Degeneration of the seminiferous epithelium
- GD:
-
Gestational day
- GSH:
-
Glutathione
- GSH-Px:
-
Glutathione peroxidase
- MDA:
-
Malondialdehyde
- MNG:
-
Multinucleated germ
- MPW:
-
Masculinization programming window
- OS:
-
Oxidative stress
- PND:
-
Postnatal day
- SEM:
-
Standard error of the mean
- SOD:
-
Superoxide dismutase
- ST:
-
Seminiferous tubule
References
Botha MH, Rajaram S, Karunaratne K (2018) Cancer in pregnancy. Int J Gynecol Obstet 143:137–142. https://doi.org/10.1002/ijgo.12621
Abdalla N, Bizoń M, Piórkowski R, Stanirowski P, Cendrowski K, Sawicki W (2017) Does chemotherapy for gynecological malignancies during pregnancy cause fetal growth restriction? BioMed Res Int. https://doi.org/10.1155/2017/7543421
Milojkovic D, Apperley JF (2014) How I treat leukemia during pregnancy. Blood 123:974–984. https://doi.org/10.1182/blood-2013-08-283580
Eastwood-Wilshere N, Turner J, Oliveira N, Morton A (2019) Cancer in Pregnancy. Asia Pac J Clin Oncol 15:296–308. https://doi.org/10.1111/ajco.13235
Bavanilathamuthiah, Lakshmanan Y, Ratnaprabha P (2016) Developmental toxicity of arsenic and its underlying mechanisms in the early embryonic development. Res J Pharm Tech 9:340–344. https://doi.org/10.5958/0974-360X.2016.00060.3
Yang Y, Zhang Z, Zhang H, Hong K, Tang W, Zhao L, Lin H, Liu D, Mao J, Wu H, Jiang H (2017) Effects of maternal acrolein exposure during pregnancy on testicular testosterone production in fetal rats. Mol Med Rep 16:491–498. https://doi.org/10.3892/mmr.2017.6624
Liu J, Liu J, Zhao D, Ma N, Luan Y (2016) Highly enhanced leukemia therapy and oral bioavailability from a novel amphiphilic prodrug of cytarabine. RSC Adv 6:35991–35999. https://doi.org/10.1039/C6RA02051H
Reese ND, Schiller GJ (2013) High-dose cytarabine (HD araC) in the treatment of leukemias: a review. Curr Hematol Malig Rep 8:141–148. https://doi.org/10.1007/s11899-013-0156-3
Hamada A, Kawaguchi T, Nakano M (2002) Clinical pharmacokinetics of cytarabine formulations. Clin Pharmacokinet 41:705–718. https://doi.org/10.2165/00003088-200241100-00002
Faruqi A, Tadi P (2021) Cytarabine. In: StatPearls. StatPearls Publishing, Treasure Island
Namoju R, Chilaka NK (2021) Alpha-lipoic acid ameliorates cytarabine-induced developmental anomalies in rat fetus. Hum Exp Toxicol 40:851–868. https://doi.org/10.1177/0960327120975114
Namoju RC, Khan S, Patel RS, Shera FY, Trivedi PP, Kushwaha S, Jena GB (2014) Pre-pubertal exposure of cytarabine-induced testicular atrophy, impaired spermatogenesis and germ cell DNA damage in SD rats. Toxicol Mech Methods 24:703–712. https://doi.org/10.3109/15376516.2014.970679
Patel RS, Rachamalla M, Chary NR, Shera FY, Tikoo K, Jena G (2012) Cytarabine induced cerebellar neuronal damage in juvenile rat: correlating neurobehavioral performance with cellular and genetic alterations. Toxicology 293:41–52. https://doi.org/10.1016/j.tox.2011.12.005
Yamauchi H, Katayama K, Ueno M, Uetsuka K, Nakayama H, Doi K (2004) Involvement of p53 in 1-beta-D-arabinofuranosylcytosine-induced trophoblastic cell apoptosis and impaired proliferation in rat placenta. Biol Reprod 70:1762–1767. https://doi.org/10.1095/biolreprod.103.026252
Magina KN, Pregartner G, Zebisch A, Wölfler A, Neumeister P, Greinix HT, Berghold A, Sill H (2017) Cytarabine dose in the consolidation treatment of AML: a systematic review and meta-analysis. Blood 130:946–948. https://doi.org/10.1182/blood-2017-04-777722
Stentoft J (1990) The toxicity of cytarabine. Drug Saf 5:7–27. https://doi.org/10.2165/00002018-199005010-00003
Zaletel LZ, Bratanic N, Jereb B (2004) Gonadal function in patients treated for leukemia in childhood. Leuk Lymphoma 45:1797–1802. https://doi.org/10.1080/1042819042000219458
Parente E, Colannino G, Picconi O, Monastra G (2017) Safety of oral alpha-lipoic acid treatment in pregnant women: a retrospective observational study. Eur Rev Med Pharmacol Sci 21:4219–4227
Moura FA, de Andrade KQ, dos Santos JCF, Goulart MOF (2015) Lipoic acid: its antioxidant and anti-inflammatory role and clinical applications. Curr Top Med Chem 15:458–483. https://doi.org/10.2174/1568026615666150114161358
Park S, Karunakaran U, Jeoung NH, Jeon JH, Lee IK (2014) Physiological effect and therapeutic application of alpha lipoic acid. Curr Med Chem 21:3636–3645. https://doi.org/10.2174/0929867321666140706141806
Gomes MB, Negrato CA (2014) Alpha-lipoic acid as a pleiotropic compound with potential therapeutic use in diabetes and other chronic diseases. Diabetol Metab Syndr 6:80. https://doi.org/10.1186/1758-5996-6-80
Micili SC, Goker A, Kuscu K, Ergur BU, Fuso A (2019) α-Lipoic acid vaginal administration contrasts inflammation and preterm delivery in rats. Reprod Sci Thousand Oaks Calif 26:128–138. https://doi.org/10.1177/1933719118766266
Monastra G, De Grazia S, Cilaker Micili S, Goker A, Unfer V (2016) Immunomodulatory activities of alpha lipoic acid with a special focus on its efficacy in preventing miscarriage. Expert Opin Drug Deliv 13:1695–1708. https://doi.org/10.1080/17425247.2016.1200556
Anto SK, Koyada N, Khan S, Jena G (2016) α-Lipoic acid attenuates transplacental nicotine-induced germ cell and oxidative DNA damage in adult mice. J Basic Clin Physiol Pharmacol 27:585–593. https://doi.org/10.1515/jbcpp-2015-0151
Koga T, Ishida T, Takeda T, Ishii Y, Uchi H, Tsukimori K, Yamamoto M, Himeno M, Furue M, Yamada H (2012) Restoration of dioxin-induced damage to fetal steroidogenesis and gonadotropin formation by maternal co-treatment with α-lipoic acid. PLoS ONE 7:e40322. https://doi.org/10.1371/journal.pone.0040322
Takeda T, Matsuo Y, Nishida K, Fujiki A, Hattori Y, Koga T, Ishii Y, Yamada H (2017) α-Lipoic acid potentially targets AMP-activated protein kinase and energy production in the fetal brain to ameliorate dioxin-produced attenuation in fetal steroidogenesis. J Toxicol Sci 42:13–23. https://doi.org/10.2131/jts.42.13
Prathima P, Venkaiah K, Pavani R, Daveedu T, Munikumar M, Gobinath M, Valli M, Sainath SB (2017) α-lipoic acid inhibits oxidative stress in testis and attenuates testicular toxicity in rats exposed to carbimazole during embryonic period. Toxicol Rep 4:373–381. https://doi.org/10.1016/j.toxrep.2017.06.009
Khan RA (2018) Natural products chemistry: the emerging trends and prospective goals. Saudi Pharm J SPJ Off Publ Saudi Pharm Soc 26:739–753. https://doi.org/10.1016/j.jsps.2018.02.015
Wu K, Li Y, Pan P, Li Z, Yu Y, Huang J, Ma F, Tian L, Fang Y, Wang Y, Lin H, Ge RS (2020) Gestational vinclozolin exposure suppresses fetal testis development in rats. Ecotoxicol Environ Saf 203:111053. https://doi.org/10.1016/j.ecoenv.2020.111053
Ohkawa H, Ohishi N, Yagi K (1979) Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem 95:351–358. https://doi.org/10.1016/0003-2697(79)90738-3
Sinha AK (1972) Colorimetric assay of catalase. Anal Biochem 47:389–394. https://doi.org/10.1016/0003-2697(72)90132-7
Kono Y (1978) Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch Biochem Biophys 186:189–195. https://doi.org/10.1016/0003-9861(78)90479-4
Ellman GL (1959) Tissue sulfhydryl groups. Arch Biochem Biophys 82:70–77. https://doi.org/10.1016/0003-9861(59)90090-6
Rotruck JT, Pope AL, Ganther HE, Swanson AB, Hafeman DG, Hoekstra WG (1973) Selenium: biochemical role as a component of glutathione peroxidase. Science 179:588–590. https://doi.org/10.1126/science.179.4073.588
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193:265–275
McCann D, Kirkish L (1985) Evaluation of free testosterone in serum. J Clin Immunoassay 8:234–236
Napso T, Hung YP, Davidge ST, Care AS, Sferruzzi-Perri AN (2019) Advanced maternal age compromises fetal growth and induces sex-specific changes in placental phenotype in rats. Sci Rep 9:16916. https://doi.org/10.1038/s41598-019-53199-x
Guzmán DC, Brizuela NO, Herrera MO, Olguín HJ, García EH, Peraza AV, Mejía GB (2016) Oleic acid protects against oxidative stress exacerbated by cytarabine and doxorubicin in rat brain. Anticancer Agents Med Chem 16:1491–1495. https://doi.org/10.2174/1871520615666160504093652
Hernández García E, Osnaya Brizuela N, Valenzuela Peraza A, Calderón Guzmán D, Ortiz Herrera M, Juárez Olguín H, Barragán Mejía G, Santamaría Del Ángel D, Rojas Ochoa A (2018) Biochemical and histological changes produced by sweeteners and cytarabine in the brain of young rats. Nutr Hosp 35:194–200. https://doi.org/10.20960/nh.1245
Laforgia N, Di Mauro A, Favia Guarnieri G, Varvara D, De Cosmo L, Panza R, Capozza M, Baldassarre ME, Resta N (2018) The role of oxidative stress in the pathomechanism of congenital malformations. Oxid Med Cell Longev. https://doi.org/10.1155/2018/7404082
Schoots MH, Gordijn SJ, Scherjon SA, van Goor H, Hillebrands JL (2018) Oxidative stress in placental pathology. Placenta 69:153–161. https://doi.org/10.1016/j.placenta.2018.03.003
Sankaran S, Kyle PM (2009) Aetiology and pathogenesis of IUGR. Best Pract Res Clin Obstet Gynaecol 23:765–777. https://doi.org/10.1016/j.bpobgyn.2009.05.003
Woods L, Perez-Garcia V, Hemberger M (2018) Regulation of placental development and its impact on fetal growth-new insights from mouse models. Front Endocrinol 9:570. https://doi.org/10.3389/fendo.2018.00570
Al Ghafli MHM, Padmanabhan R, Kataya HH, Berg B (2004) Effects of alpha-lipoic acid supplementation on maternal diabetes-induced growth retardation and congenital anomalies in rat fetuses. Mol Cell Biochem 261:123–135. https://doi.org/10.1023/b:mcbi.0000028747.92084.42
Padmanabhan R, Mohamed S, Singh S (2006) Beneficial effect of supplemental lipoic acid on diabetes-induced pregnancy loss in the mouse. Ann N Y Acad Sci 1084:118–131. https://doi.org/10.1196/annals.1372.015
Aitken RJ (2017) Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol Reprod Dev 84:1039–1052. https://doi.org/10.1002/mrd.22871
van den Driesche S, Kilcoyne KR, Wagner I, Rebourcet D, Boyle A, Mitchell R, McKinnell C, Macpherson S, Donat R, Shukla CJ, Jorgensen A, Meyts ERD, Skakkebaek NE, Sharpe RM (2017) Experimentally induced testicular dysgenesis syndrome originates in the masculinization programming window. JCI Insight 2:e91204. https://doi.org/10.1172/jci.insight.91204
Tyagi V, Scordo M, Yoon RS, Liporace FA, Greene LW (2017) Revisiting the role of testosterone: are we missing something? Rev Urol 19:16–24. https://doi.org/10.3909/riu0716
Sharpe RM (2020) Androgens and the masculinization programming window: human-rodent differences. Biochem Soc Trans 48:1725–1735. https://doi.org/10.1042/BST20200200
Salehi B, Berkay Yılmaz Y, Antika G, Boyunegmez Tumer T, Fawzi Mahomoodally M, Lobine D, Akram M, Riaz M, Capanoglu E, Sharopov F, Martins N, Cho WC, Sharifi-Rad J (2019) Insights on the use of α-lipoic acid for therapeutic purposes. Biomolecules 9:E356. https://doi.org/10.3390/biom9080356
Solmonson A, DeBerardinis RJ (2018) Lipoic acid metabolism and mitochondrial redox regulation. J Biol Chem 293:7522–7530. https://doi.org/10.1074/jbc.TM117.000259
Costantino M, Guaraldi C, Costantino D (2016) Resolution of subchorionic hematoma and symptoms of threatened miscarriage using vaginal alpha lipoic acid or progesterone: clinical evidences. Eur Rev Med Pharmacol Sci 20:1656–1663
Grandi G, Pignatti L, Ferrari F, Dante G, Neri I, Facchinetti F (2017) Vaginal alpha-lipoic acid shows an anti-inflammatory effect on the cervix, preventing its shortening after primary tocolysis. A pilot, randomized, placebo-controlled study. J Matern-Fetal Neonatal Med Off J Eur Assoc Perinat Med Fed Asia Ocean Perinat Soc Int Soc Perinat Obstet 30:2243–2249. https://doi.org/10.1080/14767058.2016.1245282
Porcaro G, Brillo E, Giardina I, Di Iorio R (2015) Alpha Lipoic Acid (ALA) effects on subchorionic hematoma: preliminary clinical results. Eur Rev Med Pharmacol Sci 19:3426–3432
Sugimura Y, Murase T, Kobayashi K, Oyama K, Hayasaka S, Kanou Y, Oiso Y, Murata Y (2009) Alpha-lipoic acid reduces congenital malformations in the offspring of diabetic mice. Diabetes Metab Res Rev 25:287–294. https://doi.org/10.1002/dmrr.947
Singh K, Bhori M, Kasu YA, Bhat G, Marar T (2018) Antioxidants as precision weapons in war against cancer chemotherapy induced toxicity—exploring the armoury of obscurity. Saudi Pharm J SPJ Off Publ Saudi Pharm Soc 26:177–190. https://doi.org/10.1016/j.jsps.2017.12.013
Acknowledgements
We would like to acknowledge Bhaskar Medical College, Hyderabad, and Bhaskar Pharmacy College, Hyderabad, for providing facilities and also Vivo Bio Tech Ltd, Pregnapur, Telangana, for the support in technological aspects.
Funding
The authors declare that this research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
RN and NKC involved in conceiving the concept and designing the study, analyzing and interpretation of data, revision, and final version approval of the manuscript. RN contributed for literature search, carried out the experiment, wrote the manuscript. NKC supervised the study. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The experimental protocol was approved by Institutional Animal Ethics Committee and all experiments were performed in compliance with the guidelines of the committee for the purpose of control and supervision of experimentation on animals.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Namoju, R., Chilaka, N.K. Maternal supplementation of α-lipoic acid attenuates prenatal cytarabine exposure-induced oxidative stress, steroidogenesis suppression and testicular damage in F1 male rat fetus. Beni-Suef Univ J Basic Appl Sci 11, 60 (2022). https://doi.org/10.1186/s43088-022-00240-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43088-022-00240-0