
Abstract
Backgrounds
Developed countries have a high mortality rate from atherosclerosis and are frequently linked to inflammation and other blood lipid disorders. MicroRNA expression can affect atherosclerotic plaque formation, lipid metabolism, inflammation, and other related processes. The search aimed to determine whether microRNA-138-5p or microRNA-132-3p expression levels are related to patient atherogenic genes.
Methods
Angiographic diagnostic method was used to select 45 healthy samples and 45 atherosclerosis patients, along with the laboratory and demographic information. After isolating peripheral blood mononuclear cells (PBMCs) from blood, the levels of miR-138 and miR-132 and the relative expression of Sirtuin 1 (SIRT1), signal transducer and activator of transcription-3 (STAT3), and CD36 genes measured using real-time PCR.
Results
miR-138 was upregulated compared to the control group in the atherosclerosis patient group (P < 0.05). In contrast, SIRT1 was downregulated in patients (P < 0.05). Our results also showed that the expression levels of miR-138 can use as a biomarker for atherosclerosis detection (P < 0.05). In addition, the expression of miR-138 with SIRT1 had a significant negative correlation (P < 0.05), and miR-132 was directly correlated with STAT3 (P < 0.01). Interestingly, STAT3 was negatively correlated with SIRT1 (P < 0.05) and positively with CD36 (P < 0.01).
Conclusion
Since atherosclerosis has no specific clinical symptoms and early diagnosis is vital, the use of miR-138 diagnostic biomarkers can play an essential role in early diagnosis. Furthermore, this study highlights the overlap of SIRT1-STAT3-CD36 signaling pathways with miR-132 and miR-138 in atherosclerosis.
Similar content being viewed by others

Introduction
Coronary artery disease (CAD) is the world's most common cardiovascular disorder. The most frequent underlying cause of CAD is atherosclerosis. Atherosclerosis is an arterial disorder causing lipoprotein accumulation in the endothelium and the oxidation of immune cells, including macrophages [1, 2]. In the subendothelial layer, monocytes transform into macrophages that develop inflammatory responses and eventually become foam cells [3]. Monocytes, dendritic cells, and lymphocytes are all components of peripheral blood mononuclear cells (PBMCs). It has been observed that inflammation and molecular mechanisms of these cells are involved in the pathogenetic processes that lead to heart disease [4].
CD36 is an 88-kDa glycoprotein found in adipocytes, cardiac myocytes, macrophages, platelets, and endothelial cells. In atherosclerosis progression, CD36 is bound by long-chain free fatty acids, making it easier for molecules like oxidized LDL to enter cells. Immune cell infiltration into arterial intima is induced by the interaction between CD36 and Ox-LDL [5, 6]. LDL, especially Ox-LDL, exhibits damage-associated molecular pattern characteristics and activates pro-atherogenic responses [7]. Recent research indicates that the signal transducer and activator of transcription 3 (STAT3) facilitates the uptake of fatty acids in cells by interacting with CD36. STAT3 signaling is activated in atherosclerosis [8, 9]. CD36 is a direct downstream target of STAT3, as overexpression of STAT 3 significantly raises its expression in cells [10]. Phosphorylation of STAT3 affects phenotypic transformation, growth, and migration of aortic vascular cells [11]. Research has demonstrated that Sirtuin 1 (SIRT1) controls STAT3's Tyr phosphorylation to reduce its rate of phosphorylation [12, 13]. SIRT1 is an essential regulator of vascular endothelial homeostasis [14] and found on the 10q21.3 chromosome. SIRT1 deacetylase activity can be influenced by factors, such as nutrition, and hormones to minimize cholesterol buildup in macrophages. Deficiency in this gene can lead pathophysiological alterations in cardiovascular diseases and is downregulated in myocardial damage [15,16,17]. SIRT1 can control CD36 expression [18] and is a target of miR-132 and miR-138 [14, 19].
MicroRNAs (miRNAs) are small non-coding RNAs that bind to 3 or 5 untranslated regions (UTRs) of messenger RNAs (mRNAs) to change post-transcriptional gene expression. Recent research has revealed that miRNAs play a crucial role in the development of cardiovascular disease [20,21,22,23]. MiR-138 is a vital miRNA family member with two putative genes for precursors, pre-miR-138–1 and pre-miR-138–2, located on chromosome 3p21.33 and 16q13, respectively [21]. MiR-138 has been implicated in heart disorders, as it regulates hypoxia-induced cell death and is expressed in hypoxic cardiomyocytes [22]. MiR-132 is a highly conserved miRNA located on human chromosome 17 that is produced during intergenic transcription and increased in cardiac tissue in response to cardiomyocyte stress [24, 25]. It has been found to have an effect on the signaling pathways controlling the growth, autophagy, handling of calcium, and contractility of cardiomyocytes [26,27,28,29]. Heart failure is caused by the progressive, adverse cardiac remodeling caused by miR-132 overactivation in cardiac tissue [30].
The function of miR-138-5p and miR-132-3p in peripheral blood mononuclear cell (PBMC) pools from patients with atherosclerosis (AS) remains unknown. The objective of this study was to investigate whether miR levels can be used to predict atherosclerotic disease and associate with atherogenic risk factors.
Materials and methods
Selection of subject populations
Ninety patients who underwent angiography and hospitalized in the cardiology departments of the Shahid Madani Hospital participated in this case–control research. The case group consisted of 45 individuals with particular atherosclerotic lesions in coronary angiography, whereas the healthy control group consisted of 45 individuals without angiographic lesions. The inclusion criteria were patients with coronary angiography diagnostic or therapeutic indication, matched age and sex, and excluded those with congenital heart disease, chronic kidney disease, pulmonary obstruction, or malignancy. Upon admission to the angiography clinic, a questionnaire was completed, asking about demographics, medical history, previous heart attacks, smoking, alcohol use, and exercise. Patients' lipid profiles and fasting blood sugar levels were assessed before atherogenic indices were computed. According to Gensini scoring, coronary artery lumen narrowing is graded as follows: One represents a narrowing of 1–25 percent; two represents attenuation of 26–50%; four represents attenuation of 51–75 percent; eight represents the attenuation of 76–90%; 16 represents the attenuation of 91–99%; and 32 represents a completely blocked vessel.
Atherogenic indices calculation
The atherogenic indices were computed using the following formulas: AIP (atherogenic index of plasma) = Log (serum triglyceride/serum HDLc); CRI-I (Castelli Risk Index II) = Serum total cholesterol/serum HDLc; CRI-II (Castelli Risk Index I) = Serum LDL cholesterol/serum HDLc; and TYG (Triglycerides-Glucose) = Ln((TG × FBS)/2).
Extraction of PBMCs
Blood samples from volunteers' peripheral veins were taken while persons were fasting and isolated PBMCs using the Ficoll–Paque density gradient method (Pharmacia, Freiburg, Germany). Blood diluted in 1:1 with phosphate-buffered saline (PBS). About 3.5 mL of Ficoll was poured into a centrifugation tube, and the samples were centrifuged at 800 g for 15 min at room temperature. After being washed with PBS twice, the PBMCs were then frozen to be used for RNA separation.
RNA extraction
Ribonucleic acid (RNA) is utilized in many molecular biology tests and is crucial for several biological processes. To extract RNA from cells, cells can initially be recovered as pellets after storage using a centrifugation procedure at 1500 g for 3 min at 4 °C. Total RNA was isolated from PBMCs according to the manufacturers' orders using an RNA isolation kit (Anacell, Iran). This RNA can be used as a substrate in various processes, such as the cDNA production of miR and mRNA. Purified and high-quality RNA, assessed spectrophotometrically at 260 nm and confirmed electrophoretically, is necessary for accurate and trustworthy gene expression data. The electrophoretic bands for the 18S and 28S rRNA genes served as evidence of RNA.
cDNA synthesis
All cDNA synthesis reactions were assumed with M-MLV. Reverse transcription first-strand complementary DNA was created by priming with oligo-dT (Yakta Tajhiz Azma) or a gene-specific miRNA primer based on the stem-loop (Zist Pooyesh) according to kit methods. After denatured at 70 °C for 5 min, RNAs and MicroRNA were then cooled in ice and added to a reverse transcription solution containing 1-µl dNTPs, 1-µl oligo-dT primer or 1.5-µl primer as miR specified, 1-µl M-MLV RT, 1-µl RNase inhibitor (Promega), and 4 µl of reaction buffer (Tris-HCl, KCl, dithiothreitol (DTT), and MgCl) in a final volume of 20 µl. The cDNA synthesis process was conducted for 60 min at 42 °C. Denaturing the reaction for 5 min at 70 °C stopped it.
Real-time PCR
Real-time PCR is a method that combines amplification and detection into a single step by continuously collecting data during the PCR process. This is accomplished by using fluorescent chemistry that relates the concentration of the PCR product to the fluorescence intensity. Real-time instrument (MIC-PCR) was used to analyze the mRNA expression. Additional file 1: Table S1 displays the distinct primer pairs. The amplifier included cDNAs made of 1-μg total RNA and used SYBR Green Real-Time PCR Master Mix (Amplicon, Denmark) with 0.8 μL of each primer. Amplification started with an initial 15 s of 94 °C incubation and was done using 40 cycles of the amplification step (94 °C 30 s and 60 °C 30 s). Results were calculated using the comparative threshold cycle Ct method (Ct) and expressed as the ratio of mRNA to GAPDH mRNA and miR to SNORD-47 microRNA as internal controls. It was possible to confirm the product identity (size and specificity) by electrophoresis on a 2 percent polyacrylamide gel.
Bioinformatics studies on miRNA targets
Using the KEGG database (http://www.genome.jp/kegg/pathway_facility), we explored signaling pathways that can be modulated by miRNA expression and were then assessed. The analysis identified two miR-regulated genes, mir-138 and miRP-132, related to angiogenesis pathways. The miRTarBase database (http://mirtarbase.mbc.nctu.edu.tw/php/download.php) was utilized to verify the targets.
Statistical analysis
GraphPad Prism version 5.0a and IBM's Statistical Package for the Social Sciences (SPSS) software were used to evaluate the data that had been collected. For statistical calculations of gene expression, the 2^ (−∆∆CT) method is used. The Kolmogorov–Smirnov test, Student's t-test, Mann–Whitney U-tests, and logistic regression analysis were used to assess the relationship between gene expression and AS. The area under the curve (AUC) was determined to evaluate the diagnostic capability of miR-132 and miR-138 mRNA expression in PBMCs for discriminating AS patients from those without the condition. The significance was set at P < 0.05.
Results
Basic characteristics of the subjects
Table 1 provides specifics about the patient's demographic and clinical parameters. The baseline traits of the two groups were almost identical. In the case and control groups, there was no apparent distinction between age, weight, sex, or body mass index (BMI). Triglycerides, fasting glucose levels, creatinine, Castelli index 2, AIP, and TYG were all significantly elevated in patients; AIP with (P value = 0.001), TYG with (P value = 0.001), creatinine with (P value = 0.037), Castelli index 2 with (P value = 0.01), fasting glucose with (P value = 0.045), and triglycerides with (P value = 0.005).
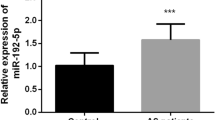
Increased miR-138-5P levels in PBMCs of AS patients
The real-time PCR assay was employed to clarify the expression of miRs. The findings showed that patients' PBMC miR-138 levels were significantly greater than those of healthy persons (P < 0.05; Fig. 1). In addition, the PBMC of the case group revealed a trend toward increased expression of the miR-132 gene, but this was not statistically significant (P > 0.05; Fig. 1). Our results imply that deregulation of miR-138 may be a critical factor in AS development.

Expression of miR-138 (A, P < 0.05) and miR-132 (B, P > 0.05) in PBMC cells of patients and controls. *Symbol means P < 0.05
Decreased SIRT1 gene expression in PBMCs of AS patients
We assessed the expression levels of two lipid metabolism-related biomarkers, SIRT1 and CD36, in the case and the control groups. Our results showed that patients' SIRT1 expression levels were lower than in the control group (P < 0.01; Fig. 2). However, the study revealed that, although not considerably, the patient group's CD36 and STAT3 levels were raised.

Expression of SIRT1 (A, P < 0.01), CD36 (B), and STAT3 (C), in PBMC cells of patients and controls. **Symbol means P < 0.01
Diagnostic potential of PBMC miR‑138‑5p in AS
A variable's potential to function as a disease biomarker can be assessed using the ROC curve. Based on sensitivity and specificity, this curve was created. The capability of the test to provide a diagnosis is represented by the area under the ROC curve. The elevated expression of miR-138 in mononuclear blood cells, as shown in Fig. 3, indicates that the biomarker is suitable for differentiating study participants with atherosclerosis from controls (P < 0.01). The area under the curve (AUC) was 0.71, the sensitivity was 56%, and the specificity was 65%. The control and case groups were distinguished using ROC curve analysis based on atherosclerosis markers. Higher values were found for TYG and AIP as predictive variables (AUC: TYG = 0.78 and AUC: AIP = 0.70), (P < 0.01).

Receiver operating characteristic (ROC) curves for models to predict atherosclerosis. AUC showed higher value in model for miR-138 as a new marker for AS detection (P < 0.05)
PBMC miR-138-5p was inversely correlated with SIRT1 gene expression, but miR-132-3p was positively correlated with STAT3 in AS patients
In PBMC, the relationships between genes and blood biochemical markers were examined further. SIRT1 and miR-138 have a statistically significant negative correlation (r: -0.42, P = 0.02). STAT3 and miR-132 positively correlated (r: 0.37, P = 0.006). Additionally, the correlation between STAT3 and CD36 was considerably positive ////(r: 0.36, P = 0.005), but the correlation between STAT3 and SIRT1 was significantly negative (r: -30, P = 0.02) (Fig. 4). miR-132 had a positive correlation with BMI (r: 0.32, P = 0.02), and the correlation of BMI with CRI-I was also positive (r: 0.46, P = 0.02).

Correlation analysis for miR-138, miR-132, SIRT1, STAT3, and CD36. Negative correlation of miR-138 and SIRT1 (A) and positive correlation of miR-132 and STAT3 (B). Negative correlation of STAT3 and SIRT1 (C). Positive correlation of STAT3 and CD36 (D)
Relationship between AS MicroRNAs and AS-associated biological signaling pathways, in silico
We used the miR-target scanning database (https://www.microrna-target.org/) and the Kegg pathway (https://www.genome.jp/kegg/pathway.html) to characterize the miR-binding biological function. Specifically, we have looked at miR-138 and its associated signaling to better understand the AS biology of this expressed miR in PBMCs. Only first-level interactions between (SIRT1 mRNA) and has-miR-138-5p (AGCUGGUGUUGUGAAUCAGGCCG) were noticed by the RNAhybrid program (https://bibiserv.cebitec.uni-bielefeld.de/rnahybrid/). ∆G was -25 (Fig. 5).

Prediction of post-transcriptional interaction between SIRT1 mRNA 3’UTR and miR-138-5p as evaluated by RNA-hybrid program. The interaction resulted in a significant minimal free energy (-25.4 mfe for SIRT1) forming thus a stable hybrid structure
Discussion
MiRNAs play a crucial role in atherosclerosis, specifically in lipid metabolism regulation, potentially initiating coronary artery disease and its diagnosis. Peripheral blood mononuclear cells (PBMCs) are a valuable source for biological research due to their easy preparation and potential involvement in the pathogenicity of heart disease [4]. The study highlights the noninvasive nature of miR-138 expression in PBMCs as a potential biomarker for AS diagnosis. Patients had higher PBMC miR-138 and lower SIRT1 levels, with a negative correlation between both. SIRT1 and STAT3 had a reverse interaction. There was a direct correlation between STAT3 with CD36 and miR-132.
The study found that miR-138 might regulate atherosclerosis progression through SIRT1 gene expression blocking. SIRT1 plays a protective role in atherosclerosis and vascular processes, exerting anti-inflammatory effects on endothelial cells and macrophages by preventing NF-kB signaling, decreasing cytokine expression, and suppressing Ox-LDL expression. It also inhibits the expression of coagulation factor III, which has antithrombotic properties [31, 32]. Sun S's 2021 research suggests that miR-138-5p, targeting SIRT1's 3'-UTR and activating p53 signaling, leads to the apoptosis of human cardiomyocytes during heart failure [33]. In 2015, Juan Xu and his team discovered that miR-138 can inhibit SIRT1 expression and promote the migration and growth of vascular smooth muscle cells in mice [14]. The study confirms a negative correlation between SIRT1 and STAT3, suggesting that a decrease in SIRT1 and loss of STAT3 control could lead to increased AS development. STAT3 is involved in physiological processes such as oxidative stress, cell proliferation, apoptosis, chronic inflammation, and myocardial infarction in various cardiovascular disorders [34,35,36,37]. STAT3 is believed to promote atherosclerosis by secreting pro-inflammatory factors from foam cells, which enhance CD36 expression and cell replication [37]. At this point, we found a positive correlation between STAT3 and CD36. One 2010 study by Iliopoulos D et al. identified putative STAT3 binding sites in miR-132-3p promoters [38]. A survey by Dongqing Li et al. in 2015 indicated that miR-132 increased STAT3 activity and phosphorylation, promoting cell growth during wound healing [39]. MiR-132 is a potential marker and therapeutic goal for detecting and controlling ischemia, regulating hypertrophy, fibrosis, cell adhesion, and the progression of ischemic heart failure [38]. The involvement of miR-132 pathway in atherosclerosis is unclear. STAT3 may be connected to the elevated levels of this gene and the increased risk of myocardial infarction. As shown in our study, the direct correlation between these two genes confirms this problem.
In summary, it is confirmed that CD36's role in regulating the proliferation of vascular smooth muscle cells (VSMCs) significantly impacts neointimal hyperplasia and the progression of vascular disorders [39]. One possible strategy for treating atherosclerosis could be to regulate and control the expression of CD36 [40]. Research shows that CD36 overexpression is facilitated by STAT3 [9]. SIRT1 negatively affects the control of pSTAT3 and STAT3 expression [41]. CD36 mediates the active uptake of Ox-LDL into macrophages [6]. SIRT1 inhibits CD36 expression, which reduces Ox-LDL absorption and prevents the development of macrophage foam cells [42]. This suggests that drugs that increase SIRT1 expression and reduce STAT3 expression might have the potential for treating or preventing CD36-related disorders. On the other hand, we found that miR-139 and miR-132 linked positively with CD36 and STAT3 and negatively with SIRT1 (Fig. 6). A mechanism for maintaining balance should be taken into account by downregulating miR-139 and miR-132 to their normal levels, enabling the combined control of SIRT1 and STAT3 to regulate CD36 and prevent foam cell formation. MicroRNAs are crucial in clinical medicine due to their ability to be detected in tissue and biological fluids, making them useful for early disease diagnosis. They could also be used to treat human diseases by returning normal microRNA levels, making them a significant factor in the future disease diagnosis and treatment [43].

The overlap of SIRT1-STAT3-CD36 signaling pathways with miR-132 and miR-138 in atherosclerosis
Conclusion
Our research revealed that atherosclerosis may be influenced by the modification of gene expression in PBMCs. The measurement and analysis of these genes' expression can be utilized to create a consistent gene expression profile instrument for early detection of atheroic lesions. Moreover, our detections indicate that the increased expression of miR-138 in PBMCs could be a novel biomarker for detecting atherosclerosis. However, this study highlights overlapping SIRT1-STAT3-CD36 signaling pathways with miR-132 and miR-138 in atherogenesis. Hence, further investigation is required to inspect this signaling in cell lines and identify therapeutic targets.
Limitations
This study has limitations, including a small sample size of healthy subjects and patients with atherosclerosis, being conducted at a single center, and potential selection bias due to the cross-sectional design. Expanding the data by increasing the number of samples from several centers or hospitals and including asymptomatic individuals in the control group will improve the study's validity in the future research.
Availability of data and materials
The datasets used analyzed during the current study are available from the corresponding author on reasonable request.
Change history
31 January 2024
A Correction to this paper has been published: https://doi.org/10.1186/s43042-024-00473-x
References
Zhang Q-Y et al (2018) Hydrogen sulfide regulating myocardial structure and function by targeting cardiomyocyte autophagy. Chin Med J 131(07):839–844
Falk E (2006) Pathogenesis of atherosclerosis. J Am Coll Cardiol 47(8S):C7–C12
Chistiakov DA, Bobryshev YV, Orekhov AN (2016) Macrophage-mediated cholesterol handling in atherosclerosis. J Cell Mol Med 20(1):17–28
Voellenkle C et al (2010) MicroRNA signatures in peripheral blood mononuclear cells of chronic heart failure patients. Physiol Genom 42(3):420–426
Silverstein RL, Li W, Park YM, Rahaman SO (2010) Mechanisms of cell signaling by the scavenger receptor CD36: implications in atherosclerosis and thrombosis. Trans Am Clin Climatol Assoc 121:206
Park YM (2014) CD36, a scavenger receptor implicated in atherosclerosis. Exp Mol Med 46(6):e99–e99
Minelli S, Minelli P, Montinari MR, Reflections on atherosclerosis: lesson from the past and future research directions. J Multidiscipl Healthcare, 2020, p. 621–633.
Rozovski U et al (2017) Overexpression of CD36, driven by STAT3, mediates free fatty acid uptake in CLL cells. Blood 130:4301
Rozovski U et al (2018) STAT3-activated CD36 facilitates fatty acid uptake in chronic lymphocytic leukemia cells. Oncotarget 9(30):21268
Su T et al (2020) Apigenin inhibits STAT3/CD36 signaling axis and reduces visceral obesity. Pharmacol Res 152:104586
Liao X-H et al (2015) STAT3 protein regulates vascular smooth muscle cell phenotypic switch by interaction with myocardin. J Biol Chem 290(32):19641–19652
Nie Y et al (2009) STAT3 inhibition of gluconeogenesis is downregulated by SirT1. Nat Cell Biol 11(4):492–500
Zhang N et al (2020) Septin4 prevents PDGF-BB-induced HAVSMC phenotypic transformation, proliferation and migration by promoting SIRT1-STAT3 deacetylation and dephosphorylation. Int J Biol Sci 16(4):708
Xu J, Li L, Yun H-F, Han Y-S (2015) MiR-138 promotes smooth muscle cells proliferation and migration in db/db mice through down-regulation of SIRT1. Biochem Biophys Res Commun 463(4):1159–1164
Houtkooper RH, Pirinen E, Auwerx J (2012) Sirtuins as regulators of metabolism and healthspan. Nat Rev Mol Cell Biol 13(4):225–238
Zané F, Ademola A, Emmanuel M (2014) Control of carbohydrate and lipid metabolism by NRF-1 and sirtuins: implications on type 2 diabetes and obesity. Chem Biol Lett 1(2):66–76
Li X (2013) SIRT1 and energy metabolism. Acta Biochim Biophys Sin 45(1):51–60
Niu B et al (2018) SIRT1 upregulation protects against liver injury induced by a HFD through inhibiting CD36 and the NF-κB pathway in mouse kupffer cells. Mol Med Rep 18(2):1609–1615
Zhou Y, Li KS, Liu L, Li SL (2020) MicroRNA-132 promotes oxidative stress-induced pyroptosis by targeting sirtuin 1 in myocardial ischaemia-reperfusion injury. Int J Mol Med 45(6):1942–1950
Xu S et al (2012) MicroRNAs as potential novel therapeutic targets and tools for regulating paracrine function of endothelial progenitor cells. Med Sci Monit Int Med J Exp Clin Res 18(7):27
Liu X et al (2009) MicroRNA-138 suppresses invasion and promotes apoptosis in head and neck squamous cell carcinoma cell lines. Cancer Lett 286(2):217–222
Zhu H, et al., MiR-138 protects cardiac cells against hypoxia through modulation of glucose metabolism by targetting pyruvate dehydrogenase kinase 1. Biosci Rep, 2017. 37(6).
Salic K, De Windt LJ (2012) MicroRNAs as biomarkers for myocardial infarction. Curr Atheroscler Rep 14:193–200
Ucar A et al (2012) The miRNA-212/132 family regulates both cardiac hypertrophy and cardiomyocyte autophagy. Nat Commun 3(1):1078
Eskildsen TV et al (2013) Angiotensin II regulates microRNA-132/-212 in hypertensive rats and humans. Int J Mol Sci 14(6):11190–11207
Foinquinos A et al (2020) Preclinical development of a miR-132 inhibitor for heart failure treatment. Nat Commun 11(1):633
Batkai S et al (2021) CDR132L improves systolic and diastolic function in a large animal model of chronic heart failure. Eur Heart J 42(2):192–201
Lu D, Thum T (2019) RNA-based diagnostic and therapeutic strategies for cardiovascular disease. Nat Rev Cardiol 16(11):661–674
Lavenniah A et al (2020) Engineered circular RNA sponges act as miRNA inhibitors to attenuate pressure overload-induced cardiac hypertrophy. Mol Ther 28(6):1506–1517
Ponikowski P et al (2016) 2016 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure. Kardiologia Polska (Polish Heart J) 74(10):1037–1147
Stein S, Matter CM (2011) Protective roles of SIRT1 in atherosclerosis. Cell Cycle 10(4):640–647
Lu C et al (2023) Novel role of the SIRT1 in endocrine and metabolic diseases. Int J Biol Sci 19(2):484–501
Sun S, Wang C, Weng J (2021) MicroRNA-138-5p drives the progression of heart failure via inhibiting sirtuin 1 signaling. Mol Med Rep 23(4):1–1
Zhang W et al (2016) Critical roles of STAT3 in β-adrenergic functions in the heart. Circulation 133(1):48–61
Zouein FA et al (2013) Role of STAT3 in angiotensin II-induced hypertension and cardiac remodeling revealed by mice lacking STAT3 serine 727 phosphorylation. Hypertens Res 36(6):496–503
Yu L et al (2011) AIP1 prevents graft arteriosclerosis by inhibiting interferon-γ–dependent smooth muscle cell proliferation and intimal expansion. Circ Res 109(4):418–427
Paulin R et al (2011) Signal transducers and activators of transcription-3/pim1 axis plays a critical role in the pathogenesis of human pulmonary arterial hypertension. Circulation 123(11):1205–1215
Xu K et al (2021) Advances in miR-132-based biomarker and therapeutic potential in the cardiovascular system. Front Pharmacol 12:751487
Yue H et al (2019) CD36 enhances vascular smooth muscle cell proliferation and development of neointimal hyperplasia. Arterioscler Thromb Vasc Biol 39(2):263–275
Tian K, Xu Y, Sahebkar A, Xu S (2020) CD36 in atherosclerosis: pathophysiological mechanisms and therapeutic implications. Curr Atheroscler Rep 22:1–10
Wang W et al (2018) JAK1-mediated Sirt1 phosphorylation functions as a negative feedback of the JAK1-STAT3 pathway. J Biol Chem 293(28):11067–11075
Hui B et al (2021) Gypenoside inhibits ox-LDL uptake and foam cell formation through enhancing Sirt1-FOXO1 mediated autophagy flux restoration. Life Sci 264:118721
Natarajan SK et al (2013) MicroRNA function in human diseases. Medical Epigenetics 1(1):106–115
Acknowledgements
We thanked the officials of Lorestan University of Medical Sciences Research Center and our dear colleagues who helped in this research.
Funding
No funding was received to assist with the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
GS has conceived the manuscript and revised it. SE and MM provided clinical data and information and performed the technical tests. FA provided data collection system design, analysis, and interpretation of data. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The present study was approved by Lorestan and Dezful University of Medical Science and the Ethics Committee, Khorramabad, Dezful, Iran (ir.dums.rec.1398.016). All the procedures performed in the studies involving human participants were in accordance with ethical standards of local ethics committee of Lorestan University of Medical Science. Written informed consent was obtained from all patient’s subjects.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original version of this article was revised: we have added author Maysam Mard-Soltani as Co-first Author of this paper
Supplementary Information
Additional file 1. Table S1.
Primer sequences.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ehsani, S., Mard‑Soltani, M., Ahmadpour, F. et al. Association between miR-138-5p, miR-132-3p, SIRT1, STAT3, and CD36 and atherogenic indices in blood mononuclear cells from patients with atherosclerosis. Egypt J Med Hum Genet 24, 84 (2023). https://doi.org/10.1186/s43042-023-00464-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43042-023-00464-4