
Abstract
Background
Efficient and selective utilization of metabolic substrates is one of the key strategies in high-altitude animals to cope with hypoxia and hypothermia. Previous findings have shown that the energy substrate utilization of highland animals varies with evolutionary history and phylogeny. The heart is a proxy for the cardiopulmonary system, and the metabolic substrate utilization in the myocardium is also under the strong selective pressure of chronically hypoxic and hypothermic environments. However, little information is available on the physiological adjustments in relation to metabolic substrate utilization in the myocardium for coping with high-altitude environments.
Methods
We compared the metabolic enzyme activities, including hexokinase (HK), phosphofructokinase (PFK), pyruvate kinase (PK), citrate synthase (CS), carnitine palmitoyl transferase 1 (CPT-1), lactic dehydrogenase (LDH), and creatine kinase (CK), and metabolic substrate contents including glucose (Glu), triglyceride (TG), and free fatty acid (FFA) in the myocardium of a typical human commensal species, Eurasian Tree Sparrows (Passer montanus) between the Qinghai-Tibet Plateau (the QTP, 3230 m) and low altitude population (Shijiazhuang, 80 m), and between sexes.
Results
Among the seven metabolic enzymes and three substrates investigated, we identified no significant differences in PK, CPT-1, HK, CS, LDH, and CK activities and TG content of the myocardium between high and low altitude populations. However, the QTP sparrows had significantly lower Glu content and PFK activities but higher FFA content relative to their lowland counterparts. In addition, male sparrows had higher myocardial HK and CS activities relative to females, independent of altitude.
Conclusions
Our results showed that the QTP sparrows elevated fatty acid utilization rather than glucose preference in the myocardium relative to lowland counterpart, which contributes to uncovering both the physiological adjustments for adapting to the extreme conditions of the QTP, intraspecifically.
Similar content being viewed by others


Background
High-altitude environments are characterized by low oxygen partial pressure and low ambient temperature (Storz et al. 2010). Theoretically, high-altitude endotherms have to cope with the hypobaric-hypoxia conditions by enhancing oxygen delivery and oxidative phosphorylation for metabolic thermogenesis relative to their low-altitude counterparts (Cheviron et al. 2014). For lowland dwellers, a suppressed overall metabolic intensity and enhanced glycolysis capacity are believed to be biochemical responses to hypoxic conditions due to the lowered blood O2-delivery capacity (Cartee et al. 1991; Zinker et al. 1994; Azevedo Jr et al. 1995; Holden et al. 1995; Roberts et al. 1996a, 1996b). In contrast to their lowland counterparts, high-altitude natives have possessed a suite of traits (e.g. bigger body size, larger lung, higher blood-O2 affinity, and more capillaries in organs) to overcome the consequences of hypoxic environments (Storz and Moriyama 2008; Qu et al. 2020). Furthermore, previous “Omics” studies have shown that strong positive selection in lipids metabolism is identified in those native endotherms of highland areas, such as birds (Ground Tit Parus humilis) (Qu et al. 2013), and mammals (Yak Bos mutus and Plateau Zokor Myospalax baileyi) (Qiu et al. 2012; Shao et al. 2015). Such an enhanced lipid metabolism is consistent with that of endotherms coping with cold, which suggests that the cold, rather than hypoxia, is the main stressful factor for free-living highland natives (Weber 2011). In humans, highland dwellers (i.e. Tibetans and Sherpas) also expressed a suite of convergent adaptations (e.g. larger motor organs and enhanced hemoglobin-O2 affinity) that are similar to those highland natives. However, they exhibited a strong positive selection in carbohydrate metabolism rather than lipid metabolism (Ge et al. 2012; Qu et al. 2015; Horscroft et al. 2017; Jing et al. 2021). Therefore, these findings suggest that evolutionary history, phylogeny, and habitat characteristics jointly play important roles in shaping the diversified strategies of high-altitude adaptations.
The heart is the core engine for metabolism, which can drive the delivery of O2 and metabolic substrates from the blood to target tissues and metabolic wastes from target tissues to excretory organs. For high-altitude animals, an efficient heart, therefore, is a proxy for the cardiopulmonary system under strong selective pressure of chronic low-oxygen environments (Ivy and Scott 2015; Sun et al. 2016; Tate et al. 2017; Qu et al. 2020). Of importance, the heart has great flexibility on substrate usage that allows it to switch back and forth between fatty acid and carbohydrate oxidation, depending on the energy requirements and nutritional state of the organism, and the availability of glycolipid metabolic substrates to myocardium (Lopaschuk et al. 2010). Whether highland animals improve their metabolic capacities in the myocardium relative to their lowland counterparts is of interest and remains largely unclear.
In the myocardium, the mitochondrial oxidative phosphorylation contributes to approximately, 95% of adenosine triphosphate (ATP) generation, and glycolysis contributes to the remainder (Ventura-Clapier et al. 2004). The carbohydrates (glucose and lactate) and fatty acids are the two primary energy substrates used by mitochondrial oxidative phosphorylation (Kodde et al. 2007). In human, carbohydrates are more efficient than fatty acids, which can generate more contractile power for any given rate of myocardial oxygen consumption (Neglia et al. 2007). An increase in the uptake of glucose (Glu) and lactate is believed to enhance myocardial metabolic efficiency under acute cardiac stress, especially the contribution of glycolysis to the myocardial ATP production (Gertz et al. 1988; Camici et al. 1991; Bagger et al. 2000). In laboratory model mammals, the myocardial ischemia induced by hypoxia can increase the glycolysis to fulfill the workload of the heart, with a relative decrease in lipids uptake (Gertz et al. 1988; Fuxe et al. 1989; Camici et al. 1991; Bagger et al. 2000). On the other hand, myocardial fatty acids account for 60–90% of the total ATP generated in the resting fasting state (van der Vusse et al. 1992), which derives from the circulating triglyceride (TG) (Wang et al. 1998). However, there is limited knowledge on the substrate preference of either Glu or lipids utilization in the myocardium of highland animals.
The cardiovascular systems of birds and mammals are highly similar in form and modulation (Burggren et al. 2011). However, birds have larger hearts, cardiac outputs, and stroke volumes but lower heart rates than mammals when body mass is controlled (Calder 1968; Grubb 1983). Furthermore, birds also have higher blood-O2 affinities (Storz and Moriyama 2008), do not have a hypoxic pulmonary vasoconstriction response (Scott et al. 2011), and are insensitive to hypercapnia (Faraci et al. 1984; Durmowicz et al. 1993; Groves et al. 1993; Ge et al. 1998; Sakai et al. 2003). These characteristics are critical for birds to fulfill O2 utilization for organs (tissue) under hypobaric hypoxia conditions.
The Qinghai-Tibet Plateau (QTP) is the largest and highest plateau worldwide, featuring typical hypobaric hypoxia conditions (Thompson et al. 2000). Previous studies have found that some birds on the QTP genetically increased hemoglobin-O2 affinity and exhibited stronger hypoxia tolerance than their lowland counterparts (Scott and Milsom 2007; Zhu et al. 2018; Hao et al. 2019). As a small-sized passerine, the Eurasian Tree Sparrow (Passer montanus) is widely distributed from the lowland areas to the QTP (Summers-Smith 2018). As a typical human commensal species, this small-sized passerine bird is ideal for studying adaptative strategies to high-altitudes, owing to having almost similar high-altitude adaptation history with the Tibetans. It is known that Eurasian Tree Sparrows on the QTP have larger body sizes and wings, greater size-corrected pectoralis and lungs (Sun et al. 2016), larger myofibers in pectoralis but not in the myocardium (Qu et al. 2020), have similar patterns of stress responses relative to their lowland counterparts (Li et al. 2011).
It was estimated that Eurasian Tree Sparrows had occupied the QTP after agricultural civilization (about 2600 years; Qu et al. 2020). Based on those findings in native Tibetans or those lowland animals acclimatizing to high-altitude environments showing a relatively lower ratio of lipid utilization but a higher ratio of carbohydrate utilization (Young et al. 1982, 1992; Sutton et al. 1983; Green et al. 1989; Brooks et al. 1991, 1992; Roberts et al. 1996a), we hypothesized that the QTP Eurasian Tree Sparrows would enhance carbohydrate metabolism in the myocardium rather than lipid metabolism than their lowland counterparts. To validate our hypothesis, we compared the maximal activities of three rate-limiting enzymes in glycolysis (hexokinase, HK; phosphofructokinase, PFK; pyruvate kinase, PK), one key enzyme of the TCA cycle (citrate synthase, CS), a rate-limiting enzyme for transporting long chain free fatty acid (FFA) into the mitochondria (carnitine palmitoyl transferase 1, CPT-1); maximal activities for lactic dehydrogenase (LDH, a measure of tissue damage) and creatine kinase (CK; a measure of cardiac functions) and metabolic substrate contents for Glu, TG, and FFA in the myocardium between the QTP and lowland Eurasian Tree Sparrows.
Methods
Study area and sample collection
A total of 29 adult Eurasian Tree Sparrows were sampled using nylon mist nets from the QTP (8 males and 9 females; Gonghe County, Qinghai Province, 36.28°N, 100.62°E, altitude of 3230 m; average temperature range: – 4 to 3 °C) and lowland area (7 males and 5 females; Pingshan County, Hebei Province, 38.25°N, 114.20°E, altitude of 80 m; average temperature range: of 3–6 °C) during the wintering stage between December 2014 and February 2015. Within 3–4 h post-capture, the birds were anesthetized by injecting with phenobarbitone (7.5 μL/g body mass) and humanely sacrificed. The left ventricle of hearts was rapidly dissected, washed with ice-cold phosphate buffer. All the samples were rapidly frozen in liquid nitrogen and stored at – 80 °C until assay.
Assays of metabolic enzyme activities and substrate concentrations
Samples of cardiac muscle were pulverized with a ceramic mortar and pestle under liquid nitrogen to powder. The powder was suspended by exacting solution (1:10, w/v) in an ice water bath. The mixed liquor was centrifuged at 11,000×g for 10 min at 4 °C, and the supernatant was transferred to the new EP tube. Total protein content was measured by the BCA method with commercially available kits (CW0014, CoWin Biotech Co. Ltd., Jiangsu, China), and using BSA as standard.
The maximal enzyme activities for all supernatant samples were measured in duplicate according to the manufacturer’s protocols. The kits for CPT-1 measurement were purchased from Enzyme-linked Biotechnology Co. Ltd. (Shanghai, China). The kits for the measurement of CS, HK, PFK, and PK were purchased from Ke-Ming Corp (Suzhou, China). Maximal enzyme activities of LDH and CK measured by an automated biochemistry analyzer (Mindray, mode BS-180, Shenzhen, China) using commercially available kits (Mindray, Shenzhen, China). The substrates were measured in 25 μL supernatant samples diluted with dH2O (1:39) using an automatic biochemical analyzer (Mindray BS-180) with commercially available kits (Mindray Corp., Shenzhen, China). All samples were measured in duplicate. Intra- and inter-assay variation were 3.1% and 8.2% (Glu), 9.6% and 10.1% (TG), 4.2% and 6.3% (FFA), respectively. Assay sensitivity was 0.3 mmol/L (Glu), 0.03 mmol/L (TG), 0. 03 mmol/L (FFA).
Statistical analysis
The effects of altitude, sex, and the interaction of altitude and sex on the maximal enzyme activities of CS, CPT-1, HK, PFK, and PK, and substrate concentrations of Glu, TG, and FFA in the myocardium were determined using a generalized linear model (GLM), respectively. If a statistically significant effect was detected, we conducted multiple comparisons in each of the two groups using Bonferroni post hoc tests. A P value < 0.05 represents a statistically significant difference. All the analyses were performed using IBM SPSS Statistics 25 (IBM Inc., NY, USA), and figures were generated using GraphPad Prism 8.4 (GraphPad Software Inc., CA, USA). The data are expressed as the mean ± SEM.
Results
Effects of altitude and sex on metabolic enzyme activities in the myocardium
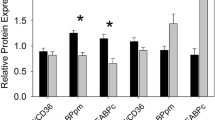
The myocardial PK, CPT-1, CK, and LDH activities of Eurasian Tree Sparrows did not vary with altitude, sex, and the interaction of altitude and sex (Table 1). The myocardial PFK did not vary with sex and the interaction of altitude and sex but varied with altitude. Post hoc results showed that high-altitude sparrows had significantly higher PFK activities relative to low-altitude ones (Table 1; Fig. 1a). The myocardial HK and CS did not vary with altitude and the interaction of altitude and sex but varied with sex. Post hoc results showed that male sparrows had significantly higher myocardial HK and CS activities than females (Table 1; Fig. 2a, b).

Comparisons of maximal phosphofructokinase (PFK) activity (a), glucose (Glu) content (b), and free fatty acid (FFA) content (c) in the myocardium of Eurasian Tree Sparrows (Passer montanus) between high altitude (3230 m) and low altitude (80 m) areas. The statistical results are shown in Table 1. The asterisk represents a significant difference (P < 0.05) between groups

Comparisons of maximal hexokinase (HK) activity (a) and citrate synthase (CS) activity (b) in the myocardium of Eurasian Tree Sparrows (Passer montanus) between sexes. The statistical results are shown in Table 1. The asterisk represents a significant difference (P < 0.05) between groups
Effects of altitude and sex on glucolipid substrates in the myocardium
The myocardial TG did not vary with altitude, sex, and the interaction of altitude and sex (Table 1). The myocardial Glu and FFA contents did not vary with sex and the interaction of altitude and sex but varied with altitude. Post hoc results showed that high-altitude sparrows had significantly lower myocardial Glu but higher FFA concentrations (Fig. 1b, c).
Discussion
In the present study, we found that maximal enzyme activities of myocardial HK and PK in Eurasian Tree Sparrows did not vary with altitude, but the QTP population had significantly lower maximal enzyme activities of PFK and glucose content in the myocardium. As a critical enzyme for phosphorylating fructose 6-phosphate to fructose 1, 6-bisphosphate of glycolysis, the PFK is negatively inhibited by ATP and citrate (fuel substrate of the TCA cycle). Theoretically, enhanced PFK activity will consume more Glu and indirectly result in a reduction of Glu levels, which will eventually lead to mobilizing other fuel substrates. Our observed lower Glu contents and decreased maximal PFK activities in the myocardium, along with lower plasma Glu levels of the QTP Eurasian Tree Sparrows relative to their lowland counterparts (Li et al. unpublished data), suggesting highland sparrows may weaken the glycolytic pathway through lowering maximal PFK activity and tend to depend on other fuel substrates.
Meanwhile, we found the QTP sparrows had comparable TG contents and maximal CPT-1 activities but increased FFA levels in the myocardium relative to lowland ones. In the hearts, the increased FFA supply through hydrolysis of circulating lipoproteins (via lipoprotein lipase) is considered as a way to compensate for the diminished contribution of glucose as an energy substrate (Pulinilkunnil and Rodrigues 2006). Therefore, our results suggest that the QTP sparrows should enhance the FFA transportation from blood to compensate for the lowered Glu contents in the heart. This preference for metabolic substrate utilization is similar to that of Deer Mice (Peromyscus leucopus), a highland native small mammalian species (Sears et al. 2006; Vaillancourt et al. 2009; Cheviron et al. 2012). Previous studies have shown that the QTP sparrows have greater body weight and heart size (Sun et al. 2016) relative to the lowland ones, which is believed to be fundamental for preventing heat dissipation and enhancing energy efficiency. Considering that lipids have a much higher energy density than carbohydrates (Berg et al. 2002; Hochachka and Somero 2002; Belitz et al. 2009), the increase in FFA and decrease in Glu influx in the myocardium might be a metabolic strategy to enhance energy efficiency for coping with the extreme environments. Whether this strategy occurs in other organs (tissues) remains to be further determined.
We detected both highland and lowland sparrows had similar maximal LDH and CK activities, which means similar maximal velocity on lactate metabolism for both populations. There are two possible explanations: (1) highland sparrows are able to counter the adverse effect of anaerobic metabolism, which is more easily induced by hypoxic environment (Samaja et al. 1997; Pilarski et al. 2009), on acid–base equilibrium in cardiomyocytes via lower glycolysis level (Burggren and Cameron 1980; Casiday and Frey 2012; Hebisz et al. 2016); (2) both population possessed similar maximal capacity on production and regeneration of ATP by lactate metabolism (Ingwall et al. 1985; Kodde et al. 2007), which indicate that the highland sparrows, compared to their lowland counterparts, could better cope with the extra demand of ATP when the cardiac workloads increased but the aerobic metabolism is suppressed. Since the leakage of LDH and CK into the cardiovascular system are documented as indexes of myocardial injury (Kaur et al. 1997), our results showed that the cardiac functions of both the highland and lowland sparrows were unlikely to be injured. Considering that limited information on cardiac functions is available between high and low altitude animals, other parameters such as heart beat rate and blood pressure are also fundamental for better understanding the physiological and ecological strategy of high-altitude adaptation, warrant to be further investigated.
Interestingly, we found male sparrows had significantly higher activities of HK and CS than females, independent of altitude. Our results are inconsistent with the finding showing no differences in CS and HK activities in the myocardium between sexes in rats (Milerová et al. 2016). Generally, the sex-based differences for animals in behavior, neuroendocrinology, and physiology are generally related to stress response, locomotor ability and body composition, etc. (Blanchard et al. 1995; Koolhaas et al. 1995; Martinez et al. 1998). Furthermore, several researchers have reported sex-based differences in the human cardiovascular system, such as in cardiac growth, O2 uptake, resting heart rate, myocardial contractile performance and modulation of arterial blood pressure (Vinet et al. 2003; Kolar and Ostadal 2013). These sex-dependent differences in the cardiovascular system are largely associated with sex hormones (Hayward et al. 2001; Lagranha et al. 2010; Murphy 2011). In our previous study, we did not detect sex-based differences in baseline and stress-induced corticosterone content (Sun et al. unpublished data) between highland and lowland Eurasian Tree Sparrows, which can rule out the metabolic impacts induced by glucocorticoids. Moreover, male sparrows tend to have longer wings, greater wing areas, and total flight muscle mass relative to females (Mónus et al. 2011; Sun et al. 2016, 2017). Whether increased overall cardiac metabolic capacity and myocardium glycolysis are associated with meeting the energy requirements of their vigorous metabolic organs for flight remains further investigation.
Conclusions
In the present study, we identified that the QTP Eurasian Tree Sparrows have lower maximal myocardial PFK activities and Glu content but higher FFA content relative to their lowland counterparts. Although this species is an invasive species on the QTP introduced along with humans, it exhibited reduced Glu utilization and enhanced FFA utilization in the myocardium, which differs from the Tibetans showing increased carbohydrate utilization but agrees with those small-sized high-altitude natives. Our results did not detect any differences in other measured metabolic enzyme activities between highland and lowland populations, indicating the maximal activities of these metabolic enzymes are highly conserved and independent of environmental conditions. These identified differences and similarities of metabolic enzymes and metabolic substrates in the myocardium of Eurasian Tree Sparrows contribute to uncovering the coping mechanisms of physiological adjustments for adapting to the extreme conditions of the QTP. Still, how highland Eurasian Tree Sparrows can express hypoxic tolerance and orchestrate energy substrate utilization in other organs and what is the underlying regulatory mechanism remains to be further determined.
Availability of data and materials
The datasets used in the present study are available from the corresponding author on reasonable request.
References
Azevedo JL Jr, Carey JO, Pories WJ, Morris PG, Dohm GL. Hypoxia stimulates glucose transport in insulin-resistant human skeletal muscle. Diabetes. 1995;44:695–8.
Bagger JP, Thomassen A, Nielsen TT. Cardiac energy metabolism in patients with chest pain and normal coronary angiograms. Am J Cardiol. 2000;85:315–20.
Belitz HD, Grosch W, Schieberle P. Food chemistry. 4th ed. Berlin: Springer; 2009.
Blanchard DC, Spencer RL, Weiss SM, Blanchard RJ, McEwen B, Sakai RR. Visible burrow system as a model of chronic social stress: behavioral and neuroendocrine correlates. Psychoneuroendocrinology. 1995;20:117–34.
Brooks GA, Butterfield GE, Wolfe RR, Groves BM, Mazzeo RS, Sutton JR, et al. Increased dependence on blood glucose after acclimatization to 4,300 m. J Appl Physiol. 1991;70:919–27.
Brooks GA, Wolfel EE, Groves BM, Bender PR, Butterfield GE, Cymerman A, et al. Muscle accounts for glucose disposal but not blood lactate appearance during exercise after acclimatization to 4,300 m. J Appl Physiol. 1992;72:2435–45.
Burggren WW, Cameron JN. Anaerobic metabolism, gas exchange, and acid–base balance during hypoxic exposure in the channel catfish Ictalurus punctatus. J Exp Zool. 1980;213:405–16.
Burggren WW, Farrell AP, Lillywhite HB. Vertebrate cardiovascular systems. In: Terjung R, editor. Comprehensive physiology, Suppl. 30. Handbook of physiology, comparative physiology. Hoboken, NJ: Wiley-Blackwell; 2011.
Calder WA. Respiratory and heart rates of birds at rest. Condor. 1968;70:358–65.
Camici PG, Marraccini P, Lorenzoni R, Buzzigoli G, Pecori N, Perissinotto A, et al. Coronary hemodynamics and myocardial metabolism in patients with syndrome X: response to pacing stress. J Am Coll Cardiol. 1991;17:1461–70.
Cartee GD, Douen AG, Ramlal T, Klip A, Holloszy JO. Stimulation of glucose transport in skeletal muscle by hypoxia. J Appl Physiol. 1991;70:1593–600.
Casiday R, Frey R. Blood, sweat, and buffers: pH regulation during exercise, acid–base equilibria experiment. St. Louis: Washington University; 2012.
Cheviron ZA, Bachman GC, Connaty AD, McClelland GB, Storz JF. Regulatory changes contribute to the adaptive enhancement of thermogenic capacity in high-altitude deer mice. Proc Natl Acad Sci USA. 2012;109:8635–40.
Cheviron ZA, Natarajan C, Projecto-Garcia J, Eddy DK, Jones J, Carling MD, et al. Integrating evolutionary and functional tests of adaptive hypotheses: a case study of altitudinal differentiation in hemoglobin function in an Andean sparrow, Zonotrichia capensis. Mol Biol Evol. 2014;31:2948–62.
Durmowicz AG, Hofmeister S, Kadyraliev TK, Aldashev AA, Stenmark KR. Functional and structural adaptation of the yak pulmonary circulation to residence at high altitude. J Appl Physiol. 1993;74:2276–85.
Faraci FM, Kilgore DL Jr, Fedde MR. Attenuated pulmonary pressor response to hypoxia in bar-headed geese. Am J Physiol. 1984;247:R402–3.
Fuxe K, Cintra A, Andbjer B, Anggård E, Goldstein M, Agnati LF. Centrally administered endothelin-1 produces lesions in the brain of the male rat. Acta Physiol Scand. 1989;137:155–6.
Ge RL, Kubo K, Kobayashi T, Sekiguchi M, Honda T. Blunted hypoxic pulmonary vasoconstrictive response in the rodent Ochotona curzoniae (pika) at high altitude. Am J Physiol. 1998;274:H1792–9.
Ge RL, Simonson TS, Cooksey RC, Tanna U, Qin G, Huff CD, et al. Metabolic insight into mechanisms of high-altitude adaptation in Tibetans. Mol Genet Metab. 2012;106:244–7.
Gertz EW, Wisneski JA, Stanley WC, Neese RA. Myocardial substrate utilization during exercise in humans. Dual carbon-labeled carbohydrate isotope experiments. J Clin Invest. 1988;82:2017–25.
Green HJ, Sutton JR, Cymerman A, Young PM, Houston CS. Operation Everest II: adaptations in human skeletal muscle. J Appl Physiol. 1989;66:2454–61.
Groves BM, Droma T, Sutton JR, McCullough RG, McCullough RE, Zhuang J, et al. Minimal hypoxic pulmonary hypertension in normal Tibetans at 3,658 m. J Appl Physiol. 1985;1993(74):312–8.
Grubb BR. Allometric relations of cardiovascular function in birds. Am J Physiol. 1983;245:H567–72.
Hao Y, Xiong Y, Cheng Y, Song G, Jia C, Qu Y, et al. Comparative transcriptomics of 3 high-altitude passerine birds and their low-altitude relatives. Proc Natl Acad Sci USA. 2019;116:11851–6.
Hayward CS, Webb CM, Collins P. Effect of sex hormones on cardiac mass. Lancet. 2001;357:1354–6.
Hebisz R, Hebisz P, Borkowski J, Zatoń M. Differences in physiological responses to interval training in cyclists with and without interval training experience. J Hum Kinet. 2016;50:93–101.
Hochachka PW, Somero GN. Biochemical adaptation. Mechanism and process in physiological evolution. New York: Oxford University Press; 2002.
Holden JE, Stone CK, Clark CM, Brown WD, Nickles RJ, Stanley C, et al. Enhanced cardiac metabolism of plasma glucose in high-altitude natives: adaptation against chronic hypoxia. J Appl Physiol. 1995;79:222–8.
Horscroft JA, Kotwica AO, Laner V, West JA, Hennis PJ, Levett DZH, et al. Metabolic basis to Sherpa altitude adaptation. Proc Natl Acad Sci USA. 2017;114:6382–7.
Ingwall JS, Kramer MF, Fifer MA, Lorell BH, Shemin R, Grossman W, et al. The creatine kinase system in normal and diseased human myocardium. N Engl J Med. 1985;313:1050–4.
Ivy CM, Scott GR. Control of breathing and the circulation in high-altitude mammals and birds. Comp Biochem Phys A. 2015;186:66–74.
Jing XP, Wang WJ, Degen AA, Guo YM, Kang JP, Liu PP, et al. Energy substrate metabolism in skeletal muscle and liver when consuming diets of different energy levels: comparison between Tibetan and Small-tailed Han sheep. Animal. 2021;15:100162.
Kaur H, Parikh V, Sharma A, Singh M. Effect of amiloride a Na+/H+ exchange inhibitor on cardioprotective effect of ischaemic preconditioning: possible involvement of resident cardiac mast cells. Pharmacol Res. 1997;36:95–102.
Kodde IF, van der Stok J, Smolenski RT, de Jong JW. Metabolic and genetic regulation of cardiac energy substrate preference. Comp Biochem Phys A. 2007;146:26–39.
Kolar F, Ostadal B. Sex differences in cardiovascular function. Acta Physiol. 2013;207:584–7.
Koolhaas JM, Meerlo P, de Boer SF, Strubbe JH, Bohus B. Social stress in rats: an animal model of depression? Acta Neuropsychiatr. 1995;7:27–9.
Lagranha CJ, Deschamps A, Aponte A, Steenbergen C, Murphy E. Sex differences in the phosphorylation of mitochondrial proteins result in reduced production of reactive oxygen species and cardioprotection in females. Circ Res. 2010;106:1681–91.
Li D, Wu J, Zhang X, Ma X, Wingfield JC, Lei F, et al. Comparison of adrenocortical responses to acute stress in lowland and highland Eurasian Tree Sparrows (Passer montanus): similar patterns during the breeding, but different during the prebasic molt. J Exp Zool A Ecol Genet Physiol. 2011;315:512–9.
Lopaschuk GD, Ussher JR, Folmes CDL, Jaswal JS, Stanley WC. Myocardial fatty acid metabolism in health and disease. Physiol Rev. 2010;90:207–58.
Martinez M, Calvo-Torrent A, Pico-Alfonso MA. Social defeat and subordination as models of social stress in laboratory rodents: a review. Aggressive Behav. 1998;24:241–56.
Milerová M, Drahota Z, Chytilová A, Tauchmannová K, Houštěk J, Ošťádal B. Sex difference in the sensitivity of cardiac mitochondrial permeability transition pore to calcium load. Mol Cell Biochem. 2016;412:147–54.
Mónus F, Szabó K, Lózsa A, Pénzes Z, Barta Z. Intersexual size and plumage differences in tree sparrows (Passer montanus)—a morphologicals study based on molecular sex determination. Acta Zool Acad Sci H. 2011;57:269–77.
Murphy E. Estrogen signaling and cardiovascular disease. Circ Res. 2011;109:687–96.
Neglia D, de Caterina A, Marraccini P, Natali A, Ciardetti M, Vecoli C, et al. Impaired myocardial metabolic reserve and substrate selection flexibility during stress in patients with idiopathic dilated cardiomyopathy. Am J Physiol Heart Circ Physiol. 2007;293:H3270–8.
Pilarski JQ, Solomon IC, Kilgore DL Jr, Hempleman SC. Effects of aerobic and anaerobic metabolic inhibitors on avian intrapulmonary chemoreceptors. Am J Physiol Regul Integr Comp Physiol. 2009;296:R1576–84.
Pulinilkunnil T, Rodrigues B. Cardiac lipoprotein lipase: metabolic basis for diabetic heart disease. Cardiovasc Res. 2006;69:329–40.
Qiu Q, Zhang G, Ma T, Qian W, Wang J, Ye Z, et al. The yak genome and adaptation to life at high altitude. Nat Genet. 2012;44:946–9.
Qu Y, Zhao H, Han N, Zhou G, Song G, Gao B, et al. Ground tit genome reveals avian adaptation to living at high altitudes in the Tibetan plateau. Nat Commun. 2013;4:2071.
Qu Y, Tian S, Han N, Zhao H, Gao B, Fu J, et al. Genetic responses to seasonal variation in altitudinal stress: whole-genome resequencing of great tit in eastern Himalayas. Sci Rep. 2015;5:14256.
Qu Y, Chen C, Xiong Y, She H, Zhang YE, Cheng Y, et al. Rapid phenotypic evolution with shallow genomic differentiation during early stages of high elevation adaptation in Eurasian Tree Sparrows. Natl Sci Rev. 2020;7:113–27.
Roberts AC, Butterfield GE, Cymerman A, Reeves JT, Wolfel EE, Brooks GA. Acclimatization to 4,300-m altitude decreases reliance on fat as a substrate. J Appl Physiol. 1996a;81:1762–71.
Roberts AC, Reeves JT, Butterfield GE, Mazzeo RS, Sutton JR, Wolfel EE, et al. Altitude and beta-blockade augment glucose utilization during submaximal exercise. J Appl Physiol. 1996b;80:605–15.
Sakai A, Matsumoto T, Saitoh M, Matsuzaki T, Koizumi T, Ishizaki T, et al. Cardiopulmonary hemodynamics of blue-sheep, Pseudois nayaur, as high-altitude adapted mammals. Jpn J Physiol. 2003;53:377–84.
Samaja M, Mariani C, Prestini A, Cerretelli P. Acid–base balance and O2 transport at high altitude. Acta Physiol Scand. 1997;159:249–56.
Scott GR, Milsom WK. Control of breathing and adaptation to high altitude in the bar-headed goose. Am J Physiol-Reg. 2007;I(293):R379–91.
Scott GR, Schulte PM, Egginton S, Scott AL, Richards JG, Milsom WK. Molecular evolution of cytochrome C oxidase underlies high-altitude adaptation in the bar-headed goose. Mol Biol Evol. 2011;28:351–63.
Sears MW, Hayes JP, O’Connor CS, Geluso K, Sedinger JS. Individual variation in thermogenic capacity affects above-ground activity of high-altitude Deer Mice. Funct Ecol. 2006;20:97–104.
Shao Y, Li JX, Ge RL, Zhong L, Irwin DM, Murphy RW, et al. Genetic adaptations of the plateau zokor in high-elevation burrows. Sci Rep. 2015;5:17262.
Storz JF, Moriyama H. Mechanisms of hemoglobin adaptation to high altitude hypoxia. High Alt Med Biol. 2008;9:148–57.
Storz JF, Scott GR, Cheviron ZA. Phenotypic plasticity and genetic adaptation to high-altitude hypoxia in vertebrates. J Exp Biol. 2010;213:4125–36.
Summers-Smith D. Eurasian tree sparrow (Passer montanus). In: del Hoyo J, Elliott A, Sargatal J, Christie DA, de Juana E, editors. Handbook of the birds of the world alive. Barcelona: Lynx Edicions; 2018. p. 217–44.
Sun YF, Ren ZP, Wu YF, Lei FM, Dudley R, Li DM. Flying high: limits to flight performance by sparrows on the Qinghai-Tibet Plateau. J Exp Biol. 2016;219:3642–8.
Sun YF, Li M, Song G, Lei FM, Li DM, Wu YF. The role of climate factors in geographic variation in body mass and wing length in a passerine bird. Avian Res. 2017;8:1.
Sutton JR, Jones NL, Griffith L, Pugh CE. Exercise at altitude. Annu Rev Physiol. 1983;45:427–37.
Tate KB, Ivy CM, Velotta JP, Storz JF, McClelland GB, Cheviron ZA, et al. Circulatory mechanisms underlying adaptive increases in thermogenic capacity in high-altitude deer mice. J Exp Biol. 2017;220:3616–20.
Thompson LG, Yao T, Mosley-Thompson E, Davis ME, Henderson KA, Lin P-N. A high-resolution millennial record of the south Asian monsoon from Himalayan ice cores. Science. 2000;289:1916–9.
Vaillancourt E, Haman F, Weber JM. Fuel selection in Wistar rats exposed to cold: shivering thermogenesis diverts fatty acids from re-esterification to oxidation. J Physiol. 2009;587:4349–59.
van der Vusse GJ, Glatz JF, Stam HC, Reneman RS. Fatty acid homeostasis in the normoxic and ischemic heart. Physiol Rev. 1992;72:881–940.
Ventura-Clapier R, Garnier A, Veksler V. Energy metabolism in heart failure. J Physiol. 2004;555:1–13.
Vinet A, Mandigout S, Nottin S, Nguyen LD, Lecoq AM, Courteix D, et al. Influence of body composition, hemoglobin concentration, and cardiac size and function of gender differences in maximal oxygen uptake in prepubertal children. Chest. 2003;124:1494–9.
Wang X, Hole DG, da Costa TH, Evans RD. Alterations in myocardial lipid metabolism during lactation in the rat. Am J Physiol. 1998;275:E265–71.
Weber JM. Metabolic fuels: regulating fluxes to select mix. J Exp Biol. 2011;214:286–94.
Young AJ, Evans WJ, Cymerman A, Pandolf KB, Knapik JJ, Maher JT. Sparing effect of chronic high-altitude exposure on muscle glycogen utilization. J Appl Physiol. 1982;52:857–62.
Zhu X, Guan Y, Signore AV, Natarajan C, DuBay SG, Cheng Y, et al. Divergent and parallel routes of biochemical adaptation in high-altitude passerine birds from the Qinghai-Tibet Plateau. Proc Natl Acad Sci USA. 2018;115:1865–70.
Zinker BA, Namdaran K, Wilson R, Lacy DB, Wasserman DH. Acute adaptation of carbohydrate metabolism to decreased arterial PO2. Am J Physiol. 1994;266:E921–9.
Acknowledgements
We thank Zhipeng Ren, Yinchao Hao, Simeng Yu, and Haiqing He for their assistance with sample collection in the field.
Funding
This work was supported by the National Natural Science Foundation of China (NSFC, No. 31971413) to DL and NSFC (No. 31770445) to YW, the Second Tibetan Plateau Scientific Expedition and Research Program (STEP, 2019QZKK0501), and the Natural Science Foundation of Hebei Province (NSFHB, C2020205038) to DL, the Foundation of Hebei Normal University (L2019B26) to CJ, and the Post-doctoral Research Programm to PD.
Author information
Authors and Affiliations
Contributions
BD and YZ: methodology, validation; YS and ML: fieldwork and investigation; GN and YW: help of writing original draft; QZ, laboratory assays; CJ: conceptualization, supervision; DL: conceptualization, supervision, funding acquisition. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The present study complies with the current laws of China. Fieldwork was carried out under permission from the Department of Wildlife Conservation (Forestry Bureau) of Hebei Province and Qinghai Province. Experimental procedures were permitted by the approval of the Institutional Animal Care and Use Committee (HEBTU2013-7) and the Ethics and Animal Welfare Committee (No. 2013-6) of Hebei Normal University, China.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ding, B., Zhao, Y., Sun, Y. et al. Coping with extremes: lowered myocardial phosphofructokinase activities and glucose content but increased fatty acids content in highland Eurasian Tree Sparrows. Avian Res 12, 44 (2021). https://doi.org/10.1186/s40657-021-00279-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40657-021-00279-0