
Abstract
Background
The capacity for thermogenesis is considered part of an animal’s adaptive strategy for survival, and basal metabolic rate (BMR) is one of the fundamental physiological standards for assessing the energy cost of thermoregulation in endotherms. BMR has been shown to be a highly flexible phenotypic trait both between, and within, species, but the metabolic mechanisms involved in the regulation of BMR, which range from variation in organ mass to biochemical adjustments, remain unclear. In this study, we investigated the relationship between organ mass, biochemical markers of metabolic tissue activity, and thermogenesis, in three species of small passerines: wild Bramblings (Fringilla montifringilla), Little Buntings (Emberiza pusilla) and Eurasian Tree Sparrows (Passer montanus), caught in Wenzhou, southeastern China.
Methods
Oxygen consumption was measured using an open-circuit respirometry system. Mitochondrial state-4 respiration and cytochrome c oxidase (COX) activity in liver and pectoral muscle were measured with a Clark electrode.
Results
Our results show that Eurasian Tree Sparrows had significantly higher BMR, digestive organ mass, mitochondrial state-4 respiration capacity and COX activity in liver and muscle, than Bramblings and Little Buntings. Furthermore, interspecific differences in BMR were strongly correlated with those indigestive tract mass, state-4 respiration and COX activity.
Conclusions
Our findings suggest that the digestive organ mass, state-4 respiration and COX activity play an important role in determining interspecific differences in BMR.
Similar content being viewed by others

Background
Adaptive variation in thermogenic capacity is critical to the survival of small birds in temperate zones (Schmidt-Nielsen 1997; Weathers 1997). To minimize the energetic cost of thermoregulation, birds use a variety of morphological and behavioral traits to adjust their rates of heat loss and heat gain, ranging from biochemical adjustments to changes in internal and whole organ mass (McKechnie 2008; Swanson 2010; Zheng et al. 2014a). Basal metabolic rate (BMR) refers to the energy expenditure of an animal at rest (i.e. thermoneutrality) during the inactive phase of the day, when it is not processing food, moulting, or reproducing (McNab 2009), and is one of the fundamental physiological standards for assessing the energetic cost of thermoregulation (McKechnie et al. 2006). BMR has been referred to as a fundamental energetic trait, in large part because it represents a fixed cost that all organisms must incur (Furness 2003). BMR can consume as much as 50–60 % of daily energy expenditure (DEE) and variation in BMR may be associated with peak, or sustained, metabolic rates, species richness and distribution, activity levels and life-history strategies (McKinney and McWilliams 2005; Wells and Schaeffer 2012). BMR is therefore an important parameter for both inter- and intraspecific comparisons of thermoregulatory ability (McKechnie et al. 2006; McNab 2009). To date, BMR has been measured in more than 500 species of birds (McKechnie 2008; McNab 2009; Smit and McKechnie 2010). Those studies show that BMR is highly flexible both between and within species (Wikelski et al. 2003; Wiersma et al. 2007; Zheng et al. 2014a). The typically lower BMR of tropical birds compared to that of their high-latitude counterparts has been explained as an adaptation to avoid heat stress and to conserve water (Wiersma et al. 2007). In turn, the higher BMR of temperate birds has been explained as a direct or indirect result of adaptation to a colder climate and a shorter breeding season, both of which would be expected to require a higher level of metabolic activity (Swanson 2010; Zheng et al. 2014b). It has been suggested that the BMR of birds that migrate to tropical latitudes in winter, but breed in colder, temperate latitudes in summer, is lower in winter than in summer (Lindström and Klaassen 2003; Zheng et al. 2013a).
What are the metabolic mechanisms underlying variation in BMR? McKechnie (2008) and Swanson (2010) identified major physiological and morphological pathways whereby metabolic rates are up- or down-regulated, namely, adjustments in organ mass, adjustments in the mass-specific metabolic intensities of specific organs, and adjustments in the transport capacities for oxygen and metabolic substrates (Zheng et al. 2014b). At the organism level, the mechanism that has received the most attention has been the positive relationship between body mass (M b) adjusted organ size and BMR (Daan et al. 1990; Chappell et al. 1999; Hammond et al. 2000). Although they represent less than 10 % of M b, internal organs such as the liver, kidneys, heart and digestive tract can contribute more than 60 % of the energy expended at the basal level (Rolfe and Brown 1997; Clapham 2012). The liver is one of the largest and most metabolically active organs in endotherms, and, under basal metabolic conditions, may contribute 25 % of total heat production (Villarin et al. 2003; Zheng et al. 2008a). Skeletal muscles have lower, mass-specific, resting metabolic rates than many central organs (Scott and Evans 1992). However, due to their large total mass, they may contribute significantly to seasonal metabolic acclimatization (Chappell et al. 1999; Zheng et al. 2008b, 2014a). At the physiological and biochemical level, changes in activities of catabolic enzymes could influence the mass-specific metabolic intensities of organs, thereby affecting BMR (Liknes and Swanson 2011; Zheng et al. 2014a). Such variation in cellular metabolic intensity is often measured by examining variation in state-4 respiration (Zheng et al. 2008b, 2013a), citrate synthase (CS) activity (Swanson 2010; Swanson et al. 2014), or cytochrome coxidase (COX) activity. CS plays a key role in the Krebs cycle whereas state-4 respiration and COX are important in oxidative phosphorylation (Zheng et al. 2014a, b). The adaptive changes that produce higher BMR in small birds are thought to have a cellular or molecular basis and levels of state-4 respiration and COX activity have been commonly used as enzymatic indicators of variation in BMR at the cellular level (Zheng et al. 2008b, 2014a; Zhou et al. 2016).
Bramblings (Fringilla montifringilla), Little Buntings (Emberiza pusilla) and Eurasian Tree Sparrows (Passer montanus) inhabit vast areas of Europe and Asia (MacKinnon and Phillipps 2000). Bramblings and Little Buntings are migratory, wintering in southern Europe, northern India, and China, whereas Eurasian Tree Sparrows are resident in China. Bramblings and Little Buntings migrate to Wenzhou only during spring and autumn migration periods (Liu et al. 2001; Zheng et al. 2013a). The Brambling and Little Bunting have relatively higher body temperatures and metabolic rates than expected based on their body masses and broad thermal neutral zones, and relatively lower critical temperatures (Liu et al. 2001, 2004). The Eurasian Tree Sparrow increases its thermogenic capacity in cold conditions mainly by increasing both respiratory enzyme activity and the level of plasma thyroid hormones (Liu et al. 2008; Zheng et al. 2008a, 2014b). The capacity to make these metabolic adjustments may be the key for this species being able to survive in relatively cold areas (Liu et al. 2004, 2008). The present study is a continuation of investigation into the thermogenic capacities of these species. We hypothesized that species-specific physiological and biochemical metabolic characteristics would contribute to interspecific variation in BMR. We predicted that species with relatively high BMR would have higher organ mass, mitochondrial respiration capacity and COX activity. In this study we tested this hypothesis by comparing BMR, organ mass and selected biochemical markers of metabolic cellular activity, in these three species.
Methods
Animals
Seven Bramblings and eight Little Buntings were live-trapped in forested parts of Wenzhou, Zhejiang Province (27°29′N, 120°51′E) in China during the spring migration period in 2011, and ten Eurasian Tree Sparrows were captured at the same time. The Wenzhou climate is warm-temperate with an average annual rainfall of 1700 mm spread across all months with slightly more precipitation during winter and spring. Mean daily maximum temperatures range from 39 °C in July to 8 °C in January (Zheng et al. 2008a, 2014a). Body mass (M b) to the nearest 0.1 g was determined immediately upon capture with a Sartorius balance (model BT25S). Bramblings, buntings and sparrows were transported to the laboratory and caged for 1 or 2 d (50 cm × 30 cm × 20 cm) outdoors under natural photoperiod (about 14:10 hours light:dark photoperiod) and temperature (18 °C) before measurements. Food and water were supplied ad libitum (Zhou et al. 2016). All experimental procedures were approved by the Animal Care and Use Committee of the Wenzhou City, Zhejiang Province, China (Wu et al. 2015; Zhou et al. 2016).
Measurement of metabolic rate
We measured oxygen consumption using an open-circuit respirometry system (S-3A/I, AEI technologies, Pittsburgh, PA, USA) (Zheng et al. 2014a). We provided a perch in respirometry chamber and allowed individual birds to rest in the 1.5-L metabolic chamber before measuring their metabolic rate (Smit and McKechnie 2010). The metabolic chamber was housed in a temperature-controlled cabinet capable of regulating temperature to ±0.5 °C (Artificial Climatic Engine BIC-300, Shanghai, China). H2O and CO2 were scrubbed from the air with a silica gel/soda lime/silica column before and after it passed through the metabolic chamber. We determined the fractional concentrations of oxygen in the inlet and outlet chamber air with an oxygen sensor (AEI technologies N-22M, USA). During the measurement of metabolic rates, we pumped dry CO2-free air through the chamber at 300 mL/min with a flow control system (AEI technologies R-1, USA) calibrated to ±1 % accuracy with a general purpose thermal mass flow-meter (TSI 4100 Series, USA), to maintain the fractional concentration of O2 in the chamber at about 20 % (McNab 2006). We obtained the baseline O2 concentration before and after each test (Li et al. 2010; Wu et al. 2015). We measured oxygen consumption rates at 30 ± 0.5 °C, which is within the thermal neutral zone for Bramblings, Little Buntings and Eurasian Tree Sparrows (Zheng et al. 2008b, 2014b). We obtained all measurements of gas exchange during the rest-phase of birds’ circadian cycles (between 20:00 and 04:00 hours) in dark chambers. We removed food 4 h before each test to create post-absorptive conditions. Measurement of oxygen consumption commenced when birds were observed perching calmly in the chamber and continued for 1 h. In general, each animal was in the metabolic chamber for at least 2 h. The oxygen consumption data were recorded every minute according to the equation 2 described by Hill (1972). We took the lowest 5 min mean oxygen consumption data over the test period to calculate BMR (Wu et al. 2015; Zhou et al. 2016). We expressed metabolic rates as mL O2/h after correcting all values to standard temperature, pressure, and dry gas (STPD) conditions (Schmidt-Nielsen 1997). We measured body temperature during metabolic measurements using a lubricated thermocouple inserted in the cloaca, and digitized the output using a thermocouple meter (Beijing Normal University Instruments Co.). We measured M b to the nearest 0.1 g before and after the experiments, and used mean M b in calculations. All measurements were taken daily between 20:00 and 04:00 hours.
Measurement of organ mass
Birds were euthanized by cervical dislocation at the end of the experiment and their pectoral muscle, heart, liver, kidneys, gizzard, small intestine and rectum extracted and weighed to the nearest 0.1 mg. Part of the muscle and liver was used to investigate state-4 respiration and COX activity (Zheng et al. 2008b, 2014a), and the other internal organs, including the remainder of the muscle and liver, were dried to a constant mass over 2 days at 65 °C, and weighed to the nearest 0.1 mg (Williams and Tieleman 2000; Liu and Li 2006; Wu et al. 2014).
Preparation of mitochondria
Liver and pectoral muscle sub-samples were placed in ice-cold, sucrose-buffered medium, cleaned of any adhering tissue, blotted, and weighed. We chopped liver samples coarsely with scissors, then rinsed and resuspended them in 5 volumes of ice-cold medium (Rasmussen et al. 2004). Pectoral muscle samples were coarsely chopped with scissors, and treated with proteinase for 5–10 min, after which the proteinase was removed and the muscle samples were resuspended in 10 volumes of ice-cold medium. Both liver and muscle preparations were homogenized with a Teflon/glass homogenizer. Homogenates were centrifuged at 600×g for 10 min at 4 °C in an Eppendorf centrifuge, and the pellets containing nuclei and cellular debris discarded. Supernatants were centrifuged at 12,000×g for 10 min at 4 °C. The resultant pellets were suspended, respun at 12,000×g, and resuspended (2:1, w/v for liver and 4:1 for muscle) in ice-cold medium (Zheng et al. 2013b). We determined the protein content of mitochondria by the Folin phenol method with bovine serum albumin as standard (Lowry et al. 1951).
Mitochondrial respiration and enzyme activity
Mitochondrial state-4 respiration in liver and pectoral muscle was measured at 30 °C in 1.96 mL of respiration medium with a Clark electrode (Hansatech Instruments LTD., England, DW-1), essentially as described by Estabrook (1967). State-4 respiration was measured over a 1 h period under substrate dependent conditions, with succinate as the substrate (Zheng et al. 2014a). State-4 respiration was expressed as mean mass-specific level [µmol O2/(min g tissue)] (Zheng et al. 2013a). Cytochrome c oxidase (COX) activity in the liver and pectoral muscle was measured polarographically at 30 °C using a Clark electrode according to Sundin et al. (1987). Enzyme activity was reported as mean mass-specific level [µmol O2/(min g tissue)] (Zheng et al. 2013b, 2014a).
Statistics
Statistical analyses were performed using the SPSS package (version 12.0). All variables were tested for normality with the Kolmogorov–Smirnov test before statistical tests were performed. Non-normal data were normalized by transforming them to their natural logarithms before conducting statistical tests. M b among different groups was compared using a one-way ANOVA. The significance of differences in BMR and organ mass was determined with a one-way ANCOVA with M b as a covariate. We used Tukey’s HSD post hoc test to determine which species differed significantly from others. The statistical significance of differences in mitochondrial protein, mitochondrial state-4 respiration and COX activity in the liver and muscle was tested with a one-way ANOVA. Least-squares linear regression was used to evaluate the relationship between log BMR and log M b, and between log BMR, log state-4 respiration and log COX. Data are reported as mean ± SE, unless otherwise noted. The p values <0.05 were considered statistically significant.
Results
Body mass (M b) and basal metabolic rate
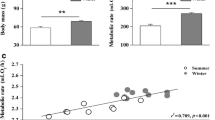
There were significant differences in M bs among the three species (F 2,22 = 21.303, p < 0.001; Fig. 1A; Table 1). There were also significant differences in BMR (mL O2/h) among the three species (F 2,22 = 26.772, p < 0.001; Fig. 1B); mean BMR was significantly higher in Eurasian Tree Sparrows than in Bramblings (19.0 %) and Little Buntings (74.4 %). Corrected for M b, BMR still differed significantly among the three species (F 2,21 = 5.402, p < 0.05, Table 1). There was a positive correlation between M b and BMR (r = 0.768, p < 0.001; Fig. 1C). No significant differences were found between the three species in body temperature (data not shown).

Comparison of M b (A), basal metabolic rate (B) and the relationship between log M b and log basal metabolic rate (C) among Bramblings (Fringilla montifringilla), Little Buntings (Emberiza pusilla) and Eurasian Tree Sparrows (Passer montanus). Data are shown as mean ± SE, bars with different letters are significantly different
Organ and muscle mass
Gizzard mass differed significantly among the three species (F 2,21 = 8.852, p < 0.01; Table 1). The mean gizzard mass of Eurasian Tree Sparrows was heavier than that of Bramblings, but there was no significant difference in gizzard mass between Eurasian Tree Sparrows and Little Buntings, or between that of Little Buntings and Bramblings. The three species also differed significantly in rectal mass (F 2,21 = 5.815, p < 0.01; Table 1). Eurasian Tree Sparrows and Bramblings had a higher average rectal mass than Little Buntings, but there was no significant difference in rectal mass between Eurasian Tree Sparrows and Bramblings. Eurasian Tree Sparrows had a higher average total digestive tract mass than Bramblings and Little Buntings (F 2,21 = 21.358, p < 0.001), but there was no significant difference in this variable between the latter two species (p > 0.05). No significant between-species differences were apparent in the dry mass of the heart, liver, kidneys, small intestine, or muscle (Table 1). Partial correlations between log organ mass and log M b were positive for all organs, and the dry mass of the heart, liver, kidneys, gizzard, total digestive tract, and muscle, were significantly correlated with M b (Table 2). For each of these organs, the slopes of the respective regression lines exceeded 1.0, indicating that organ mass increased with body size at a faster rate than overall M b (Table 2). BMR residuals were only significantly, positively correlated with the total digestive tract dry mass residuals (Table 2).
Protein content, mitochondrial state-4 respiration, and cytochrome c oxidase (COX) activity in liver and muscle
There were no significant interspecific differences in the protein content of different organs (liver, F 2,22 = 1.007, p > 0.05; muscle, F 2,22 = 0.360, p > 0.05; Fig. 2A), but Eurasian Tree Sparrows had higher mitochondrial state-4 respiration (liver, F 2,22 = 4.374, p < 0.05; muscle, F 2,22 = 15.108, p < 0.001; Fig. 2B) and COX activity (liver, F 2,22 = 9.615, p < 0.01; muscle, F 2,22 = 8.492, p < 0.01; Fig. 2C) than Bramblings and Little Buntings. No significant differences were found in these variables between the latter two species (p > 0.05). Log BMR was positively correlated with log COX activity in the liver (r = 0.388, p < 0.05; Fig. 3b), log state-4 respiration (r = 0.568, p < 0.01; Fig. 3c) and log COX activity (r = 0.548, p < 0.01; Fig. 3d) in muscle.

Differences in mitochondrial protein (A), state-4 respiration (B), and cytochrome c oxidase (C) in the liver and pectoral muscle among Bramblings (Fringilla montifringilla), Little Buntings (Emberiza pusilla) and Eurasian Tree Sparrows (Passer montanus). Data are shown as mean ± SE, bars with different letters are significantly different

Correlations between log metabolic rate and state-4 respiration in the liver (a), and cytochrome c oxidase (COX) activity in the liver (b), state-4respiration in pectoral muscle (c) and cytochrome c oxidase (COX) activity in pectoral muscle (d), among Bramblings (Fringilla montifringilla), Little Buntings (Emberiza pusilla) and Eurasian Tree Sparrows (Passer montanus)
Discussion
The results of this study indicate significant differences in organ mass, and some biochemical markers of metabolic tissue activity, among the three species, which could partly account for the observed interspecific differences in BMR (Guderley et al. 2005).
Comparison of internal organ mass and thermogenic properties
With respect to metabolic traits, the BMR of an animal is the sum of the metabolic rates of its organs and other metabolically active tissues (Zheng et al. 2008a; Swanson 2010; Clapham 2012). The selective pressures that influence metabolism may, however, be complex and act on metabolic rate through multiple avenues. Two of these potential avenues are to alter the sizes of tissues or organs and to alter the density of mitochondria and the concentration of enzymes in aerobic catabolic pathways (Brand et al. 2003; Else et al. 2004). However, it is not clear whether large, energetically expensive organs are responsible for higher BMR, or whether they are necessary to support a higher BMR. Thus, the relationship between BMR and organ mass remains purely correlative, which is cause and which is effect remains unresolved (Steyermark et al. 2005). What are the ecological and evolutionary implications of having larger visceral organs for higher BMR birds? It has been suggested that much of the energy used in basal metabolism is consumed by the visceral organs (Daan et al. 1990; Piersma et al. 1996). Williams and Tieleman (2000) hypothesized that natural selection adjusts the size of the internal organs to match energy requirements, and that body size independent variation in BMR reflects the relative size of internal organs. These include the digestive tract, which performs digestion and absorption, the heart, which transports oxygen to the tissues, the liver which performs catabolism, and the kidneys, which eliminate nitrogenous and other wastes (Kersten and Piersma 1987; Daan et al. 1990; Hammond et al. 2001). We found no significant interspecific differences in heart, liver, kidneys, or muscle mass, and consequently no evidence to support the hypothesis that the mass of these organs should be greater in species with higher BMR. However, compared to Bramblings and Little Buntings, Eurasian Tree Sparrows had a heavier gizzard, rectum and total digestive tract. These findings suggest that the mass of the digestive organs could be related to the observed between-species differences in BMR. The ecological implications of having a larger digestive tract are increased food consumption, which could, in turn, stimulate the enlargement of organs such as the gizzard and small intestine (Zheng et al. 2008b; Lv et al. 2014). For example, Zheng et al. (2013b) acclimated Chinese Bulbuls (Pycnonotus sinensis) to either 10 or 30 °C for 4 weeks, measured their BMR, and then determined the dry mass of their internal organs. Bulbuls acclimated to 10 °C had a significantly higher BMR, and a markedly larger liver and intestine than those acclimated to 30 °C. Eurasian Tree Sparrows also had a significantly higher BMR in winter than in summer, and had a larger liver, smaller intestine and entire digestive tract compared to birds examined in summer (Liu and Li 2006; Zheng et al. 2008b). Changes in the size of digestive organs in response to elevated daily energy intake could therefore result in elevated BMR (Williams and Tieleman 2000).
Comparison of biochemical indices and thermogenic properties
Interspecific differences in metabolic intensity are linked with differences in mitochondrial densities, oxidative capacities and mitochondrial proton leaks (Else et al. 2004; Guderley et al. 2005). A strong correlation between metabolic rate, mitochondrial respiration, and proton leaks has been reported (Brookes et al. 1998; Li et al. 2010). The liver is one of the largest, and most metabolically active, organs in endotherms, and is considered to make an important contribution to BMR (Villarin et al. 2003; Zheng et al. 2008b). Mechanisms of heat generation in the liver include the uncoupling of oxidative phosphorylation, futile cycling of substrates and high mass-specific metabolic intensity (Brand et al. 2003; Zheng et al. 2014a). For example, Else et al. (2004) compared the respiration rate of hepatocytes in five birds and found that these approximated the basal metabolic rate–body mass relationship. Similar results have also been obtained in small mammals. For example, in addition to higher BMR, Brandt’s Voles (Lasiopodomys brandtii) also had higher mitochondrial state-4 respiration capacity and COX activity in the liver than Mongolian Gerbils (Meriones unguiculatus), suggesting that there is a relationship between these metabolic process and BMR (Li et al. 2010). In the present study, we found significant interspecific differences in state-4 respiration and COX activity in the liver, and significant, positive correlations between BMR and COX activity. These results suggest that the higher metabolic activity in the liver of Eurasian Tree Sparrows may contribute to the observed interspecific differences in BMR. This finding is in agreement with the results of our previous studies which show that seasonal and latitudinal variation in Eurasian Tree Sparrows was correlated not only with variation in BMR, but also in state-4 respiration and COX activity in the liver (Zheng et al. 2008b, 2014b).
Because skeletal muscle mass comprises nearly 40 % of M b, it is an important contributor to thermogenesis via shivering, and even nonshivering thermogenesis (Bicudo et al. 2001; Pitit and Vézina 2014). Furthermore, adjustment of cellular aerobic capacity in muscle potentially involves modulation of the activities of key catabolic enzymes in oxidative pathways, and, or, the activities of enzymes and transporters involved in substrate mobilization and delivery pathways (Marsh et al. 1990; Swanson 2010; Zheng et al. 2008b, 2014a). The results of this study demonstrate that Eurasian Tree Sparrows had higher mitochondrial state-4 respiration and COX activity than Bramblings and Little Buntings, and that there was a positive relationship between BMR, state-4 respiration and COX activity in these three species. This suggests that biochemical metabolic markers may be useful indicators of interspecific variation in BMR.
Conclusions
The selective pressures that influence metabolism may be complex and influence metabolic rate via multiple avenues. Our results show that Eurasian Tree Sparrows had significantly higher BMR, digestive organ mass, mitochondrial state-4 respiration capacity and COX activity in the liver and muscle, than Bramblings and Little Buntings. This suggests that digestive organ mass and the above biochemical markers of metabolic activity are both strongly correlated with BMR in these species, and play an important role in the determination of BMR. Future studies could add to these results by measuring thyroid hormones (thyroxine and triiodothyronine), which affect adaptive thermogenesis by substrate cycling, ion cycling, and mitochondrial proton leakage (Yen 2001; Liu et al. 2006; Mullur et al. 2014). Additional avenues for further research on the mechanisms underlying BMR variations include quantifying inter- and intraspecific variation in avian uncoupling protein (avUCP), proton conductance, and myostatin, all of which can affect the basal thermogenesis of tissues (Dridi et al. 2004; Swanson 2010).
References
Bicudo JE, Vianna CR, Chaui-Berlinck JG. Thermogenesis in birds. Biosci Rep. 2001;21:181–8.
Brand MD, Turner N, Ocloo A, Else PL, Hulbert AJ. Proton conductance and fatty acyl composition of liver mitochondria correlates with body mass in birds. Biochem J. 2003;376:741–8.
Brookes PS, Buckingham JA, Tenreiro AM, Hulbert AJ, Brand MD. The proton permeability of the inner membrane of liver mitochondria from ectothermic and endothermic vertebrates and from obese rats: correlations with standard metabolic rate and phospholipid fatty acid composition. Comp Biochem Phys B. 1998;119:325–34.
Chappell MA, Bech C, Buttemer WA. The relationship of central and peripheral organ masses to aerobic performance variation in house sparrows. J Exp Biol. 1999;202:2269–79.
Clapham JC. Central control of thermogenesis. Neuropharmacology. 2012;63:111–23.
Daan S, Masman D, Groenewold A. Avian basal metabolic rates: their association with body composition and energy expenditure in nature. Am J Physiol. 1990;259:R333–40.
Dridi S, Onagbesan O, Swennen Q, Buyse J, Decuypere E, Taouis M. Gene expression, tissue distribution and potential physiological role of uncoupling protein in avian species. Comp Biochem Phys A. 2004;139:273–83.
Else PL, Brand MD, Turner N, Hulbert AJ. Respiration rate of hepatocytes varies with body mass in birds. J Exp Biol. 2004;207:2305–11.
Estabrook RW. Mitochondrial respiratory control and the polarographic measurement of ADP: O ratio. Method Enzymol. 1967;10:41–7.
Furness RW. It’s in the genes. Nature. 2003;425:479–80.
Guderley H, Turner N, Else PL, Hulbert AJ. Why are some mitochondria more powerful than others: insights from comparisons of muscle mitochondria from three terrestrial vertebrates. Comp Biochem Phys A. 2005;142:172–80.
Hammond KA, Chappell MA, Cardullo RA, Lin RS, Johnsen TS. The mechanistic basis of aerobic performance variation in red jungle fowl. J Exp Biol. 2000;203:2053–64.
Hammond KA, Szewczak J, Krόl E. Effects of altitude and temperature on organ phenotypic plasticity along an altitudinal gradient. J Exp Biol. 2001;204:1991–2000.
Hill RW. Determination of oxygen consumption by use of the paramagnetic oxygen analyzer. J Appl Physiol. 1972;33:261–3.
Kersten M, Piersma T. High levels of energy expenditure in shorebirds; metabolic adaptations to an energetically expensive way of life. Ardea. 1987;75:175–87.
Li YG, Yang ZC, Wang DH. Physiological and biochemical basis of basal metabolic rates in Brandt’s voles (Lasiopodomys brandtii) and Mongolian gerbils (Meriones unguiculatus). Comp Biochem Phys A. 2010;157:204–11.
Liknes ET, Swanson DL. Phenotypic flexibility of body composition associated with seasonal acclimatization in passerine birds. J Ther Biol. 2011;36:363–70.
Lindström Å, Klaassen M. High basal metabolic rates in shorebirds: a circumpolar view. Condor. 2003;105:420–7.
Liu JS, Li M. Phenotypic flexibility of metabolic rate and organ masses among tree sparrows Passer montanus in seasonal acclimatization. Acta Zool Sin. 2006;52:469–77.
Liu JS, Zhang ZY, Ma H, Hou ZS. Characteristics of resting metabolic rate in little bunting (Emberiza pusilla) and chestnut bunting (E. rutila). Acta Zool Sin. 2001;47:347–50.
Liu JS, Wang DH, Wang Y, Chen MH, Song CG, Sun RY. Energetics and thermoregulation of the Carpodacus roseus, Fringilla montifringilla and Acanthis flammea. Acta Zool Sin. 2004;50:357–63.
Liu JS, Chen YQ, Li M. Thyroid hormones increase liver and muscle thermogenic capacity in the little buntings (Emberiza pusilla). J Therm Biol. 2006;31:386–93.
Liu JS, Li M, Shao SL. Seasonal changes in thermogenic properties of liver and muscle in tree sparrows Passer montanus. Acta Zool Sin. 2008;54:777–84.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with Folin phenol reagent. J Biol Chem. 1951;193:265–75.
Lv JW, Xie ZL, Sun YR, Sun CR, Liu LR, Yu TF, Xu XJ, Shao SL, Wang CH. Seasonal plasticity of duodenal morphology and histology in Passer montanus. Zoomorphology. 2014;133:435–43.
MacKinnon J, Phillipps K. A field guide to the birds of China. London: Oxford University Press; 2000.
Marsh RL, Dawson WR, Camilliere JJ, Olson JM. Regulation of glycolysis in the pectoralis muscles of seasonally acclimatized American goldfinches exposed to cold. Am J Physiol. 1990;258:R711–7.
McKechnie AE. Phenotypic flexibility in basal metabolic rate and the changing view of avian physiological diversity: a review. J Comp Phys B. 2008;178:235–47.
McKechnie AE, Freckleton RP, Jetz W. Phenotypic plasticity in the scaling of avian basal metabolic rate. Proc R Soc B Biol Sci. 2006;273:931–7.
McKinney RA, McWilliams SR. A new model to estimate daily energy expenditure for wintering waterfowl. Wilson Bull. 2005;117:44–55.
McNab BK. The relationship among flow rate, chamber volume and calculated rate of metabolism in vertebrate respirometry. Comp Biochem Phys A. 2006;145:287–94.
McNab BK. Ecological factors affect the level and scaling of avian BMR. Comp Biochem Phys A. 2009;152:22–45.
Mullur R, Liu YY, Brent GA. Thyroid hormone regulation of metabolism. Physiol Rev. 2014;94:355–82.
Piersma T, Bruinzeel L, Drent R, Kersten M, Van der Meer J, Wiersma P. Variability in basal metabolic rate of a long-distance migrant shorebird (Red Knot, Calidris canutus) reflects shifts in organ sizes. Physiol Zool. 1996;69:191–217.
Pitit M, Vézina F. Phenotype manipulations confirm the role of pectoral muscles and haematocrit in avian maximal thermogenic capacity. J Exp Biol. 2014;217:824–30.
Rasmussen UF, Vielwerth SE, Rasmussen VH. Skeletal muscle bioenergetics: a comparative study of mitochondria isolated from pigeon pectoralis, rat soleus, rat biceps brachii, pig biceps femoris and human quadriceps. Comp Biochem Phys A. 2004;137:435–46.
Rolfe DF, Brown GC. Cellular energy utilization and molecular origin of standard metabolic rate in mammals. Physiol Rev. 1997;77:731–58.
Schmidt-Nielsen K. Animal physiology: adaptation and environment. London: Cambridge University Press; 1997.
Scott I, Evans PR. The metabolic output of avian (Sturnus vulgaris, Calidris alpina) adipose tissue, liver and skeletal muscle: implications for BMR/body massrelationships. Comp Biochem Phys A. 1992;103:329–32.
Smit B, McKechnie AE. Avian seasonal metabolic variation in a subtropical desert: basal metabolic rates are lower in winter than in summer. Funct Ecol. 2010;24:330–9.
Steyermark AC, Miamen AG, Feghahati HS, Lewno AW. Physiological and morphological correlates of among-individual variation in standard metabolic rate in the leopard frog Rana pipiens. J Exp Biol. 2005;208:1201–8.
Sundin U, Moore G, Nedergaard J, Cannon B. Thermogenin amount and activity in hamsterbrown fat mitochondria: effect of cold acclimation. Am J Physiol. 1987;252:R822–32.
Swanson DL. Seasonal metabolic variation in birds: functional and mechanistic correlates. In: Thompson CF, editor. Current ornithology. Berlin: Springer; 2010. p. 75–129.
Swanson DL, Zhang YF, Liu JS, Merkord CL, King MO. Relative roles of temperature and photoperiod as drivers of metabolic flexibility in dark-eyed juncos. J Exp Biol. 2014;217:866–75.
Villarin JJ, Schaeffer PJ, Markle RA, Lindstedt SL. Chronic cold exposure increases liver oxidative capacity in the marsupial Monodelphis domestica. Comp Biochem Phys A. 2003;136:621–30.
Weathers WW. Energetics and thermoregulation by small passerines of the humid, lowland tropics. Auk. 1997;114:341–53.
Wells ME, Schaeffer PJ. Seasonality of peak metabolic rate in non-migrant tropical birds. J Avian Biol. 2012;43:481–5.
Wiersma P, Muñoz-Garcia A, Walker A, Williams JB. Tropical birds have a slow pace of life. Proc Natl Acad Sci. 2007;104:9340–5.
Wikelski M, Spinney L, Schelsky W, Scheuerlein A, Gwinner E. Slow pace of life in tropical sedentary birds: a common-garden experiment on four stonechat populations from different latitudes. Proc R Soc B Biol Sci. 2003;270:2383–8.
Williams J, Tieleman BI. Flexibility in basal metabolic rate and evaporative water loss among hoopoe larks exposed to different environmental temperatures. J Exp Biol. 2000;203:3153–9.
Wu MS, Xiao YC, Yang F, Zhou LM, Zheng WH, Liu JS. Seasonal variation in body mass and energy budget in Chinese bulbuls (Pycnonotus sinensis). Avian Res. 2014;5:4.
Wu MX, Zhou LM, Zhao LD, Zhao ZJ, Zheng WH, Liu JS. Seasonal variation in body mass, body temperature and thermogenesis in the Hwamei, Garrulax canorus. Comp Biochem Phys A. 2015;179:113–9.
Yen PM. Physiological and molecular basis of thyroid hormone action. Physiol Rev. 2001;81:1097–142.
Zheng WH, Liu JS, Jang XH, Fang YY, Zhang GK. Seasonal variation on metabolism and thermoregulation in Chinese bulbul. J Therm Biol. 2008a;33:315–9.
Zheng WH, Li M, Liu JS, Shao SL. Seasonal acclimatization of metabolism in Eurasian tree sparrows (Passer montanus). Comp Biochem Phys A. 2008b;151:519–25.
Zheng WH, Lin L, Liu JS, Xu XJ, Li M. Geographic variation in basal thermogenesis in little buntings: relationship to cellular thermogenesis and thyroid hormone concentrations. Comp Biochem Phys A. 2013a;164:240–6.
Zheng WH, Lin L, Liu JS, Pan H, Cao MT, Hu YL. Physiological and biochemical thermoregulatory responses of Chinese bulbuls Pycnonotus sinensis to warm temperature: phenotypic flexibility in a small passerine. J Therm Biol. 2013b;38:483–90.
Zheng WH, Liu JS, Swanson DL. Seasonal phenotypic flexibility of body mass, organ masses, and tissue oxidative capacity and their relationship to RMR in Chinese bulbuls. Physiol Biochem Zool. 2014a;87:432–44.
Zheng WH, Li M, Liu JS, Shao SL, Xu XJ. Seasonal variation of metabolic thermogenesis in Eurasian tree sparrows Passer montanus over a latitudinal gradient. Physiol Biochem Zool. 2014b;87:704–18.
Zhou LM, Xia SS, Chen Q, Wang RM, Zheng WH, Liu JS. Phenotypic flexibility of thermogenesis in the Hwamei (Garrulax canorus): responses to cold acclimation. Am J Physiol. 2016;310:R330–6.
Authors’ contributions
JL provided the research idea and designed the experiments. MB, XU and KC conducted the experiments and collected the data. MB and XU finished the data analysis, compiled the results and wrote the first draft of the article. JL and WZ supervised the research and revised the draft. All authors read and approved the final manuscript.
Acknowledgements
We thank Dr. Ron Moorhouse revising the English and giving some suggestions, and all the members of Animal Physiological Ecology Group, Wenzhou University Institute of Applied Ecology, for their helpful suggestions. This study was financially supported by Grants from the National Natural Science Foundation of China (No. 31470472), the National Undergraduate “Innovation” Project and Zhejiang Province’s “Xinmiao” Project.
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Bai, M., Wu, X., Cai, K. et al. Relationships between interspecific differences in the mass of internal organs, biochemical markers of metabolic activity, and the thermogenic properties of three small passerines. Avian Res 7, 11 (2016). https://doi.org/10.1186/s40657-016-0046-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40657-016-0046-1