
Abstract
Background
The reliance on chemical inputs to support high yields is the Achilles’ heel of modern crop production. The soil organic matter management is as old as agriculture itself. Recently, the use of soluble humic substances as plant growth promoters has been brought to attention due to their effects on nutrient uptake and water use efficiency. Humic substances applied directly at low concentrations can trigger different molecular, biochemical, and physiological processes in plants. However, how humic substances exert this plethoric regulatory action remains unclear. The objective of this study was to evaluate changes in the transcription level of genes coding cell receptors, phosphatases, synthesis, and function of different plant hormones and transcription factors.
Materials and methods
After seven days of humic acid treatment, we used RNAseq in maize root seedlings. The level of gene transcription was compared with control plants.
Results
Plant kinase receptors and different phosphatases were regulated by humic acids. Likewise, genes related to plant hormones (auxin, gibberellin, ethylene, cytokinin, abscisic acid, brassinosteroids, jasmonic and salicylic acids) were transcript in differential levels in maize root seedlings as well as the expression of a hundred of transcription factors modifying the signal transduction pathway via alterations of the subsequent gene response.
Conclusion
We showed a general mechanism for simultaneously regulating the activity of several hormones where humic acids act as a key regulatory hub in plant responses integrating hormonal signalling and response pathways.
Similar content being viewed by others

Introduction
Soil organic matter has a central role in human civilization. The relationship among humus content, soil fertility, and social development is not a coincidence and was essential for the development of the food-producing economy of Neolithic farmers [1]. Today, the challenge is feeding an increasingly demanding and growing population, concomitant to reducing external inputs and minimization of environmental impacts, even under variable and enhanced weather conditions expected in the future [2]. Reliance on the use of chemical inputs to support high yields is perhaps the environmental drawback of modern crop production. The overuse of fertilizers does not correspond to a significant increase in yields and is responsible for polluting waters and soils [3]. Pesticides were detected in 97% of stream water samples in agricultural areas of the USA [4]. While different efforts to reduce excessive use of agricultural chemicals have been exerted [5], plant growth promoters, and plant biostimulants may have a relevant role in this strategy.
Humic substances (HSs) are the major component of organic matter from soils, waters, and sediments [6] and are regarded as a complex non-covalent supramolecular association of relative small heterogeneous molecules that survive microbial degradation of plant and animal tissues and are held together by weak dispersive forces, hydrogen bonds, and metal-bridged intermolecular electrostatic bonds [7]. Humic substances became the most common material employed in biostimulants [8]. Plant metabolism and morphology are influenced by humic substances that modulate various biochemical mechanisms and physiological processes, stimulating growth, and increasing nutrient uptake [9]. Moreover, the best performance of humic substances was observed under stress conditions [10]. It was previously reported that HS significantly improves plant resistance to abiotic stresses [11,12,13,14,15]. This HS ability can be attributed to the interaction of multiple effects among which biosynthesis and concentration of secondary metabolites directly involved in stress alleviation, like phenols [16], expression of genes involved in plant responses to abiotic stress [17,18,19], including protective enzymes include catalase (CAT), ascorbate peroxidase (APX), superoxide dismutase (SOD), and polyphenol oxidase (PPO) [15, 20], and regulation of reactive oxygen species accumulation and metabolism [11,12,13,14]. Furthermore, it is well known that HS promotes several interconnected, hormone-mediated signalling pathways related to plant growth and defence [21]. Several plant hormones, such as ethylene [21,22,23], abscisic acid [24], salicylic acid [25], gibberellins [26], cytokinins [27], auxin [28], jasmonates [29], and brassinosteroids [30], were advocated to be involved in stress signalling. Furthermore, hormone-like activity is one of the humic substances' most suggested physiological effects [31]. Hormone-like responses of humic substances emulating auxins [32, 33], gibberellins [34, 35], cytokinins [36, 37], jasmonate [38, 39], alkamides [40], and nitric oxide [41] have been reported. However, how the plant cell perceives humic substances remains unclear.
According to Shah et al. [42], humic substances trigger various molecular processes in plant cells, manifesting their effects in cells through genetic, post-transcriptional, and post-translational modifications of signalling entities that promote different molecular, biochemical, and physiological processes. Nevertheless, how this happens remains again a matter of speculation. Our work hypothesizes that kinase receptors perceive humic substances that initialize a complex interactome via phosphorylation and downstream cascade by using hormone signalling pathways and regulation of transcription factors acting as key hub agents. In other words, plants perceive soluble humic substances as a typical environmental cue or stress agent that triggers hormonal crosstalk.
The objective of this work was to challenge this hypothesis by identifying the differential transcription level of hormonal signalling, transcription factors, and protein kinases codifying genes in the RNAseq for maize seedlings roots treated or untreated with humic acids.
Materials and methods
Humic acids
A solution of 0.5 M NaOH was added under shaking to earthworm compost (10:1, v/v) under N2 atmosphere. After 12 h, the suspension was centrifuged at 5000 ×g, and the humic acids (HA) were precipitated by adding 6 M HCl until pH 1.5. After centrifugation (5000 ×g) for 15 min, the sample was repeatedly washed with water until chloride free. Subsequently, the sample was dialysed against deionized water using a 1000-Da cut-off membrane (Thomas Scientific, Swedesboro, NJ, USA) and lyophilized. The HA solution was prepared by solubilizing HA powder in 1 mL of 0.01 M NaOH, followed by pH adjustment to 6.5 with 0.1 M HCl. After freeze drying, the carbon content was analysed by dry combustion (CHN analyser Perkin Elmer series 2400, Norwalk, CT, USA). The molecular composition of HA was evaluated by Cross-polarization magic-angle spinning (CP/MAS) 13C nuclear magnetic resonance (13C-NMR). The spectrum was acquired from the solid sample with a Bruker Avance 500 MHz (Bruker, Karlsruhe, Germany), equipped with a 4-mm-wide bore MAS probe, operating at a 13C-resonating frequency of 75.47 MHz. The spectra were integrated over the chemical shift (ppm) resonance intervals of 0 to 46 ppm (alkyl C, mainly CH2 and CH3 sp3 carbons), 46 to 65 ppm (methoxy and N alkyl C from OCH3, C–N, and complex aliphatic carbons), 65 to 90 ppm (O-alkyl C, such as alcohols and ethers), 90 to 108 ppm (anomeric carbons in carbohydrate-like structures), 108 to 145 ppm (phenolic carbons), 145 to 160 ppm (aromatic and olefinic sp2 carbons), 160 to 185 ppm (carboxyl, amides, and esters), and 185 to 225 ppm (carbonyls).
Plant treatment
Maize seeds (Zea mays L., var. Dekalb 177) were surface-sterilized by soaking in 0.5% NaClO for 30 min, rinsing, and then soaking in water for 6 h. Then, the seeds were sown in 2.0-L pots filled with washed and sterilized sand wetted with 1/3 strength Furlani nutrient solution [43] (μmol L−1: 3.527 Ca; 2.310 K; 855 Mg; 45 P; 587 S; 25 B; 77 Fe; 9.1 Mn; 0.63 Cu; 0.83 Mo; 2.29 Zn; 1.74 Na; and 75 EDTA), with the N content adjusted to a low concentration (100 μmol L−1 NO3 + NH4). Six replicates were used in a randomized statistical design. After 1 week, the solution was changed for one-half of the ionic force. At 7 days after planting, the maize seedlings were treated with solutions containing humic acids diluted with low N Furlani nutrient solution at 0 and 4 mM C L−1: Seven days after treatment, imposing three plants per pot were collected, and root tissues were analysed individually for RNAseq, and the mean value was considered. Three replicates were used, with a total of nine plants analysed. The experiment was entirely repeated twice.
Transcriptional analysis of humic acid-treated maize root plants
For RNA extraction, 100 mg of control roots and HA-treated roots, using the best dose for root growth at 4 mM C HA L−1, was macerated in liquid nitrogen. According to the manufacturer's instructions, the total RNA of the samples (3 biological replicates per treatment) was extracted with the RNeasy Plant Mini Kit (Qiagen). Total RNA was quantified using the Nanodrop 1000 spectrophotometer. The RNA was eluted in DEPC-treated water (total amount of 4–10 μg RNA) digested with DNAse and depleted ribosomal RNA using the GOTAQ® 1-STEP RT-QPCR (PROMEGA). Subsequently, a 1% free RNAse agarose gel was made to analyse the RNA extracted. According to the manufacturer's protocol, sequencing libraries were prepared using the Whole Transcriptome Analysis kit (Applied Biosystems). Libraries were sequenced on the Illumina platform by LacTad company—Brazil. The reads obtained from the RNAseq technology were analysed to identify ribosomal RNA (rRNA) sequences in two steps: 1- rRNA sequences of Zea mays were downloaded from NCBI, and an index file of rRNA was created using Novoalign v3.06.05. (http://www.novocraft.com/products/novoalign/). Then reads were mapped on index file using Novoalign; 2- All fastq files were converted into Fasta, and BLASTN analysis was performed against downloaded rRNA sequences. Identified rRNA sequences were removed, and reads were cleaned. Further, the quality of all reads was accessed by running the FastQC software [64], and high-quality cleaned reads were aligned on Z. mays genome using Novoalign. Gene expression levels were normalized as reads per kilobase of transcript per million mapped reads (RPKM). The differential gene expression between control and HA-treated samples was determined by using Cuffdiff v2.2.1. The genes with differences of at least one-fold change along with adjusted p-value (FDR) ≤ 0.05 were considered to be significantly differentially expressed. Functional classification analysis was executed with MapMan version 3.6.0RC1 (https://mapman.gabipd.org/).
Results
Receptor protein kinases (RPKs) are a diverse group of single-pass transmembrane proteins with extracellular and cytosolic domains that allow cells to recognize and respond to their extracellular environment. Table 1 shows that 261 related kinases receptors, located at the cell wall, membranes, and cytosol, were differentially expressed in maize root seedlings when treated with HA. The majority RPKs found in a higher transcript level contained leucine-rich repeat (LRR) sequences implicated in protein–protein interactions. In addition, another group was found to contain an S-domain, and several others contained unique features, such as tyrosine and chitin kinase.
Following RPKs modulation by HA, we expected that also protein phosphorylation might be induced since protein phosphorylation is the most common mechanism for regulating and controlling protein activity and function [44]. Table 2 shows the main phosphatases up and down-regulated by HA grouped by the functional category.
Phosphatases and kinases control multiple cellular events, including cell proliferation, differentiation, and stress responses, by regulating reversible protein phosphorylation, the most important post-translational modification. The activation of genes codified different cell wall acid phosphatases, representing a group of essential proteins capable of hydrolysing and solubilizing organic soil phosphate independently of soil microbial activity [45]. Purple acid phosphatases (PAPs) belong to the family of binuclear metallohydrolases, which catalyse a wide range of phosphomonoesters and are involved in plants' phosphate uptake [46]. Nine PAPs were found in a larger significant transcription level than for control, seven of which were inhibited and two up-regulated (Table 2). Other phosphatases with metal ion binding functions were found, including different inorganic pyrophosphatases. The cell wall is characterized by a significantly significant presence of a stem 28 kDa glycoprotein [47] that was down-regulated by HA treatment in root seedlings. Several cytosolic phosphatases with differential transcription were found, including those linked to carbohydrate metabolism. Putative ribose-5-phosphate isomerase and similar proteins were both down- and up-regulated by HA. These enzymes catalyse the reversible conversion between ribose-5-phosphate and ribulose-5-phosphate in the pentose phosphate pathway [48]. Glucose-6-phosphate dehydrogenase (G6PDH) provides nicotinamide adenine dinucleotide phosphate (NADPH) and intermediate metabolites for the biosynthesis of several compounds that control the flux through this non-reversible branch of the oxidative pentose phosphate pathway [49] and were found in differential transcription level in maize root seedlings treated with HA (Table 2). The 2-hydroxy-3-oxopropionate reductase (garR) is involved in galactarate degradation and catalyses the reduction of tatronate semialdehyde to D-glycerate and is involved in the carbohydrate acid metabolism. This enzyme was also found down- and up-regulated in respect to control. Different DNA-binding phosphatases were found for the HA treatment, including glyoxylate reductase (NADP) activity (GLYRs) as glyoxylate/hydroxypyruvate reductase (HPR3), glyoxylate/succinic semialdehyde reductase 2 chloroplastic, glyoxylate reductase, and 2-hydroxy-3-oxopropionate reductase. These enzymes catalyse the reduction of glyoxylate to glycolate by using the NADH or NADPH cofactor and were involved in detoxifying aldehydes during stress and contributing to the redox balance [50].
Our experiment found two dose-dependent cell cycle regulator 2 (Dcr2) phosphatases up-regulated by the HA treatment (Table 2). These enzymes played a positive role in cell cycle progression and stress response, acting as an antagonist of unfolded protein response (UPR) [51]. Sit4 was down-regulated by HA, implying a phosphatase's involvement in various processes, such as transcription, translation, bud formation, glycogen metabolism, monovalent ion homeostasis, and H + transport, including telomere function being functionally linked to the ubiquitin–proteasome system. Lipid phosphatases were found in differential transcription levels, including lipid phosphate phosphatase 2, which was involved in abscisic acid signalling by dephosphorylation of diacylglycerol pyrophosphate (DGPP) and phosphatidic acid (PA), which are known secondary messengers [52]. Phosphatidylinositol was found in different transcription levels and is responsible for transferring the phosphorylinositol group from phosphatidylinositol (PI) to phytoceramide, an essential step in sphingolipid biosynthesis, and is believed to be associated with self-defence through the promotion of sphingolipid metabolism and regulation of ceramide accumulation. Different inositol phosphate synthases were coupled to lipid metabolism and plant stress response by using lipids phosphorylation [53]. Myo-inositol 1-phosphate synthase and Myo-inositol-3-phosphate were implied in high transcription levels. They were involved in various biochemical and physiological processes, such as intracellular signal transduction, membrane construction and trafficking, membrane-related protein anchoring, and cell wall construction [54]. Mitogen-activated protein kinase (MAPK) cascades acted as signal transduction pathways to translate external stimuli into cellular responses, including hormones responses. Table 3 shows the MAPK enzymes found in significantly differential transcription levels in respect to control, most of which were inhibited by HA treatment.
Auxins showed the highest number of genes up- and down-regulated for root tissues exposed to HA (Table 4). Sixty-three genes related to metabolism and auxin synthesis were found at a high transcription level, with 26 down-regulated and 37 up-regulated control.
The second plant hormone with a transcriptional level modified by HA was ethylene (50 genes), of which 28 was down- and 22 up-regulated. ABA [39] was another plant hormone with more genes [25] down-regulated than up-regulated [14] by HA treatment in respect to control. There were 22 genes involved in gibberellin synthesis, signalling and metabolism with high transcript levels, compared to the control, with 13 down- and 9 up-regulated. We found 18 proteins related to brassinosteroids (BR) and jasmonates (JA) biosynthesis and metabolic processes. However, in BR, most genes were up-regulated, while in JA, the HA treatment displayed more genes down-regulated in respect to control. Finally, 14 genes were found to be related to CK and 4 linked to SA synthesis and metabolism. It is the first time evidence has been presented on humic matter capacity to modify gene responses linked to BR signalling in plants.
Table 5 shows the relationship among genes linked to plant hormones in significant transcriptional levels, considering positive or inhibitory effects. Although the classical literature about the hormone-like activity of humic substances report each specific and separately hormone-like activity, it is evident that HA is promoting changes in the overall hormone balance (Table 5).
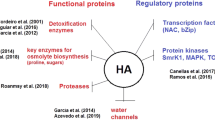
Extensive interactions among genes in both synergist and inhibitory pathways were observed for different plant hormones. Genes related to auxin synthesis, metabolism, and action interplaying with other plant hormones are well explored in the literature (Table 5). However, it is also clear that HA induces changes in all plant hormone responses. The ultimate interaction of cell receptors, phosphatases, MAPK, and hormonal signalling is the activation of transcription factors (TF) and induction of gene responses. It was observed that 948 different TFs were transcripted in different levels in response to HA, with 330 up-regulated and 518 down-regulated. Figure 1 summarizes the main families of TFs induced by HA treatments, while Table 6 reports TFs up and down-regulated related to plant hormone gene response. HA inhibited most TF, but a wider redundancy indicates that almost all TF are inhibited at a significant and positive expression.

Hormonal Transcription factor and regulatory genes integration network induced by humic acid (HA) treatment. Abbreviations: NAC: NAC (NAM, ATAF1,2, and CUC2; MYB: MYB Proto-Oncogene; MYC Proto-Oncogene; bZIP: Basic leucine zípper; WRKY: WRKY Family; AP2/ERF: APETALA2/Ethylene-responsive element-binding protein family; B3: B3 domain-containing protein; ARR: Two-component response regulator; REMORIN: Remorin family protein; Aux/IAA: Auxin/indole-3-acetic acid protein Family; ARF: Auxin Response Factor Family; SUVR5: Histone-lysine N-methyltransferase; bHLH: Basic Helix-Loop-Helix Family; HDAC: Histone deacetylase; PPR: Putative pentatricopeptide repeat-containing protein; F-box: F-box protein; LSD: Lysine-specific demethylase; PLATZ: Plant AT-rich sequence and zinc-binding proteins; CAMTA: Calmodulin-binding transcription activators; Ring-type E3: RING-type E3 ubiquitin transferase; PAT: S-acyltransferase; GRAS: GRAS transcription factor family (Scarecrow); CRF: Chromatin Remodelling Factors; AGO: Argonaute; GTE: Transcription factor group E; MTREF: Mitochondrial transcription termination factor family protein; G-box: G-box protein; MOB: MOB kinase activator like; UBX2: Ubiquitin regulatory X domain-containing protein 2; SWI/SNF: SWItch/Sucrose Non-Fermentable; EIN3: EIN3-like (EIL) transcription factor Family; FREE1: fyve domain protein required for endosomal sorting 1; TCP: TCP protein domain; LOB: LOB domain-containing protein; Alba: Alba DNA/RNA-binding protein; SEUSS: Transcriptional corepressor SEUSS; C3H Type: C3H-type transcription factor; GRFs: GRF transcription factor; ABH: Alpha/beta hydrolase; CtBP: C-terminal-binding protein; ALF: Alfin-like transcription factor; HB: Homeobox transcription factor Family
Discussion
Due to the significant molecular heterogeneity and superstructural complexity of HA, it is unlikely that specific plant cell receptors may be specifically elicited. It would be more reasonable to imagine general cell pseudo-receptors being sensitized. One of the central and well-known wide energy cell sensors is the sucrose non-fermenting-1 (SNF-1) kinases family. In fact, differential expression of the SnRK2.2 protein kinase genes (serine/threonine kinase related to non-fermentative sucrose SNF) induced by HA was previously observed by RTq-PCR [73]. Other general pseudo-receptors are proteins linked to H+-ATPase that change plasma membrane potential after being challenged by various types of signals to initiate a Ca2+-mediated signal transduction cascade [74]. The effect of HA on H+-ATPase activity is one of the most studied physiological effects [9]. However, more than 250 RPKs were elicited by HA, including cell wall, transmembrane, and cytosolic receptors (Table 1), which revealed a putative network dedicated to perceiving and transmitting the HA cue. This humic effect is reported here for the first time, opening a new perspective on how plants perceive biostimulants based on HA applied at low concentrations.
The subsequent event after cell perception is the amplification of the signal, which is manifested by the specific signal transduction cascade pathways. Post-translational modification of Ser, Thr, and Tyr residues by protein kinases and phosphatases is a major transduction route for many signals [75]. According to Trevawas [76], protein phosphorylation is a common means of manipulating connection strength. There are about 1000 protein kinases and hundreds of protein phosphatases in plants with differing degrees of specificity and control for constructing a phosphoproteome. Almost a quarter of these proteins were differentially transcribed in the HA-treated maize root seedlings showing a strong effect on signal transduction (Tables 1 and 2). In a previous proteomic study with maize seedlings treated with HA isolated from vermicompost, 24 up-regulated proteins linked to protein modifications were found, but few were phosphatases [77]. One of the most important groups of protein kinases comprises the MAPK cascade involved in transduction signals triggered by plant hormones. Interestingly, the treatment inhibited most MAPK and MAPKK (Table 3). However, the genes codifying proteins related to synthesis, functioning, and regulation of plant hormones were found to be both up- and down-regulated by HA (Table 4).
Indole-3-acetic acid-amido synthetase (GH3) was both up- and down-regulated by the HA treatment. The conjugation of indole-3-acetic acid (IAA) to amino acids by GH3 is an essential part of regulation for auxin level, thus providing a mechanism for the plant to cope with excess auxin. According to Ding et al. [78], GH3 activates disease resistance in salicylic acid signalling and jasmonic acid signalling-independent pathways. The IAA-amino acid hydrolase ILR1-like 4 is a family of hydrolase genes initially isolated in Arabidopsis thaliana and involved in regulating auxin levels [79] and hydrolysing amino acids acid conjugates with IAA, including IAA-Ala, IAA-Asn, IAA-Cys, IAA-Glu, IAA-Met, IAA-Ser, and IAA-Gly [79]. ILR1 hydrolyses amino acids that conjugate with jasmonic and hydroxy jasmonic acids and is also induced by jasmonic acid (JA).
The small Auxin Up-regulated RNA (SAUR) family is controlled by HA and consists of early auxin-responsive genes with an overall effect on auxin signalling-regulated plant growth and development. These genes can be readily induced by exogenous auxin [80] and are involved in plant growth by inhibiting PP2CD phosphatases, which activate plasma membrane (PM) H+-ATPases promoting cell expansion [80]. The effect of HA on PM H+-ATPase activity was previously reported [70,71,72]. Besides auxin, hormonal regulation of SAUR expression was observed with other hormones, such as brassinosteroids (BR), gibberellins (GA), jasmonate (AJ), and abscisic acid (ABA) [84, 85]. HA treatment increased the differential transcription level of specific SAUR11, SAUR32, and SAUR36 compared to control.
Parker et al. [86] have identified two recessive allelic mutants in Arabidopsis, designated as a continuous vascular ring (cov1), that display a dramatic increase in vascular tissue development and were induced by auxin. A high auxin level induces polar auxin transport, resulting in vascular differentiation. PIN1 protein is essential to auxin transport and has a high transcription level in HA-treated seedlings. PIN proteins induce polar auxin transport because of their asymmetric subcellular localizations [87]. Moreover, it was found that genes differentially expressed coding proteins linked to a ubiquitination process that is directed by auxins, such as F-box FBX14, BIG (BIG) binding/ubiquitin-protein ligase/zinc ion binding, auxin signalling F-BOX 3, DCN1-like protein 2 (Defective in cullin neddylation protein), and TIR1 (TRANSPORT INHIBITOR RESPONSE 1). Ubiquitination of proteins is a widely employed mechanism for hormonal regulation. Ubiquitin is covalently attached to an internal lysine residue or the N terminus of a substrate protein, as those conjugated with auxins promoted by GH3 and ILR1. Additional ubiquitin moieties are attached to the initial ubiquitin, resulting in the formation of ubiquitin chains. In most cases, these chains are recognized by the 26S proteasome, which degrades the tagged substrates. It is possible that HA can regulate auxin signalling by an interaction between TRANSPORT INHIBITOR RESPONSE 1/AUXIN SIGNALINGF-BOX proteins (TIR1/AFBs) and AUXIN/INDOLE-3-ACETIC ACID (Aux/IAA) coreceptor proteins and the subsequent transcriptional regulation. The underlined mechanism was extensively studied, showing that the process of auxin perception by TIR1 (an F-box protein) forms an SKP1–Cullin–F-box (SCF) ubiquitin ligase complex inducing further complexation with the AUXIN/INDOLE-3-ACETIC ACID (AUX/IAA) transcription regulator and results in proteasomal degradation of AUX/IAA, thereby promoting the expression of auxin-inducible genes [88]. Cullin-based E3 ubiquitin ligases are activated by modifying the cullin subunit with the ubiquitin-like protein Nedd8. DCN1 regulates cullin neddylation and thus ubiquitin ligase activity [89] and was found in a high transcription level in root seedlings treated with HA. The inhibition of various auxin-responsive proteins (IAA4, IAA9, IAA13, IAA24) can be interpreted as one additional evidence of promoting the ubiquitination process induced by HA.
Degradation of the Aux/IAAs activates the auxin response factor (ARF) family of transcription factors, whose activities in regulating auxin-responsive genes are otherwise inhibited by the Aux/IAA proteins. This mechanism is mediated by the TIR1 ubiquitin ligase [90]. HA treatments also induced other genes related to mechanisms of auxin homeostasis regulation. Glycosylation is one of the general homeostatic mechanisms of metabolism of small molecules, and glycosyltransferases recognize a great diversity of substrates, including hormones (auxin and cytokinins), secondary metabolites, and xenobiotics as pesticides and herbicides [91]. In addition, O-fucosyltransferases are glycosyltransferase regulated by the HA treatment. Finally, we found a particular mechanism of cell homeostasis triggered by HA involving the redox process. It is well known that plants can detoxify reactive oxygen species and reactive aldehydes induced by stress, to less toxic compounds, such as alcohols or carboxylic acids, by up-regulating the expression of a few enzymes with oxidoreductase activity. Aldo–keto reductases (AKRs), belonging to the oxidoreductase superfamily, are stress-regulated genes and play an essential role in the cellular response to electrophilic, osmotic, and oxidative stress depending on the presence of coenzyme NAD (P)(H) (Nicotinamide Adenine Dinucleotide Phosphate). The presence of many auxin and abscisic acid response elements in the promoters of MtAKRs indicates potential functions in plant growth and responses to environmental stresses.
The MtAKR2 promoter contains both auxin and MeJA response elements [92]. Apoplastic reductants act as an antioxidant barrier and modulate oxidative signals, thus contributing to cell homeostasis. Ascorbate occurs at 10–4 to 10–3 M concentrations in the apoplast, representing the primary pool of low-molecular-mass antioxidants. Preger et al. [93] reported that the trans-PM electron transfer from cytosolic ascorbate to apoplastic MDA is affected by a cytochrome b561. The major ascorbate-reducible cytochrome b is associated with the PM of soybean codified by AIR12 (for auxin-induced in root cultures). A DOMON-plus-cytochrome b561 protein was found to be induced by HA treatment. In this respect, the increase in ascorbate concentration in plants treated with HA was previously reported [94].
We found 39 genes promoted by HA linked to ABA activity. Among them, 25 were negatively regulated, and 14 were positively regulated. ABA response element-binding factor has a DNA-binding transcription factor activity displaying abscisic acid-activated signalling pathway. Both are up- and down-regulated by HA. bZIP (basic zipper) leucine was up-regulated in the HA treatment. The primary target of the bZIP class of transcription factors is the ABRE motif, an important one for its proper regulation by ABA. ABF4 (ABRE BINDING FACTOR 4) was found in a high level of transcription regarding control. At the same time, ABA INSENSITIVE(ABI)5/ABA Response Element (ABRE)-binding factor (ABF/AREB) is another clade of basic leucine zipper (bZIP) transcription factors that are generally induced by ABA in response to dehydrating stresses promoted by the HA treatment. Lynch et al. [95] reported that these bZIPs must be phosphorylated to be active and are inhibited by protein phosphatases and ubiquitin ligases, which affect their activity and stability, respectively. Putative GEM-like protein 8 belongs to GRAM domain-containing gene family has a multiple ABRE cis-element [96], which was termed the ABA-responsive protein (ABR) family and was up-regulated by HA. These domains are implicated in functional protein association networks. ABI1 was also shown to bind weakly with the Sucrose non-fermenting Related Kinase, SnRK3.1, that participates in global stress responses [97]. Protein serine/threonine phosphatase was up-regulated by HA and are receptors that start ABA signalling cascade by the phosphorylation of the C-terminus of the H+-ATPase to release the protein from autoinhibition, thus allowing access by 14–3-3 proteins to maintain the proton pump in the active conformation [98]. ABA counteracts the blue-light-dependent activity of the H+-ATPase and locks the membrane potential to depolarize state, thereby allowing the continuous effluxes of cations and anions [99, 100].
A critical rate-limiting step in ABA synthesis is the cleavage of the carotenoids 9-cisneoxanthin and 9-cis-violaxanthin in the chloroplast to yield xanthoxin, which is exported to the cytoplasm and metabolized to ABA [101]. This carotenoid cleavage reaction is catalysed by the 9-cisepoxycartenoid dioxygenases (NCEDs) strongly down-regulated by the HA treatment. Furthermore, AAO (Arabidopsis Aldehyde Oxidase) promotes the last step of ABA biosynthesis, catalysing the oxidation of abscisic aldehyde [102], and was also down-regulated by HA. Besides NCED, others called carotenoid cleavage dioxygenases (CCD) genes down-regulated by HA and may be involved in another hormone biosynthesis. Indole-3-acetaldehyde oxidase is an enzyme involved in the redox process and the abscisic acid and auxin biosynthetic process. This enzyme belongs to the family of oxidoreductases, specifically acting on the aldehyde or oxo group of donors with oxygen as acceptors. The oxidation of abscisic aldehyde in the last step of ABA biosynthesis is catalysed by aldehyde oxidase (EC 1.2.3.1) [103], which is also involved in tryptophan synthesis [104]. Abscisic acid-induced-like protein (HVA22) was both up- and down-regulated by HA and involved ABA perception. Polyadenylation is an alternative post-transcriptional regulatory mechanism that is important in generating new transcript isoforms and, therefore, new gene functions. Polyadenylation is a highly regulated process that involves the activity of FIP1, which provides a bridge between poly(A) polymerase (PAP) and other subunits of the cleavage and polyadenylation factors [94]. Genes possessing polyadenylation sites within protein-coding regions or introns are significantly associated with stress responses. According to Tellez-Robledo et al. [105], FIP1 is needed for the correct polyadenylation of transcripts that display ABA signalling responses and stresses, such as salt or cadmium genes. In addition, Wang et al. [106] revealed that FIP1 negatively regulated the expression of CIPK8 and CIPK23, two protein kinases involved in nitrate signalling. FIP1 was both up- and down-regulated by the HA treatment.
The homeobox gene GLABRA2 (GL2) has been considered a key component in the root-hair pattern determined by a regulatory circuit composed of transcription factor genes. GL2 exerts its regulatory effect on root-hair development by modulating phospholipid signalling [107]. ABA INSENSITIVE 8 (ABI8]/ELONGATION EFFECTIVE 1 (ELD1]/KOBITO1 (KOB1] promoted the expression of genes involved in cell elongation and cellulose synthesis in addition to the traditional light response, encoding a glycosyltransferase-like protein in Arabidopsis previously that is implicated in cellulose biosynthesis and hypocotyl elongation [108].
Gibberellin 2-oxidases (GA2oxs) are a group of 2-oxoglutarate-dependent dioxygenases that catalyse the deactivation of bioactive GA or its precursors through the 2β-hydroxylation reaction [109]. We found seven genes linked to this family, with six down-regulated and one up-regulated by the HA treatment. Gibberellins (GAs) are a large group of diterpenoid natural products that regulate various developmental and growth processes and impact crop yields. When gibberellin synthesis is inhibited, more precursors in the terpenoid pathway accumulate, potentially increasing the production of other molecules, such as abscisic acid. Some plant growth regulators used in agriculture, such as Paclobutrazol, inhibit the GA synthesis inducing anti-stress response. The main target of this kind of plant growth regulator is the inhibition of the Cyt P450 necessary for the kaurene oxidase activity. Ent-kaurene oxidase catalyses three steps of GA biosynthesis [110], and both ent-kaurene oxidase and kaurene oxidase 2 were down-regulated by HA. However, the intermediate steps that oxidize ent-kaurene to GA12 are catalysed by several Cyt P450 monooxygenases found at high transcriptional levels in roots seedlings treated with HA.
Furthermore, ent-copalyl diphosphate synthases also have a distinct ent-kaurene synthase activity associated with the same protein molecule. The suppression of ent-kaurene synthase activity of the protein leads to the build-up of ent-copalyl pyrophosphate found in high transcript levels by the HA treatment. In addition, ent-copalyl as precursors for several classes of phytoalexins involved in stress response besides GAs [111]. Phytoalexins are low-molecular-weight compounds produced after exposure to microorganism attacks and other elicitors, and it has been suggested that they serve as plant antibiotics. GA signalling is mediated by GIBBERELLIN-INSENSITIVE DWARF1 (GID1) and DELLA proteins in collaboration with a GA-specific F-box protein. The DELLA protein belongs to the GRAS superfamily of putative transcription factors, a well-characterized factor involved in the GA signalling pathway. The DELLA protein functions as a negative regulator of GA signalling and is rapidly degraded when plants are treated with GA [112]. These authors also reported that GIBBERELLININSENSITIVE DWARF2(GID2) and SLEEPY1 (SLY1), candidate F-box components of Skp1-Cullin-F box protein (SCF) E3 ubiquitin ligases, are responsible for targeting DELLA proteins to the proteasome. It is found that HA down-regulated genes that encode DELLA and SLEEPPY proteins. The binding of GA to its receptor GA INSENSITIVE DWARF1 (GID1) enhances the GID1–DELLA interaction, which, in turn, leads to the rapid proteolysis of DELLA through the ubiquitin–proteasome pathway and allows transcriptional reprogramming of GA-responsive genes. The mechanism of this regulation is not clear, although SCARECROW (SCR) appeared to be implicated as a regulatory protein [113]. GASA (gibberellic acid stimulated in Arabidopsis) proteins are small peptides localized in the apoplast or cell wall, and it is found that HA up-regulated their expression. The role of GASA in the interaction between different hormonal signalling pathways in redox regulation and several aspects of plant biology was reported [114, 115].
Ethylene is an essential regulator of many developmental and physiological processes and plays a vital role in the plant’s defence against biotic and abiotic stress factors [116]. The biosynthetic pathway of the ethylene is well characterized, and two enzymes involved in the pathway, ACC synthase (EC 4.4.1.14) and ACC oxidase (EC 1.4.3), have been identified [117]. Furthermore, the genes capable of codifying both proteins were down-regulated by HA treatment. Ethylene response factors (ERFs) are integral to the ethylene signalling and response pathway. ERFs are transcription factors that can bind to cis-acting elements, such as GCC-box motifs and dehydration-responsive elements (DREs) [118]. Ethylene is sensed by receptors localized at the endoplasmic reticulum membrane, including Ethylene Response1 [ETR], and results to be down- [ETR40] and up- [ETR1] regulated by HA. ERFs are key regulators in abiotic stress tolerance, such as drought, salinity, light stress, and cold and heat treatments. ERF3 was up-regulated, while ERF8 and ERF1B were down-regulated. The APETALA2/ethylene-responsive element-binding protein (AP2/EREBP) superfamily is one of the largest groups of plant-specific transcription factors. They play vital roles in plant growth, development, and response to abiotic and biotic stresses. As ERF, AP2/EREB was found to be implicated in various hormone-related signal transduction pathways, including ABA, CK, and JA [119]. Another candidate to make a bridge between stress and transcription factors are the Multiprotein Bridging Factor 1 [MBF1) family of proteins [120]. According to these authors, some genes related to abiotic and biotic stress response regulated by ET (and other plant hormones, like JA and SA) were up-regulated in the MBF1a overexpression in plants, suggesting that MBF1a could be protecting the plant against pathogen attack via ethylene and jasmonic acid-dependent pathways. All three MBF1 forms were found negatively regulated by the HA treatment. The Arabidopsis thaliana zinc finger transcription factor (ZF-TF), S-nitrosothiol (SNO) Regulated 1 (SRG1), is a central target of NO bioactivity during plant immunity. SRG1 appears to act as a transcriptional repressor utilizing its putative ERF-associated amphiphilic repression (EAR) domain to recruit the corepressor TOPLESS, thus contributing to plant defence responses [121]. HA was found to down-regulate SRG1. Finally, DeSI-like protein belongs to an ethylene-responsive element-binding protein with ubiquitinyl hydrolase activity. Its function on hormone homeostasis is linked to protein modification by small protein removal. These proteins were found to be regulated by the HA treatment.
Brassinosteroids (BRs) are a group of plant steroidal hormones ubiquitous in the plant kingdom and regulate various physiological responses. The BRASSINAZOLE-RESISTANT 1 (BZR1) transcription factor family is essential in plant brassinosteroid (BR) signalling. The well-known negative regulation of abscisic acid inhibits root growth by brassinosteroids and is partially mediated via direct suppression of ABSCISIC ACID INSENSITIVE 5 expression by BRASSINAZOLE RESISTANT 1 [122]. The transcription level of BZR1 was significantly down-regulated in the roots treated with HA. BRs are perceived at the cell surface, and their biosynthesis occurs on the endoplasmic reticulum (ER) using different pathways. [123]. Two different BRs receptors were down-regulated by HA (Brassinazole-resistant 1 protein and Brassinosteroid-responsive/RING-H2). However, several proteins involved in plant steroid biosynthesis were up-regulated by HA, including Delta (24)-sterol reductase, Steroid 5-alpha-reductase (DET2), Polyprenol reductase, Squalene monooxygenase, Sterol 14-demethylase, and 3-Epi-6-deoxocathasterone 23-monooxygenase (cytochrome P450 90C1/CYP90D1), among others.
Cytokinin (CK) plays an important role in regulating shoot and root growth, leaf senescence, chloroplast development, stress response, and pathogen resistance [124]. Isopentenyl transferase1 (IPT1) catalysed the first step in the Ck biosynthesis and was up-regulated by HA. Moreover, Ck oxidases/dehydrogenase catalyses the irreversible degradation of Ck. CKX4 are strongly inhibited by the HA treatment, while CKX6 was up-regulated. The Ck signal is perceived in Arabidopsis by histidine kinase receptors that were up- and down-regulated by HA addition to maize root seedlings. The common Ck signalling system has a membrane-bound receptor histidine kinase, which senses the signal and autophosphorylates. A response regulator activates the transcription of its target genes upon phosphorylation by the receptor kinase or initiates another output reaction [125]. The histidine kinase was found to be regulated by the HA treatment (Table 1).
JAs (jasmonates) are cyclopentanone compounds derived from linolenic acid via an octadecanoid pathway consisting of several enzymatic steps. The early steps of this pathway occur in chloroplasts, where linolenic acid is converted to 12-oxo-phytodienoic acid (OPDA) using three enzymes, lipoxygenase, allene oxide synthase (AOS), and allene oxide cyclase, and is subsequently reduced in a cyclopentenone ring by a peroxisome-localized enzyme, 12-oxophytodienoic acid reductase 3 [126]. All these proteins involved in JA biosynthesis were negatively regulated in the HA treatment, except for lipoxygenase (LOX11).
The integration of different plant hormones is a well-known process in system biology studies, and its magnitude is often dependent on plant development and physiological processes. For example, auxins are a pivotal plant hormone that commands rooting [127], and the auxin-like activity is the most reported effect of HA [21]. Here, we integrated the HA and plant hormone network using transcriptomic data.
The keystone of environmental and hormonal effects in plants is the signal perception and transduction cascade. Our results showed that HA alters genes' expression, codifying proteins related to the perception, metabolism, and signalling of ABA, CK, AUX, BR, CH, ET, GA, JA, and SA, thereby changing the hormonal crosstalk. These genes have multiple regulations and are both inhibited or promoted by hormones indicating putative complex crosstalk. Table 5 shows the main known hormonal crosstalk previously defined in the scientific literature involving the genes significantly regulated by HA. Genes stimulated by HA include regulatory proteins that further alter gene expression and possibly function in hormonal crosstalk response. They comprise several transcription factors (TFs), emphasizing the role of various transcriptional regulatory mechanisms in the hormone signal transduction pathways. The TFs interact with cis-elements in the promoter regions of several genes and thus up-regulate the expression of many downstream genes, resulting in imparting hormone response. Table 6 reports the relationship among TFs found in differential transcription level and plant hormone crosstalk based on literature, while Table 7 reports the interaction between TFs and plant hormones.
The phytohormone auxin is the central regulator of root architecture, particularly lateral root emergence in plants, and coordinates a complex gene regulatory network enabling plants to cope with decreased water and nutrient availability. HA was found to change root development and its response to environmental cues. These responses include changes in transcript levels of different plant hormones (Table 2). Since auxin is a master regulator of root traits, we put this hormone in focus in this discussion. Genes for various transcription factors containing typical DNA-binding motifs, such as MYB, bZIP, MYC, ERF/AP2, and Zinc fingers, have been inducible by HA (Fig. 1).
MYB, Zinc finger family, WRKY, AP2/EREB, and NAC were the TFs that held more differential transcription levels induced by HA treatment that, in turn, promoted both up- and down-regulation, with the majority being negatively regulated. Myeloblastosis viral oncogene homolog (MYB) transcription factors are active players in abiotic stress signalling [177] and correspond to one of the most prominent TF families in plants. MYB is induced by drought and other abiotic stresses and positively controls the drought resistance by inducing stress and auxin-responsive genes [167]. MYB TFs also affect signal transduction and phytohormone biosynthesis, including auxin, gibberellic acid, methyl jasmonate, and ABA [179], becoming a strong candidate for a mediator of the integration among different hormonal signals. The role of MYBs in root development and differentiation has been previously reported [180]. It involves the control of cell cycle progression at the root tips [129] and the expression of auxin-inducible genes regulating lateral root formation [181]. In addition, it was observed that MYB TF can regulate lateral root meristem activation under drought conditions via ABA–auxin signalling crosstalk [182]. MYB are also critical regulators of the synthesis of phenylpropanoid-derived compounds [183]. Plant phenylpropanoid-derived compounds are a large family of phenylalanine-derived secondary metabolites, including monolignols, flavonoids (anthocyanins, proanthocyanidins, flavonols), flavones, flavanones, isoflavonoids, and phlobaphenes), various phenolic acids, and stilbenes. Humic substances can induce the first step of synthesizing phenylpropanoid-derived compounds catalysed by phenylalanine ammonia lyase [184]. MYB proteins can act as transcriptional activators and repressors that control the synthesis of the phenylpropanoid-derived compounds.
WRKY TFs have been reported to play pivotal roles in defence signalling and in regulating plants' growth and developmental processes [185]. For example, Yang et al. [186] have shown that rice ABI5-Like1 (ABL1) regulates ABA and auxin responses through ABRE-containing WRKY genes, suppressing auxin signalling while enhancing ABA signalling, thereby providing insights into ABA and auxin crosstalk. Furthermore, according to Tripathi et al. [187], the interaction of bZIP transcription factors with promoters from some WRKY gene promoters is crucial for ABA and auxin signalling.
The NAC [for NAM-ATAF1/2-CUC2) transcription factor constitutes one of the most prominent transcription factor families in plant genomes. Roles of many NAC transcription factors have been demonstrated in diverse processes, such as cell cycle control [188] and AtNAC2 functioning in root development [18]. According to Park et al. [189], some NAC proteins are membrane associated, and controlled proteolytic activation of the membrane-bound NAC transcription factors has been proposed to serve as an adaptive strategy that ensures rapid transcriptional responses to abrupt environmental changes. Xie et al. [190] demonstrated that NAC1 is induced by auxin and mediates auxin signalling to promote lateral root development. That is the other typical effect of HA on root traits [70]. Moreover, NAC2 integrates auxin and cytokinin pathways to modulate root development in rice [191], binding of promoters of IAA inactivation-related genes (GH3.6 and GH3.8), IAA signalling-related gene (ARF25), and a cytokinin oxidase gene (CKX4). Phytohormones influence signalling responses by acting in conjunction with or in opposition to each other to maintain cellular homeostasis. Nuruzzaman et al. [192] showed that besides the response to biotic and abiotic stress NAC TFs are also crucial in the phytohormone signalling pathway and reported the NAC involvement on ABA and JA regulation, ABA, and JA/ET and in the ABA-independent pathway of abiotic stress and in regulating biotic stress via an antagonistic JA and SA pathway and can regulate GA3-mediated salt signalling in seed germination [192, 193].
Hormonal crosstalk occurs between Auxin/GA/Cytokinin/ABA/Ethylene/SA through TFs. Both ABA-independent and ABA-dependent signal transduction pathways convert the initial HA signalling to cellular responses. The TF family members involved in both ABA-independent (AP2/ERF, bHLH, and NAC) and ABA-dependent (MYB, bZIP, and MYC) pathways were up-and down-regulated in maize root seedlings by HA, showing high and wide redundancy. A way to interpret and unify this vast redundancy of both genes encoding proteins related to the various hormones and TFs is to consider HA per se as one agent of abiotic stress when applied directly to plant surface [62, 183]. For different reasons, such as the presence of aromatic compounds, dissociation of organic acids, compounds with structures, like plant hormones, and general changes in membrane potential, HA can boost the plant signalling, which finally leads to enhanced growth and crop yield. Srivastava et al. [195] proposed a unified mechanism of action of different plant bioregulators that includes plant redox homeostasis control, which regulates root growth to improve plant water/nutrient status, photosynthetic efficiency, and source–sink homeostasis for enhanced crop yield and metabolism for an overall improvement of plant growth.
The potential role of HA in preventing and restoring the cytosolic redox homeostasis was described by García et al. [12, 14]. After exposure of HA, the general mechanism is triggered to prevent cellular damage by reactive oxygen species (ROS) that induces the synthesis of antioxidative enzymes, such as superoxide dismutase, peroxidases, catalase, and ascorbate peroxidase, and that of non-enzymatic production of scavenging compounds, like ascorbate, tocopherol, and phenolics, are induced. In addition, compatible solutes, such as proline, are also produced to protect cells against ROS accumulation. Lamar [196] elegantly showed the electron shuttling capacity of HA. The HA interacts with plant response via redox activity by relating their electron-donating capacities with processes, such as stimulation of H+-ATPase activity, increase in [Ca2+]cyt, and polarization-dependent Ca2+ channels and membrane depolarization. Finally, Monda et al. [197] also observed that the HA redox effect was determinant for maintaining cell homeostasis in tomatoes under nutritional stress and improving nutrient use efficiency.
Conclusion
Here we proposed a unified mechanism to plant stimulation showing that HA can act as a key regulatory hub in plant hormone crosstalk by modifying plant receptors, phosphatases activity, and changing expression of different TFs. Among the hormones most affected by HA, auxins showed the highest number of up-regulated genes, corroborating previous studies demonstrating the auxin activity of HA. On the other hand, ethylene, ABA, and jasmonic acid, which are hormones commonly involved in stress response, displayed the highest number of down-regulated genes, suggesting a role of HA as a plant growth stimulant instead of an integrator of stress responses. Several specific and non-specific (proton pumps) membrane receptors were sensitized by HA, and different phosphatases were found in the differential transcription level. In addition, most of the synthesis and metabolism of genes encoding hormones have also been elicited by HA, and an expected response on the main Transcription Factors (TFs) and other regulatory genes was observed. Therefore, HA, acting as external cues, mediates plant hormone response using intricately interconnected signalling pathways and facilitates the generation of gene responses regulating plant hormones crosstalk.
Availability of data and materials
Not applicable.
Abbreviations
- AAMT1:
-
Anthranilate O-methyltransferase
- ABA:
-
Abscisic acid
- ABI3:
-
Abscisic acid insensitive 3
- ACS4:
-
1-Aminocyclopropane-1-Carboxylic acid synthase 4
- ACS5:
-
1-Aminocyclopropane-1-Carboxylic acid synthase 5
- AGO:
-
Argonaute
- GTE:
-
Transcription factor group E
- AHK:
-
Histidine kinases
- AHP6:
-
Histidine phosphotransfer protein 6
- Alba:
-
Alba DNA/RNA-binding protein
- ALF:
-
Alfin-like transcription factor
- AP2/ERF:
-
APETALA2/Ethylene-responsive element-binding protein family
- ARF:
-
Auxin Response Factor Family
- ARF:
-
Auxin response factor
- ARR:
-
Two-component response regulator
- ARR1:
-
Two-component response regulator/ARR1 (Arabidopsis Response Regulator)
- ARR5:
-
Two-component response regulator/ARR5 (Arabidopsis Response Regulator)
- AS:
-
Salicylic acid
- Aux/IAA:
-
Auxin/indole-3-acetic acid protein Family
- AUX/IAA:
-
Auxin/indole-3-acetic acid
- AUX:
-
Auxins
- B3:
-
B3 domain-containing protein
- bHLH:
-
Basic Helix-Loop-Helix Family
- BIN2:
-
Brassinosteroid insensitive 2
- BR:
-
Brassinosteroids
- BTB:
-
BTB domain-containing protein
- bZIP:
-
Basic leucine zípper
- BZR1:
-
Brassinazole-resistant 1
- C3H Type:
-
C3H-type transcription factor
- CAMTA:
-
Calmodulin-binding transcription activators
- CK:
-
Cytokinin
- CKX:
-
Cytokinin oxidases/dehydrogenases
- CPK4:
-
Calcium-dependent Protein Kinase 4
- CRF:
-
Chromatin Remodelling Factors
- CtBP:
-
C-terminal-binding protein
- ALF:
-
Alfin-like transcription factor
- DELLA:
-
DELLA protein Family
- EIN3:
-
EIN3-like (EIL) transcription factor Family
- EIN3:
-
Ethylene insensitive 3
- ERF:
-
Ethylene response factor
- ET:
-
Ethylene
- ETR1:
-
Ethylene response1
- F-box:
-
F-box protein
- FLS:
-
Flavonol synthase
- FREE1:
-
Fyve domain protein required for endosomal sorting 1
- GA:
-
Gibberellic acid
- GA20ox:
-
Gibberellin 3-oxidase 1
- GRAS:
-
GRAS transcription factor family (Scarecrow)
- GRFs:
-
GRF transcription factor
- ABH:
-
Alpha/beta hydrolase
- HA:
-
Humic acids
- HB:
-
Homeobox transcription factor Family
- HDAC:
-
Histone deacetylase
- IAA19:
-
Auxin-responsive protein IAA19
- IAA5:
-
Auxin-responsive protein IAA5
- IAR3:
-
Iaa-alanine resistant 3
- IPT:
-
Adenylate isopentenyl transferase
- JA:
-
Jasmonate
- JAZ9:
-
Jasmonate zim-domain 9
- LOB:
-
LOB domain-containing protein
- LOB:
-
LOB domain-containing protein
- Alba:
-
Alba DNA/RNA-binding protein
- LOX:
-
Lipoxygenase
- LSD:
-
Lysine-specific demethylase
- MOB:
-
MOB kinase activator-like
- MTREF:
-
Mitochondrial transcription termination factor family protein
- G-box:
-
G-box protein
- MYC2:
-
MYC2 Proto-Oncogene
- NAC:
-
NAC (NAM, ATAF1,2, and CUC2
- MYB:
-
MYB Proto-Oncogene
- PAT:
-
S-acyltransferase
- PIN:
-
Auxin efflux carrier component 1
- PLATZ:
-
Plant AT-rich sequence and zinc-binding proteins
- PLS:
-
Polaris
- PLT1/2:
-
AP2-like ethylene-responsive transcription factor (plethoras)
- PP2C:
-
2C protein phosphatases
- PPR:
-
Putative pentatricopeptide repeat-containing protein
- PR1:
-
Pathogenesis-related 1
- REMORIN:
-
Remorin family protein
- Ring type E3:
-
RING-type E3 ubiquitin transferase
- SEUSS:
-
Transcriptional corepressor
- SEUSS; C3H Type:
-
C3H-type transcription factor
- SHY2:
-
Auxin-responsive protein
- SUVR5:
-
Histone-lysine N-methyltransferase
- SWI/SNF:
-
SWItch/Sucrose Non-Fermentable
- TCP:
-
TCP protein domain
- TCP:
-
TCP protein domain
- TIR1:
-
Transport inhibitor response
- UBX2:
-
Ubiquitin regulatory X domain-containing protein 2
- WRKY:
-
WRKY Family
References
Zong Y, Chen Z, Innes JB, Chen C, Wang Z, Wang H. Fire and flood management of coastal swamp enabled first rice paddy cultivation in east China. Nature. 2007;449(7161):459–62. https://doi.org/10.1038/nature06135.
Pittelkow CM, Liang X, Linquist BA, van Groenigen KJ, Lee J, Lundy ME, van Gestel N, Six J, Rodney T, Venterea RT, van Kessel C. Productivity limits and potentials of the principles of conservation agriculture. Nature. 2014;517(7534):365–8. https://doi.org/10.1038/nature13809.
Matson PA, Naylor R, Ortiz-Monasterio I. Integration of environmental, agronomic, and economic aspects of fertilizer management. Science. 1998;280:112–5. https://doi.org/10.1126/science.280.5360.112.
Gilliom RJ. Pesticides in U.S. streams and groundwater. Environ Sci Technol. 2007;41(10):3408–14. https://doi.org/10.1021/es072531u.
Chen XP, Cui ZL, Vitousek PM, Cassman KG, Matson PA, Bai JS, Meng QF, Hou P, Yue SC, Römheld V, Zhang FS. Integrated soil-crop system management for food security. Proc Natl Acad Sci USA. 2011;108(16):6399–404. https://doi.org/10.1073/pnas.1101419108.
Hayes MH, Swift RS. Vindication of humic substances as a key component of organic matter in soil and water. Adv Agron. 2020;163:1–37. https://doi.org/10.1016/bs.agron.2020.05.001.
Piccolo A. The supramolecular structure of humic substances. A novel understanding of humus chemistry and implications in soil science. Adv Agron. 2002;75:57–134. https://doi.org/10.1016/S0065-2113(02)75003-7.
Du Jardin P. Plant biostimulants: definition, concept, main categories and regulation. Sci Hortic. 2015;196:3–14. https://doi.org/10.1016/j.scienta.2015.09.021.
Nardi S, Carletti P, Pizzeghello D, Muscolo A. Biological activities of humic substances. In: Senesi N, Xing B, Huang PM, editors. Biophysico-chemical processes involving natural nonliving organic matter in environmental systems, vol. 2. Hoboken: Wiley; 2009. p. 305–9.
Rose MT, Patti AF, Little KR, Brown AL, Jackson WR, Cavagnaro TR. A meta-analysis and review of plant-growth response to humic substances. Adv Agron. 2014. https://doi.org/10.1016/B978-0-12-800138-7.00002-4.
Berbara RL, García AC. Humic substances and plant defense metabolism. In: Ahmad P, Wani M, editors. Physiological mechanisms and adaptation strategies in plants under changing environment. New York: Springer; 2014. https://doi.org/10.1007/978-1-4614-8591-9_11.
García AC, Santos LA, Izquierdo FG, Sperandio ML, Castro RN, Berbara RLL. Vermicompost humic acids as an ecological pathway to protect rice plant against oxidative stress. Ecol Eng. 2012;47:203–8. https://doi.org/10.1016/j.ecoleng.2012.06.011.
García AC, Santos LA, Izquierdo FG, Rumjanek VM, Castro RN, dos Santos FS, Souza LGA, Berbara RLL. Potentialities of vermicompost humic acids to alleviate water stress in rice plants (Oryza sativa L.). J Geochem Explor. 2014;136:48–54. https://doi.org/10.1016/j.gexplo.2013.10.005.
García AC, Santos LA, Ambrósio de Souza LG, Tavares OCH, Zonta E, Gomes ETM, García-Mina JM, Berbara RLL. Vermicompost humic acids modulate the accumulation and metabolism of ROS in rice plants. J Plant Physiol. 2016;192:56–63. https://doi.org/10.1016/j.jplph.2016.01.008.
Aguiar NO, Medici LO, Olivares FL, Dobbss LB, Torres-Netto A, Silva SF, Canellas LP. Metabolic profile and antioxidant responses during drought stress recovery in sugarcane treated with humic acids and endophytic diazotrophic bacteria. Ann Appl Biol. 2016;168:203–13. https://doi.org/10.1111/aab.12256.
Schiavon M, Pizzeghello D, Muscolo A, Vaccaro S, Francioso O, Nardi S. High molecular size humic substances enhance phenylpropanoid metabolism in maize (Zea mays L.). J Chem Ecol. 2010;36:662–9. https://doi.org/10.1007/s10886-010-9790-6.
Trevisan S, Botton A, Vaccaro S, Vezzaro A, Quaggiotti S, Nardi S. Humic substances affect Arabidopsis physiology by altering the expression of genes involved in primary metabolism, growth and development. Environ Exp Bot. 2011;74:45–55. https://doi.org/10.1016/j.envexpbot.2011.04.017.
Jannin L, Arkoun M, Ourry A, Laîneì P, Goux D, Garnica M, et al. Microarray analysis of humic acid effects on Brassica napus growth: involvement of N C and S metabolisms. Plant Soil. 2012;359:297–319. https://doi.org/10.1007/s11104-012-1191-x.
Canellas LP, Canellas NOA, Irineu LE, Olivares FL, Piccolo A. Plant chemical priming by humic acids. Chem Biol Technol Agric. 2020;7:12. https://doi.org/10.1186/s40538-020-00178-4.
Cordeiro FC, Santa-Catarina C, Silveira V, de Souza SR. Humic acid effect on catalase activity and the generation of reactive oxygen species in corn ( I L). Biosci Biotechnol Biochem. 2011;75:70–4. https://doi.org/10.1271/bbb.100553.
Garcia AC, Olaetxea M, Santos LA, Mora V, Baigorri R, Fuentes M, Zamarreño AM, Berbara RL, Garcia-Mina JM. Involvement of hormone- and ROS signaling pathways in the beneficial action of humic substances on plants growing under normal and stressing conditions. BioMed Res Int. 2016. https://doi.org/10.1155/2016/3747501.
Zhao X-C, Schaller GE. Effect of salt and osmotic stress upon expression of the ethylene receptor ETR1 in Arabidopsis thaliana. FEBS Lett. 2004;562(1–3):89–192. https://doi.org/10.1016/S0014-5793(04)00238-8.
Cela J, Chang C, Munné-Bosch S. Accumulation of g- rather than a-tocopherol alters ethylene signaling gene expression in the vte4 mutant of Arabidopsis thaliana. Plant Cell Physiol. 2011;52:1389–400. https://doi.org/10.1093/pcp/pcr085.
Cheng MC, Liao PM, Kuo WW, Lin TP. The Arabidopsis ETHYLENE RESPONSE FACTOR1 regulates abiotic stress-responsive gene expression by binding to different cis-acting elements in response to different stress signals. Plant Physiol. 2013;162(3):1566–82. https://doi.org/10.1104/pp.113.221911.
Wu L, Chen X, Ren H, Zhang Z, Zhang H, Wang J, Wang XC, Huang R. ERF protein JERF1 that transcriptionally modulates the expression of abscisic acid biosynthesis-related gene enhances the tolerance under salinity and cold in tobacco. Planta. 2007;226(4):815–25. https://doi.org/10.1007/s00425-007-0528-9.
Jayakannan M, Bose J, Babourina O, Shabala S, Massart A, Poschenrieder C, Rengel Z. The NPR1-dependent salicylic acid signalling pathway is pivotal for enhanced salt and oxidative stress tolerance in Arabidopsis. J Exp Bot. 2015;66(7):1865–75. https://doi.org/10.1093/jxb/eru528.
Magome H, Yamaguchi S, Hanada A, Kamiya Y, Oda K. The DDF1 transcriptional activator upregulates expression of a gibberellin-deactivating gene, GA2ox7, under high-salinity stress in Arabidopsis. Plant J. 2008;56(4):613–26. https://doi.org/10.1111/j.1365-313X.2008.03627.x.
Wu X, He J, Chen J, Yang S, Zha D. Alleviation of exogenous 6-benzyladenine on two genotypes of eggplant (Solanum melongena Mill.) growth under salt stress. Protoplasma. 2014;251(1):169–76. https://doi.org/10.1007/s00709-013-0535-6.
He X-J, Mu R-L, Cao W-H, Zhang Z-G, Zhang J-S, Chen S-Y. AtNAC2, a transcription factor downstream of ethylene and auxin signaling pathways, is involved in salt stress response and lateral root development. Plant J. 2005;44(6):903–16. https://doi.org/10.1111/j.1365-313X.2005.02575.x.
Creelman RA, Mullet JE. Jasmonic acid distribution and action in plants: regulation during development and response to biotic and abiotic stress. Proc Natl Acad Sci USA. 1995;92(10):4114–9. https://doi.org/10.1073/pnas.92.10.4114.
Divi UK, Rahman T, Krishna P. Brassinosteroid-mediated stress tolerance in Arabidopsis shows interactions with abscisic acid, ethylene and salicylic acid pathways. BMC Plant Biol. 2010;10(1):151. https://doi.org/10.1186/1471-2229-10-151.
Nardi S, Pizzeghello D, Ertani A. Hormone-like activity of the soil organic matter. Appl Soil Ecol. 2018;123:517–20. https://doi.org/10.1016/j.apsoil.2017.04.020.
Nardi S, Panuccio MR, Abenavoli MR, Muscolo A. Auxin-like effect of humic substances extracted from faeces of Allolobophora caliginosa and A. rosea. Soil Biol Biochem. 1994;26(10):1341–6. https://doi.org/10.1016/0038-0717(94)90215-1.
Muscolo A, Cutrupi S, Nardi S. IAA detection in humic substances. Soil Biol Biochem. 1998;30:1199–201. https://doi.org/10.1016/S0038-0717(98)00005-4.
Pizzeghello D, Nicolini G, Nardi S. Hormone-like activity of humic substances in Fagus sylvaticae forests. New Phytol. 2001;151(3):647–57. https://doi.org/10.1046/j.0028-646x.2001.00223.x.
Pizzeghello D, Nicolini G, Nardi S. Hormone-like activities of humic substances in different forest ecosystems. New Phytol. 2002;155(3):393–402. https://doi.org/10.1046/j.1469-8137.2002.00475.x.
Mora V, Bacaicoa E, Zamarreno AM, Aguirre E, Garnica M, Fuentes M, Garcia-Mina JM. Action of humic acid on promotion of cucumber shoot growth involves nitrate-related changes associated with the root-to-shoot distribution of cytokinins, polyamines and mineral nutrients. J Plant Physiol. 2010;167:633–42. https://doi.org/10.1016/j.jplph.2009.11.018.
Pizzeghello D, Francioso O, Ertani A, Muscolo A, Nardi S. Isopentenyladenosine and cytokinin-like activity of different humic substances. J Geochem Explor. 2013;129:70–5. https://doi.org/10.1016/j.gexplo.2012.10.007.
Ali AYA, Ibrahim MEH, Zhou G, Nimir NEA, Jiao X, Zhu G, Elsiddig AMI, Suliman MSE, Elradi SBM, Yue W. Exogenous jasmonic acid and humic acid increased salinity tolerance of sorghum. Agron J. 2020. https://doi.org/10.1002/agj2.20072.
De Hita D, Fuentes M, Fernández V, Zamarreño AM, Olaetxea M, García-Mina JM. Discriminating the short-term action of root and foliar application of humic acids on plant growth: emerging role of jasmonic acid. Front Plant Sci. 2020;11:493. https://doi.org/10.3389/fpls.2020.00493.
Zandonadi DB, Matos CRR, Castro RN, Spaccini R, Olivares FL, Canellas LP. Alkamides: a new class of plant growth regulators linked to humic acid bioactivity. Chem Biol Technol Agric. 2019;6:23. https://doi.org/10.1186/s40538-019-0161-4.
Zandonadi DB, Santos MP, Dobbss LB, Olivares FL, Canellas LP, Binzel ML, Okorokova-Façanha AL, Façanha AR. Nitric oxide mediates humic acids-induced root development and plasma membrane H+-ATPase activation. Planta. 2010;231(5):1025–36. https://doi.org/10.1007/s00425-010-1106-0.
Furlani PR, Clark RB. Screening sorghum for aluminum tolerance in nutrient solutions1. Agron J. 1981;73:587–94.
Shah ZH, Rehman HM, Akhtar T, Alsamadany H, Hamooh BT, Mujtaba T, Daur I, Al Zahrani Y, Alzahrani HAS, Ali S, Yang SH, Chung G. Humic substances: determining potential molecular regulatory processes in plants. Front Plant Sci. 2018. https://doi.org/10.3389/fpls.2018.00263.
Chen Y, Hoehenwarter W. Changes in the phosphoproteome and metabolome link early signaling events to rearrangement of photosynthesis and central metabolism in salinity and oxidative stress response in Arabidopsis. Plant Physiol. 2015;169:3021–33. https://doi.org/10.1104/pp.15.01486.
Crasnier M, Noat G, Ricard J. Purification and molecular properties of acid phosphatase from sycamore cell walls. Plant Cell Environ. 1980;3(3):17–224. https://doi.org/10.1111/1365-3040.ep11581714.
Schenk G, Mitić N, Hanson GR, Comba P. Purple acid phosphatase: a journey into the function and mechanism of a colorful enzyme. Coord Chem Rev. 2013;257(2):473–82. https://doi.org/10.1016/j.ccr.2012.03.020.
Sergeant K, Printz B, Guerriero G, Renaut J, Lutts S, Hausman JF. The Dynamics of the cell wall proteome of developing alfalfa stems. Biology (Basel). 2019;8(3):60. https://doi.org/10.3390/biology8030060.
Howles PA, Birch RJ, Collings DA, et al. A mutation in an Arabidopsis ribose 5-phosphate isomerase reduces cellulose synthesis and is rescued by exogenous uridine. Plant J. 2006;48(4):606–18. https://doi.org/10.1111/j.1365-313X.2006.02902.x.
Hauschild R, Schaewen AV. Differential regulation of glucose-6-phosphate dehydrogenase isoenzyme activities in potato. Plant Physiol. 2003;133(1):47–62. https://doi.org/10.1104/pp.103.025676.
Allan WL, Clark SM, Hoover GJ, Shelp BJ. Role of plant glyoxylate reductases during stress: a hypothesis. Biochem J. 2009;423(1):15–22. https://doi.org/10.1042/BJ20090826.
Guo J, Polymenis M. Dcr2 targets Ire1 and downregulates the unfolded protein response in Saccharomyces cerevisiae. EMBO Rep. 2006;7(11):1124–7. https://doi.org/10.1038/sj.embor.7400813.
Paradis S, Villasuso AL, Aguayo SS, Maldiney R, Habricot Y, Zalejski C, Machado E, Sott B, Miginia E, Jeannette E. Arabidopsis thaliana lipid phosphate phosphatase 2 is involved in abscisic acid signalling in leaves. Plant Physiol Biochem. 2011;49(3):357–62. https://doi.org/10.1016/j.plaphy.2011.01.010.
Munnik T, Vermeer JEM. Osmotic stress-induced phosphoinositide and inositol phosphate signalling in plants. Plant Cell Environ. 2010;33(4):655–69. https://doi.org/10.1111/j.1365-3040.2009.02097.x.
Luo Y, Qin G, Zhang J, Liang Y, Song Y, Zhao M, Qu L-J. d-myo-inositol-3-phosphate affects phosphatidylinositol-mediated endomembrane function in Arabidopsis and is essential for auxin-regulated embryogenesis. Plant Cell. 2011;23(4):1352–72. https://doi.org/10.1105/tpc.111.083337.
Lenk C, Köllner TG, Erb M, Degenhardt J. Two enzymes responsible for the formation of herbivore-induced volatiles of maize, the methyltransferase AAMT1 and the terpene synthase TPS23, are regulated by a similar signal transduction pathway. Entomol Exp Appl. 2012;144(1):86–92. https://doi.org/10.1111/j.1570-7458.2012.01240.x.
Brady SM, Sarkar SF, Bonetta D, McCourt P. The ABSCISIC ACID INSENSITIVE 3 (ABI3) gene is modulated by farnesylation and is involved in auxin signaling and lateral root development in Arabidopsis. Plant J. 2003;34(1):67–75. https://doi.org/10.1046/j.1365-313x.2003.01707.x.
El-Showk S, Ruonala R, Helariutta Y. Crossing paths: cytokinin signalling and crosstalk. Development. 2013;140(7):1373–83. https://doi.org/10.1242/dev.086371.
Liu J, Rowe J, Lindsey K. Hormonal crosstalk for root development: a combined experimental and modeling perspective. Front Plant Sci. 2014;5:116. https://doi.org/10.3389/fpls.2014.00116.
Liu J, Moore S, Chen C, Lindsey K. Crosstalk complexities between auxin, cytokinin, and ethylene in arabidopsis root development: from experiments to systems modeling, and back again. Mol Plant. 2017;10(12):1480–96. https://doi.org/10.1016/j.molp.2017.11.002.
Jang G, Yoon Y, Choi YD. Crosstalk with jasmonic acid integrates multiple responses in plant development. Int J Mol Sci. 2020;21(1):305. https://doi.org/10.3390/ijms21010305.
Liscum E, Reed JW. Genetics of Aux/IAA and ARF action in plant growth and development. Plant Mol Biols. 2002;49(3–4):387–400. https://doi.org/10.1007/978-94-010-0377-3_10.
Galstyan A, Nemhauser JL. Auxin promotion of seedling growth via ARF5 is dependent on the brassinosteroid-regulated transcription factors BES1 and BEH4. Plant Direct. 2019. https://doi.org/10.1002/pld3.166.
Vert G, Walcher CL, Chory J, Nemhauser JL. Integration of auxin and brassinosteroid pathways by auxin response factor 2. Proc Natl Acad Sci USA. 2008;105(28):9829–34. https://doi.org/10.1073/pnas.0803996105.
Kono A, Yin Y. Updates on BES1/BZR1 regulatory networks coordinating plant growth and stress responses. Front Plant Sci. 2020;11: 617162. https://doi.org/10.3389/fpls.2020.617162.
Nishiyama R, Watanabe Y, Fujita Y, et al. Analysis of cytokinin mutants and regulation of cytokinin metabolic genes reveals important regulatory roles of cytokinins in drought, salt and abscisic acid responses, and abscisic acid biosynthesis. Plant Cell. 2011;23(6):2169–83. https://doi.org/10.1105/tpc.111.087395.
Müller M. Foes or friends: ABA and ethylene interaction under abiotic stress. Plants (Basel). 2021;10(3):448. https://doi.org/10.3390/plants10030448.
Song S, Qi T, Wasternack C, Xie D. Jasmonate signaling and crosstalk with gibberellin and ethylene. Curr Opin Plant Biol. 2014;21:112–9. https://doi.org/10.1016/j.pbi.2014.07.005.
Lewis DR, Ramirez MV, Miller ND, et al. Auxin and ethylene induce flavonol accumulation through distinct transcriptional networks. Plant Physiol. 2011;156(1):144–64. https://doi.org/10.1104/pp.111.172502.
Kou E, Huang X, Zhu Y, Kou SuW, Liu H, Sun G, Song S. Crosstalk between auxin and gibberellin during stalk elongation in flowering Chinese cabbage. Sci Rep. 2021;11:3976. https://doi.org/10.1038/s41598-021-83519-z.
Chandler JW. Auxin as compère in plant hormone crosstalk. Planta. 2009;231(1):1–12. https://doi.org/10.1007/s00425-009-1036-x.
Manohar M, Wang D, Manosalva PM, Choi HW, Kombrink E, Klessig DF. Members of the abscisic acid co-receptor PP2C protein family mediate salicylic acid-abscisic acid crosstalk. Plant Direct. 2017;1(5): e00020. https://doi.org/10.1002/pld3.20.
Zubo YO, Schaller GE. Role of the cytokinin-activated type-B response regulators in hormone crosstalk. Plants (Basel). 2020;9(2):166. https://doi.org/10.3390/plants9020166.
Baía DC, Olivares FL, Zandonadi DB, De Paula SC, Spaccini R, Canellas LP. Humic acids trigger the weak acids stress response in maize seedlings. Chem Biol Technol Agric. 2020;7(1):1–13. https://doi.org/10.1186/s40538-020-00193-5.
Trewavas A, Read N, Campbell AK, Knight M. Transduction of Ca2+ signals in plant cells and compartmentalization of the Ca2+ signal. Biochem Soc Trans. 1996;24(4):971–4. https://doi.org/10.1042/bst0240971.
Trewavas A. Post-translational modification of proteins by phosphorylation. Annu Rev Plant Physiol. 1976;27(1):349–74. https://doi.org/10.1146/annurev.pp.27.060176.
Trewavas AJ. The foundations of plant intelligence. Interface Focus. 2017;7:20160098. https://doi.org/10.1098/rsfs.2016.0098.
Nunes RO, Domiciano GA, Alves WS, Melo ACA, Nogueira FCS, Canellas LP, Soares MR. Evaluation of the effects of humic acids on maize root architecture by label-free proteomics analysis. Sci Rep. 2019. https://doi.org/10.1038/s41598-019-48509-2.
Ding X, Cao Y, Huang L, Zhao J, Xu C, Li X, Wang S. Activation of the Indole-3-acetic acid-amido synthetase GH3-8 suppresses expansin expression and promotes salicylate- and jasmonate-independent basal immunity in rice. Plant Cell. 2008;20(1):228–40. https://doi.org/10.1105/tpc.107.055657.
LeClere S, Tellez R, Rampey RA, Matsuda SPT, Bartel B. Characterization of a family of IAA-amino acid conjugate hydrolases from Arabidopsis. J Biol Chem. 2002;277:20446–52. https://doi.org/10.1074/jbc.M111955200.
Jain M, Tyagi AK, Khurana JP. Genome-wide analysis, evolutionary expansion, and expression of early auxin-responsive SAUR gene family in rice (Oryza sativa). Genomics. 2006;88(3):360–71. https://doi.org/10.1016/j.ygeno.2006.04.008.
Canellas LP, Façanha AO, Olivares FL, Façanha AR. Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence, and plasma membrane H+-ATPase activity in maize roots. Plant Physiol. 2002;130:1951–7. https://doi.org/10.1104/pp.007088.
Quaggiotti S, Ruperti B, Pizzeghello D, Francioso O, Tugnoli V, Nardi S. Effect of low molecular size humic substances on nitrate uptake and expression of genes involved in nitrate transport in maize (Zea mays L.). J Exp Bot. 2004;55(398):803–13. https://doi.org/10.1093/jxb/erh085.
Zandonadi DB, Canellas LP, Façanha AR. Indolacetic and humic acids induce lateral root development through a concerted plasmalemma and tonoplast H+ pumps activation. Planta. 2007;225(6):1583–95. https://doi.org/10.1007/s00425-006-0454-2.
He JX, Gendron JM, Sun Y, et al. BZR1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses. Science. 2005;308(5729):1743. https://doi.org/10.1126/science.1107580.
Yin Y, Vafeados D, Tao Y, Yoshida S, Asami T, Chory J. A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell. 2005;20:249–59. https://doi.org/10.1016/j.cell.2004.11.044.
Parker G, Schofield R, Sundberg B, Turner S. Isolation of COV1, a gene involved in the regulation of vascular patterning in the stem of Arabidopsis. Development. 2003;130(10):2139–48. https://doi.org/10.1242/dev.00441.
Adamowski M, Friml J. PIN-dependent auxin transport: action, regulation, and evolution. Plant Cell. 2015;27:20–32. https://doi.org/10.1105/tpc.114.134874.
Yamada R, Murai K, Uchida N, Takahashi K, Iwasaki R, Tada Y, Kinoshita T, Itami K, Torii KU, Hagihara S. A super strong engineered auxin–TIR1 pair. Plant Cell Physiol. 2018;59:1538–44. https://doi.org/10.1093/pcp/pcy127.
Kurz T, Chou Y-C, Willems AR, Meyer-Schaller N, Hecht M-L, Tyers M, Peter M, Sicheri F. Dcn1 functions as a scaffold-type E3 ligase for cullin neddylation. Mol Cell. 2008;29(1):23–35. https://doi.org/10.1016/j.molcel.2007.12.012.
Tan X, Calderon-Villalobos LIA, Sharon M, Zheng C, Robinson CV, Estelle M, Zheng N. Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature. 2007;446(7136):640–5. https://doi.org/10.1038/nature05731.
Lim EK, Bowles DJ. A class of plant glycosyltransferases involved in cellular homeostasis. EMBO J. 2004;23(15):2915–22. https://doi.org/10.1038/sj.emboj.7600295.
Yu J, Sun H, Zhang J, Hou Y, Zhang T, Kang J, Wang Z, Long R. Analysis of aldo-keto reductase gene family and their responses to salt, drought, and abscisic acid stresses in Medicago truncatula. Int J Mol Sci. 2020;21(3):754. https://doi.org/10.3390/ijms21030754.
Preger V, Tango N, Marchand C, Lemaire SD, Carbonera D, Di Valentin M, Costa A, Pupillo P, Trost P. Auxin-responsive genes AIR12 code for a new family of plasma membrane b-type cytochromes specific to flowering plants. Plant Physiol. 2009;150(2):606–20. https://doi.org/10.1104/pp.109.139170.
Aguiar NO, Olivares FL, Novotny EH, Canellas LP. Changes in metabolic profiling of sugarcane leaves induced by endophytic diazotrophic bacteria and humic acids. PeerJ. 2018;6:5445. https://doi.org/10.7717/peerj.5445.
Lynch T, Erickson BJ, Finkelstein RR. Direct interactions of ABA-insensitive(ABI)-clade protein phosphatase(PP)2Cs with calcium-dependent protein kinases and ABA response element-binding bZIPs may contribute to turning off ABA response. Plant Mol Biol. 2012;80(6):647–58. https://doi.org/10.1007/s11103-012-9973-3.
Liu L, Li N, Yao C, Meng S, Song C. Functional analysis of the ABA-responsive protein family in ABA and stress signal transduction in Arabidopsis. Chin Sci Bull. 2013;58(31):3721–30. https://doi.org/10.1007/s11434-013-5941-9.
Guo Y, Xiong L, Song CP, Gong D, Halfter U, Zhu JK. A calcium sensor and its interacting protein kinase are global regulators of abscisic acid signaling in Arabidopsis. Dev Cell. 2002;3(2):233–44. https://doi.org/10.1016/s1534-5807(02)00229-0.
Kinoshita T, Shimazaki K. Blue light activates the plasma membrane H(+)-ATPase by phosphorylation of the C-terminus in stomatal guard cells. EMBO J. 1999;18(20):5548–58. https://doi.org/10.1093/emboj/18.20.5548.
Shimazaki K, Iino M, Zeiger E. Blue light-dependent proton extrusion by guard-cell protoplasts of Vicia faba. Nature. 1986;319:324–6. https://doi.org/10.1038/319324a0.
Schroeder JI, Allen GJ, Hugouvieux V, Kwak JM, Waner D. Guard cell signal transduction. Annu Rev Plant Physiol Plant Mol Biol. 2001;52:627–58. https://doi.org/10.1146/annurev.arplant.52.1.627.
Finkelstein R. Abscisic acid synthesis and response. Arabidopsis Book. 2013;11: e0166. https://doi.org/10.1199/tab.0166.
Seo M, Peeters AJ, Koiwai H, et al. The Arabidopsis aldehyde oxidase 3 (AAO3) gene product catalyzes the final step in abscisic acid biosynthesis in leaves. Proc Natl Acad Sci USA. 2000;97(23):12908–13. https://doi.org/10.1073/pnas.220426197.
Seo M, Koiwai H, Akaba S, et al. Abscisic aldehyde oxidase in leaves of Arabidopsis thaliana. Plant J. 2000;23(4):481–8. https://doi.org/10.1046/j.1365-313x.2000.00812.x.
Koshiba T, Matsuyama H. An in vitro system of indole-3-acetic acid formation from tryptophan in maize (Zea mays) coleoptile extracts. Plant Physiol. 1993;102(4):1319–24. https://doi.org/10.1104/pp.102.4.1319.
Téllez-Robledo B, Manzano C, Saez A, et al. The polyadenylation factor FIP1 is important for plant development and root responses to abiotic stresses. Plant J. 2019;99(6):1203–19. https://doi.org/10.1111/tpj.14416.
Wang C, Zhang W, Li Z, et al. FIP1 plays an important role in nitrate signaling and regulates CIPK8 and CIPK23 expression in Arabidopsis. Front Plant Sci. 2018;9:593. https://doi.org/10.3389/fpls.2018.00593.
Ohashi Y, Oka A, Rodrigues-Pousada R, et al. Modulation of phospholipid signaling by GLABRA2 in root-hair pattern formation. Science. 2003;300(5624):1427–30. https://doi.org/10.1126/science.1083695.
Wang X, Jing Y, Zhang B, Zhou Y, Lin R. Glycosyltransferase-like protein ABI8/ELD1/KOB1 promotes Arabidopsis hypocotyl elongation through regulating cellulose biosynthesis. Plant Cell Environ. 2015;38(3):411–22. https://doi.org/10.1111/pce.12395.
Wuddineh WA, Mazarei M, Zhang J, et al. Identification and overexpression of gibberellin 2-oxidase (GA2ox) in switchgrass (Panicum virgatum L.) for improved plant architecture and reduced biomass recalcitrance. Plant Biotechnol J. 2015;13(5):636–47. https://doi.org/10.1111/pbi.12287.
Helliwell CA, Poole A, James Peacock W, Dennis ES. Arabidopsisent-kaurene oxidase catalyzes three steps of gibberellin biosynthesis. Plant Physiol. 1999;119(2):507–10. https://doi.org/10.1104/pp.119.2.507.
Otomo K, Kenmoku H, Oikawa H, et al. Biological functions of ent- and syn-copalyl diphosphate synthases in rice: key enzymes for the branch point of gibberellin and phytoalexin biosynthesis. Plant J. 2004;39(6):886–93. https://doi.org/10.1111/j.1365-313X.2004.02175.x.
Hirano K, Nakajima M, Asano K, et al. The GID1-mediated gibberellin perception mechanism is conserved in the Lycophyte Selaginella moellendorffii but not in the Bryophyte Physcomitrella patens. Plant Cell. 2007;19(10):3058–79. https://doi.org/10.1105/tpc.107.051524.
Zhang Z-L, Ogawa M, Fleet CM, Zentella R, Hu J, Heo J-O, Lim J, Kamiyab Y, Yamaguchi S, Sun T. SCARECROW-LIKE 3 promotes gibberellin signaling by antagonizing master growth repressor DELLA in Arabidopsis. Proc Natl Acad Sci. 2011;108(5):2160–5. https://doi.org/10.1073/pnas.1012232108.
Nahirñak V, Almasia NI, Hopp HE, Vazquez-Rovere C. Snakin/GASA proteins. Plant Signal Behav. 2012;7(8):1004–8. https://doi.org/10.4161/psb.20813.
Zhong C, Xu H, Ye S, Wang S, Li L, Zhang S, Wang X. AtGASA6 serves as an integrator of gibberellin-, abscisic acid- and glucose-signaling during seed germination in Arabidopsis. Plant Physiol. 2015. https://doi.org/10.1104/pp.15.00858.
Van de Poel B, Smet D, Van Der Straeten D. Ethylene and hormonal cross talk in vegetative growth and development. Plant Physiol. 2015;169:61–72. https://doi.org/10.1104/pp.15.00724.
Houben M, Van de Poel B. 1-aminocyclopropane-1-carboxylic acid oxidase (ACO): the enzyme that makes the plant hormone ethylene. Front Plant Sci. 2019;10:695. https://doi.org/10.3389/fpls.2019.00695.
Müller M, Munné-Bosch S. Ethylene response factors: a key regulatory hub in hormone and stress signaling. Plant Physiol. 2015;169:32–41. https://doi.org/10.1104/pp.15.00677.
Chen HY, Hsieh EJ, Cheng MC, Chen CY, Hwang SY, Lin TP. ORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47) regulates jasmonic acid and abscisic acid biosynthesis and signaling through binding to a novel cis-element. New Phytol. 2016;211(2):599–613. https://doi.org/10.1111/nph.13914.
Jaimes-Miranda F, Chávez Montes RA. The plant MBF1 protein family: a bridge between stress and transcription. J Exp Bot. 2020;71(6):1782–91. https://doi.org/10.1093/jxb/erz525.
Cui B, Pan Q, Clarke D, Villarreal MO, Umbreen S, Yuan B, Shan W, Jiang J, Loake GJ. S-nitrosylation of the zinc finger protein SRG1 regulates plant immunity. Nat Commun. 2018. https://doi.org/10.1038/s41467-018-06578-3.
Yang X, Bai Y, Shang J, Xin R, Tang W. The antagonistic regulation of abscisic acid-inhibited root growth by brassinosteroids is partially mediated via direct suppression of ABSCISIC ACID INSENSITIVE 5 expression by BRASSINAZOLE RESISTANT 1. Plant Cell Environ. 2016;39(9):1994–2003. https://doi.org/10.1111/pce.12763.
Nolan TM, Vukašinović N, Liu D, Russinova E, Yin Y. Brassinosteroids: multidimensional regulators of plant growth, development, and stress responses. Plant Cell. 2020;32(2):295–318. https://doi.org/10.1105/tpc.19.00335.
Mok DW, Mok MC. Cytokinin metabolism and action. Annu Rev Plant Physiol Plant Mol Biol. 2001;52:89–118. https://doi.org/10.1146/annurev.arplant.52.1.89.
West AH, Stock AM. Histidine kinases and response regulator proteins in two-component signaling systems. Trends Biochem Sci. 2001;26(6):369–76. https://doi.org/10.1016/s0968-0004(01)01852-7.
Taki N, Sasaki-Sekimoto Y, Obayashi T, et al. 12-oxo-phytodienoic acid triggers expression of a distinct set of genes and plays a role in wound-induced gene expression in Arabidopsis. Plant Physiol. 2005;139(3):1268–83. https://doi.org/10.1104/pp.105.067058.
Druege U, Franken P, Hajirezaei MR. Plant hormone homeostasis, signaling, and function during adventitious root formation in cuttings. Front Plant Sci. 2016;7:381. https://doi.org/10.3389/fpls.2016.00381.
Ambawat S, Sharma P, Yadav NR, Yadav RC. MYB transcription factor genes as regulators for plant responses: an overview. Physiol Mol Biol Plants. 2013;19(3):307–21. https://doi.org/10.1007/s12298-013-0179-1.
Sawa S, Ohgishi M, Goda H, Higuchi K, Shimada Y, Yoshida S, Koshiba T. The HAT2 gene, a member of the HD-Zip gene family, isolated as an auxin inducible gene by DNA microarray screening, affects auxin response in Arabidopsis. Plant J. 2002;32:1011–22. https://doi.org/10.1046/j.1365-313x.2002.01488.x.
Johannesson H, Wang Y, Hanson J, Engström P. The Arabidopsis thaliana homeobox gene ATHB5 is a potential regulator of abscisic acid responsiveness in developing seedlings. Plant Mol Biol. 2003;51:719–29. https://doi.org/10.1023/A:1022567625228.
Lin Z, Hong Y, Yin M, Li C, Zhang K, Grierson D. A tomato HD-Zip homeobox protein, LeHB-1, plays an important role in floral organogenesis and ripening. Plant J. 2008;55(2):301–10. https://doi.org/10.1111/j.1365-313X.2008.03505.x.
Yin J, Chang X, Kasuga T, Bui M, Reid MS, Jiang CZ. A basic helix-loop-helix transcription factor, PhFBH4, regulates flower senescence by modulating ethylene biosynthesis pathway in petunia. Hortic Res. 2015;2:15059. https://doi.org/10.1038/hortres.2015.59.
Kumar P, Parveen A, Sharma H, et al. Understanding the regulatory relationship of abscisic acid and bZIP transcription factors towards amylose biosynthesis in wheat. Mol Biol Rep. 2021;48(3):2473–83. https://doi.org/10.1007/s11033-021-06282-4.
Alves M, Dadalto S, Gonçalves A, De Souza G, Barros V, Fietto L. Plant bZIP transcription factors responsive to pathogens: a review. Int J Mol Sci. 2013;14(4):7815–28. https://doi.org/10.3390/ijms14047815.
Hirsch S, Oldroyd GED. GRAS-domain transcription factors that regulate plant development. Plant Signal Behav. 2009;4(8):698–700. https://doi.org/10.4161/psb.4.8.9176.
Liu M, Sun W, Li C, Yu G, Li J, Wang Y, Wang X. A multilayered cross-species analysis of GRAS transcription factors uncovered their functional networks in plant adaptation to the environment. J Adv Res. 2020;29:191–205. https://doi.org/10.1016/j.jare.2020.10.004.
Kumari P, Kakkar M, Gahlaut V, Jaiswal V, Kumar S. Multifarious roles of GRAS transcription factors in plants. Preprints Org. 2021. https://doi.org/10.20944/preprints202103.0066.v1.
Phukan UJ, Jeena GS, Tripathi V, Shukla RK. Regulation of Apetala2/ethylene response factors in plants. Front Plant Sci. 2017. https://doi.org/10.3389/fpls.2017.00150.
Nan H, Gao L. Genome-wide analysis of WRKY genes and their response to hormone and mechanic stresses in carrot. Front Genetics. 2019. https://doi.org/10.3389/fgene.2019.00363.
Li W, Wang H, Yu D. Arabidopsis WRKY transcription factors WRKY12 and WRKY13 oppositely regulate flowering under short-day conditions. Mol Plant. 2016;9(11):1492–503. https://doi.org/10.1016/j.molp.2016.08.003.
Li S-B, Xie Z-Z, Hu C-G, Zhang J-Z. A review of auxin response factors (ARFs) in plants. Front Plant Sci. 2016. https://doi.org/10.3389/fpls.2016.00047.
Xing H, Fu X, Yang C, et al. Genome-wide investigation of pentatricopeptide repeat gene family in poplar and their expression analysis in response to biotic and abiotic stresses. Sci Rep. 2018;8(1):2817. https://doi.org/10.1038/s41598-018-21269-1.
Yuan X, Zhang S, Sun M, Liu S, Qi B, Li X. Putative DHHC-cysteine-rich domain S-acyltransferase in plants. PLoS ONE. 2013;8(10): e75985. https://doi.org/10.1371/journal.pone.0075985.
Jensen MK, Kjaersgaard T, Petersen K, Skriver K. NAC genes: time-specific regulators of hormonal signaling in Arabidopsis. Plant Signal Behav. 2010;5(7):907–10. https://doi.org/10.4161/psb.5.7.12099.
Caro E, Stroud H, Greenberg MV, Bernatavichute YV, Feng S, Groth M, Vashisht AA, Wohlschlegel J, Jacobsen SE. The SET-domain protein SUVR5 mediates H3K9me2 deposition and silencing at stimulus response genes in a DNA methylation-independent manner. PLoS Genet. 2012;8(10): e1002995. https://doi.org/10.1371/journal.pgen.1002995.
Horák J, Grefen C, Berendzen KW, Hahn A, Stierhof Y-D, Stadelhofer B, Stahl M, Koncz C, Harter K. The Arabidopsis thaliana response regulator ARR22 is a putative AHP phospho-histidine phosphatase expressed in the chalaza of developing seeds. BMC Plant Biol. 2008;8:77. https://doi.org/10.1186/1471-2229-8-77.
Badawi MA, Agharbaoui Z, Zayed M, et al. Genome-wide identification and characterization of the wheat remorin (TaREM) family during cold acclimation. Plant Genome. 2019. https://doi.org/10.3835/plantgenome2018.06.0040.
Zhang Y, Li Z, Ma B, Hou Q, Wan X. Phylogeny and functions of LOB domain proteins in plants. Int J Mol Sci. 2020;21(7):2278. https://doi.org/10.3390/ijms21072278.
Ahmad B, Zhang S, Yao J, Rahman MU, Hanif M, Zhu Y, Wang X. Genomic organization of the B3-domain transcription factor family in grapevine (Vitis vinifera L.) and expression during seed development in seedless and seeded cultivars. Int J Mol Sci. 2019;20(18):4553. https://doi.org/10.3390/ijms20184553.
Misra A, McKnight TD, Mandadi KK. Bromodomain proteins GTE9 and GTE11 are essential for specific BT2-mediated sugar and ABA responses in Arabidopsis thaliana. Plant Mol Biol. 2018;96:393–402. https://doi.org/10.1007/s11103-018-0704-2.
Toyoda Y, Matsunaga S. Lysine-specific demethylase epigenetically regulates human and plant phenomena. Cytologia. 2019;84(4):295–8. https://doi.org/10.1508/cytologia.84.295.
Fu Y, Cheng M, Li M, Guo X, Wu Y, Wang J. Identification and characterization of PLATZ transcription factors in wheat. Int J Mol Sci. 2020;21(23):8934. https://doi.org/10.3390/ijms21238934.
Yin X, Gao Y, Song S, Hassani D, Lu J. Identification, characterization and functional analysis of grape (Vitis vinifera L.) mitochondrial transcription termination factor (mTERF) genes in responding to biotic stress and exogenous phytohormone. BMC Genomics. 2021;22(1):136. https://doi.org/10.1186/s12864-021-07446-z.
Mandadi KK, Misra A, Ren S, McKnight TD. BT2, a BTB protein, mediates multiple responses to nutrients, stresses, and hormones in Arabidopsis. Plant Physiol. 2009;150(4):1930–9. https://doi.org/10.1104/pp.109.139220.
Verma JK, Wardhan V, Singh D, Chakraborty S, Chakraborty N. Genome-wide identification of the alba gene family in plants and stress-responsive expression of the rice alba genes. Genes (Basel). 2018;9(4):183. https://doi.org/10.3390/genes9040183.
Bao F, Azhakanandam S, Franks RG. SEUSS and SEUSS-LIKE transcriptional adaptors regulate floral and embryonic development in Arabidopsis. Plant Physiol. 2010;152(2):821–36. https://doi.org/10.1104/pp.109.146183.
Kayum MA, Kim HT, Nath UK, Park JI, Kho KH, Cho YG, I-Sup N,. Research on biotic and abiotic stress related genes exploration and prediction in Brassica rapa and B. oleracea: a review. Plant Breed Biotech. 2016;4(2):135–44. https://doi.org/10.9787/PBB.2016.4.2.135.
Yu H, Wu J, Xu N, Peng M. Roles of F-box proteins in plant hormone responses. Acta Biochim Biophys Sin (Shanghai). 2007;39(12):915–22. https://doi.org/10.1111/j.1745-7270.2007.00358.x.
Liu C, Xin Y, Xu L, et al. Arabidopsis ARGONAUTE 1 binds chromatin to promote gene transcription in response to hormones and stresses. Dev Cell. 2018;44(3):348-361.e7. https://doi.org/10.1016/j.devcel.2017.12.002.
Sorin C, Bussell JD, Camus I, et al. Auxin and light control of adventitious rooting in Arabidopsis require ARGONAUTE1. Plant Cell. 2005;17(5):1343–59. https://doi.org/10.1105/tpc.105.031625.
Liu X, Yang S, Zhao M, et al. Transcriptional repression by histone deacetylases in plants. Mol Plant. 2014;7(5):764–72. https://doi.org/10.1093/mp/ssu033.
Yue R, Lu C, Sun T, et al. Identification and expression profiling analysis of calmodulin-binding transcription activator genes in maize (Zea mays L.) under abiotic and biotic stresses. Front Plant Sci. 2015;6:576. https://doi.org/10.3389/fpls.2015.00576.
Liu L, Xu W, Hu X, Liu H, Lin Y. W-box and G-box elements play important roles in early senescence of rice flag leaf. Sci Rep. 2016. https://doi.org/10.1038/srep20881.
Lin PC, Pomeranz MC, Jikumaru Y, Kang SG, Hah C, Fujioka S, Kamiya Y, Jang JC. The Arabidopsis tandem zinc finger protein AtTZF1 affects ABA- and GA-mediated growth, stress and gene expression responses. Plant J. 2011;65(2):253–68. https://doi.org/10.1111/j.1365-313X.2010.04419.x.
Khatun K, Robin AHK, Park J-I, Nath UK, Kim CK, Lim K-B, Nou IS, Chung M-Y. Molecular characterization and expression profiling of tomato GRF transcription factor family genes in response to abiotic stresses and phytohormones. Int J Mol Sci. 2017;18(5):1056. https://doi.org/10.3390/ijms18051056.
Maury S, Sow MD, Le Gac AL, Genitoni J, Lafon-Placette C, Mozgova I. Phytohormone and chromatin crosstalk: the missing link for developmental plasticity? Front Plant Sci. 2019;10:395. https://doi.org/10.3389/fpls.2019.00395.
Seo KI, Song E, Chung S, Lee JH. Roles of various cullin-RING E3 ligases involved in hormonal and stress responses in plants. J Plant Biol. 2012;55:421–8. https://doi.org/10.1007/s12374-012-0902-4.
Song Y, You J, Xiong L. Characterization of OsIAA1 gene, a member of rice Aux/IAA family involved in auxin and brassinosteroid hormone responses and plant morphogenesis. Plant Mol Biol. 2009;70:297–309. https://doi.org/10.1007/s11103-009-9474-1.
Potuschak T, Lechner E, Parmentier Y, Yanagisawa S, Grava S, Koncz C, Genschik P. EIN3-dependent regulation of plant ethylene hormone signaling by two arabidopsis F box proteins: EBF1 and EBF2. Cell. 2003;115(6):679–89. https://doi.org/10.1016/S0092-8674(03)00968-1.
Cui X, Guo Z, Song L, Wang Y, Cheng Y. NCP1/AtMOB1A plays key roles in auxin-mediated arabidopsis development. PLoS Genet. 2016;12(3): e1005923. https://doi.org/10.1371/journal.pgen.1005923.
Roosjen M, Paque S, Weijers D. Auxin response factors: output control in auxin biology. J Exp Bot. 2018;69(2):179–88. https://doi.org/10.1093/jxb/erx237.
Reyes JC. The many faces of plant SWI/SNF complex. Mol Plant. 2014;7(3):454–8. https://doi.org/10.1093/mp/sst147.
Sirko A, Wawrzyńska A, Brzywczy J, Sieńko M. Control of ABA signaling and crosstalk with other hormones by the selective degradation of pathway components. Int J Mol Sci. 2021;22(9):4638. https://doi.org/10.3390/ijms22094638.
Mindrebo JT, Nartey CM, Seto Y, Burkart MD, Noel JP. Unveiling the functional diversity of the alpha/beta hydrolase superfamily in the plant kingdom. Curr Opin Struct Biol. 2016;41:233–46. https://doi.org/10.1016/j.sbi.2016.08.005.
Xie M, Zhang J, Yao T, Bryan AC, Pu Y, Labbé J, Pelletier DA, Engle N, Morrell-Falvey JL, Schmutz J, Ragauskas AJ, Tschaplinski TJ, Chen F, Tuskan GA, Muchero W, Chen JG. Arabidopsis C-terminal binding protein ANGUSTIFOLIA modulates transcriptional co-regulation of MYB46 and WRKY33. New Phytol. 2020;228(5):1627–39. https://doi.org/10.1111/nph.16826.
Ortigosa A, Fonseca S, Franco-Zorrilla JM, et al. The JA-pathway MYC transcription factors regulate photomorphogenic responses by targeting HY5 gene expression. Plant J. 2020;102(1):138–52. https://doi.org/10.1111/tpj.14618.
Li C, Ng CK-Y, Fan L-M. MYB transcription factors, active players in abiotic stress signaling. Environ Exp Bot. 2015;114:80–91. https://doi.org/10.1016/j.envexpbot.2014.06.014.
Wang RK, Cao ZH, Hao YJ. Overexpression of a R2R3 MYB gene MdSIMYB1 increases tolerance to multiple stresses in transgenic tobacco and apples. Physiol Plant. 2014;150:76–87. https://doi.org/10.1111/ppl.12069.
Han Z, Shang X, Shao L, Wang Y, Zhu X, Fang W, Ma Y. Meta-analysis of the effect of expression of MYB transcription factor genes on abiotic stress. PeerJ. 2021;9:11268. https://doi.org/10.7717/peerj.11268.
Mu RL, Cao YR, Liu YF, et al. An R2R3-type transcription factor gene AtMYB59 regulates root growth and cell cycle progression in Arabidopsis. Cell Res. 2009;19(11):1291–304. https://doi.org/10.1038/cr.2009.83.
Shin R, Burch AY, Huppert KA, et al. The Arabidopsis transcription factor MYB77 modulates auxin signal transduction. Plant Cell. 2007;19(8):2440–53. https://doi.org/10.1105/tpc.107.050963.
Seo PJ, Park C-M. Auxin homeostasis during lateral root development under drought condition. Plant Signal Behav. 2009;4(10):1002–4. https://doi.org/10.4161/psb.4.10.9716.
Liu J, Osbourn A, Ma P. MYB transcription factors as regulators of phenylpropanoid metabolism in plants. Mol Plant. 2015;8(5):689–708. https://doi.org/10.1016/j.molp.2015.03.012.
Schiavon M, Pizzeghello D, Muscolo A, Vaccaro S, Francioso O, Nardi S. High molecular size humic substances enhance phenylpropanoid metabolism in maize (Zea mays L.). J Chem Ecol. 2010;36(6):662–9. https://doi.org/10.1007/s10886-010-9790-6.
Rushton PJ, Somssich IE, Ringler P, Shen QJ. WRKY transcription factors. Trends Plant Sci. 2010;15(5):247–58. https://doi.org/10.1016/j.tplants.2010.02.006.
Yang X, Yang YN, Xue LJ, et al. Rice ABI5-Like1 regulates abscisic acid and auxin responses by affecting the expression of ABRE-containing genes. Plant Physiol. 2011;156(3):1397–409. https://doi.org/10.1104/pp.111.173427.
Tripathi P, Rabara RC, Rushton PJ. A systems biology perspective on the role of WRKY transcription factors in drought responses in plants. Planta. 2014;239(2):255–66. https://doi.org/10.1007/s00425-013-1985-y.
Kim YS, Kim SG, Park JE, et al. A membrane-bound NAC transcription factor regulates cell division in Arabidopsis. Plant Cell. 2006;18(11):3132–44. https://doi.org/10.1105/tpc.106.043018.
Park J, Kim YS, Kim SG, Jung JH, Woo JC, Park CM. Integration of auxin and salt signals by the NAC transcription factor NTM2 during seed germination in Arabidopsis. Plant Physiol. 2011;156(2):537–49. https://doi.org/10.1104/pp.111.177071.
Xie Q, Frugis G, Colgan D, Chua NH. Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development. Genes Dev. 2000;14(23):3024–36. https://doi.org/10.1101/gad.852200.
Mao C, He J, Liu L, et al. OsNAC2 integrates auxin and cytokinin pathways to modulate rice root development. Plant Biotechnol J. 2020;18(2):429–42. https://doi.org/10.1111/pbi.13209.
Nuruzzaman M, Sharoni AM, Kikuchi S. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants. Front Microbiol. 2013. https://doi.org/10.3389/fmicb.2013.00248.
Kim SG, Lee AK, Yoon HK, Park CM. A membrane-bound NAC transcription factor NTL8 regulates gibberellic acid-mediated salt signaling in Arabidopsis seed germination. Plant J. 2008;55(1):77–88. https://doi.org/10.1111/j.1365-313X.2008.03493.x.
Srivastava AK, Pasala R, Minhas PS, Suprasanna P. Plant bioregulators for sustainable agriculture: integrating redox signaling as a possible unifying mechanism. Adv Agron. 2016. https://doi.org/10.1016/bs.agron.2015.12.002.
Lamar RT. Possible role for electron shuttling capacity in elicitation of pb activity of humic substances on plant growth enhancement. In: Geelen D, Xu L, Stevens CV, editors. The chemical biology of plant biostimulants. Hoboken: Wiley Press; 2020. p. 97–121. https://doi.org/10.1002/9781119357254.ch4.
Monda H, McKenna AM, Fountain R, Lamar RT. Bioactivity of humic acids extracted from shale ore: molecular characterization and structure-activity relationship with tomato plant yield under nutritional stress. Front Plant Sci. 2021;12: 660224. https://doi.org/10.3389/fpls.2021.660224.
Acknowledgements
Not applicable.
Funding
This work was supported by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) and Conselho Nacional de Desenvolvimento de Pesquisa e Tecnologia (CNPq).
Author information
Authors and Affiliations
Contributions
LPC, FLO, AP, and LEPP conceived the experiments. ACS carried out the experiments. FLO supervised the RNAseq experiment. LPC wrote the first version of the manuscript. All the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This manuscript is an original paper and has not been published in other journals. The authors agreed to keep the copyright rule.
Consent for publication
The authors agreed to the publication of the manuscript in this journal.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Souza, A.C., Olivares, F.L., Peres, L.E.P. et al. Plant hormone crosstalk mediated by humic acids. Chem. Biol. Technol. Agric. 9, 29 (2022). https://doi.org/10.1186/s40538-022-00295-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40538-022-00295-2