
Abstract
Regulatory T cells are crucial in controlling various functions of effector T cells during experimental autoimmune encephalomyelitis. While regulatory T cells are reported to exert their immunomodulatory effects in the peripheral immune organs, their role within the central nervous system (CNS) during experimental autoimmune encephalomyelitis is unclear. Here, by combining a selectively timed regulatory T cells depletion with 2-photon microscopy, we report that regulatory T cells exercise their dynamic control over effector T cells in the CNS. Acute depletion of regulatory T cells exacerbated experimental autoimmune encephalomyelitis severity which was accompanied by increased pro-inflammatory cytokine production and proliferation of effector T cells. Intravital microscopy revealed that, in the absence of regulatory T cells, the velocity of effector T cells was decreased with simultaneous increase in the proportion of stationary phase cells in the CNS. Based on these data, we conclude that regulatory T cells mediate recovery from experimental autoimmune encephalomyelitis by controlling cytokine production, proliferation and motility of effector T cells in the CNS.
Similar content being viewed by others
Introduction
CD4+Foxp3+ regulatory T cells (Treg) have a well-characterized role in promoting peripheral immunological tolerance throughout life by suppressing deleterious inflammatory responses [1]. Lack of Treg due to mutations in the FOXP3 gene in humans results in aggressive multi-organ autoimmunity called IPEX (immunodysregulation, polyendocrinopathy, enteropathy, X-linked) syndrome [2]. Similarly, scurfy mice, which harbor mutations in the Foxp3 gene, or Foxp3-gene deficient mice suffer from a massive lymphoproliferative syndrome [3],[4]. Targeted depletion of Treg also resulted in severe multi-organ autoimmunity [5],[6]. Intriguingly, however, no spontaneous central nervous system (CNS) inflammation was observed in Foxp3 mutant mice or after targeted depletion of Foxp3+Treg cells in wild type mice [7].
Treg have been demonstrated to be capable of controlling CNS autoimmunity in several Experimental Autoimmune Encephalomyelitis (EAE) models. The frequencies of Treg population within the CNS were elevated during the recovery phase of actively induced EAE [8]-[10]. Moreover, several studies described that the transfer of CD25+ Treg ameliorated EAE symptoms [9],[11]-[13]. In addition, non-specific ablation of natural Treg by anti-CD25 antibodies has been reported to exacerbate EAE [9],[14]-[16]. Furthermore, Treg have been shown to prevent spontaneous EAE development [17],[18] or delay spontaneous EAE onset [19].
Where and how do Treg exert their control over myelin-specific T cells? In principle, Treg could suppress effector T cells (Teff) in the periphery or within the target organ, CNS. One report demonstrated that Treg accumulate in the CNS at the peak of EAE but were unable to suppress CNS-derived Teff in vitro[8]. In contrast, Treg isolated from the recovery phase of the disease were still capable of suppressing Teff[10]. Furthermore, another study reported that, in the absence of Treg, there is an enhanced migration of Teff from the periphery [19]. Treg are known to limit the inflammatory reactions using several mechanisms that include soluble mediators, cell-to-cell contact with Teff or inhibiting antigen presenting cells (APCs) [1]. Treg influence EAE by affecting the priming and polarization of Teff[11],[20]. Among soluble cytokines produced by Treg, IL-10 is important in containing Teff proliferation in vitro[16]. Treg can also set a threshold for activation of autoreactive Teff by inhibiting their contacts with antigen-loaded dendritic cells (DCs) in the lymph nodes [21]-[23]. Furthermore, Treg have been shown to contact and inhibit DCs in vitro via CTLA-4 [24]. However, the mode of action of Treg during CNS autoimmunity, in particular within the target organ, still remains unclear.
To address those principal outstanding issues, in the present study, we combine targeted and acute depletion of Treg with intravital two-photon microscopy to investigate the functional role of Treg in the CNS during EAE. We found that Treg limit autoimmune inflammation by controlling the Teff proliferation and motility within the CNS.
Material and methods
Animals
DEREG [6] and T-Red [25] mice with the C57BL/6 genetic background were used. All mice were bred in the animal facility of the Max Planck Institute of Neurobiology and all experiments were conducted according to the guidelines of the committee on animals of the Max Planck Institute of Neurobiology and were approved by the Regierung von Oberbayern.
EAE induction and diphtheria toxin treatment
EAE was induced by injecting the mice subcutaneously into the flanks with 200 μl of emulsion containing 200 μg MOG35–55 peptide (MEVGWYRSPFSRVVHLYRNGK) and 500 μg M. tuberculosis strain H37 Ra (Difco) in incomplete Freund Adjuvant oil (Difco). In addition, the mice received 400 ng pertussis toxin (List Biological Laboratories) intraperitoneally (i.p.) on days 0 and 2 after immunization. Clinical signs of EAE were assessed daily according to the standard 5 point scale [26]. For depletion of Treg in DEREG mice, diphtheria toxin (Sigma-Aldrich) was injected both i.p. and i.v. (200 ng respectively) on day 4 post EAE onset.
Cell isolation and flow cytometry
Cells from lymph nodes and spinal cord were isolated as described before [26]. For detection of cell surface markers, cells were stained in FACS buffer (PBS containing 1% BSA and 0.1% NaN3) with the following fluorochrome labeled monoclonal antibodies: anti-CD45 (30-F11), anti-CD4 (RM4-5), anti-CD25 (PC61) and anti-CD44 (IM7). For intracellular cytokine staining, cells were incubated for 16 hours with anti-CD3 (0.5 μg/ml). Next, cells were fixed and permeabilized by incubation with Foxp3 Fixation/Permeabilization Buffer (eBioscience) and stained in Permeabilization Buffer (eBioscience) with the following fluorochrome labeled monoclonal antibodies: anti-Foxp3 (FJK-16s), anti-IL-17 (eBio17B7) and anti-IFNγ (XMG1.2). All antibodies were purchased from BD Pharmigen or eBioscience. For cell number quantification, 104 FACSuite FC Beads (BD) were added per sample prior to acquisition. Samples were acquired on FACS Verse (BD). FACS data were analyzed using FlowJo 7.6.5 software (TreeStar).
EdU proliferation assay
For in vivo proliferation experiments, 400 μg EdU (Life Technologies) were injected i.p. to mice ~16 hours before their sacrification. The Click-iT® EdU Alexa Fluor® 647 Flow Cytometry Assay Kit (Life Technologies) was used for staining for flow cytometry according to manufacturer’s instructions.
Immunofluorescence
The organ sections were prepared as described previously [27]. The following monoclonal antibodies were used for staining: biotin-anti-CD4 (RM4-5; BD), Alexa Fluor 647-anti-CD11b (M1/70; Biolegend), Alexa Fluor 488-anti-Foxp3 (FJK-16 s; eBioscience), and Alexa Fluor 568-streptavidin (Invitrogen). Images were acquired on a SP5 confocal microscope (Leica), using 20x air-immersion (N.A. 0.70) or 63x oil-immersion (N.A. 1.4) objective. Images were processed using Image J (NIH) and Photoshop CS5 software (Adobe Systems).
In vivo IL-2 blocking
MOG35–55/CFA-immunized DEREG B6 mice were treated with DTx, as described above. Purified anti-IL2 (JES6-1A12) monoclonal antibody or isotype control antibody (J1.2) was injected i.v. on day 4 (400 μg) and day 6 (200 μg) post EAE onset.
Intravital imaging
The technical setup of the 2-photon microscopy was as described before [28]. The pulsed laser was tuned to 880 nm and routed through a 25× water immersion objective (N.A. 0.95, Leica). Typically, a field of 360 × 360 μm was scanned, and 40–80 μm z-stacks were acquired using a 3–6 μm z-step. The acquisition rate was set to 25.219 s intervals, with images line-averaged twice. The fluorescence signals were detected using non-descanned photomultiplier tube (PMT) detectors (Hamamatsu) equipped with 525/50 nm (for detection of Alexa Fluor 488) and 630/69 nm (for detection of dsRedII) band-pass filters (Semrock). Mice were anesthetized and imaging in the spinal cord was performed as described previously [28]. For labeling of perivascular meningeal APC, we performed local instillation of Alexa Fluor 488–conjugated dextran (10 ng/μl, 10 kDa; Life Technologies) 20 min prior to imaging, as described before [29]. Image analysis was performed as described previously [29].
Statistical analysis
Statistical evaluations were performed as indicated in figure legends using GraphPad Prism software.
Results and discussion
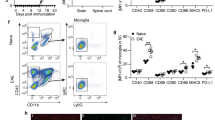
Our approach to analyze the role of Treg in vivo during EAE differs from previous attempts which have used anti-CD25 antibodies [9],[14]-[16]. Since these antibodies persist in the circulation for long time, their effect on CD25-expressing activated Teff cannot be excluded. To investigate the functional role of Treg in the CNS during peak EAE, we chose DEREG mice which express a diphtheria toxin (DTx) receptor-enhanced GFP fusion protein, under the control of the Foxp3 gene locus, permitting specifically timed depletion of Treg by treatment with DTx [6]. We immunized DEREG mice with MOG35–55 in CFA and treated them with DTx or PBS during peak of the disease. We monitored the efficiency of Treg depletion in peripheral immune organs and the CNS by flow cytometry. Staining for Foxp3 revealed that Treg population was almost completely absent in DTx-treated DEREG mice compared to control animals in all the organs tested (Figure 1A,B). Next, immunized and treated mice were monitored daily for clinical score to assess the effect of Treg depletion on EAE pathogenesis. While PBS-treated DEREG mice partially recovered from EAE, DTx-treated DEREG mice not only failed to recover from the disease, but also developed severe and, eventually, fatal EAE (Figure 1C). To rule out any adverse effect of DTx on EAE disease course, we have treated MOG35–55 immunized wild type mice with DTx during peak EAE. Unlike in DEREG mice, the EAE course was not affected compared to control mice (Additional file 1: Figure S1). These findings suggest that Treg contribute to recovery from EAE and perhaps have an essential function within the CNS.

Exacerbation of EAE after acute T reg depletion at the peak of the disease. Representative flow cytometry plots (A) or mean frequency (± SEM) of Foxp3+ in CD45+CD4+ T cells (B) isolated from indicated organs of DEREG mice two days after DTx or PBS treatment are shown (n = 11 mice per group, pooled data from four independent experiments). (C) Mean clinical score (± SEM) of mice following immunization and DTx or PBS treatment (day 4 post EAE onset). (n = 6-7 mice per group, representative data from two independent experiments. **P < 0.01, ****P < 0.0001, 2way ANOVA).
To learn how Treg ablation led to dramatic disease exacerbation, we stained spinal cord sections of the mice two days after the treatment with DTx or PBS. We observed highly increased numbers of Teff, as well as macrophages, in the spinal cord infiltrates in Treg-depleted mice compared to the control animals (Figure 2A-F). Next, we quantified the numbers of CD45+CD4+Foxp3− T cells (Teff) in the draining lymph nodes (LN), as well as in the spinal cord, using flow cytometry. Interestingly, lymph nodes and spinal cord of Treg-depleted mice contained significantly elevated numbers of Teff (defined as CD45+CD4+Foxp3− T cells) compared to control mice (Figure 2G). In addition, DTx-treated mice displayed higher frequencies of pro-inflammatory cytokine IFNγ-secreting Teff in the CNS and LN (Figure 2H). In contrast, we detected similar frequencies of IL-17+ Teff in both groups. We also did not observe differences in the percentage of activated CD25+CD44high Teff in DTx-treated mice (Additional file 1: Figure S2). Therefore, the exacerbated EAE pathology observed in Treg-depleted mice could be attributed to increased fraction of IFNγ-producing Teff in the CNS.

Increased numbers and enhanced proliferation of T eff in T reg -depleted mice. (A), (D) Representative panoramic pictures of spinal cord cryosections from mice treated with DTx (A) or PBS (D) stained with anti-Foxp3 (green), anti-CD4 (red) and anti-CD11b Ab (grey) (scale bar: 200 μm). (B), (C), (E), (F) Magnified pictures of the indicated regions (A) and (D) (scale bar: 50 μm). (G) Mean absolute numbers (± SEM) of Teff (CD45+CD4+Foxp3− cells) isolated from LN and spinal cord of DEREG mice two days after DTx or PBS treatment (n = 11 mice per group, pooled data from three independent experiments). (H) Mean frequency (± SEM) of IFNγ+ cells in Teff isolated from LN and spinal cord of DEREG mice two days after DTx or PBS treatment (n = 5 mice per group, representative data from three independent experiments) (I) EdU was injected i.p. one day after DTx or PBS treatment and 16 hours later lymphocytes were isolated and stained for EdU. Mean frequency (± SEM) of EdU+ cells within Teff (n = 13-14 mice per group, pooled data from four independent experiments). (*P < 0.05, **P < 0.01, t-test).
Treg are known to suppress the proliferation and activation of Teff cells through multiple mechanisms [1]. Lack of functional Treg results in a lymphoproliferative disease, as in scurfy mutant mice [30]. Similar fatal lymphoproliferative disease was observed after chronic depletion of Treg in adult and neonatal mice [5],[6]. To determine if the elevated numbers of Teff were a result of increased T cell proliferation, we assessed the in vivo proliferation of Teff in the presence or absence of Treg. One day after the DTx or PBS treatment of immunized DEREG mice, we injected EdU (5-ethynyl-2´-deoxyuridine), a thymidine analogue which is readily incorporated into cellular DNA during DNA replication, and examined the EdU incorporation in T cells by flow cytometry (Figure 2I). The fraction of EdU+ Teff was significantly higher in both LN and the spinal cord of DTx-treated mice compared to control mice, suggesting that the Teff proliferation during EAE is enhanced in the absence of Treg.
Since we observed an increased proliferation of Teff in the CNS, we focused on the role of IL-2, a pivotal cytokine for T cell proliferation. Treg express high levels of high affinity IL-2 receptor α (CD25), thereby restricting the availability of IL-2 by direct consumption to restrain the activation of proliferating T cells [31]. We hypothesized that the enhanced Teff proliferation that we observed after elimination of Treg could be attributed to increased availability of IL-2. To test this hypothesis, we quantified the IL-2 protein levels in LN and spinal cord tissue extracts from Treg-depleted and Treg-intact mice with EAE. However, both groups exhibited similar levels of IL-2 (Additional file 1: Figure S3A). Furthermore, administration of anti-IL-2 blocking antibodies in parallel with DTx treatment did not prevent EAE exacerbation (Additional file 1: Figure S3B). These findings suggest that IL-2 deprivation is not a major mechanism used by Treg to control Teff proliferation in vivo within the CNS during EAE.
We considered the possibility of direct or indirect interactions of Treg with Teff and APCs to mediate suppression of Teff in the CNS during EAE recovery. Previous 2-photon imaging studies in LN have shown that Treg can limit the contacts between Teff and DCs [21]-[23]. However, the effect of Treg on the migratory behavior of Teff within the CNS during EAE is not known. We sought to investigate how the ablation of Treg can affect the dynamic behavior of Teff in the CNS using intravital two-photon imaging. To this end, we crossed T-Red mice, in which T cells express the red fluorescent protein dsRedII [32], to DEREG mice. Subsequently, we treated MOG-immunized T-Red x DEREG mice with DTx or PBS at the peak of EAE and performed intravital two-photon imaging in the spinal cord meninges.
Cell tracking of the dsRedII-expressing Teff revealed that Teff displayed more confined trajectories in the CNS of DTx-treated mice than PBS-treated mice (Figure 3A,B). Indeed, analysis of the Teff tracks showed that the cells had significantly reduced mean velocity and linearity index compared to the control animals in the CNS (Figure 3C,D). In parallel, the stationary phase of Teff was increased (Figure 3E). Collectively, these findings suggest that the absence of Treg results in decreased motility of Teff, which indicates enhanced interactions with potential APCs within the inflamed spinal cord meninges.

T eff dynamics in the CNS of T reg -depleted mice. Intravital two-photon imaging was performed in the spinal cord meninges of T-Red x DEREG mice two days after DTx or PBS treatment. (A) Trajectories (white lines) of dsRedII+ Teff overlaid with snapshots from representative videos. One representative out of three independent experiments per treatment condition is shown (red: Teff, green: APC, scale bar: 30 μm) (B) Superimposed trajectories of Teff movements after DTx or PBS treatment. Time points with contacts with APC are indicated in red or blue, respectively. One representative out of three independent experiments per treatment condition is shown. (C) Average velocity, (D) linearity index (sum of the total displacement divided by the path length of a cell), (E) stationary phase (part of the track with velocity <3 μm/min) (± SEM) of Teff after DTx or PBS treatment (pooled data from three independent experiments (**P < 0.01, t-test) (pooled data from three independent experiments).
In summary, using 2-photon imaging, we showed that Treg exert dynamic control over Teff within the CNS during effector phase of EAE. This finding doesn’t exclude additional actions mediated by Treg in the periphery. Our results are in agreement with many reports which showed that ablation of Treg population (by treatment with anti-CD25 antibody) exacerbates EAE [9],[14]-[16]. However, a major disadvantage of this approach is that CD25 is not a Treg-specific marker, but is also expressed by activated Teff complicating the interpretation of these findings. Our approach using DEREG mice circumvents these issues by specifically timed depletion of Treg. This is also a first study in an active EAE which uses specific Treg depletion. Our results are compatible with a recent report which showed that selective Treg depletion resulted in an increased incidence and accelerated disease onset in a spontaneous EAE model [19].
While the importance of Treg during CNS autoimmunity is unequivocally shown, where and in which phase of the disease they are important is not clear. The main conclusion from our study is that the Treg exert their regulatory control over Teff within the CNS in addition to their known peripheral effects. At first glance, our results are in contrast to a report by Korn et al., which suggested that regulatory T cells accumulate in the CNS but are unable to control CNS infiltrating Teff during peak of the disease [8]. The conclusions were drawn based on the inability of CNS derived Treg to suppress Teff proliferation. We, however, have followed the behavior of Teff cells in their “native” environment. Moreover, several studies reported that natural recovery from EAE correlating with increasing Treg numbers suggests that Treg are essential to mediate recovery [8]-[10]. Concerning the potential mode of action, we observed that the exacerbation of EAE was preceded by an increase in the numbers of Teff due to local proliferation in the absence of Treg. Earlier reports using two-photon microscopy have demonstrated that CD4+CD25− T cells established longer contacts with DCs in lymph node in the absence of Treg (defined as CD4+CD25+ T cells), while in Treg-sufficient environment these contacts were inhibited [22],[23]. Moreover, Treg have been recently described to suppress the T cell movements in the LN during EAE in a PSGL-1-dependent mechanism [21]. Our results show that there is an increase in the motility of Teff in Treg-depleted mice and there was an increase in the stationary phase of Teff, indicating increased contacts with APCs in the CNS. In conclusion, our findings suggest that Treg are indispensable for recovery from EAE through their actions within and outside of the CNS.
Additional file
References
Josefowicz SZ, Lu LF, Rudensky AY: Regulatory T cells: Mechanisms of differentiation and function. Annu Rev Immunol 2012, 30: 531–564. doi:10.1146/annurev.immunol.25.022106.141623 10.1146/annurev.immunol.25.022106.141623
Bennett CL, Christie J, Ramsdell F, Brunkow ME, Ferguson PJ, Whitesell L, Kelly TE, Saulsbury FT, Chance PF, Ochs HD: The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nat Genet 2001, 27: 20–21. 10.1038/83713
Brunkow ME, Jeffery EW, Hjerrild KA, Paeper B, Clark LB, Yasayko SA, Wilkinson JE, Galas D, Ziegler SF, Ramsdell F: Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of the scurfy mouse. Nat Genet 2001, 27: 68–73. doi:10.1038/83784 10.1038/83784
Fontenot JD, Gavin MA, Rudensky AY: Foxp3 programs the development and function of CD4 + CD25+ regulatory T cells. Nat Immunol 2003, 4: 330–336. doi:10.1038/ni904 10.1038/ni904
Kim JM, Rasmussen JP, Rudensky AY: Regulatory T cells prevent catastrophic autoimmunity throughout the lifespan of mice. Nat Immunol 2007, 8: 277–284. doi:10.1038/ni1428 10.1038/ni1437
Lahl K, Loddenkemper C, Drouin C, Freyer J, Arnason J, Eberl G, Hamann A, Wagner H, Huehn J, Sparwasser T: Selective depletion of Foxp3+ regulatory T cells induces a scurfy-like disease. J Exp Med 2007, 204: 57–63. 10.1084/jem.20061852
Krishnamoorthy G, Holz A, Wekerle H: Experimental models of spontaneous autoimmune disease in the central nervous system. J Mol Med 2007, 85: 1161–1173. doi:10.1007/s00109–007–0218-x 10.1007/s00109-007-0218-x
Korn T, Reddy J, Gao WD, Bettelli E, Awasthi A, Petersen TR, Bäckström BT, Sobel RA, Wucherpfennig KW, Strom TB, Oukka M, Kuchroo VK: Myelin-specific regulatory T cells accumulate in the CNS but fail to control autoimmune inflammation. Nat Med 2007, 13: 423–431. 10.1038/nm1564
McGeachy MJ, Stephens LA, Anderton SM: Natural recovery and protection from autoimmune encephalomyelitis: Contribution of CD4 + CD25 + regulatory cells within the central nervous system. J Immunol 2005, 175: 3025–3032. 10.4049/jimmunol.175.5.3025
O'Connor RA, Malpass KH, Anderton SM: The inflamed central nervous system drives the activation and rapid proliferation of Foxp3 + regulatory T cells. J Immunol 2007, 179: 958–966. 10.4049/jimmunol.179.2.958
Kohm AP, Carpentier PA, Anger HA, Miller SD: Cutting edge: CD4 + CD25+ regulatory T cells suppress antigen-specific autoreactive immune responses and central nervous system inflammation during active experimental autoimmune encephalomyelitis. J Immunol 2002, 169: 4712–4716. 10.4049/jimmunol.169.9.4712
Mekala DJ, Alli RS, Geiger TL: IL-10-dependent infectious tolerance after the treatment of experimental allergic encephalomyelitis with redirected CD4 + CD25 + T lymphocytes. Proc Natl Acad Sci U S A 2005, 102: 11817–11822. 10.1073/pnas.0505445102
Stephens LA, Malpass KH, Anderton SM: Curing CNS autoimmune disease with myelin-reactive Foxp3(+) Treg. Eur J Immunol 2009, 39: 1108–1117. 10.1002/eji.200839073
Gärtner D, Hoff H, Gimsa U, Burmester GR, Brunner-Weinzierl MC: CD25 regulatory T cells determine secondary but not primary remission in EAE: Impact on long-term disease progression. J Neuroimmunol 2006, 172: 73–84. 10.1016/j.jneuroim.2005.11.003
Montero E, Nussbaum G, Kaye JF, Perez R, Lage A, Ben-Nun A, Cohen IR: Regulation of experimental autoimmune encephalomyelitis by CD4 +, CD25 + and CD8 + T cells: analysis using depleting antibodies. J Autoimmun 2004, 23: 1–7. 10.1016/j.jaut.2004.05.001
Zhang X, Koldzic DN, Izikson L, Reddy J, Nazareno RF, Sakaguchi S, Kuchroo VK, Weiner HL: IL-10 is involved in the suppression of experimental autoimmune encephalomyelitis by CD25 + CD4+ regulatory T cells. Int Immunol 2004, 16: 249–256. 10.1093/intimm/dxh029
Hori S, Haury M, Coutinho A, Demengeot J: Specificity requirements for selection and effector functions of CD25 + 4 + regulatory T cells in anti-myelin basic protein T cell receptor transgenic mice. Proc Natl Acad Sci U S A 2002, 99: 8213–8218. 10.1073/pnas.122224799
Olivares-Villagómez D, Wang Y, Lafaille JJ: Regulatory CD4 + T cells expressing endogenous T cell receptor chains protect myelin basic protein-specific transgenic mice from spontaneous autoimmune encephalomyelitis. J Exp Med 1998, 188: 1883–1894. 10.1084/jem.188.10.1883
Lowther DE, Chong DL, Ascough S, Ettorre A, Ingram RJ, Boyton RJ, Altmann DM: Th1 not Th17 cells drive spontaneous MS-like disease despite a functional regulatory T cell response. Acta Neuropathol 2013, 126: 501–515. doi:10.1007/s00401–013–1159–9 10.1007/s00401-013-1159-9
Reddy J, Waldner H, Zhang XM, Illés Z, Wucherpfennig KW, Sobel RA, Kuchroo VK: CD4+CD25+ regulatory T cells contribute to gender differences in susceptibility to experimental autoimmune encephalomyelitis. J Immunol 2005, 175: 5591–5595. 10.4049/jimmunol.175.9.5591
Angiari S, Rossi B, Piccio L, Zinselmeyer BH, Budui S, Zenaro E, Della Bianca V, Bach SD, Scarpini E, Bolomini-Vittori M, Piacentino G, Dusi S, Laudanna C, Cross AH, Miller MJ, Constantin G: Regulatory T cells suppress the late phase of the immune response in lymph nodes through P-selectin glycoprotein ligand-1. J Immunol 2013, 191: 5489–5500. doi:10.4049/jimmunol.1301235 10.4049/jimmunol.1301235
Tadokoro CE, Shakhar G, Shen SQ, Ding Y, Lino AC, Maraver A, Lafaille JJ, Dustin ML: Regulatory T cells inhibit stable contacts between CD4+ T cells and dendritic cells in vivo. J Exp Med 2006, 203: 505–511. 10.1084/jem.20050783
Tang QZ, Adams JY, Tooley AJ, Bi MY, Fife BT, Serra P, Santamaria P, Locksley RM, Krummel MF, Bluestone JA: Visualizing regulatory T cell control of autoimmune responses in nonobese diabetic mice. Nat Immunol 2006, 7: 83–92. 10.1038/ni1289
Onishi Y, Fehérvári Z, Yamaguchi T, Sakaguchi AY: Foxp3 + natural regulatory T cells preferentially form aggregates on dendritic cells in vitro and actively inhibit their maturation. Proc Natl Acad Sci U S A 2008, 105: 10113–10118. 10.1073/pnas.0711106105
Mempel TR, Pittet MJ, Khazaie K, Weninger W, Weissleder R, von Boehmer H, von Andrian UH: Regulatory T Cells Reversibly Suppress Cytotoxic T Cell Function Independent of Effector Differentiation. Immunity 2006, 25: 129–141. http://dx.doi.org/10.1016/j.immuni.2006.04.015 10.1016/j.immuni.2006.04.015
Domingues HS, Mues M, Lassmann H, Wekerle H, Krishnamoorthy G (2010) Functional and pathogenic differences of Th1 and Th17 cells in experimental autoimmune encephalomyelitis. PLoS One 5:e15531. doi:10.1371/journal.pone.0015531
Berer K, Mues M, Koutroulos M, Al Rasbi Z, Boziki M, Johner C, Wekerle H, Krishnamoorthy G: Commensal microbiota and myelin autoantigen cooperate to trigger autoimmune demyelination. Nature 2011, 479: 538–541. doi:10.1038/nature10554 10.1038/nature10554
Mues M, Bartholomaus I, Thestrup T, Griesbeck O, Wekerle H, Kawakami N, Krishnamoorthy G: Real-time in vivo analysis of T cell activation in the central nervous system using a genetically encoded calcium indicator. Nat Med 2013, 19: 778–783. doi:10.1038/nm.3180 10.1038/nm.3180
Pesic M, Bartholomaus I, Kyratsous NI, Heissmeyer V, Wekerle H, Kawakami N: 2-photon imaging of phagocyte-mediated T cell activation in the CNS. J Clin Invest 2013, 123: 1192–1201. doi:10.1172/JCI67233 10.1172/JCI67233
Godfrey VL, Wilkinson JE, Rinchik EM, Russell LB: Fatal lymphoreticular disease in the scurfy ( sf ) mouse requires T cells that mature in the sf thymic environment: Potential model for thymic education. Proc Natl Acad Sci U S A 1991, 88: 5528–5532. 10.1073/pnas.88.13.5528
Pandiyan P, Zheng L, Ishihara S, Reed J, Lenardo MJ: CD4 + CD25 + Foxp3+ regulatory T cells induce cytokine deprivation-mediated apoptosis of effector CD4+ T cells. Nat Immunol 2007, 8: 1353–1362. doi:10.1038/ni1536 10.1038/ni1536
Pittet MJ, Mempel TR: Regulation of T-cell migration and effector functions: insights from in vivo imaging studies. Immunol Rev 2008, 221: 107–129. 10.1111/j.1600-065X.2008.00584.x
Acknowledgments
We thank Irene Arnold-Ammer, Sabine Kosin and Birgit Kunkel for technical support. We thank Ingo Bartholomäus for providing analysis tools for imaging data. We thank Tim Sparwasser for providing DEREG mice. This work was funded by the Hertie foundation, SFB/CRC 128 (Deutsche Forschungsgemeinschaft), KKNMS (Klinische Kompetenznetz Multiple Sklerose; BMBF) and the Max Planck Society. H.W. is an incumbent of a Hertie senior professorship. N.K. is supported by the Novartis Foundation for Therapeutic Research and LMU Munich.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Electronic supplementary material
40478_2014_163_MOESM1_ESM.pdf
Additional file 1: Figure S1.: DTx injection does not affect EAE course in non-transgenic mice. Mean clinical score (± SEM) of mice following immunization and DTx or PBS treatment (day 4 post EAE onset). (n = 5-6 mice per group, representative data from two independent experiments. Figure S2. Expression of IL-17 and T cell activation markers in Treg-depleted mice. (A) Mean frequency (± SEM) of IL-17+ cells in Teff isolated from LN and spinal cord of DEREG mice two days after DTx or PBS treatment (n = 5 mice per group, representative data from three independent experiments) (B) Mean frequency (± SEM) of CD25+CD44high cells within Teff isolated from LN and spinal cord of DEREG mice two days after DTx or PBS treatment (n = 8 mice per group, pooled data from two independent experiments). Figure S3. Treg do not use IL-2 deprivation to limit Teff proliferation. (A) IL-2 protein levels (± SEM) in LN and spinal cord of DEREG mice two days after DTx or PBS treatment (n = 5 mice per group, pooled data from two independent experiments). (B) Mean clinical score (± SEM) of mice following immunization and DTx treatment (day 4 post EAE onset) as well as anti-IL2 antibody treatment on days 4 and 6 post EAE onset (see arrow) (n = 3-4 mice per group, representative data from two independent experiments). (PDF 32 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

{kind=link}
{kind=link}
{kind=link}
Cite this article
Koutrolos, M., Berer, K., Kawakami, N. et al. Treg cells mediate recovery from EAE by controlling effector T cell proliferation and motility in the CNS. acta neuropathol commun 2, 163 (2014). https://doi.org/10.1186/s40478-014-0163-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40478-014-0163-1