
Abstract
Exosomes, ranging in size from 30 to 150 nm as identified initially via electron microscopy in 1946, are one of the extracellular vesicles (EVs) produced by many cells and have been the subject of many studies; initially, they were considered as cell wastes with the belief that cells produced exosomes to maintain homeostasis. Nowadays, it has been found that EVs secreted by different cells play a vital role in cellular communication and are usually secreted in both physiological and pathological conditions. Due to the presence of different markers and ligands on the surface of exosomes, they have paracrine, endocrine and autocrine effects in some cases. Immune cells, like other cells, can secrete exosomes that interact with surrounding cells via these vesicles. Immune system cells-derived exosomes (IEXs) induce different responses, such as increasing and decreasing the transcription of various genes and regulating cytokine production. This review deliberate the function of innate and acquired immune cells derived exosomes, their role in the pathogenesis of immune diseases, and their therapeutic appliances.
Similar content being viewed by others

Introduction
Exosomes are produced from various cells like the immune cells and secreted into the extracellular environment. They are generally present in mammalian peripheral blood, urine, human saliva, semen, breast milk, bronchoalveolar fluid, cerebrospinal fluid, and amniotic fluid [1, 2]. Exosomes produced from various cells have similarities and differences in size, composition, function, and surface markers [3].
Exosomes can increase the level of biomimetism by simulating what is happening in nature between cells. Although extracellular vesicles (EVs) are released from different cells that include both microvesicles (MVs) and exosomes, only exosomes have the optimal size that may be considered suitable for potential therapeutic applications [4]. It is proposed that exosomes released by normal cells always positively affect their target cells. In contrast, those released by cells under pathological conditions, such as tumors and hyperactivated immune cells, may exert dangerous, adverse, and unknown effects [4].
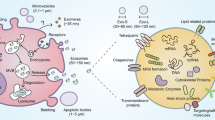
IEXs have a broad spectrum of functions in the immune system, including expression of several types of genes, modulation of the immune system, antigen presentation, antitumor immunity, and suppressing the immune system, besides playing a critical role in the pathogenesis of certain immune-related diseases such as asthma [5] and cardiovascular disease [6]. Exosomes can be isolated using ultracentrifugation, differential centrifugation, filtration, chromatography, immunoaffinity, and deploying kits [7]. It has been shown that the dendritic cell-derived exosomes (DEX) can enhance antitumor immunity and activate specific T cells to combat against tumor cells [8]. Using methods to upregulate and downregulate exosome production by immune cells is a new way to regulate immunity against tumor and infected cells and immune reaction in some autoimmune and allergic diseases [9, 10]. Also, tumor cells secrete exosomes into their microenvironment and suppress the immune responses in this region [11]. Furthermore, inhibition of tumor cells exosome production may reduce tumor immune suppression as well as their other functions, such as angiogenesis [12]. The analysis of exosome contents leads to their characterization and determines biological applications [13]; the content of exosomes can vary depending on the origin’s cell condition, such as the stage of maturation and cell type. Exosome content is composed of three groups of macromolecules, comprising proteins, lipids, and nucleic acids [14, 15]. Exosomes’ internal compounds are divided into two groups, the first group being common for all exosomes [16]. In contrast, the second group comprises specific substances that characterize the properties and functions of exosomes [17]. Biogenesis of exosomes is initiated by inward budding of the late endosomal membrane to form multivesicular bodies (MVBs). Intraluminal vesicles (ILVs) are formed during this process in the lumen of late endosomes [18]. As shown in Fig. 1, exosome formation in different cells occurs through two pathways, including endosomal sorting complex required for transport (ESCRT) dependent and independent pathways [19, 20]. After the formation of exosomes in the late endosomes and their conversion to MVBs, they move toward the cell membrane and, after integration, exosomes release out of the cell. The cargos carried by the exosomes are transmitted to the target cell in two ways [21].

Exosome’s contents, its formation, and its capture by the target cell are shown. Interestingly, although many exosomes do not enter the phagocytic pathway and degradation, some of them are release to the cytoplasm and are destroyed by lysosomal degradation. As shown in the figure, the exosomes bind and integrate by target cell in different ways
In some cases, exosome binding is sufficient to exert modifications on the target cell, but in other cases, the exosome must be internalized in addition to the binding. The initiation of exosome interaction with the target cell requires the interaction of two high-affinity surface molecules [22]. This interaction is mediated by a wide range of cell surface receptors such as integrins, cadherins, and tetraspanin [23]. Cargo transfer by exosomes to target cells modifies their functions and is performed in various ways, including soluble signaling, juxtacrine signaling, fusion, phagocytosis, macropinocytosis, receptor-mediated endocytosis, and lipid-mediated endocytosis [23, 24]. Figure 1 summarizes the biogenesis, composition, and different methods of exosomes uptake by the target cell (Fig. 1).
Immune cells-derived exosomes (IEXs)
Immune cells, like other cells, produce exosomes to reflect their properties and create an optimal microenvironment for immune cell function where they perform their function in paracrine and autocrine forms. IEXs can activate other immune cells, inhibit immune responses, and participate in the licensing phenomenon of antigen-presenting cells (APCs) [25]. To better understand, the characteristics of the exosomes of each cell involved in acquired and innate immunity are discussed separately below.
Dendritic cells-derived exosomes (DEXs)
DCs produce exosomes with diverse properties based on the activation nature and the presence of cytokines in the environment. If DCs are activated in inflammatory conditions, they produce exosomes that activate the innate immune system through stimulating the expression of NKG2D ligands (present on the surface of NK cells) such as MIC-A, MIC-B (MHC class I polypeptide-related sequence A and B), and ULBP-1 (UL16 binding protein-1) [26]. Also, these exosomes can activate specific T cells to detect antigens in both direct and indirect pathways [27]. In the direct pathway, DEXs can express the MHCII-peptide complexes and costimulatory molecules and bind to T cell receptors. In the indirect pathway, DEXs can deliver the MHC II-peptide complex to other DCs, a process named MHC-dressing. Besides, exosomes are able to transfer antigenic peptides from activated to inactivated DCs, which increases the number of MHCII-peptide complexes on the surface of DCs to activate the T cells. The results of new studies show that exosomes can form inside microbial compounds containing phagosomes (different types of antigens) and then be released into the immune microenvironment and participate in the antigen presentation [28]. It has also been shown that DCs-derived EVs can affect T cell responses by transferring miR-155 [29].
When DCs activated in a tolerogenic environment, they produce IL-10 and express Fas ligand (FAS-L) on their surface. Exosomes produced from these cells suppress autoimmune responses and the activity of both the innate and adaptive immune cells [30]. The expression of some proteins in the activated and inactivated DEXs are the same, such as clathrin, HSC70, annexin-2, and CD9. However, the expression levels of some proteins in the activated and activated DC-derived exosomes may differ. In the activated DC-derived exosomes, the expression levels of some molecules like MHCII, costimulatory molecules such as CD80, CD86, and CD40, as well as adhesion molecules, namely ICAM-1 increases but the expression of some molecules like MFG-E8 (Milk fat globule-epidermal growth factor-8) and MHC-I decrease [31]. Due to the unique characteristics of these exosomes, they have been investigated in various studies such as cancer immunotherapy, recruitment of mesenchymal stem cells (MSCs) [32], osteogenic differentiation of MSCs [33, 34], vaccine production for autoimmune diseases, and modulating the immune responses [35].
Macrophages-derived exosomes
Analysis based on mass spectrophotometry has revealed the presence of ~ 5100 proteins inside the macrophages-derived exosomes, and their protein content changes after activation. For example, in the lipopolysaccharide (LPS) activated macrophages, the NOD-like receptor (NLR) related proteins will increase. Activated macrophage exosomes induce inflammation in the target cell by activating the NLRP3 receptor-dependent inflammasome, TOLL-like receptors (TLR) and TNF-related signaling pathways. Also, the levels of some cytokines and chemokines such as CXCL2, IL-17, TNF-α, CCL3, and CXCL10, are significantly increased [36] and the levels of some molecules, such as mir-17, which regulate ICAM-1 negatively, decreases in exosomes produced from activated macrophages [37]. Macrophage-derived exosomes due to the presence of mir-155 reduced the proliferation and increased the inflammation in cardiac fibroblast cells. Mir-155 has two binding sites on the 3’UTR region of the SOS1 gene. The product of this gene, a major mediator of Ras activation, is involved in proliferation by activating MAPKs (mitogen-activated protein kinases) [38], which inhibits upon binding to mir-155, and reduces cell proliferation. Likewise, mir-155 binds to the SOCS1 gene, which inhibits the secretion of inflammatory cytokines, impedes their function, and enhances the production of anti-inflammatory cytokines in cardiac fibroblast cells [39]. Macrophage-derived exosomes that activated in the inflammatory conditions can induce inflammatory conditions in liver cells [36], placenta, and vascular endothelial cells [37]. Similarly, macrophage-derived exosomes can inhibit endothelial and tumour cells’ migration by inhibiting the expression of β1 integrins [40].
Exosomes produced from nonclassical macrophages (M2) regulate tumor cells’ migration, proliferation, invasion, and angiogenesis as they contain high levels of miR-21-5p and miR-155-5p molecules [41]. These molecules have a binding site on the 3′-UTR region of the BRG1 gene (Brahma-related gene-1) and, after integrating the exosome with the tumor cell membrane, inhibit the expression of this gene. Since this gene’s product inhibits metastasis, M2 derived-exosomes increase tumor cell metastasis by inhibiting the expression of this gene [42]. Besides, miR-21-5p enhances the cellular growth and drug resistance of tumor cells by targeting PTEN (phosphatase and tensin homolog), P21 (account for cell cycle arrest), PCD (programmed cell death), and apoptotic protease activating factor 1 (APAF1) [43]. It has also been shown that miR-155-5p in M2 macrophages derived exosomes after transfer to colon cancer cells by targeting zinc-finger-type-containing 12B (ZC3H12B) leads to increased IL-6 expression, immune escape and tumor progression in colon cancer [44].
Neutrophils-derived exosomes
Mass spectrophotometric analyses have identified 271 unique proteins within neutrophil exosomes, and their amounts vary in activated and non-activated neutrophil-derived exosomes [45]. As neutrophils are one of the first cells present at the site of inflammation, their exosomes play an important role in innate immunity. Innate immune response to infections requires the interaction of activated neutrophils with platelets which bind to circulating neutrophils via the interaction of P-selectin by glycoprotein ligand-1 (PSGL-1). This binding induces neutrophil exosome production through platelet Iba glycoprotein (GPIba). These exosomes, which contain high levels of arachidonic acid, are then transferred to intracellular structures rich in cyclooxygenase-1 (COX-1) when integrating with the platelet membrane. Exosomal arachidonic acids are converted to thromboxane A2 (TxA2) by COX-1 function. Platelet-produced TxA2 enhances the neutrophil responses by inducing endothelial cells to express intracellular adhesion molecule-1(ICAM-1), stimulating rolling, and enhancing the neutrophil migration out of vessels. Inhibition of the COX-1 function suppresses neutrophil migration and inhibits innate immunity. As a COX-1 inhibitor drug, aspirin has been widely used to reduce platelet function, which also leads to neutrophil inhibition and fever reduction [46]. Moreover, activated neutrophil-derived exosomes can interact with airway smooth muscle cells and alter their proliferative properties. Some proteins are identified in these cells’ exosomes can stimulate proliferation in their target cell, thus suggesting that these exosomes may play a role in airway structural changes in asthma [45]. Recent studies also show that neutrophil-derived exosomes by transfer miR-30d-5p induces polarization of M1 macrophages and initiate macrophage pyroptosis by activating NF-κB signaling. This study indicates that neutrophil-derived exosomes contribute to the pathogenesis of sepsis-related acute lung injury (ALI) by inducing inflammatory responses in macrophages [47].
Mast cells-derived exosomes
Mast cell-derived exosomes carry external antigens with HSP-60 and HSC-70 as complexes and can perform diverse functions under different conditions. Mast cells can activate B and T lymphocytes by their produced exosomes under both, in vivo and in vitro conditions. External antigens present in mast cell exosomes are delivered to T cells and activates them through cross-presentation by DCs. CD91, as an endocytic receptor for this complex expressed in DCs, captures the HSP-60 or HSC-70-antigen complex. After antigen processing, these antigens are presented on cell surfaces through the MHC-II pathway. Furthermore, as mast cell exosomes contain HSP-60 and HSC-70, the expression of costimulatory molecules such as CD80, CD86, and CD40 is increased after integration with DCs. These exosomes can stimulate the maturation of DCs and converts them into better cells for antigen presentation [48]. Likewise, mast cells-derived exosomes can activate T cells through OX40L-OX40 interaction in vitro by a non-MHC-dependent pathway and stimulate their differentiation into Th2 cells [49]. These exosomes increase plasminogen activator inhibitor-1 (PAI-1) gene expression by affecting the endothelial cells due to their TNF-a precursor, angiotensin, and proteins involved in thrombin production. The product of this gene is a serine protease inhibitor and increases the risk of thrombosis and atherosclerosis [50]. Other actions performed by mast cell-derived exosomes include stimulating tumor cell proliferation in lung adenocarcinoma. These exosomes increase the expression level of cyclin D1 and accelerate tumor cell proliferation by inducing the transfer of KIT protein that binds to the SCF in the tumor cells and leads to the PI3K/AKT signaling pathway [51].
Eosinophils-derived exosomes (EOXs)
These cells produce exosomes that have autocrine effects on the eosinophils themselves. One of the EOXs functions is to stimulate the production of reactive oxygen species (ROS) and nitric oxide (NO) by eosinophils [52]. These mediators increase inflammation by augmenting the TNF-α, CCL26, and POSTN gene expression; POSTN is a gene that can produce periostin protein and function as a ligand for some integrin receptors responsible for the eosinophilic migration. EOXs consequently prolong inflammatory conditions, leading to damage to the airway epithelium, thus playing a role in the pathology of asthma [53]. Further, EOXs carries chemotactic factors such as S100A8, S100A9, EDN, and C3a that can influence the expression of some adhesion molecules such as ICAM-1 and B1integrins family, which leads to increased cell adhesion and eosinophil migration [52]. Studies on EOXs role in the pathology of asthma have revealed that the composition of EOXs in patients with asthma and healthy individuals are similar in size and carries important enzymes such as EPO (Eosinophil peroxidase), MBP (Major basic protein), and ECP (Eosinophil cationic protein) [54]. But the critical point is that in patients with asthma, eosinophils produce more amount of exosomes compared to healthy people, which mediate tissue damage in asthma [55].
NK cells-derived exosomes
Resting and activated NK cells constantly produce and secrete exosomes [56], and activated NK cells derived exosomes can activate non-activated NK cells. These exosomes express common NK cell markers like CD56, NKG2D, NCRs [57] and killer-related proteins such as perforin and FAS-L (membrane-bounded and soluble forms). Analyses have shown that the expression levels of perforin and FAS-L in the activated and inactivated NK exosomes are the same. Interestingly, however, the molecular weight of FAS-L is normally about 40 kDa, but in NK cells exosomes, the molecular weight of this molecule is 50 kDa [56]. The acidic microenvironment of tumors may suppress immune responses and reduce chemotherapeutic efficacy. Nevertheless, on the other hand, this acidic environment could increase the accumulation of exosomes in the tumor microenvironment, thereby enhancing the fusion of the exosome membrane with the tumor cell membrane [58]. The uptake of NK cell-derived exosomes by tumor cells induces apoptosis through perforin and Fas-L in them [59]. Likewise, NK cell exosomes play a cytotoxic role against the activated immune cells, thereby maintaining the homeostasis of immune responses [60]. The Yutaka Enomoto et al. study results show that stimulation of NK cells by a combination of IL-15 and IL-21 leads to an increase in their cytotoxic activity. However, further studies have shown that this increased cytotoxic capacity is not related to cytotoxic granules and is mediated by their EVs, including exosomes. Finally, mass spectrometric analyzes in different experimental groups showed that the level of CD226 (DNAM-1) on the surface of activated NK cells (by IL-15 + IL-21) derived exosomes was significantly increased. These exosomes are captured by tumor cells through macropinocytosis, and blocking CD226 (using monoclonal antibodies) reduces the cytolytic activity of activated NK cells exosomes and EVs [61].
B lymphocytes derived-exosomes
B lymphocytes activated through B cell receptor (BCR) or TLR secrete more exosomes than the inactivated B cells [62]. These exosomes express MHC-I and II and CD40 CD54, CD63, CD80, CD81, and CD86. In addition, B cell-derived exosomes express CD19, which is a specific marker of B cells [63]. Also, they carry the MHCII-peptide complex and stimulate T lymphocytes activation through binding to them. Similarly, these exosomes can bind to follicular dendritic cells (FDCs) as they contain α4β1 integrins that interact with VCAM-1 on the surface of FDCs. These cells do not produce MHC-II molecules, but they are observed in inactive and secondary forms on FDC surface. After binding of B lymphocyte-derived exosomes to FDCs, MHC-II molecules are expressed by dressing phenomenon on their surfaces. This molecule on the FDC surface is necessary to clonal select in the secondary follicle light zone to induce the affinity maturation phenomenon in B lymphocytes [64]. These exosomes also express β1 and β2 integrins. By the expression of β1 integrins, they can bind to TNF-a-activated fibroblasts and collagen type I and fibronectin in the extracellular matrix (ECM) [65]. Some studies have shown that B lymphocyte exosomes can directly stimulate T lymphocytes activation. Still, B cell-derived exosome-mediated activation of T cells has led to the production of cells that have little proliferation and cytokine production compared to the usual mode of T lymphocyte activation [66].
In contrast, other studies have shown that the activatory potential of B cell-derived exosomes is due to their interaction with APCs and their stimulation [27]. Direct activation of T lymphocytes by B lymphocyte exosomes depends on several factors, namely the affinity of T cell receptor (TCR) to the MHC-II-peptide complex on exosomes, the concentration of exosomes, the expression levels of costimulatory molecules on the surface of exosomes, and also the cell type and phenotype of exosome origin and target cells [63]. Environmental changes can affect the B lymphocytes derived exosomes’ internal cargos. For example, in response to heat stress, B cells produce exosomes with a high amount of heat shock proteins (HSPs) such as HSP-27, HSC-70, HSP-70, and HSP-90 increases [67].
T lymphocytes-derived exosomes
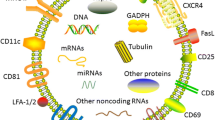
Naïve T lymphocytes produce exosomes that modulate the activity of immune cells and express the specific TCR and various adhesion molecules [68]. Besides, T cell exosomes express other markers such as CD3, CD2, CD4, CD8, CD11c, CD25, CD69, LFA-1 (lymphocyte function-associated antigen1), CXCR4, FASL, and GITR (glucocorticoid-induced TNF receptors family-related gene) [69]. Exosomes produced from activated T-cell with IL-2 increase the proliferation of resting autologous cells [70]; they carry a large amount of microRNAs and affect the functions of target cells upon integration with them. Immunological synapses enhance T cell exosome transferring efficacy between T-cell and APCs. During this process, T cell exosomes convert APCs into better cells for antigen-presenting by transferring miRNAs [71]. Exosomes produced from activated CD8+ T cells can activate ERK and NF-κB signaling pathways in melanoma cells, increasing the metastasis of cancerous cells in vitro by increasing matrix metalloproteinase-9 (MMP9) expression [72].
Regulatory T lymphocytes (Treg) produce and secrete large amounts of exosomes which express markers such as CD25, CTLA-4, and CD73 and suppress the immune system through various mechanisms. CD73, on the surface of these exosomes, plays an important role in suppressing immune system responses by inducing adenosine production (role in the anti-inflammatory response) [73]. Treg exosomes prevent the formation of Th1 and IFN-γ production by transferring let-7b, let-7d, and microRNA-155 to the target cells. Likewise, many of the inhibitory functions of these exosomes are due to the presence of IL-10 and TGF-β cytokines [74]; Treg derived exosomes can also regulate the function of DCs by transmitting miRNAs. These exosomes increase the production of IL-10 by reducing the production of IL-6 and TNF-a by transferring miR-150-5p and miR-142-3p to DCs [75].
Microarray analyses have identified 257 different types of lncRNAs in exhausted and non-exhausted TCD8+ cell exosomes. Functional analyses show that these lncRNAs regulate various TCD8+ processes, such as metabolism and gene expression. Since exhausting and normal T cell exosomes have different lncRNAs, their function is different. Exhausted T cell exosomes are captured by the normal TCD8+ cells and alter their function [76]. During this transfer, the rate of proliferation and production of cytokines in non-exhausted TCD8+ cells decreases, and the levels of inhibitory markers such as PD-1 and TIM-3 increase at their surfaces [76, 77]. Because lncRNAs are the main factor in this action, exosomes must integrate with the target cell membrane [78].
Due to the associated problems with CAR-T cell therapy, namely CAR-T cell cytokine release syndrome (CRS), which can lead to hypotension, nausea, tachycardia, and headache [79], some researchers have suggested the application of their exosomes as therapeutic agents. Because normal T-cell exosomes express TCR, it is suggested that CAR-T cell exosomes can also express the chimeric receptor and mimic CAR-T cell receptor characteristics; they have advantages over the use of CAR-T cells. First, they can transfer granzyme to the target cell along with lysosomal enzymes without the need for perforin by fusion with the target cell or via endocytosis [80]. Second, exosomes being very small in size, can cross the blood-brain barrier and tumor-forming blood barrier and perform their functions on the target cell [81]. Third, as antitumor drugs can be loaded into these exosomes, the effectiveness of the combat against the tumor is significantly increased as these exosomes can deliver anticancer drugs specifically to tumor cells [82]. In addition to the above, exosomes tend to bind to the ECM of solid tumors, which increases their therapeutic efficacy [72].
According to the above, exosomes derived from each immune cell can affect the function of other immune and non-immune cells. Figure 2 summarizes the intercellular communication of immune system cells by exosomes and their effects (Fig. 2).

Interrelationship between immune cells by producing exosomes. Each immune cell secret exosomes and release them into the microenvironment. These exosomes can affect other immune cells’ functions by suppressing or activating them and so affects immune responses
Role of immune-cell derived exosomes in diseases
Immune system cells play a crucial role in the pathogenesis of various diseases by producing exosomes. Exosomes from different cell sources transmit different cargos to the target cell and can change their behavior and functions. Since many of these cells trigger inflammatory responses to remove pathogens, their exosomes can play a role in inflammatory diseases such as tissue destruction, autoimmunity, and allergies. Further, some immune cells may help modulate responses and homeostasis by suppressing immune responses. These regulatory cells produced exosomes can contribute to the development of cancer and infectious diseases. Table 1 summarizes the role of IEXs in the pathogenesis of various diseases (Table 1).
Role of IEX in cancer
In the tumor microenvironment, diverse cells such as immune cells actively interact with their surroundings through EVs and facilitate tumor malignancy by regulating malignant cascades such as immune modulation, angiogenesis, drug resistance, cell proliferation and metastasis [21]. Immune cells located in these environments are altered by exosomes derived from tumor cells and contribute to tumor progression [21, 83].
Role of IEX in tumor enlargement by increasing tumor cell proliferation
Microarray analyses have shown that M2 macrophages contain large amounts of miR-21-5p, miR-24-3p, and miR-221-3p. Because miR-221-3p directly suppresses the cyclin-dependent kinase inhibitor 1B (CDKN1B), these exosomes can resolve the cell cycle arrest in the G1/S and help tumor cells proliferate. Furthermore, in vivo analyses indicate that M2 macrophage-derived exosomes play a role in epithelial ovarian cancer (EOC) [84].
Role of IEX in preventing inflammatory responses in immune cells
Treg/Th17 cell imbalance occurs in many diseases such as human systemic lupus, diabetes, and some tumors. As mentioned, M2 macrophage exosomes contain a variety of miRs. A study by Zhou et al. shows that M2 macrophage exosomes transfer miR-29a-3p and miR-21-5p (with a synergistic mechanism) to TCD4+ cells. These exosomes disrupt the Treg/Th17 balance by stimulating the differentiation of Naive cells to Treg by inhibiting the STAT3 transcription factor. During this procedure, the production of inflammatory cytokines such as IL4, IL6, and TNF-a decreases, and anti-inflammatory cytokines like IL10 increase. Thus, these exosomes suppress T cell anti-tumor responses by stimulating the differentiation of TCD4+ cells into Treg and contribute to tumor progression [85].
Myeloid-derived suppressor cells (MDSCs) in the tumor microenvironment are more immune suppressive than MDSCs in peripheral lymphatic organs [86]. In vivo and in vitro analyses show that exosomes produced from MDSCs can suppress immune system cells such as macrophages and T cells in the tumour environment. A study by Rashid et al. found that the use of MDSC exosomes increases activating markers (CD69, CD44) and also exhausting markers (PD-1/CD279) by repetitive stimulation of TCD8+ cells, which leads to induction of a state of non-responsivity (exhausting). Repetitive activation of TCD8+ cells leads to increased ROS production in these cells. Furthermore, since MDSC exosomes express high Fas-L levels at their surface, they can induce activation-induced apoptosis (AICD) in TCD8+ cells. Thus, in general, MDSCs-derived exosomes inhibit TCD8+ cell function by inducing exhausting conditions, ROS production, and apoptosis induction through Fas/FasL (CD95/CD95L) interaction and contributing to tumor tumours progression [87].
Role of IEX in the migration/invasion of cancer cells
The M2 macrophage-derived exosomes carry miR-21-5p and miR-155-5p, enhance colon cancer cells’ migration and metastasis, and contribute to tumor progression. These miRs inhibit the expression of Brahma-related gene 1 (BRG1) by binding to this gene. Since BRG1 plays a vital role in inhibiting these cells’ metastasis, its inhibition leads to increased metastasis and invasion of colon tumor cells [42]. Yang et al. illustrated that macrophages derived from monocytes treated with IL-4 produce exosomes containing larger amounts of miR-223 and inhibit the expression of myocyte-enhancing factor- 2c (Mef-2c) by binding to 2 target sites in 3′-UTR after transfer to a breast cancer cell. Because Mef-2c depletion is associated with β-catenin nuclear accumulation, the rate of cell migration and metastasis in breast cancer cells increases [88]. Mass spectrometric analyses of M2 macrophages-derived exosomes indicate apolipoprotein E (ApoE) on their surface and increase metastasis in gastric cancer cells after their fusion and transfer of ApoE to them. ApoE is an important regulator of tumor progression through increased cell adhesion, proliferation, and migration with cytoskeletal rearrangement through the PI3K/Akt/mTOR signaling pathway [89].
The TCD8+ cells in the tumor microenvironment are transformed into a non-responsive state through factors secreted by the tumor cells. Cai et al. demonstrated that these cells produce FasL+ and CD8+ exosomes in the tumor microenvironment and enhance the metastasis in the B16 melanoma cell line. These exosomes activate the NF-κB, ERK, and c-FLIPL signaling pathways through the Fas/FasL-dependent trail, leading to MMP2, MMP9, and COX2 increased expression in B16 cancer cells, which play a critical role in cancer cell metastasis [72].
Role of IEX in drug resistance and reduction of tumor cell apoptosis
M2 macrophages, DCs, and T cells derived exosomes carry large amounts of miR-21, which can induce drug resistance in cancer cells by binding to APAF1. The binding of APAF1 to cytochrome c leads to caspases 9 and 3 activations and apoptosis initiation. Thus, the inactivation of APAF1 function can lead to drug resistance in cancer cells. However, APAF1 cannot be entirely responsible for the effect of miR-21 on increasing paclitaxel resistance in ovarian cancer [43]. Other studies have shown that miR-21 can increase the survival and drug resistance to tamoxifen, faslodex, and topotecan in breast cancer therapy by targeting other genes such as PTEN, PDCD4, and P21 [90].
A 2019 study by Chen et al. showed that M2 macrophage derived-exosomes contain a lncRNA named long non-coding RNA that stabilizes HIF-1α (HISLA). These exosomes inhibit the interaction between the prolyl hydroxylase 2 (PHD2) and hypoxia-inducible factor 1-alpha (HIF-1α) and prevent the hydroxylation and degradation of HIF-1α after fusion with breast cancer cells via HISLA transfer [91]; HIF-1α is an oxygen-sensing transcription factor that determines the type of glucose consumption by oxidation or glycolysis [92]. Therefore, increasing the level of HIF-1α leads to increased aerobic glycolysis and enhanced resistance to apoptosis in cancer cells [93].
Role of IEXs in transplant rejection
EVs, including exosomes, play an important role in transplant rejection by delivering antigens to immune cells. Exosomes produced by APCs are abundant in MHC and T cell-associated adhesion molecules on their surface. Allograft derived exosomes (Allo-Exo) released by donor leukocytes “throw out” from the connective tissue and pass through the pores of the recipient’s lymphatic capillaries into the transplanted drainage lymph tissue [94].
Many studies show that donor-derived DCs levels in transplanted lymph nodes are undetectable in acute allograft rejection; therefore, it is likely that other mechanisms are involved in this type of transplant rejection. Recent studies have shown that donor DEXs mimic the characteristics of DCs (having MHC-I and MHC-II and helper molecules) and migrate to the draining lymph node, and present the MHC to alloreactive T cells to stimulate the anti-alloantigen responses [95].
Exosomes have a limited ability to activate T cells due to the low number of MHC-peptide complexes and costimulatory molecules as APCs capture them in phagocytosis or other ways [96]. These cross-dressed APCs express exosome-derived MHC-peptide complexes on their surface and activate alloreactive T cells with higher efficiency [97]. Recent studies show that the innate immune system cells are also involved in transplant rejection, wherein monocyte-derived DCs are produced continuously to participate in the transplant rejection process [98] in mice with defective T cells, B cells, and innate lymphocytes. Since Allo-Exo’s main role in transplant rejection has not been precisely elucidated, several actions can be attributed to them, such as increasing macrophage phagocytic activity, providing antigen for presentation, and stimulating the monocytes’ maturation to inflammatory DCs [99, 100].
IEXs role in the pathogenesis of asthma and allergic diseases
Asthma is a chronic inflammatory disease of the respiratory tract with a complex pathophysiology and is classified as a type 1 hypersensitivity disease [101]. Exosomes play an important role in the pathogenesis of asthma and intercellular signaling by transporting various cargoes to target cells. They can increase inflammation by regulating the immune cells’ function (increasing recruitment, activation, or differentiation) [102].
Previous studies have shown that DCs and macrophages derived exosomes contains lipid mediators derived from arachidonic acid such as leukotriene A4 (LTA4) [54] and participate in the pathogenesis of some allergic diseases and chronic inflammation. Besides, DEXs contain enzymes that can convert LTA4 to other leukotrienes (LTs), such as LTB4 and LTC4. Since LTB4 is also a chemotactic factor, it increases inflammation by attracting other immune cells to the inflammation site [5]. Thymic stromal lymphopoietin (TSLP)-activated DCs also induce T cell responses and their differentiation to Th2 through the interaction between OX40-OX40-L with T cells, which can help in IgE production [103]. Lymphocytes are key players in the inflammatory response in allergies and asthma. B cell-derived exosomes present allergen-derived peptides such as birch peptide (Bet v1) to T cells and induce their differentiation to the Th2 phenotype [63]. Mast cell exosomes stimulate IgE production in B cells through the CD40-CD40L interaction in the absence of T cells. Besides, it has been shown that these exosomes can stimulate the production of pro-inflammatory cytokines by T lymphocytes and bronchial smooth muscle cells (BSMCs) and contribute to the progression of asthma [48].
Role of IEXs in the progression of infectious diseases
Exosomes produced by infectious agents can lead to the spread of infection and prevent immune system responses [104]. Also, infected cells with intracellular pathogens can secrete exosomes that play an important role in the infection’s fate. They can directly carry damaging pathogen related molecules and contribute to infectious disease pathology, thus leading to the spread of infectious diseases [105].
Human T-lymphotropic virus type 1 (HTLV-1) infected T cells produce exosomes containing various viral proteins that contribute to the pathogenesis of HTLV-1-induced disease. These exosomes contain Tax proteins and transcripts of viral mRNAs such as Tax, HBZ, and Env [106], which are translated into proteins by the host cell and exert their functions; Tax is an important protein for regulating the dynamics of the viral life cycle and regulating various genes and can activate the NF-κB, and Akt pathways which is two main cell survival pathways in target cells.
Furthermore, Tax can play a role in the disease’s pathogenesis by damaging cellular DNA in two ways. This molecule disrupts DNA repair mechanisms such as base exchange repair (BER), nucleotide exchange repair (NER), and mismatch repair (MMR). Also, Tax directly damages cellular DNA by stimulating ROS production in host cells. Thus, exosomes produced from HTLV-1 infected T cells can lead to genetic damage in other Naive T cells and prevent their function [107]. HIV-1 infected T cells also produce exosomes containing the p24 and Nef proteins and a double-stranded non-coding RNA named TAR-RNA. These exosomes can transfer viral related proteins to adjacent cells and contribute to the pathogenesis of the disease; Nef is one of the HIV-1 associated proteins and helps to virus replication and progression of the disease [108]. Monocytes are among the most important sources of HIV-1 infection by this virus. Analyses results show that Nef-expressing monocytes produce AATK, SLC27A1, and CDKAL mRNAs containing exosomes. Transfer of This mRNA to surrounding TCD4+ cells induces apoptosis, disrupts fatty acid metabolism, and thus participates in the pathogenesis of HIV-1 [109]. Another study showed during infection with human herpesvirus-6 (HHV-6), infected cells such as TCD4+ [110] produce exosomes that have a number of complete and mature viruses and are transmitted to other cells, thus, leading to the spread of infection [111]. Figure 3 summarizes the role of IEXs as double-edged swords in disease progression as well as their therapeutic applications (Fig. 3).

IEX has a role in the pathogenesis of the disease. Also, these exosomes play a role in various disease treatments. As shown in the figure, IEX therapeutic application is an exciting subject used for autoimmune disease, cancer, asthma, allergy, transplantation, and infection treatment
Role of IEXs in the pathogenesis of autoimmune disease
Type 1 diabetes (T1D)
Guay et al. showed that human and rodent T cells derived exosomes carry the miR-155, miR-142-5p, and miR-142-3p, which can stimulate the secretion of different chemokines including CCL2, CCL7, and CXCL10 and regulate the NF-κB related signaling pathway and apoptosis in pancreatic beta cells in T1D. These chemokines may promote immune cells’ recruitment and increase the selective death of insulin-secreting cells. Inactivation of these miRs in NOD mice increases insulin levels, decreases insulitis scores and inflammation, and also prevents the progression of diabetes [112].
Type 2 diabetes (T2D)
Zhang et al. found that cytokines secreted by adipose tissue macrophages (ATMs) significantly altered the adipocyte function, inducing inflammatory responses and stimulating M1 macrophage differentiation. Exosomes derived from these macrophages in the presence of insulin in adipose tissue reduce the insulin-dependent phosphorylation of Akt by increasing NF-κB activation. This action blocks glucose uptake by reducing the transfer of GLUT4 to the cell surface. Thus, these exosomes increase insulin resistance due to obesity in adipose tissue, which is a significant component in T2D pathogenesis [113].
Guillain-Barré syndrome
Du et al. found that M1 macrophages-derived exosomes intensified the autoimmune neuritis (EAN), the animal model of Guillain-Barré syndrome, by increasing Th1 responses. Guillain-Barré syndrome is the most common acute paralysis neuropathy, which results from an immune system attacking the peripheral nervous system and demyelination of neurons [114]. M1 macrophage exosomes in the in vivo studies directly increase the ratio of IFN-γ and IL-17 producing CD4+ T cells in the lymph nodes and spleen, which promotes EAN progression. Evidence also suggests that these exosomes induce the expression of pro-inflammatory cytokines such as IL-6 and IL-12 in DCs and macrophages, which in turn amplifies the Th1 and Th17 responses via indirect pathways. These exosomes induce in vitro proliferation, differentiation, and effective function in CD4+ and CD8+ T cells that produce IFN-γ. Experimental results suggest that M1 macrophage-derived exosomes may affect T cells by regulating the T-bet transcription factor [115].
Multiple sclerosis (MS)
Studies show that exosomes secreted by T cells contain large amounts of miR-326, which, by targeting and suppressing the Ets-1 gene product as a negative regulator of Th17 differentiation, increases the Th17 differentiation in vitro and in vivo and thus exacerbates MS [116]. Studies show that microglia cells secrete exosomes that contain IL1-β, MHC-II, MMPs, and caspase-1 into the cerebrospinal fluid, which can cross the blood-brain barrier (BBB). These exosomes are distributed throughout the body and contribute to antigens’ rapid release and their delivery to immune cells. The enzymes in these exosomes damage the BBB and cause neuroinflammation by immune system cells migrating increase, resulting in MS. Exosomes derived from active T cells increase ICAM-1 and Mac-1 (macrophage-1 antigen) expression in monocytes. The expression of these two molecules increases the migration of active monocytes and intensifies MS [117]. It has also been shown that some of the miRs involved in the pathogenesis of MS in T cell-derived exosomes, such as miR-326, are significantly higher than in healthy individuals [116]. miR-326 helps to promote MS and myelin degradation by inhibiting CD47 expression in brain resident cells that inhibit the phagocytic activity of macrophages [118].
Immune cell-derived exosomes (IEX) - therapeutic applications
The ability of IEX to enhance or suppress immune responses has led to their therapeutic usage. Because in certain diseases, the function of some immune cells and the production of exosomes from them increases, they can be used as diagnostic markers [6]. Also, as mentioned earlier, the exosomes from different cells of the immune system cells have dissimilar contents that lead to different responses by affecting the target cells. These exosomes can be isolated from the patient’s immune cells and be injected as an autologous graft. Table 2 summarizes some new studies pertaining to IEXs and their deployment in treating various diseases (Table 2).
IEX therapeutic application in cancer
Tumor tissue and cells have evolved to suppress immune responses. The tumor microenvironment contains molecules and factors that suppress the immune system and prevent the development of DCs and T lymphocytes. Therefore, establishing specific, strong, and long-lasting immune responses against cancer cells and reducing immune suppressants are critical for cancer effective treatment. The results of a study by Pete et al. show that the use of DEXs has several advantages over DCs applications in cancer immunotherapy [32, 132]. At present, exosomes are preferred to cell therapy for Five reasons. (1) These vesicles can be stored for a long time without losing immune activity; (2) Exosomes, due to their membranes’ similarity to body cells, integrate with the target cell more efficiently than soluble factors produced by cells [133]; (3) They are smaller than cells and can easily pass through different capillaries [134]; (4) Ability to easily manipulate exosomes and produce engineered exosomes [135]; (5) Easier injection compared to cell therapy (possibility of nasal administration) [135].
Transformation of cancer cells into cells with higher immunogenicity
Because DEX carries many DC-related immune-stimulating molecules, tumor cells’ uptake can turn tumor cells into better immune targets. A study by Romagnoli et al. showed that in vitro treatment of SK-BR-3, U87, and K562 cell lines by mature DCs exosomes is associated with increased expression levels of MHC-I, MHC-II, CD86, CD11c, CD54, and CD18 in them. This makes the tumor cells more suitable to activate the immune system. Interestingly, when DEXs are exposed to anti-CD9 antibodies before treatment, they fuse to a lesser extent with tumor cells, indicating this molecule’s role in integrating the exosome with the tumor cell [136]. Human breast adenocarcinoma cells (SK-BR-3) treated with DEX could be used to stimulate CD3+ T cells. It has been shown (by ELISPOT analysis) that the number of IFN-γ-producing T cells in the DEX-treated tumor cells group is much more than in the untreated control group. These data suggest that treating tumor cells with DEXs increases their ability to activate T cells for a more efficient response [137].
Stimulation of tumor antigen-specific T cells responses
DEXs carry the molecules needed to activate tumor-specific immune cell response and perform as an anti-tumor vaccine alone [138]. For this purpose, DCs are first isolated from patients, and after activation with tumor-specific antigen and proliferation, these cells are injected via various routes. It has been shown that DEXs can be used as acellular vaccines that express melanoma-associated antigens (MAGE) on MHC-I and MHC-II. In a study, IFN-γ-stimulated monocytes derived exosomes increased the activity of NK cells and were associated with tumor regression and recovery in patients with lung cancer [139].
In a study by Lu et al., it has been shown that alpha phytoprotein (AFP) activated DCs derived exosomes can stimulate antigen-specific immune responses and increase tumor regression in mice with hepatocellular carcinoma (HCC) [140]. Also, the measurement of splenic IFN-γ producing TCD3+ cells by fluorescence-activated cell sorting (FACS) reveals a significant increase in the number of these cells in the DEX-AFP-treated group compared to the control group. This study illustrated that T cells’ production of inflammatory cytokines such as IFN-γ and IL-2 has increased, and the level of inhibitory cytokines such as TGF-β1 and interleukin-10 increased as well [140, 141]. Stimulation of DCs by TLRs ligands such as poly I:C (TLR-3 ligand), LPS (TLR-4 ligand), and CpG DNA (TLR-9 ligand) lead to the production of DEXs that activate the immune system and significantly increase their ability for anti-tumor therapy. The use of melanoma B16F10 cell lysates combined with poly (I: C) to stimulate DCs results in DEX production that can stimulate CD8+ T cells; these exosomes also increase recruitment cytotoxic CD8+ T, NKT, and NK cells to the tumor site. As a result, tumor growth is significantly reduced, and the patient’s survival rate increases [142].
Martina Damo et al. reveal that exosomes isolated from matured DCs with albumin purified antigen and poly (I:C) antigens (DEX (OVA + pIC)) supernatant stimulate more robust Th1 responses by secretion of IFN-γ and TNF-α as proinflammatory cytokines, compared to exosomes isolated from DCs matured by LPS and OVA (DEX (OVA + LPS)) as well as DCs matured by CpG and OVA (DEX (OVA + CpG)). These cellular immune responses stimulate the tumor regression in the absence of OVA-specific antibodies, indicating that DEXs do not play a role in producing tumor-specific antibodies [142]. Another study by Chen et al. confirmed the results of a previous investigation wherein HPV E749-57 antigens in combination with poly (I:C) had been used to stimulate DCs maturation to produce DEX (Dex (E7 + pIC)) for the treatment of cervical tumors. The use of this Dex (E7 + pIC) based vaccine induces a specific CTL response against E749-57 [143].
Stimulation of apoptosis in tumor cells
Previous studies have shown that hematopoietic and tumor cells may secrete HSPs into the bloodstream through granulomas, exosomes, or exocytosis [144]. These extracellular HSPs stimulate innate immune responses through TLRs [145]. Treatment of DCs with CO2 hyperthermia (HT-CO2) results in exosome production in which HSP70 levels are elevated. Evaluation of these exosomes’ effect by analyzing the activity of caspase-3 and measuring CCK-8 indicates an increase in apoptosis and a decrease in proliferation in the AGS cell as a gastric cancer cell line [146].
miR-let-7a-5p is present in large amounts in the M1 macrophages exosomes, and this miR targets four genes, including BCL2-like protein 1 (BCL2-L1), insulin-like growth factor 1 receptor (IGF1R), mitogen-activated protein kinase 8 (MAPK8), and Fas [147]. These genes are functionally divided into two groups involved in autophagy, including BCL2-L1, IGF1R, and MAPK8, and the group of genes involved in apoptosis, encompassing BCL2-L1, Fas, and MAPK8. Duan et al. showed that let-7a-5p decreased BCL2-L1 gene expression by stimulating the PI3Kγ signaling pathway and increasing autophagy and apoptosis in lung cancer cells [148].
NK cell exosomes express the DNAX-1 accessory molecule (DNAM1) on their surface, enabling exosomes to integrate with the tumor cell membrane by binding to PVR and Nectin-2. Integration of NK exosomes containing perforin and granzyme leads to the onset of apoptosis in the NALM-18 cell line (associated with acute lymphoblastic leukaemia in childhood) due to caspase-8 activation [149]. Exosomes derived from NK cells also contain granulysin that disrupts the tumor cell membrane integrity [150]. These exosomes induce apoptosis in tumor cells by damaging the mitochondria and activating caspase 9 and 12 [151]. In a study by Jong and colleagues, it has been shown that NK cell exosomes induce several caspase pathways by using different cytotoxic molecules in SupB15 cells associated with acute lymphoblastic leukaemia (ALL) and neuroblastoma cancer cell lines [152].
Inhibition of metastasis, migration, and proliferation in tumor cells
Given that approximately 70% of ovarian cancer cells express high levels of epidermal growth factor receptor (EGFR) and the involvement of this receptor in angiogenesis and metastasis processes, the inhibition of this receptor could offer a potential treatment for ovarian cancer [153, 154]. Macrophages that are activated by weakly induced apoptosis from the TNF family cytokine (TWEAK) [155] produce exosomes that contain some more microRNA-7 (miR-7) relative to macrophages that are activated commonly. These exosomes are captured by ovarian cancer cells, and miR-7 decreases these cells’ metastasis by inhibiting the EGFR/AKT/ERK1/2 signaling pathway [156, 157].
Due to the significant role of tumor-derived MSCs in angiogenesis and the invasion of tumor cells [158], the use of mechanisms that lead to the death of these cells reduces the rate of tumor angiogenesis and metastasis. Studies by Seo et al. reveal that exosomes produced by CD8+ T cells containing miR-298–5p reduce these cells’ population. Further, the study also showed that the transfer of CD8+ T cell exosomes produced by healthy mice could kill tumour-derived MSCs in the same species without H-2 (mouse MHC-1) restriction [159].
The results of new studies show that M1 macrophage-derived exosomes suppress proliferation, migration and invasion but lead to the induction of apoptosis in tumor cells. M1 exosomes affect tumor cell function by transferring lncRNA HOXA transcript at the distal tip (HOTTIP) to head and neck squamous cell carcinoma (HNSCC) [160]. However, the antitumor function is attenuated by HOTTIP-knockdown exosomes, indicating that HOTTIP acts as a key molecule in M1 exosomes. Further analysis of the role of HOTTIP lncRNA in tumor cells showed that overexpression of HOTTIP prevents proliferation, migration, and invasion but induces apoptosis of cancer cells in vitro [161]. Finally, the relevant data showed that exosomal lncRNA derived from M1 macrophages suppressed the progression of HNSCC by positively regulating the TLR5/NF-κB signaling pathway through competitive sponging miR-19a-3p and miR-19b-3p [161]. Also, it has been proven M1 macrophage-derived exosomes containing HOTTIP can induce circulating monocytes polarization to the M1 antitumor phenotype [161]. Therefore, it is suggested that the use of these exosomes may contribute to anti-tumor immunity. Also, Müller et al. Showed that NK cells derived exosomes could lead to the apoptosis of tumor cells. Further analysis showed that these exosomes inhibited the growth of MYCN- amplified neuroblastoma cells proliferation by transporting miR-186 [132, 162].
Decrease of drug resistance in tumor cells
Due to the inflammatory properties of M1 macrophages derived exosomes, they can be used in the treatment of various tumors. In a study by Zhang et al., cisplatin was loaded in exosomes (exoCIS) by sonication after separating from M1 macrophages. This study shows that the application of drug encapsulated exosomes leads to an increase in apoptosis of A2780/DDP (drug-resistant) and A2780 (drug-sensitive) cell lines compared to the use of cisplatin alone. Interestingly, cytotoxicity in drug-resistant cells has been significantly increased compared to sensitive cells. It seems that the two mechanisms are involved in the anti-cancer activity of exoCIS: (1) Preferred accumulation of cisplatin by exosome in cancer cells and (2) Intrinsic properties of exosomes derived from M1 macrophages to kill tumor cells [163].
IEX therapeutic application in autoimmune diseases
Autoimmune diseases occur when abnormal immune system responses damage the body tissues and severely affect a patient’s quality of life. Due to immune system exosomes’ role in modulating immune responses, they are now used to treat many autoimmune diseases such as multiple sclerosis, rheumatoid arthritis, inflammatory bowel disease, and myasthenia gravis.
Improve multiple sclerosis (MS) and experimental autoimmune encephalomyelitis (EAE)
In vivo analysis show that the use of IFN-γ-induced DCs derived exosomes (IFN-γ-DEX) through the transfer of various miRNAs increases the central nervous system (CNS) myelination in patients with MS and demyelination syndromes. Among the miRs involved in this process is miR-219, which plays a crucial role in myelin formation and its maintenance [164], miR-9, which is produced during oligodendrocyte maturation [165], and increases the oligodendroglial differentiation, and miR-17–92 that increase the proliferation of oligodendrocyte progenitor cells [166]. Finally, it can be articulated that these exosomes stimulate CNS myelination by increasing the proliferation and differentiation of oligodendrocyte progenitor cells, increasing the antioxidant activity of microglia, and modulating the inflammatory environment [167].
Yu’s study showed that using membrane TGF-β1 expressing DEXs (mTGF-β1-EXOs) in mice with EAE inhibited the progression and development of the disease. Treatment of mice with these exosomes also reduced Ag-specific Th1 and IL-17 mediated responses. It increased IL-10 related responses in ex vivo as they help cure the disease by increasing Treg cells’ differentiation and reducing the inflammation-associated damages [168]. Given that T cell inflammatory responses are one of the important factors in the development of MS, the use of Treg derived exosomes to inhibit their function can be helpful; studies reveal that exosomes produced from Treg in people with MS have fewer immunomodulatory effects compared to healthy people. Therefore, exosomes derived from the Treg of healthy individuals can effectively treat autoimmune diseases such as MS and EAE. These exosomes reduce proliferation in conventional T cells and induce apoptosis to prevent T cell reactions [169].
Myasthenia gravity (MG)
A study by Bu et al. in 2015 showed that the use of immature dendritic cell exosomes (imDEX) prevented the progression of the experimental autoimmune Myasthenia Gravity (EAMG) in mice as they suppress the responses of B cells (producing anti-AchR antibody) and AchR specific T cells (producing inflammatory cytokines) [170]. In a study by Yin, miR-146a was used to produce tolerogenic DCs. These cells produce exosomes that express a reduced amount of CD80 and CD86 molecules, while the amount of MHC-II molecules on these exosomes’ surfaces has not changed. The results show that exosomes derived antigen-specific immature DCs reduce the clinical signs of EAMG mice by decreasing the serum levels of anti-AchR, IgG1, and IgG2b, as well as shifting the T cell responses from Th1/Th17 to Th2/Treg [171].
Inflammatory bowel disease (IBD)
Due to the anti-inflammatory properties of MDSC, they can be deployed in the treatment of autoimmune diseases [172], as illustrated in a study where this cell’s exosomes have been used to treat dextran sodium sulfate(DSS)-induced colitis. Due to the presence of high levels of arginase-1 (Arg-1) in MDSC-Exo, these exosomes suppress inflammatory responses and help treat induced colitis in mice by reducing the number of Th1 cells and increasing Treg. Also, it has been observed that serum levels of IFN-γ and TNF-α were decreased significantly in exosome-treated mice compared to controls. Interestingly, the inhibition of Arg-1 activity in MDSC-Exo significantly reduces the exosome therapy’s recovery effect [173].
Rheumatoid arthritis (RA)
RA is a chronic and systemic autoimmune disease in which chronic inflammation of the joints causes joint damage. The use of DEXs due to their characteristics is one way to reduce inflammation and treat this disease. Various studies show that some strategies have been applied to produce inflammation suppressing DCs (TolDCs) that produce such anti-inflammatory exosomes. These include genetically modified DCs for IL-4 expression [174], treatment of DCs with IL-10 [175], and enzyme indoleamine 2,3 deoxygenase (IDO) containing DCs [176]. These exosomes can modulate the APCs and T cells’ activity through the MHC-II- and FasL/Fas dependent mechanisms. Besides, they can also suppress delayed-type hypersensitivity (DTH)-related inflammatory responses and improve collagen-induced osteoarthritis [174]. Furthermore, various studies have shown that IDO-expressing DCs derived exosomes suppress the CD8+ T cells’ activity by reducing tryptophan dose and producing cytotoxic metabolites, leading to the enhanced differentiation and activation of Treg cells [176].
IEX therapeutic application in transplant rejection
Some immune cells with tolerogenic and immunosuppressive properties produce exosomes that can be applied to prevent tissue transplant rejection in the host. For example, PD-L1 expressing TolDCs deliver the donor MHC molecules to the recipient by tolerogenic methods and suppress the response of effector T cells produced against transplantation through PD-L1 [177]. In a study by Yu et al., serum and histological analysis showed that using Treg CD4+ CD25+ exosomes leads to renal allografts’ long-term survival in the mouse model. Also, in vitro analyses revealed that the use of these exosomes suppresses T cell proliferation. Therefore, according to the results of in vivo and in vitro analyses, exosomes released from Tregs can be considered a potential immunosuppressive agent to prevent transplant rejection [178]. Furthermore, the exosomes produced by Tregs express inhibitory molecules such as CTLA-4 and CD25 on their surface [73]. After fusion of these exosomes with APCs, these molecules are placed on the surface of them and can inhibit the activation of T cells by removing IL-2 from the environment and reducing its amount by CD25 to inhibit the initiation of T cell responses by CTLA-4 [179].
IEX therapeutic application in infectious diseases
IEXs play an important role in antimicrobial responses where these exosomes operate as “bridges” between different cells and prevent the spread of viral and bacterial infections. During hepatitis C virus (HCV) infection, the interaction between resident macrophages and liver cells is a critical component of the liver’s innate immunity. In 2016, Zhou et al. showed that the transfer of TLR-3 activated macrophages supernatants (that contain exosomes) to Huh7 cells (human liver cell line type) prevents the proliferation of HCV within them; these exosomes carry some members of the miRNA-29 family such as miR-29a, 29b, and 29c and have an anti-HCV role [180].
LPS activated mature CD8+ DCs secrete exosomes with high levels of MHC-1 and ICAM-1 at their surface, which are captured by other DCs through LFA-1-ICAM-1 interaction, and transfer the processed antigen and the peptide-MHC complex to the target cell. These DCs cannot reprocess antigens, but they can enhance T cell activation in the presence of exosome-associated MHC-peptide complex and the delivery of exogenous antigens. By this mechanism, activated DCs increase the number of APCs to T cells [181]. DEXs can also carry microbial TLR ligands and transfer them to other DCs to activate them and increase the antigen presentation and production of inflammatory cytokines such as TNF-α. Further, these exosomes increase NK cell function and differentiation of CD4+ T cells into Th1 [182]. Since more than 90% of healthy individuals produce effective responses against mycobacterium tuberculosis, and because infected DCs and macrophages have a limited ability to deliver antigens, other mechanisms such as exosome production play a very effective role in the activation of T cells [183]. The inhibition of exosome production by tuberculosis antigens presenting DCs and macrophages leads to decreased immune response and increased bacterial load. Defects in exosome production, such as those that occur in Rab27a-deficient mice, significantly reduce the bacterial antigen trafficking and immune system responses. So, DC and macrophage-derived exosomes can be used to stimulate T-cell mycobacterium tuberculosis-specific responses to and prevent the disease progression [184]. Another study that confirms the therapeutic function of IEXs in the treatment of microbial infections pertains to Helicobacter pylori-infected DCs and macrophages-derived exosomes. These exosomes carry miR-155 in large quantities and transport them to other macrophages during vesicular trafficking. Transfer of these exosomes to macrophages increases the production of inflammatory cytokines such as TNF-a, IL-6, IL-23, and costimulatory molecules such as CD40, CD63, CD81, and MHC-I [185, 186].
IEX therapeutic application in asthma and allergies
Exosomes produced by mast cells contain receptors with a high affinity for IgE, and they can bind to free IgE in serum via the FC𝜀R1 and decrease the concentration of this antibody in the serum. This reduction in serum IgE limits the activation of other mast cells and prevents allergic reactions. The results of a study by Xie et al. reveal that the use of bone marrow-derived mast cell in mice with chronic asthma reduces respiratory tract inflammation and has a role in the remodelling and regeneration of lung tissue; accordingly, mast cell exosomes can be used as an anti-IgE agent in various allergic diseases [187, 188].
In 2020, Li et al. described that M2 macrophage-derived exosomes could prevent the progression of asthma. In this study, to create a model of asthma in C57BL/6 mice, they used ovalbumin (OVA) and Freund adjuvant. The outcome reveals that these exosomes carry a large amount of miR-370, inhibiting the FGF1/MAPK/STAT1 signaling axis in airway epithelial cells (AECs) [189]; they inhibit cell apoptosis, pulmonary fibrosis, OVA-induced inflammation in mice, and ectopic hyperplasia in AECs. Further studies illustrate the number of granulocytes, such as neutrophils and eosinophils, and the levels of IL-1β, IL-6, TNF-α, and MCP-1 in OVA-treated mice in bronchoalveolar lavage fluid (BALF) are reduced. These findings provide a new understanding of the involved molecules in asthma development and suggest the deployment of exosomes as an alternative treatment [190]. Due to the importance of IEXs clinical applications, Table 3 summarizes the cellular sources of exosomes, mediators, mechanism, type of disease, and their application results (Table 3).
Conclusions and future prospects
As shown in Table 4, despite all the advantages of IEXs, only a handful of clinical trials have exploited these nanovesicles. Due to the existence of some unresolved limitations, the clinical application of exosomes has been challenging [191]. For example, the biological distribution of these vesicles in the individuals treated by them is not well understood, which may lead to difficult challenges in managing the use of exosomes in standardization methods related to their separation, quantification, and analysis of results [83]. Also, the difficulty in exosome storage due to their aggregation and the destruction of their cargo during the freeze-thaw process is another limitation of their limitations [192]. This problem can be circumvented by adding stabilizers such as sucrose, trehalose, and glucose [193]. The choice of an appropriate source for exosome isolation in therapeutic applications and as a delivery system for each molecule and drug is a significant issue that is often completely ignored and can greatly affect treatment outcomes [4].
Moreover, the clinical trials using IEXs show that naive exosomes are often insufficient to produce strong responses in vivo [194]. The focus has now shifted to improving the effects of exosomes by engineering them for different treatment approaches. One of these strategies is to load drugs in IEXs [195]. An example of this type of exosome is the treatment of tumors using doxorubicin encapsulated DEXs [196].
The interaction between immune system cells through the exosomes is very complex. Exosome production by immune cells leads to change in the function of adjacent cells (paracrine effect), and sometimes the cells themselves (autocrine effect), and turn them into better cells to perform their functions. Besides, the IEXs can be responsible for tissue damage caused by many diseases, such as asthma. Therefore, the function of these exosomes is not always beneficial to the body and sometimes disrupts the function of the other cells. However, due to the unique features of exosomes, including biocompatibility, low clearance rate, and ability to transfer drugs, they can be used as drug delivery vehicles [132]. Understanding the properties of diverse cell exosomes is crucial for selecting an appropriate exosome for therapeutic applications. Most of IEXs can induce inflammation, and hence they can be used to enhance immune responses to infections and tumors. In contrast, some IEXs have inhibitory roles on immune system responses that can be used to prevent graft rejection and treatment of hypersensitivity and autoimmune diseases.
Finally, it can be said that studies in this field are in their beginning, and more research is needed on the wide therapeutic application of IEX in the treatment of various diseases.
Availability of data and materials
The data supporting the conclusions of this article are all online.
Abbreviations
- IEXs:
-
Immune cells-derived exosomes
- DEXs:
-
Dendritic cell-derived exosomes
- APCs:
-
Antigen-presenting cells
- MVBs:
-
Multivesicular bodies
- ILVs:
-
Intraluminal vesicles
- ESCRT:
-
Endosomal sorting complex required for transport
- MHC:
-
Major histocompatibility complex
- HSPs:
-
Heat shock proteins
- DCs:
-
Dendritic cells
- FAS-L:
-
Fas ligand
- LPS:
-
Lipopolysaccharide
- TLR:
-
TOLL-like receptor
- TNF-a:
-
Tumour necrosis factor-alpha
- PI3-k:
-
Phosphoinositide 3-kinase
- FDCs:
-
Follicular dendritic cells
- ECM:
-
Extracellular matrix
- TCR:
-
T cell receptor
- MMP:
-
Matrix metalloproteinase
- PTEN:
-
Phosphatase and tensin homolog
- lncRNA:
-
Long non-coding RNA
- HIF-1a:
-
Hypoxia-inducible factor 1-alpha
References
Asea A, Jean-Pierre C, Kaur P, Rao P, Linhares IM, Skupski D, et al. Heat shock protein-containing exosomes in mid-trimester amniotic fluids. J Reprod Immunol. 2008;79(1):12–7.
Chen J, Li P, Zhang T, Xu Z, Huang X, Wang R, et al. Review on strategies and Technologies for Exosome Isolation and Purification. Front Bioeng Biotechnol. 2022;9:811971.
Jeppesen DK, Fenix AM, Franklin JL, Higginbotham JN, Zhang Q, Zimmerman LJ, et al. Reassessment of exosome composition. Cell. 2019;177(2):428–45 e418.
Logozzi M, Di Raimo R, Mizzoni D, Fais S. What we know on the potential use of exosomes for nanodelivery. In: Seminars in Cancer Biology. Academic Press; 2021.
Mortaz E, Alipoor SD, Varahram M, Jamaati H, Garssen J, Mumby SE, et al. Exosomes in severe asthma: update in their roles and potential in therapy. BioMed Res Int. 2018;2018.
Wu R, Gao W, Yao K, Ge J. Roles of exosomes derived from immune cells in cardiovascular diseases. Front Immunol. 2019;10:648.
Skottvoll FS, Berg HE, Bjørseth K, Lund K, Roos N, Bekhradnia S, et al. Ultracentrifugation versus kit exosome isolation: nanoLC–MS and other tools reveal similar performance biomarkers, but also contaminations. Future science OA. 2018;5(1):FSO359.
Tian H, Li W. Dendritic cell-derived exosomes for cancer immunotherapy: hope and challenges. Ann Transl Med. 2017;5(10).
Wu Z, He D, Li H. Bioglass enhances the production of exosomes and improves their capability of promoting vascularization. Bioact Mater. 2021;6(3):823–35.
Kita S, Shimomura I. Stimulation of exosome biogenesis by adiponectin, a circulating factor secreted from adipocytes. J Biochemistry. 2021;169(2):173–9.
Zhang H-G, Grizzle WE. Exosomes and cancer: a newly described pathway of immune suppression. Clin Cancer Res. 2011;17(5):959–64.
Wulf G. Exosome inhibiting agents and uses thereof. In: Google Patents; 2013.
Théry C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses. Nat Rev Immunol. 2009;9(8):581–93.
Villarroya-Beltri C, Baixauli F, Gutiérrez-Vázquez C, Sánchez-Madrid F, Mittelbrunn M. Sorting it out: regulation of exosome loading. In: Seminars in cancer biology: Elsevier; 2014. p. 3–13.
György B, Szabó TG, Pásztói M, Pál Z, Misják P, Aradi B, et al. Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles. Cell Mol Life Sci. 2011;68(16):2667–88.
Doyle LM, Wang MZ. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells. 2019;8(7):727.
Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9(6):654–9.
Huotari J, Helenius A. Endosome maturation. EMBO J. 2011;30(17):3481–500.
Amini H, Rezabakhsh A, Heidarzadeh M, Hassanpour M, Hashemzadeh S, Ghaderi S, et al. An examination of the putative role of melatonin in exosome biogenesis. Front Cell Dev Biol. 2021;9:686551.
Kenific CM, Zhang H, Lyden D. An exosome pathway without an ESCRT. Cell Res. 2021;31(2):105–6.
Yokoi A, Ochiya T. Exosomes and extracellular vesicles: rethinking the essential values in cancer biology. In: Seminars in Cancer Biology: Elsevier; 2021. p. 79–91.
Zhang Y, Liu Y, Liu H, Tang WH. Exosomes: biogenesis, biologic function and clinical potential. Cell Biosci. 2019;9(1):1–18.
McKelvey KJ, Powell KL, Ashton AW, Morris JM, McCracken SA. Exosomes: mechanisms of uptake. J Circulating Biomarkers. 2015;4:7.
Gurung S, Perocheau D, Touramanidou L, Baruteau J. The exosome journey: from biogenesis to uptake and intracellular signalling. Cell Communication and Signaling. 2021;19(1):1–19.
Arenaccio C, Chiozzini C, Columba-Cabezas S, Manfredi F, Affabris E, Baur A, et al. Exosomes from human immunodeficiency virus type 1 (HIV-1)-infected cells license quiescent CD4+ T lymphocytes to replicate HIV-1 through a Nef-and ADAM17-dependent mechanism. J Virol. 2014;88(19):11529–39.
Viaud S, Terme M, Flament C, Taieb J, André F, Novault S, et al. Dendritic cell-derived exosomes promote natural killer cell activation and proliferation: a role for NKG2D ligands and IL-15Rα. PLoS One. 2009;4(3):e4942.
Théry C, Duban L, Segura E, Véron P, Lantz O, Amigorena S. Indirect activation of naïve CD4+ T cells by dendritic cell–derived exosomes. Nat Immunol. 2002;3(12):1156–62.
Lindenbergh MF, Wubbolts R, Borg EG, van’t Veld EM, Boes M, Stoorvogel W. Dendritic cells release exosomes together with phagocytosed pathogen; potential implications for the role of exosomes in antigen presentation. J Extracell Vesicles. 2020;9(1):1798606.
Lindenbergh MF, Koerhuis DG, Borg EG, van’t Veld EM, Driedonks TA, Wubbolts R, et al. Bystander T-cells support clonal T-cell activation by controlling the release of dendritic cell-derived immune-stimulatory extracellular vesicles. Front Immunol. 2019;10:448.
Kim SH, Bianco N, Menon R, Lechman ER, Shufesky WJ, Morelli AE, et al. Exosomes derived from genetically modified DC expressing FasL are anti-inflammatory and immunosuppressive. Mol Ther. 2006;13(2):289–300.
Miyasaka K, Hanayama R, Tanaka M, Nagata S. Expression of milk fat globule epidermal growth factor 8 in immature dendritic cells for engulfment of apoptotic cells. Eur J Immunol. 2004;34(5):1414–22.
Pitt JM, André F, Amigorena S, Soria J-C, Eggermont A, Kroemer G, et al. Dendritic cell–derived exosomes for cancer therapy. J Clin Invest. 2016;126(4):1224–32.
Silva AM, Almeida MI, Teixeira JH, Maia AF, Calin GA, Barbosa MA, et al. Dendritic cell-derived extracellular vesicles mediate mesenchymal stem/stromal cell recruitment. Sci Rep. 2017;7(1):1–15.
Malekpour K, Hazrati A, Zahar M, Markov A, Zekiy AO, Navashenaq JG, et al. The potential use of mesenchymal stem cells and their derived exosomes for orthopedic diseases treatment. Stem Cell Rev Rep. 2021:1–19. https://doi.org/10.1007/s12015-021-10185-z.
Bianco NR, Kim S-H, Morelli AE, Robbins PD. Modulation of the immune response using dendritic cell-derived exosomes. In: Immunological Tolerance: Springer; 2007. p. 443–55.
Wang G, Jin S, Ling X, Li Y, Hu Y, Zhang Y, et al. Proteomic profiling of LPS-induced macrophage-derived exosomes indicates their involvement in acute liver injury. Proteomics. 2019;19(3):1800274.
Osada-Oka M, Shiota M, Izumi Y, Nishiyama M, Tanaka M, Yamaguchi T, et al. Macrophage-derived exosomes induce inflammatory factors in endothelial cells under hypertensive conditions. Hypertens Res. 2017;40(4):353–60.
Depeille P, Henricks LM, Van De Ven RA, Lemmens E, Wang C-Y, Matli M, et al. RasGRP1 opposes proliferative EGFR–SOS1–Ras signals and restricts intestinal epithelial cell growth. Nat Cell Biol. 2015;17(6):804–15.
Wang C, Zhang C, Liu L, Xi A, Chen B, Li Y, et al. Macrophage-derived mir-155-containing exosomes suppress fibroblast proliferation and promote fibroblast inflammation during cardiac injury. Mol Ther. 2017;25(1):192–204.
McDonald MK, Tian Y, Qureshi RA, Gormley M, Ertel A, Gao R, et al. Functional significance of macrophage-derived exosomes in inflammation and pain. PAIN®. 2014;155(8):1527–39.
Hung Y-Y, Chou C-K, Yang Y-C, Fu H-C, Loh E-W, Kang H-Y. Exosomal let-7e, miR-21-5p, miR-145, miR-146a and miR-155 in predicting antidepressants response in patients with major depressive disorder. Biomedicines. 2021;9(10):1428.
Lan J, Sun L, Xu F, Liu L, Hu F, Song D, et al. M2 macrophage-derived exosomes promote cell migration and invasion in colon cancer. Cancer Res. 2019;79(1):146–58.
Yeung CLA, Co N-N, Tsuruga T, Yeung T-L, Kwan S-Y, Leung CS, et al. Exosomal transfer of stroma-derived miR21 confers paclitaxel resistance in ovarian cancer cells through targeting APAF1. Nat Commun. 2016;7(1):1–14.
Ma Y-S, Wu T-M, Ling C-C, Yu F, Zhang J, Cao P-S, et al. M2 macrophage-derived exosomal microRNA-155-5p promotes the immune escape of colon cancer by downregulating ZC3H12B. Mol Ther Oncolytics. 2021;20:484–98.
Vargas A, Roux-Dalvai F, Droit A, Lavoie J-P. Neutrophil-derived exosomes: a new mechanism contributing to airway smooth muscle remodeling. Am J Respir Cell Mol Biol. 2016;55(3):450–61.
Rossaint J, Kühne K, Skupski J, Van Aken H, Looney MR, Hidalgo A, et al. Directed transport of neutrophil-derived extracellular vesicles enables platelet-mediated innate immune response. Nat Commun. 2016;7(1):1–14.
Jiao Y, Zhang T, Zhang C, Ji H, Tong X, Xia R, et al. Exosomal miR-30d-5p of neutrophils induces M1 macrophage polarization and primes macrophage pyroptosis in sepsis-related acute lung injury. Crit Care. 2021;25(1):1–15.
Skokos D, Botros HG, Demeure C, Morin J, Peronet R, Birkenmeier G, et al. Mast cell-derived exosomes induce phenotypic and functional maturation of dendritic cells and elicit specific immune responses in vivo. J Immunol. 2003;170(6):3037–45.
Li F, Wang Y, Lin L, Wang J, Xiao H, Li J, et al. Mast cell-derived exosomes promote Th2 cell differentiation via OX40L-OX40 ligation. J Immunol Res. 2016;2016.
Al-Nedawi K, Szemraj J, Cierniewski CS. Mast Cell–Derived Exosomes Activate Endothelial Cells to Secrete Plasminogen Activator Inhibitor Type 1. Arterioscler Thromb Vasc Biol. 2005;25(8):1744–9.
Xiao H, Lässer C, Shelke GV, Wang J, Rådinger M, Lunavat TR, et al. Mast cell exosomes promote lung adenocarcinoma cell proliferation–role of KIT-stem cell factor signaling. Cell Commun Signal. 2014;12(1):1–10.
Cañas JA, Sastre B, Mazzeo C, Fernández-Nieto M, Rodrigo-Muñoz JM, González-Guerra A, et al. Exosomes from eosinophils autoregulate and promote eosinophil functions. J Leukoc Biol. 2017;101(5):1191–9.
Cañas J, Sastre B, Rodrigo-Muñoz J, Fernández-Nieto M, Barranco P, Quirce S, et al. Eosinophil-derived exosomes contribute to asthma remodelling by activating structural lung cells. Clin Exp Allergy. 2018;48(9):1173–85.
Cañas JA, Sastre B, Rodrigo-Muñoz JM, Del Pozo V. Exosomes: a new approach to asthma pathology. Clin Chim Acta. 2019;495:139–47.
Mazzeo C, Cañas JA, Zafra MP, Marco AR, Fernández-Nieto M, Sanz V, et al. Exosome secretion by eosinophils: a possible role in asthma pathogenesis. J Allergy Clin Immunol. 2015;135(6):1603–13.
Lugini L, Cecchetti S, Huber V, Luciani F, Macchia G, Spadaro F, et al. Immune surveillance properties of human NK cell-derived exosomes. J Immunol. 2012;189(6):2833–42.
Shoae-Hassani A, Hamidieh AA, Behfar M, Mohseni R, Mortazavi-Tabatabaei SA, Asgharzadeh S. NK cell–derived exosomes from NK cells previously exposed to neuroblastoma cells augment the antitumor activity of cytokine-activated NK cells. J Immunother. 2017;40(7):265–76.
Parolini I, Federici C, Raggi C, Lugini L, Palleschi S, De Milito A, et al. Microenvironmental pH is a key factor for exosome traffic in tumor cells. J Biol Chem. 2009;284(49):34211–22.
Zhu L, Kalimuthu S, Gangadaran P, Oh JM, Lee HW, Baek SH, et al. Exosomes derived from natural killer cells exert therapeutic effect in melanoma. Theranostics. 2017;7(10):2732.
Fais S. NK cell-released exosomes: natural nanobullets against tumors. Oncoimmunology. 2013;2(1):e22337.
Enomoto Y, Li P, Jenkins LM, Anastasakis D, Lyons GC, Hafner M, et al. Cytokine-enhanced cytolytic activity of exosomes from NK Cells. Cancer Gene Ther. 2021:1–16.
Knight AM. Regulated release of B cell-derived exosomes: Do differences in exosome release provide insight into different APC function for B cells and DC? Eur J Immunol. 2008;38(5):1186–9.
Admyre C, Bohle B, Johansson SM, Focke-Tejkl M, Valenta R, Scheynius A, et al. B cell–derived exosomes can present allergen peptides and activate allergen-specific T cells to proliferate and produce TH2-like cytokines. J Allergy Clin Immunol. 2007;120(6):1418–24.
Wubbolts R, Leckie RS, Veenhuizen PT, Schwarzmann G, Möbius W, Hoernschemeyer J, et al. Proteomic and biochemical analyses of human B cell-derived exosomes potential implications for their function and multivesicular body formation. J Biol Chem. 2003;278(13):10963–72.
Clayton A, Turkes A, Dewitt S, Steadman R, Mason MD, Hallett MB. Adhesion and signaling by B cell-derived exosomes: the role of integrins. FASEB J. 2004;18(9):977–9.
Admyre C, Johansson SM, Paulie S, Gabrielsson S. Direct exosome stimulation of peripheral humanT cells detected by ELISPOT. Eur J Immunol. 2006;36(7):1772–81.
Clayton A, Turkes A, Navabi H, Mason MD, Tabi Z. Induction of heat shock proteins in B-cell exosomes. J Cell Sci. 2005;118(16):3631–8.
Ventimiglia LN, Alonso MA. Biogenesis and function of T cell-derived exosomes. Front Cell Dev Biol. 2016;4:84.
Lu J, Wu J, Tian J, Wang S. Role of T cell-derived exosomes in immunoregulation. Immunol Res. 2018;66(3):313–22.
Wahlgren J, Karlson TDL, Glader P, Telemo E, Valadi H. Activated human T cells secrete exosomes that participate in IL-2 mediated immune response signaling. PLoS One. 2012;7(11):e49723.
Mittelbrunn M, Gutiérrez-Vázquez C, Villarroya-Beltri C, González S, Sánchez-Cabo F, González MÁ, et al. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat Commun. 2011;2(1):1–10.
Cai Z, Yang F, Yu L, Yu Z, Jiang L, Wang Q, et al. Activated T cell exosomes promote tumor invasion via Fas signaling pathway. J Immunol. 2012;188(12):5954–61.
Smyth LA, Ratnasothy K, Tsang JY, Boardman D, Warley A, Lechler R, et al. CD73 expression on extracellular vesicles derived from CD4+ CD25+ Foxp3+ T cells contributes to their regulatory function. Eur J Immunol. 2013;43(9):2430–40.
Okoye IS, Coomes SM, Pelly VS, Czieso S, Papayannopoulos V, Tolmachova T, et al. MicroRNA-containing T-regulatory-cell-derived exosomes suppress pathogenic T helper 1 cells. Immunity. 2014;41(1):89–103.
Tung SL, Boardman DA, Sen M, Letizia M, Peng Q, Cianci N, et al. Regulatory T cell-derived extracellular vesicles modify dendritic cell function. Sci Rep. 2018;8(1):1–12.
Wang X, Shen H, He Q, Tian W, Xia A, Lu X-J. Exosomes derived from exhausted CD8+ T cells impaired the anticancer function of normal CD8+ T cells. J Med Genet. 2019;56(1):29–31.
Chen J, Song Y, Miao F, Chen G, Zhu Y, Wu N, et al. PDL1-positive exosomes suppress antitumor immunity by inducing tumor-specific CD8+ T cell exhaustion during metastasis. Cancer Sci. 2021;112(9):3437.
Yao H, Tian L, Yan B, Yang L, Li Y. LncRNA TP73-AS1 promotes nasopharyngeal carcinoma progression through targeting miR-342-3p and M2 polarization via exosomes. Cancer Cell Int. 2022;22(1):1–15.
Obstfeld AE, Frey NV, Mansfield K, Lacey SF, June CH, Porter DL, et al. Cytokine release syndrome associated with chimeric-antigen receptor T-cell therapy: clinicopathological insights. Blood. 2017;130(23):2569–72.
Peters PJ, Borst J, Oorschot V, Fukuda M, Krähenbühl O, Tschopp J, et al. Cytotoxic T lymphocyte granules are secretory lysosomes, containing both perforin and granzymes. J Exp Med. 1991;173(5):1099–109.
Yang T, Martin P, Fogarty B, Brown A, Schurman K, Phipps R, et al. Exosome delivered anticancer drugs across the blood-brain barrier for brain cancer therapy in Danio rerio. Pharm Res. 2015;32(6):2003–14.
Tang X-J, Sun X-Y, Huang K-M, Zhang L, Yang Z-S, Zou D-D, et al. Therapeutic potential of CAR-T cell-derived exosomes: a cell-free modality for targeted cancer therapy. Oncotarget. 2015;6(42):44179.
Jella KK, Nasti TH, Li Z, Malla SR, Buchwald ZS, Khan MK. Exosomes, their biogenesis and role in inter-cellular communication, tumor microenvironment and cancer immunotherapy. Vaccines. 2018;6(4):69.
Li X, Tang M. Exosomes released from M2 macrophages transfer miR-221-3p contributed to EOC progression through targeting CDKN1B. Cancer Med. 2020;9(16):5976–88.
Zhou J, Li X, Wu X, Zhang T, Zhu Q, Wang X, et al. Exosomes released from tumor-associated macrophages transfer miRNAs that induce a Treg/Th17 cell imbalance in epithelial ovarian cancer. Cancer Immunol Res. 2018;6(12):1578–92.
Chan CK, Gulati GS, Sinha R, Tompkins JV, Lopez M, Carter AC, et al. Identification of the human skeletal stem cell. Cell. 2018;175(1):43–56 e21.
Rashid MH, Borin TF, Ara R, Piranlioglu R, Achyut BR, Korkaya H, et al. The critical immunosuppressive effect of MDSC-derived exosomes in the tumor microenvironment. In: bioRxiv; 2020.
Yang M, Chen J, Su F, Yu B, Su F, Lin L, et al. Microvesicles secreted by macrophages shuttle invasion-potentiating microRNAs into breast cancer cells. Mol Cancer. 2011;10(1):117.
Zheng P, Luo Q, Wang W, Li J, Wang T, Wang P, et al. Tumor-associated macrophages-derived exosomes promote the migration of gastric cancer cells by transfer of functional apolipoprotein E. Cell Death Dis. 2018;9(4):1–14.
Si M, Zhu S, Wu H, Lu Z, Wu F, Mo Y. miR-21-mediated tumor growth. Oncogene. 2007;26(19):2799–803.
Chen F, Chen J, Yang L, Liu J, Zhang X, Zhang Y, et al. Extracellular vesicle-packaged HIF-1α-stabilizing lncRNA from tumour-associated macrophages regulates aerobic glycolysis of breast cancer cells. Nat Cell Biol. 2019;21(4):498–510.
DeBerardinis RJ, Chandel NS. Fundamentals of cancer metabolism. Sci Adv. 2016;2(5):e1600200.
Hsu PP, Sabatini DM. Cancer cell metabolism: Warburg and beyond. Cell. 2008;134(5):703–7.
Marino J, Babiker-Mohamed MH, Crosby-Bertorini P, Paster JT, LeGuern C, Germana S, et al. Donor exosomes rather than passenger leukocytes initiate alloreactive T cell responses after transplantation. Sci Immunol. 2016;1(1):aaf8759.
Liu Q, Rojas-Canales DM, Divito SJ, Shufesky WJ, Stolz DB, Erdos G, et al. Donor dendritic cell–derived exosomes promote allograft-targeting immune response. J Clin Invest. 2016;126(8):2805–20.
Mallegol J, Van Niel G, Lebreton C, Lepelletier Y, Candalh C, Dugave C, et al. T84-intestinal epithelial exosomes bear MHC class II/peptide complexes potentiating antigen presentation by dendritic cells. Gastroenterology. 2007;132(5):1866–76.
Hwang B, Bryers J, Mulligan MS. Potential role of exosome-based allorecognition pathways involved in lung transplant rejection: Elsevier; 2020.
Lin J, Wang H, Liu C, Cheng A, Deng Q, Zhu H, et al. Dendritic cells: versatile players in renal transplantation. Front Immunol. 2021;12:1669.
Lakkis FG, Li XC. Innate allorecognition by monocytic cells and its role in graft rejection. Am J Transplant. 2018;18(2):289–92.
Chow KV, Delconte RB, Huntington ND, Tarlinton DM, Sutherland RM, Zhan Y, et al. Innate allorecognition results in rapid accumulation of monocyte-derived dendritic cells. J Immunol. 2016;197(5):2000–8.
Brusselle GG, Koppelman GH. Biologic therapies for severe asthma. N Engl J Med. 2022;386(2):157–71.
Engeroff P, Vogel M. The potential of exosomes in allergy immunotherapy. Vaccines. 2022;10(1):133.
Huang L, Zhang X, Wang M, Chen Z, Yan Y, Gu W, et al. Exosomes from thymic stromal lymphopoietin-activated dendritic cells promote Th2 differentiation through the OX40 ligand. Pathobiology. 2019;86(2-3):111–7.
Lahiri D, Nag M, Dey A, Sarkar T, Pattnaik S, Ghosh S, et al. Exosome-associated host–pathogen interaction: a potential effect of biofilm formation. J Analyt Sci Technol. 2021;12(1):1–8.
Rezaie J, Aslan C, Ahmadi M, Zolbanin NM, Kashanchi F, Jafari R. The versatile role of exosomes in human retroviral infections: from immunopathogenesis to clinical application. Cell Biosci. 2021;11(1):1–15.
Jaworski E, Narayanan A, Van Duyne R, Shabbeer-Meyering S, Iordanskiy S, Saifuddin M, et al. Human T-lymphotropic virus type 1-infected cells secrete exosomes that contain tax protein. J Biol Chem. 2014;289(32):22284–305.
Matsuoka M, Jeang K-T. Human T-cell leukemia virus type 1 (HTLV-1) and leukemic transformation: viral infectivity, Tax, HBZ and therapy. Oncogene. 2011;30(12):1379–89.
Arien KK, Verhasselt B. HIV Nef: role in pathogenesis and viral fitness. Curr HIV Res. 2008;6(3):200–8.
Aqil M, Mallik S, Bandyopadhyay S, Maulik U, Jameel S. Transcriptomic analysis of mRNAs in human monocytic cells expressing the HIV-1 Nef protein and their exosomes. Biomed Res Int. 2015;2015:492395.
Li L, Gu B, Zhou F, Chi J, Wang F, Peng G, et al. Human herpesvirus 6 suppresses T cell proliferation through induction of cell cycle arrest in infected cells in the G2/M phase. J Virol. 2011;85(13):6774–83.
Mori Y, Koike M, Moriishi E, Kawabata A, Tang H, Oyaizu H, et al. Human herpesvirus-6 induces MVB formation, and virus egress occurs by an exosomal release pathway. Traffic. 2008;9(10):1728–42.
Guay C, Kruit JK, Rome S, Menoud V, Mulder NL, Jurdzinski A, et al. Lymphocyte-derived exosomal micrornas promote pancreatic β cell death and may contribute to type 1 diabetes development. Cell Metab. 2019;29(2):348–61 e346.
Zhang Y, Shi L, Mei H, Zhang J, Zhu Y, Han X, et al. Inflamed macrophage microvesicles induce insulin resistance in human adipocytes. Nutr Metab. 2015;12(1):1–14.
Laman JD, Huizinga R, Boons G-J, Jacobs BC. Guillain-Barré syndrome: expanding the concept of molecular mimicry. Trends Immunol. 2022;43(4):296–308.
Du T, Yang C-L, Ge M-R, Liu Y, Zhang P, Li H, et al. M1 macrophage derived exosomes aggravate experimental autoimmune neuritis via modulating Th1 response. Front Immunol. 2020;11:1603.
Azimi M, Ghabaee M, Moghadasi AN, Izad M. Altered expression of miR-326 in T cell-derived exosomes of patients with relapsing-remitting multiple sclerosis. Iran J Allergy Asthma Immunol. 2019;18(1):108–13.
Wu W-C, Song S-J, Zhang Y, Li X. Role of extracellular vesicles in autoimmune pathogenesis. Front Immunol. 2020;11:2310.
Junker A, Krumbholz M, Eisele S, Mohan H, Augstein F, Bittner R, et al. MicroRNA profiling of multiple sclerosis lesions identifies modulators of the regulatory protein CD47. Brain. 2009;132(12):3342–52.
Dai Phung C, Pham TT, Nguyen HT, Nguyen TT, Ou W, Jeong J-H, et al. Anti-CTLA-4 antibody-functionalized dendritic cell-derived exosomes targeting tumor-draining lymph nodes for effective induction of antitumor T-cell responses. Acta Biomater. 2020;115:371–82.
Zhu J, Chen Z, Wang Q, Guo J, Ma L, Shi H, et al. Exosome-derived circITGB1 regulates dendritic cell maturation and cardiac inflammation via miR-342-3p/NFAM1; 2020.
Elashiry M, Elashiry MM, Elsayed R, Rajendran M, Auersvald C, Zeitoun R, et al. Dendritic cell derived exosomes loaded with immunoregulatory cargo reprogram local immune responses and inhibit degenerative bone disease in vivo. J Extracell Vesicles. 2020;9(1):1795362.
Liu S, Chen J, Shi J, Zhou W, Wang L, Fang W, et al. M1-like macrophage-derived exosomes suppress angiogenesis and exacerbate cardiac dysfunction in a myocardial infarction microenvironment. Basic Res Cardiol. 2020;115(2):1–17.
Yan W, Li T, Yin T, Hou Z, Qu K, Wang N, et al. M2 macrophage-derived exosomes promote the c-KIT phenotype of vascular smooth muscle cells during vascular tissue repair after intravascular stent implantation. Theranostics. 2020;10(23):10712.
Li L, Zuo X, Xiao Y, Liu D, Luo H, Zhu H. Neutrophil-derived exosome from systemic sclerosis inhibits the proliferation and migration of endothelial cells. Biochem Biophys Res Commun. 2020;526(2):334–40.
Matsumoto H. Bronchiectasis in severe asthma and asthmatic components in bronchiectasis. Respir Investig. 2021.
Sastre B, Cañas JA, Rodrigo-Muñoz JM, Fernandez-Nieto M, Barranco P, Quirce S, et al. Eosinophil-derived exosomes contribute to asthma remodeling by activating structural lung cells. J Allergy Clin Immunol. 2018;141(2):AB72.
Li D, Wang Y, Jin X, Hu D, Xia C, Xu H, et al. NK cell-derived exosomes carry miR-207 and alleviate depression-like symptoms in mice. J Neuroinflammation. 2020;17:1–19.
Kornbluth J. Treatment of cancer and infectious diseases with natural killer (nk) cell-derived exosomes. In: Google Patents; 2020.
Nisticò N, Maisano D, Iaccino E, Vecchio E, Fiume G, Rotundo S, et al. Role of chronic lymphocytic leukemia (CLL)-derived exosomes in tumor progression and survival. Pharmaceuticals. 2020;13(9):244.
Hu H, Wu J, Cao C, Ma L. Exosomes derived from regulatory T cells ameliorate acute myocardial infarction by promoting macrophage M2 polarization. IUBMB Life. 2020;72(11):2409–19.
Fu W, Lei C, Liu S, Cui Y, Wang C, Qian K, et al. CAR exosomes derived from effector CAR-T cells have potent antitumour effects and low toxicity. Nat Commun. 2019;10(1):1–12.
Srivastava A, Rathore S, Munshi A, Ramesh R. Organically derived exosomes as carriers of anticancer drugs and imaging agents for cancer treatment. In: Seminars in Cancer Biology: Elsevier; 2022.
Markov O, Oshchepkova A, Mironova N. Immunotherapy based on dendritic cell-targeted/−derived extracellular vesicles—a novel strategy for enhancement of the anti-tumor immune response. Front Pharmacol. 2019;10:1152.
Ye Q, Wang B, Mao J. The pathogenesis and treatment of theCytokine Storm'in COVID-19. J Infect. 2020;80(6):607–13.
Hosseini NF, Dalirfardouei R, Aliramaei MR, Najafi R. Stem cells or their exosomes: which is preferred in COVID-19 treatment? Biotechnol Lett. 2022:1–19.
Romagnoli GG, Toniolo PA, Migliori IK, Caldini ÉG, Ferreira MA, Pizzo CR, et al. Tumour cells incorporate exosomes derived from dendritic cells through a mechanism involving the tetraspanin CD9. Exosomes Microvesicles. 2013;1:4.
Romagnoli GG, Zelante BB, Toniolo PA, Migliori IK, Barbuto JAM. Dendritic cell-derived exosomes may be a tool for cancer immunotherapy by converting tumor cells into immunogenic targets. Front Immunol. 2015;5:692.
Choi S-J, Cho H, Yea K, Baek M-C. Immune cell-derived small extracellular vesicles in cancer treatment. BMB Rep. 2022;55(1):48.
Elashiry M, Elsayed R, Cutler CW. Exogenous and endogenous dendritic cell-derived exosomes: lessons learned for immunotherapy and disease pathogenesis. Cells. 2021;11(1):115.
Li J, Huang S, Zhou Z, Lin W, Chen S, Chen M, et al. Exosomes derived from rAAV/AFP-transfected dendritic cells elicit specific T cell-mediated immune responses against hepatocellular carcinoma. Cancer Manag Res. 2018;10:4945.
Lu Z, Zuo B, Jing R, Gao X, Rao Q, Liu Z, et al. Dendritic cell-derived exosomes elicit tumor regression in autochthonous hepatocellular carcinoma mouse models. J Hepatol. 2017;67(4):739–48.
Damo M, Wilson DS, Simeoni E, Hubbell JA. TLR-3 stimulation improves anti-tumor immunity elicited by dendritic cell exosome-based vaccines in a murine model of melanoma. Sci Rep. 2015;5(1):1–15.
Chen S, Ou R, Tang J, Deng X, Wu Y, van Velkinburgh JC, et al. Enhanced anti-tumor effects of HPV16E749–57-based vaccine by combined immunization with poly (I: C) and oxygen-regulated protein 150. Cancer Epidemiol. 2013;37(2):172–8.
Kim HP, Morse D, Choi AM. Heat-shock proteins: new keys to the development of cytoprotective therapies. Expert Opin Ther Targets. 2006;10(5):759–69.
Tamura Y, Torigoe T, Kukita K, Saito K, Okuya K, Kutomi G, et al. Heat-shock proteins as endogenous ligands building a bridge between innate and adaptive immunity. Immunotherapy. 2012;4(8):841–52.
Wang J, Wang Z, Mo Y, Zeng Z, Wei P, Li T. Effect of hyperthermic CO2-treated dendritic cell-derived exosomes on the human gastric cancer AGS cell line. Oncol Lett. 2015;10(1):71–6.
Zhang L, Hao C, Zhai R, Wang D, Zhang J, Bao L, et al. Downregulation of exosomal let-7a-5p in dust exposed-workers contributes to lung cancer development. Respir Res. 2018;19(1):1–8.
Zhang L, Duan S, Yu S, Yuan T, Yao S. Exogenous let-7a-5p induces A549 lung cancer cell death through BCL2L1-mediated PI3Kγ signaling pathway. Front Oncol. 2019;9:808.
Pace ALD, Tumino N, Besi F, Alicata C, Conti LA, Munari E, et al. Characterization of human NK cell-derived exosomes: role of DNAM1 receptor in exosome-mediated cytotoxicity against tumor. Cancers. 2020;12(3):661.
Lettau M, Janssen O. Intra-and extracellular effector vesicles from human T and NK cells: same-same, but different? Front Immunol. 2021;12:804895.
Saini RV, Wilson C, Finn MW, Wang T, Krensky AM, Clayberger C. Granulysin delivered by cytotoxic cells damages endoplasmic reticulum and activates caspase-7 in target cells. J Immunol. 2011;186(6):3497–504.
Jong AY, Wu C-H, Li J, Sun J, Fabbri M, Wayne AS, et al. Large-scale isolation and cytotoxicity of extracellular vesicles derived from activated human natural killer cells. J Extracell Vesicles. 2017;6(1):1294368.
Guo T, Zhao L, Zhang Y, Liu G, Yao Y, Li H. A monoclonal antibody targeting the dimer interface of epidermal growth factor receptor (EGFR). Immunol Lett. 2016;180:39–45.
Qiu L, Zhou C, Sun Y, Di W, Scheffler E, Healey S, et al. Crosstalk between EGFR and TrkB enhances ovarian cancer cell migration and proliferation. Int J Oncol. 2006;29(4):1003–11.
Maecker H, Varfolomeev E, Kischkel F, Lawrence D, LeBlanc H, Lee W, et al. TWEAK attenuates the transition from innate to adaptive immunity. Cell. 2005;123(5):931–44.
Gu L, Dai L, Cao C, Zhu J, Ding C, Xu HB, et al. Functional expression of TWEAK and the receptor Fn14 in human malignant ovarian tumors: possible implication for ovarian tumor intervention. PLoS One. 2013;8(3):e57436.
Hu Y, Li D, Wu A, Qiu X, Di W, Huang L, et al. TWEAK-stimulated macrophages inhibit metastasis of epithelial ovarian cancer via exosomal shuttling of microRNA. Cancer Lett. 2017;393:60–7.
Ochs K, Sahm F, Opitz CA, Lanz TV, Oezen I, Couraud P-O, et al. Immature mesenchymal stem cell-like pericytes as mediators of immunosuppression in human malignant glioma. J Neuroimmunol. 2013;265(1-2):106–16.
Seo N, Shirakura Y, Tahara Y, Momose F, Harada N, Ikeda H, et al. Activated CD8+ T cell extracellular vesicles prevent tumour progression by targeting of lesional mesenchymal cells. Nat Commun. 2018;9(1):1–11.
Guo W, Li Y, Pang W, Shen H. Exosomes: a potential therapeutic tool targeting communications between tumor cells and macrophages. Mol Ther. 2020;28(9):1953–64.
Jiang H, Zhou L, Shen N, Ning X, Wu D, Jiang K, et al. M1 macrophage-derived exosomes and their key molecule lncRNA HOTTIP suppress head and neck squamous cell carcinoma progression by upregulating the TLR5/NF-κB pathway. Cell Death Dis. 2022;13(2):1–15.
Neviani P, Wise PM, Murtadha M, Liu CW, Wu C-H, Jong AY, et al. Natural killer–derived exosomal miR-186 inhibits neuroblastoma growth and immune escape mechanisms. Cancer Res. 2019;79(6):1151–64.
Zhang X, Liu L, Tang M, Li H, Guo X, Yang X. The effects of umbilical cord-derived macrophage exosomes loaded with cisplatin on the growth and drug resistance of ovarian cancer cells. Drug Dev Ind Pharm. 2020;46(7):1150–62.
Dugas JC, Cuellar TL, Scholze A, Ason B, Ibrahim A, Emery B, et al. Dicer1 and miR-219 are required for normal oligodendrocyte differentiation and myelination. Neuron. 2010;65(5):597–611.
Lau P, Verrier JD, Nielsen JA, Johnson KR, Notterpek L, Hudson LD. Identification of dynamically regulated microRNA and mRNA networks in developing oligodendrocytes. J Neurosci. 2008;28(45):11720–30.
Budde H, Schmitt S, Fitzner D, Opitz L, Salinas-Riester G, Simons M. Control of oligodendroglial cell number by the miR-17-92 cluster. Development. 2010;137(13):2127–32.
Pusic AD, Pusic KM, Clayton BL, Kraig RP. IFNγ-stimulated dendritic cell exosomes as a potential therapeutic for remyelination. J Neuroimmunol. 2014;266(1-2):12–23.
Yu L, Yang F, Jiang L, Chen Y, Wang K, Xu F, et al. Exosomes with membrane-associated TGF-β1 from gene-modified dendritic cells inhibit murine EAE independently of MHC restriction. Eur J Immunol. 2013;43(9):2461–72.
Azimi M, Ghabaee M, Moghadasi AN, Noorbakhsh F, Izad M. Immunomodulatory function of Treg-derived exosomes is impaired in patients with relapsing-remitting multiple sclerosis. Immunol Res. 2018;66(4):513–20.
Bu N, Wu H-Q, Zhang G-L, Zhan S-Q, Zhang R, Fan Q-Y, et al. Immature dendritic cell exosomes suppress experimental autoimmune myasthenia gravis. J Neuroimmunol. 2015;285:71–5.
Yin W, Ouyang S, Luo Z, Zeng Q, Hu B, Xu L, et al. Immature exosomes derived from microRNA-146a overexpressing dendritic cells act as antigen-specific therapy for myasthenia gravis. Inflammation. 2017;40(4):1460–73.
Zöller M. Janus-faced myeloid-derived suppressor cell exosomes for the good and the bad in cancer and autoimmune disease. Front Immunol. 2018;9:137.
Wang Y, Tian J, Tang X, Rui K, Tian X, Ma J, et al. Exosomes released by granulocytic myeloid-derived suppressor cells attenuate DSS-induced colitis in mice. Oncotarget. 2016;7(13):15356.
Kim SH, Bianco NR, Shufesky WJ, Morelli AE, Robbins PD. Effective treatment of inflammatory disease models with exosomes derived from dendritic cells genetically modified to express IL-4. J Immunol. 2007;179(4):2242–9.
Blois S, Tometten M, Kandil J, Hagen E, Klapp BF, Margni RA, et al. Intercellular adhesion molecule-1/LFA-1 cross talk is a proximate mediator capable of disrupting immune integration and tolerance mechanism at the feto-maternal interface in murine pregnancies. J Immunol. 2005;174(4):1820–9.
Bianco NR, Kim SH, Ruffner MA, Robbins PD. Therapeutic effect of exosomes from indoleamine 2, 3-dioxygenase–positive dendritic cells in collagen-induced arthritis and delayed-type hypersensitivity disease models. Arthritis Rheum. 2009;60(2):380–9.
Ono Y, Perez-Gutierrez A, Nakao T, Dai H, Camirand G, Yoshida O, et al. Graft-infiltrating PD-L1hi cross-dressed dendritic cells regulate antidonor T cell responses in mouse liver transplant tolerance. Hepatology. 2018;67(4):1499–515.
Yu X, Huang C, Song B, Xiao Y, Fang M, Feng J, et al. CD4+ CD25+ regulatory T cells-derived exosomes prolonged kidney allograft survival in a rat model. Cell Immunol. 2013;285(1-2):62–8.
Agarwal A, Fanelli G, Letizia M, Tung SL, Boardman D, Lechler R, et al. Regulatory T cell-derived exosomes: possible therapeutic and diagnostic tools in transplantation. Front Immunol. 2014;5:555.
Bandyopadhyay S, Friedman RC, Marquez RT, Keck K, Kong B, Icardi MS, et al. Hepatitis C virus infection and hepatic stellate cell activation downregulate miR-29: miR-29 overexpression reduces hepatitis C viral abundance in culture. J Infect Dis. 2011;203(12):1753–62.
Segura E, Guérin C, Hogg N, Amigorena S, Théry C. CD8+ dendritic cells use LFA-1 to capture MHC-peptide complexes from exosomes in vivo. J Immunol. 2007;179(3):1489–96.
Sobo-Vujanovic A, Munich S, Vujanovic NL. Dendritic-cell exosomes cross-present Toll-like receptor-ligands and activate bystander dendritic cells. Cell Immunol. 2014;289(1-2):119–27.
Yao Y, Fu C, Zhou L, Mi Q-S, Jiang A. DC-derived exosomes for cancer immunotherapy. Cancers. 2021;13(15):3667.
Smith VL, Cheng Y, Bryant BR, Schorey JS. Exosomes function in antigen presentation during an in vivo mycobacterium tuberculosis infection. Sci Rep. 2017;7:43578.
Wang J, Deng Z, Wang Z, Wu J, Gu T, Jiang Y, et al. MicroRNA-155 in exosomes secreted from helicobacter pylori infection macrophages immunomodulates inflammatory response. Am J Transl Res. 2016;8(9):3700.
Rodrigues M, Fan J, Lyon C, Wan M, Hu Y. Role of extracellular vesicles in viral and bacterial infections: pathogenesis, diagnostics, and therapeutics. Theranostics. 2018;8(10):2709.
Lecce M, Molfetta R, Milito ND, Santoni A, Paolini R. FcεRI signaling in the modulation of allergic response: role of mast cell-derived exosomes. Int J Mol Sci. 2020;21(15):5464.
Xie G, Yang H, Peng X, Lin L, Wang J, Lin K, et al. Mast cell exosomes can suppress allergic reactions by binding to IgE. J Allergy Clin Immunol. 2018;141(2):788–91.
Li C, Deng C, Zhou T, Hu J, Dai B, Yi F, et al. MicroRNA-370 carried by M2 macrophage-derived exosomes alleviates asthma progression through inhibiting the FGF1/MAPK/STAT1 axis. Int J Biol Sci. 2021;17(7):1795.
Li C, Dai B, Hu J, Shang Y. M2 macrophage-derived exosomes carry microRNA-370 to alleviate asthma progression through inhibiting the FGF1/MAPK/STAT1 axis. Exp Cell Res. 2020:112285.
Yamashita T, Takahashi Y, Takakura Y. Possibility of exosome-based therapeutics and challenges in production of exosomes eligible for therapeutic application. Biol Pharm Bull. 2018;41(6):835–42.
Jeyaram A, Jay SM. Preservation and storage stability of extracellular vesicles for therapeutic applications. AAPS J. 2018;20(1):1.
Lim SK. Method for lyophilising an exosome. In: Google Patents; 2019.
Veerman RE, Akpinar GG, Eldh M, Gabrielsson S. Immune cell-derived extracellular vesicles–functions and therapeutic applications. Trends Mol Med. 2019;25(5):382–94.
Peng H, Ji W, Zhao R, Yang J, Lu Z, Li Y, et al. Exosome: a significant nano-scale drug delivery carrier. J Mater Chem B. 2020;8(34):7591–608.
Tian Y, Li S, Song J, Ji T, Zhu M, Anderson GJ, et al. A doxorubicin delivery platform using engineered natural membrane vesicle exosomes for targeted tumor therapy. Biomaterials. 2014;35(7):2383–90.
Acknowledgments
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
AH and KM wrote the manuscript, M M and A R prepared the figure, and the tables, S S and S M H supervised the study and correspondence during the paper preparation and submission, and RSV read and edited the final submission. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Hazrati, A., Soudi, S., Malekpour, K. et al. Immune cells-derived exosomes function as a double-edged sword: role in disease progression and their therapeutic applications. Biomark Res 10, 30 (2022). https://doi.org/10.1186/s40364-022-00374-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40364-022-00374-4