
Abstract
Metastasis is still poorly understood and thus further research must be conducted to provide insight into the driving factors. Novel research has revealed the significance of the microenvironment in the delegation of metastasis, expanding the field of cancer metastasis to cells and cell environments surrounding the migrated tumor cells. Research on hepatic metastasis is an ever-growing domain of this field, as several primary tumors can metastasize to the liver. The two features within the liver that promote metastasis—cellular and acellular—are found in the current interpretation of liver microenvironment. Novel findings of both are included in this review. Different hypotheses detailing the methods by which metastasis can occur must be included to understand the significance of the microenvironment, as well as a brief overview of the methods that can be used during research. This review aims to highlight the importance of liver microenvironment on the development or potential regression of hepatic metastasis through discussing both acellular and cellular components of liver microenvironment and their interaction with metastasis.
Similar content being viewed by others


Introduction
Organ microenvironments are an undeniable component for metastasis, enabling proliferation of cancer cells to organ systems external to the primary tumor. Migration of cancer cells requires compatibility with the destination, particularly with the microenvironment. Hepatic metastases can arise from the primary cancers in different locations within the human body. Both the acellular-such as extracellular matrix (ECM) proteins (i.e. collagen) and the cellular components of the liver such as Kupffer cells (KCs), hepatic stellate cells (HSCs), and liver sinusoidal endothelial cells (LSECs) contribute to the metastatic ability of tumors of different origins. Common incidences of liver metastasis occur in colorectal cancers, as these cells can take advantage of both the proximity (as a great amount of venous drainage is to the liver) and highly vascular nature of the liver [1]. A statistical analysis revealed that of 4399 patients with cancer, 41% of them experienced metastasis to the liver, marking it as the second greatest metastatic site, just behind nonspecific lymph node metastases [2].
The ECM is the major acellular component contributing to the liver microenvironment, and consequently the microenvironment of the tumor. The matrix of biological tissue serves as a framework by which cells organize, intertwined with the local vasculature. The ECM also contains domains that allow proteins to bind, which are important for cell–cell communication as well as specific function. ECM proteins can interact with growth factors, such as hepatocyte growth factor (HGF) which promotes cell migration and vascular endothelial growth factor (VEGF) which can enable metastasis progression through angiogenesis [3]. The major cellular components, such as Kupffer cells, liver sinusoidal endothelial cells, hepatocytes, etc. are also involved, primarily through communication with matrix and intracellular proteins, to promote metastasis.
Though the liver microenvironment can promote metastasis, the liver will initially respond in ways to react to inflammation or damage. This premetastatic niche can protect the liver, using cellular signaling to promote immune responses. Both cellular and acellular provide these dueling roles, such as KCs and cytokines, though eventually the prometastatic elements outweigh the opposition as metastasis occurs [4]. Table 1 summarizes major cellular and acellular components play role in hepatic metastasis identified in previous studies. The initial microenvironment can play opposing roles in the development of metastases, but once tumor growth occurs, that environment is subject to changes that ultimately support that growth.
Acellular components
The major acellular components that support or are involved in metastatic niche formation are as follows; the cell-adhesion molecule carcinoembryonic antigen (CEA) and other cell adhesion molecules (CAMs), CXC motif-chemokines (CXCLs), VEGF, MAPK, and NF-κB, Citrullinated proteins/PAD, Spermine pullulan, matrix metalloproteinases, and collagen proteins (Table 1).
Carcinoembryonic antigen
Abdul-Wahid et al. reported that the CEA, which can be expressed on the surface of colon circulating tumor cells (CTCs), contributes to the subsequent attachment to fibronectin of the liver ECM, resulting in increased levels of metastasis [5]. This is supported in the review by Rizeq et al., which describes in further detail of human CEA and other cell adhesion molecules, showing its role in cancer progression [6]. It reiterates the ability of these molecules to increase the binding of fibronectin in the ECM to cancer cells, as well as participating in more direct proliferative measures such as apoptosis, all of which mark the beginning of metastasis [6].
CXC motif-chemokines
It is known that CXC motif-chemokines (CXCL) 1, 2, 3, and 5 contribute to colorectal carcinoma, but novel exploration expanded this understanding to metastasis to the liver. Knock-down of Interleukin-8 (IL-8, or CXCL8), which can be produced by macrophages, was shown to inhibit tumor growth in colorectal liver metastasis. This was associated with the reduction in cell proliferation and viability in samples deficient in CXCL8 [7].
VEGF, MAPK, and NF-κB
VEGF expression levels were also decreased during knock-down of CXCL8, suggesting a mechanism for the decreased proliferation [8]. VEGF, along with MAP Kinase (MAPK) and NF-κB, were reported to contribute to liver metastasis of breast cancer [9]. A study identified different genes associated with patients that developed liver metastases from a primary breast tumor, and found that these signaling pathways were highly conserved among patients, since VEGF promotes angiogenesis (required for tumor growth and expansion), MAPK promotes epithelial-to-mesenchymal transition (EMT) associated with TGF-β, and NF-κB is a regulatory transcription factor for the immune response and is associated with EMT as well [10].
Citrullinated proteins and PAD
Recently, ECM proteins were shown to have a greater incidence of citrullinated proteins, catalyzed by peptidylarginine deiminase proteins (PAD), which is specifically seen in liver metastases. Citrullination occurs on the arginine, producing a neutrally charged product. It was suggested in earlier studies that citrullination of certain proteins, which are prevalent and detectable in tumor cells, contribute to the progression of the tumor [11]. A more recent study focused on PAD4 and colorectal cancer metastasis to the liver, showing that the downregulation of PAD4 reduced metastatic growth, suggesting the possibility of citrullination as a contributor to metastasis [11].
Spermine pullulan
Polysaccharide spermine modified pullulan (SP), which is produced by the fungus Aureobasidium pullulans, can be utilized to polarize macrophages towards M1, which can ultimately inhibit metastasis by increasing inflammation [12].
Matrix metalloproteinases
In an experiment involving the treatment of murine models with chemotherapy, periostin and matrix-metalloproteinase 2 (MMP-2) were increased when murine models were treated with the drug cisplatin. Therefore, they concluded that cisplatin may induce liver metastasis of murine melanoma cells via increase in MMPs, which in turn can manipulate the extracellular environment by protein turnover [13]. The study also looked at how treatment of the chemotherapy drug vincristine could aid in the metastasis of human neuroblastoma to liver, via an increase in MMP-9 expression [13]. The results of this study suggest that the microenvironment interactions with chemotherapy can not only increase understanding of metastasis, but provide valuable information regarding treatment expectations.
Collagen proteins
Observational data show colorectal liver metastasis (CRLM) patients have elevated levels of type I collagen in the urine and plasma [14]. This report prompted further exploration of other ECM collagens [14]. Elevated levels for CRLM patients compared to controls were analyzed with a P value < 0.0001. Collagen peptides and alpha chains were identified, and the majority were upregulated in CRLM patients. This study underwent extensive data analysis, finding possible markers as well as drivers for metastasis. It was additionally hypothesized that abnormal ECM protein synthesis and degradation occurred, as it was determined that collagen turnover related proteins were upregulated in CRLM patients as well [14]. Data analysis on breast cancer metastasis determined that the addition of type I collagen altered the metastatic properties of tumor epithelia, allowing for a greater incidence of lung metastasis as well as an increase in the number of CTCs [13]. Though different mechanisms are involved in metastasis to the lung as opposed to metastasis to the liver, this provides an alternate view, accentuating the significance of collagen type I. Collagen type I was reviewed as a possible instigator of metastasis in these particular studies, and other experiments looked at metastasis occurring with other collagen types. A study on collagen IV and liver metastasis revealed the significance of type IV collagen expression, showing how upregulation of collagen IV is a possible driver of metastasis, and that downregulating this ECM protein reduces metastasis [15]. These articles collectively address the importance of matrix components in relation to the metastatic capacity of the tumor.
Cellular components
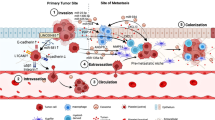
Cellular components of the liver are as follows: bone-marrow-derived macrophages such as tumor-associated macrophages (TAMs), Kupffer cells (KCs), liver sinusoidal endothelial cells (LSECs), immune cells like tumor-associated neutrophils (TANs), myeloid-derived suppressor cells (MDSC), fibroblasts, hepatocytes, and hepatic stellate cells (HSCs) [4] (Fig. 1, Table 1).

Cellular factors playing role in formation and development of metastatic niche within liver microenvironment. This figure depicts the schematic representation of the major cellular components involved in a multistep process in the formation and development of the metastatic niches in the liver microenvironment. (TEM transendothelial migration, MMP metalloproteinases, CCL5 chemokine ligand 5)
Macrophages and Kupffer cells
Macrophages can either be resident like KCs, or monocyte-derived, both of which can undergo M1 to M2 repolarization [16]. This transition circumvents the pro-immunogenic response and anti-tumorigenic qualities of these immune cells, assisting tumor growth. TAMs which take on the M2 phenotype, will prevent immunogenic and inflammatory responses and enhance cancer progression [12]. In addition, M2 phenotype can combat chemotherapy, thus preventing macrophage M2 polarization (thereby inducing M1 polarization) is a plausible area for treatment. Both cell types can contribute to metastasis [4], however some studies have suggested a larger role for KCs in their prevention of metastasis. A study by Kimura et al. reports lectin receptors (Dectins 1–3) can suppress tumor growth by promoting KCs and natural killer cells (NK cells) [17]. KCs are able to promote a viable environment for cancer cells, as they can indirectly promote the production of fibronectin [18]. However, KCs are a double edged sword, known for anti-metastatic properties as well [18]. Since the relationship of KCs to cancer progression is complex, the downstream effects of these cells needs to be further identified to enable probable therapeutic outcomes.
Sinusoidal endothelial cells
Liver sinusoidal endothelial cells (LSECs) can provide both anti-metastatic and pro-metastatic functions. For example, LSECs can undergo apoptosis or release toxins to aid in elimination of tumors at the site of metastasis, or these cells can increase angiogenesis or EMT to promote metastasis [4].
Immune cells
Different immune cells, such as dendritic cells, tumor-associated neutrophils (TAN), myeloid-derived suppressor cells (MDSC), and TAMs are all associated with different cytokines/chemokines or other factors that contribute to metastasis, summarized in Table 1 of the review by Smith and Kang [19]. Many of these factors are acellular components, though all acellular components must be considered within the context of the cellular origin or target. This is particularly important when discussing the role of immune cells, which frequently must intercommunicate with cytokines/chemokines.
Fibroblasts
As mentioned, chemokines can contribute to metastasis by immune system suppression. The behavior of macrophages and other immune cells towards cancer is dependent on these cell-signal molecules. SDF-1 (CXCL12), or stromal cell derived factor, is known to allow for TAM movement, and can be secreted directly from cancer cells [20]. SDF-1 production is the result of activation of HSCs, which are thought to be progenitors to α-smooth muscle actin (α-SMA)-positive myofibroblasts [21] This phenotype is usually a marker for cancer-associated fibroblasts, which are tumor-activated forms of fibroblasts that secrete matrix proteins such as collagen, and can enhance the metastatic properties of the tumor [22]. The interaction between SDF-1 and CXCR4 can be interrupted as well, and targeting CXCR4 has shown to reduce metastasis, as SDF-1/CXCR4 expression and binding induces migration of cancer cells [22, 23]. Treatment with a CXCR4 antagonist was able to reduce metastasis in endometrial cancer (induced by cancer-associated fibroblasts) [22]. A recent study showed that the addition of a CXCR4 antagonist also disturbs metastasis of colorectal cancer to the liver, and connected certain HSC-derived factors to the increased expression of CXCR4 [24]. Hypoxia-inducing factor-1(HIF-1), can stimulate the transition of macrophages from M1 to M2 [20]. The conversion back to M1 can be induced through signaling molecules such as tumor necrosis factor alpha (TNF-α) and lipopolysaccharide (LPS) [4], as well as SP (earlier stated); however, this could prolong the inflammatory response systemically. Insulin-like growth factor 1 (IGF-1) has been shown to induce metastasis and the signaling pathways involved in the promotion of EMT. Therefore, depletion of IGF-1 and prevented polarization of TAN to their pro-metastatic (N2) phenotype [25, 26].
Cancer stem cells and tumor microenvironment
Cancer stem cells (CSC) are thought to be a portion of the population of a tumor, and can be differentiated based on their surroundings. The tumor microenvironment is crucial to this differentiation. CSCs can interact with both acellular and cellular components of the microenvironment. The review by Lau et al. describes the connections between CSC pathways and ECM changes, hypoxia, growth factors, etc., which all contribute to metastasis [27]. The development of CSCs and their role in metastasis needs to be studied further, though many hypotheses have been developed to explain this relationship. One particular hypothesis in regards to the relationship between CSCs and the EMT, which is currently not completely understood, involves the requirement of the microenvironment to provide signals and structure to which the CSCs can react [28, 29]. A specific example involves the migration of pancreatic adenocarcinoma to the liver, in which Knaack et al. studied in vitro, manipulating CSCs as well as the microenvironment by incorporating HSCs and hepatic myofibroblasts (HMF) to mimic physiological and inflammatory conditions. The data expressed the importance of HSCs and HMF in the formation of disseminated pancreatic ductal epithelial cell (PDEC) colonies, showing a marked relationship between liver microenvironment and pancreatic CTCs [30]. In addition, these colonies expressed a greater amount of Nestin, a CSC-marker, than their counterparts, which reveals the increased metastatic capabilities of these cells [30].
The role of exosome in establishing liver metastasis
Recent studies have shown that pancreatic ductal adenocarcinoma (PDAC)-derived exosomes stimulate liver pre-metastatic niche formation (PMN). Costa-Vista et al. showed that uptake of PDAC-derived exosomes by Kupffer cells caused TGF-β secretion and upregulation of fibronectin production by HSCs [31]. They found that macrophage migration inhibitory factor (MIF) was highly expressed in PDAC-derived exosomes, and its blockade impeded liver pre-metastatic niche formation and metastasis [31]. This group also showed that exosomes from pancreatic cancer cell lines that metastasize selectively to the liver fused preferentially with KCs in the liver. Interestingly, Costa-Silva et al. found that exosomal MIF upregulation in mice with pretumoral pancreatic lesions, and high levels of exosomal MIF in plasma were also identified in patients with stage I PDAC. These results suggest that these exosomes could be formed at very early stages of cancer development [31].
Epithelial-to-mesenchymal transition and circulating tumor cells
It is assumed that EMT is a primary method of metastasis, as the transition to mesenchymal cells allows for migration to the bloodstream, which acts as a highway for tumor cells. The reverse, mesenchymal-to-epithelial transition (MET), can halt tumor cells at a certain location. Individual cells detected in patient serum are known as circulating or disseminated tumor cells (DTCs), and it is thought that EMT plays a role in tumor cell dissemination [32]. The path of a CTC following EMT for liver metastasis is the infiltration of that cell into the sinusoids and ultimately extravasation, assuming survival of the CTC [33]. Monitoring CTCs in serum is difficult, as there are approximately 5 cells per 10 mL of blood [32]. However, discussion of CTCs in the formulation of a mechanism for cancer metastasis is still important. Interestingly, CTCs can differ from the primary tumor genetically, which increase the difficulty in detection, as DNA analysis can be used as a method of characterization [32]. Without proper characterization, the interaction between CTCs and the liver microenvironment is concealed.
Methods for studying microenvironment and metastasis
The tumor and organ microenvironment is not as easily manipulated in laboratory settings; in vitro is not primarily involved in the components surrounding individual cells. This barrier prompted the exploration into new methods of visualization and manipulation. Both two-dimensional (2D) and three-dimensional (3D) models can be utilized, and have their respective advantages and disadvantages [34]. In vitro 2D models are relatively limited in the study of metastasis, especially in the context of the surrounding microenvironment, thus 3D models are typically used. However, in vivo models are expensive and inefficient, thus prompting exploration of different methods of cancer metastasis visualization that allow for environmental control. Organoids, aptly named to indicate its three-dimensional nature, are clusters of cells that represent a fraction of a particular tissue environment [35]. Use of organoids to represent hepatic tissue contain a greater representation of the liver proteins and genetic components than two-dimensional structures, and can serve as an advantageous method to study liver microenvironment [36]. Organoids were generated in a study of colorectal cancer liver metastasis, and this supported the use of organoids to study metastasis as certain biomarkers were retained, supplemental growth was established, and drug screening was successful [37]. Decellularization of tissue is a relatively new method of in vitro research, and can be used to study how the microenvironment of a particular organ interacts with different cell lines [38]. The decellularized tissue, known colloquially as biomatrix, can be pulverized and plated, which increases trial efficiency, or the structure of the organ can be retained, keeping this aspect of the microenvironment constant during experimentation [39]. In a study was done by Tian et al. in our lab, an ex vivo engineered metastatic model was developed on lung and liver biomatrix which can be used as new in vivo model for tissue-specific metastasis [40]. The review by Clark et al. discusses ex vivo systems, which can mimic in vivo and avoid certain limitations of murine models [33].
Conclusions
Ultimately, the microenvironment of the liver is crucial to the development of hepatic metastases. Different growth factors, cell types, posttranslational modifications, signaling molecules, etc. are all necessary to understand, including their interactions, when deciphering the elements of metastatic outgrowth. The accumulation of observational and experimental studies contribute to the knowledge required to battle cancer metastasis, and the methods to understand this metastasis in laboratory settings continues to progress.
Abbreviations
- ECM:
-
extracellular matrix
- KCs:
-
Kupffer cells
- HSCs:
-
hepatic stellate cells
- LSEC:
-
liver sinusoidal endothelial cells
- HGF:
-
hepatocyte growth factor
- VEGF:
-
vascular endothelial growth factor
- CEA:
-
carcinoembryonic antigen
- CTC:
-
circulating tumor cells
- CEACAM6:
-
carcinoembryonic antigen-related cell adhesion molecule 6
- CXCL:
-
CXC ligand
- MAPK:
-
mitogen-activated protein kinase
- NF-κB:
-
nuclear factor kappa-light-chain-enhancer of activated B cells
- TGF-β:
-
transforming growth factor beta
- EMT:
-
epithelial-to-mesenchymal transition
- PAD:
-
peptidylarginine deaminase
- SP:
-
spermine pullulan
- MMP:
-
matrix metalloproteinase
- CRLM:
-
colorectal liver metastasis
- TAM:
-
tumor-associated macrophage
- NK cells:
-
natural killer cells
- TAN:
-
tumor-associated neutrophils
- MDSC:
-
myeloid-derived suppressor cells
- SDF:
-
stromal cell-derived factor
- HSC:
-
hepatic stellate cell
- HIF-1:
-
hypoxia-inducing factor-1
- TNF-α:
-
tumor necrosis factor alpha
- IGF-1:
-
insulin-like growth factor -1
- CSC:
-
cancer stem cells
- HMF:
-
hepatic myofibroblasts
- PDAC:
-
pancreatic ductal adenocarcinoma
- PDEC:
-
pancreatic ductal epithelial cell
- MET:
-
mesenchymal-to-epithelial transition
- DTC:
-
disseminated tumor cells
- 2D:
-
two-dimensional
- 3D:
-
three-dimensional
References
Sheth KR, Clary BM (2005) Management of hepatic metastases from colorectal cancer. Clin Colon Rectal Surg 18(3):215–223
Hess KR, Varadhachary GR, Taylor SH, Wei W, Raber MN, Lenzi R, Abbruzzese JL (2006) Metastatic patterns in adenocarcinoma. Cancer 106:1624–1633
Arriazu E, Ruiz de Galarreta M, Cubero FJ, Varela-Rey M, Pérez de Obanos MP, Leung TM, Lopategi A, Benedicto A, Abraham-Enachescu I et al (2014) Extracellular matrix and liver disease. Antioxid Redox Signal 21(7):1078–1097
Brodt P (2016) Role of the microenvironment in liver metastasis: from pre- to prometastatic niches. Clin Cancer Res 22:5971–5982
Abdul-Wahid A, Cydzik M, Fischer NW, Prodeus A, Shively JE, Martel A, Alminawi S, Ghorab Z, Berinstein NL, Gariépy J (2018) Serum-derived carcinoembryonic antigen (CEA) activates fibroblasts to induce a local re-modeling of the extracellular matrix that favors the engraftment of CEA-expressing tumor cells. Int J Cancer 143:1963–1977
Rizeq B, Zakaria Z, Ouhtit A (2017) Towards understanding the mechanisms of actions of carcinoembryonic antigen-related cell adhesion molecule 6 in cancer progression. Cancer Sci 109(1):33–42
Sharma I, Singh A, Siraj F, Saxena S (2018) IL-8/CXCR7/2 signalling promotes tumor cell proliferation, invasion and vascular mimicry in glioblastoma. J Biomed Sci 25(1):62
Kumar A, Cherukumilli M, Mahmoudpour SH, Brand K, Bandapalli OR (2018) ShRNA-mediated knock-down of CXCL8 inhibits tumor growth in colorectal liver metastasis. Biochem Biophys Res Commun 500(3):731–737
Chen X, Zheng Z, Chen L, Zheng H (2017) MAPK, NFκB, and VEGF signaling pathways regulate breast cancer liver metastasis. Oncotarget 8(60):101452–101460
Jiang Z, Cui Y, Wang L, Zhao Y, Yan S, Chang X (2013) Investigating citrullinated proteins in tumour cell lines. World J Surg Oncol 11:260
Yuzhalin AE, Gordon-Weeks AN, Tognoli ML, Jones K, Markelc B, Konietzny R, Fischer R, Muth A, O’Neill E, Thompson PR, Venables PJ, Kessler BM, Lim SY et al (2018) Colorectal cancer liver metastatic growth depends on PAD4-driven citrullination of the extracellular matrix. Nat Commun 9(1):4783
Xie L, Yang Y, Meng J, Wen T, Liu J, Xu H (2019) Cationic polysaccharide spermine-pullulan drives tumor associated macrophage towards M1 phenotype to inhibit tumor progression. Int J Biol Macromol 123:1012–1019
Zenitani M, Nojiri T, Hosoda H, Kimura T, Uehara S, Miyazato M et al (2018) Chemotherapy can promote liver metastasis by enhancing metastatic niche formation in mice. J Surg Res 22:50–57
Huizen NA, Braak RC, Doukas M, Dekke LJ, Ijzermans J, Luider TM (2018) Up-regulation of collagen proteins in colorectal liver metastasis compared with normal liver tissue. J Biol Chem 294(1):281–289
Burnier JV, Wang N, Michel RP, Hassanain M, Li S, Lu Y et al (2011) Type IV collagen-initiated signals provide survival and growth cues required for liver metastasis. Oncogene 30(35):3766–3783
Shapouri-Moghaddam A, Mohammadian S, Vazini H et al (2018) Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol 233:6425–6440
Kimura Y, Inoue A, Hangai S, Saijo S, Negishi H, Nishio J, Yamasaki S, Iwakura Y, Yanai H et al (2016) The innate immune receptor Dectin-2 mediates the phagocytosis of cancer cells by Kupffer cells for the suppression of liver metastasi. Proc Natl Acad Sci USA 113(49):14097–14102
Keirsse J, Damme HV, Geeraerts X, Beschin A, Raes G, Ginderachter JA (2018) The role of hepatic macrophages in liver metastasis. Cell Immunol 330:202–215
Smith HA, Kang Y (2013) The metastasis-promoting roles of tumor-associated immune cells. J Mol Med 91(4):411–429
Rezaeeyan H, Shirzad R, McKee TD, Saki N (2018) Role of chemokines in metastatic niche: new insights along with a diagnostic and prognostic approach. APMIS 126:359–370
Matsusue R, Kubo H, Hisamori S et al (2009) Hepatic stellate cells promote liver metastasis of colon cancer cells by the action of SDF-1/CXCR23 Axis. Ann Surg Oncol 16(9):2645–2653
Erdogan B, Webb DJ (2017) Cancer-associated fibroblasts modulate growth factor signaling and extracellular matrix remodeling to regulate tumor metastasis. Biochem Soc Trans 45(1):229–236
Ramsey DM, Mcalpine SR (2013) Halting metastasis through CXCR25 inhibition. Bioorg Med Chem Lett 23(1):20–25
Benedicto A, Romayor I, Arteta B (2018) CXCR26 receptor blockage reduces the contribution of tumor and stromal cells to the metastatic growth in the liver. Oncol Rep 39(4):2022–2030
Li H, Batth IS, Qu X, Xu L, Song N, Wang R, Liu Y (2017) IGF-IR signaling in epithelial to mesenchymal transition and targeting IGF-IR therapy: overview and new insights. Molecular Cancer 16(1):6
Rayes RF, Milette S, Fernandez MC, Ham B, Wang N, Bourdeau F, Perrino S, Yakar S et al (2018) Loss of neutrophil polarization in colon carcinoma liver metastases of mice with an inducible, liver-specific IGF-I deficiency. Oncotarget 9(21):15691–15704
Lau EY, Ho NP, Lee TK (2017) Cancer stem cells and their microenvironment: biology and therapeutic implications. Stem Cells Int 2017:3714190
Shiozawa Y, Nie B, Pienta KJ, Morgan TM, Taichman RS (2013) Cancer stem cells and their role in metastasis. Pharmacol Ther 138(2):285–293
Shibue T, Weinberg RA (2017) EMT, CSCs, and drug resistance: the mechanistic link and clinical implications. Nat Rev Clin Oncol 14(10):611–629
Knaack H, Lenk L, Philipp LM, Miarka L, Rahn S, Viol F, Hauser C, Egberts JH, Gundlach JP, Will O, Tiwari S, Mikulits W, Schumacher U, Hengstler JG et al (2018) Liver metastasis of pancreatic cancer: the hepatic microenvironment impacts differentiation and self-renewal capacity of pancreatic ductal epithelial cells. Oncotarget 9(60):31771–31786
Costa-Silva B, Aiello NM, Ocean AJ, Singh S, Zhang H, Thakur BK, Becker A et al (2015) Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver. Nat Cell Biol 17(6):816–826
Pantel K, Alix-Panabières C (2010) Circulating tumour cells in cancer patients: challenges and perspectives. Trends Mol Med 16(9):398–406
Clark AM, Ma B, Taylor DL, Griffith L, Wells A (2016) Liver metastases: microenvironments and ex-vivo models. Exp Biol Med 241(15):1639–1652
Kapałczyńska M, Kolenda T, Przybyła W, Zajączkowska M, Teresiak A, Filas V, Ibbs M, Bliźniak R, Łuczewski Ł et al (2016) 2D and 3D cell cultures—a comparison of different types of cancer cell cultures. Arch Med Sci 14(4):910–919
Nantasanti S, de Bruin A, Rothuizen J, Penning LC, Schotanus BA (2016) Concise review: organoids are a powerful tool for the study of liver disease and personalized treatment design in humans and animals. Stem Cells Transl Med 5(325–330):37
Wang L, Liu L, Ge J, Yuan Y, Sun L, Xu H et al (2018) A multiple-cell microenvironment in a 3-dimensional system enhances direct cellular reprogramming into hepatic organoids. Transpl Proc 50(9):2864–2867
Buzzelli JN, Ouaret D, Brown G, Allen PD, Muschel RJ (2018) Colorectal cancer liver metastases organoids retain characteristics of original tumor and acquire chemotherapy resistance. Stem Cell Res 27:109–120
Gilbert T, Sellaro T, Badylak S (2006) Decellularization of tissues and organs. Biomaterials 27(19):3675–3683
Wang Y, Reid LC, Yamauchi M, Cui C, Wang AZ, Werner ME (2017) U.S. Patent No US 2017/0015974 A1. U.S. Patent and Trademark Office, Washington, DC
Tian X, Werner ME, Roche KC, Hanson AD, Foote HP, Yu SK et al (2018) Organ-specific metastases obtained by culturing colorectal cancer cells on tissue-specific decellularized scaffolds. Nat Biomed Eng 2(6):443–452
Barcus CE, O’Leary KA, Brockman JL, Rugowski DE, Liu Y, Garcia N, Yu M, Keely PJ, Eliceiri KW et al (2017) Elevated collagen-I augments tumor progressive signals, intravasation and metastasis of prolactin-induced estrogen receptor alpha positive mammary tumor cells. Breast Cancer Res 19(1):9
Wen SW, Ager EI, Christophi C (2013) Bimodal role of Kupffer cells during colorectal cancer liver metastasis. Cancer Biol Ther 14(7):606–613
Acknowledgements
Research reported in this publication was supported by the National Cancer Institute of the National Institutes of Health under Award Number T32CA196589. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Author information
Authors and Affiliations
Contributions
TV wrote the first draft and assisted with the initial revision. NS designed and drew the schematic figure and assisted in revision of the manuscript. HS developed the idea of the mini review and intoduced the subjects needed to be discussed in the paper, revised and finalized the manuscript submission. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Williamson, T., Sultanpuram, N. & Sendi, H. The role of liver microenvironment in hepatic metastasis. Clin Trans Med 8, 21 (2019). https://doi.org/10.1186/s40169-019-0237-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40169-019-0237-6