
Abstract
In this paper, we study a new SEIRS epidemic model describing nonlinear incidence with a more general form and the transmission of influenza virus with disease resistance. The basic reproductive number \(\Re_{0}\) is obtained by using the method of next generating matrix. If \(\Re_{0}<1\), the disease-free equilibrium is globally asymptotically stable, and if \(\Re_{0}>1\), by using the geometric method, we obtain some sufficient conditions for global stability of the unique endemic equilibrium. Finally, numerical simulations are provided to support our theoretical results.
Similar content being viewed by others


1 Introduction
There are lots of people dying because of infectious diseases every day. From an epidemiological viewpoint, it is important to study the global stability of disease transmission. Mathematical models describing the infectious disease dynamics have played an important role and provided the preventive strategies in a period. The SEIRS epidemic model is an important model. It shows that the total population is divided into four classes: the susceptible S, the exposed E, the infectious I and the removed R. This model has been studied by many authors [1–11]; however, many literature works did not consider disease resistance in humans. With the development and progress of society, people begin to understand the importance of health and exercise. In other words, people’s resistance has improved greatly. So, disease resistance has become an indispensable factor in the study of infectious disease models. Nguyen Huu Khanh considered the disease resistance and formulated a mathematical model [3]. In the model, a person in the exposed group or infected group can return to the susceptible group without treatment.
In fact, the disease incidence plays an important role in the study of mathematical epidemiological model. The general form of incidence rate is written as \(\beta U(N)\frac{S}{N}I\), where \(U(N)\) is usually called the contact rate. In many articles, the adequate contact rate takes two forms frequently, the corresponding disease incidence is the bilinear incidence rate \(\beta SI\) (\(U(N)=N\)) and the standard incidence rate \(\beta\frac{S}{N}I\) (\(U(N)=1\)). Between the two contact rates, there is a more realistic saturated contact rate \(U(N)=\frac{\alpha N}{1+\omega N}\) [12]. Heesterbeek et al. considered the saturated contact rate forming \(U(N)=\frac{\alpha N}{1+b N+\sqrt{1+2bN}}\) [13]. The above specific contact rates have the following common characteristic:
Based on this characteristic, more general incidence rates have also been proposed, for example, \(\beta I^{p} S^{q} \) [14, 15], \(\beta Sg(I)\) [16], \(f(S,N,I)\) [17], \(f(S,I)\) [4, 18]. Moreover, Qi and Cui established a new SEIRS epidemic model including a general incidence forming \(\beta h(S)I\) [6], and it also satisfied the above characteristics.
In real life, many infectious diseases are transmitted through both exposed individuals and infected individuals, for example, HIV, HBV and influenza. For the convenience of mathematical research, we assume that the exposed and infected individuals have the same infection rate. It is assumed that the nonlinear incidence is to be of the form \(\beta h(S)(E+I)\), where h satisfies
- (\(A_{1}\)):
-
for \(x\geq0\), \(h(x)\geq0\), with equality if and only if \(x=0\), \(h'(x)>0\) and \(h''(x)\leq0\) (where ′ represents differentiation with respect to x).
Our paper is organized as follows. In Section 2, we formulate an SEIRS mathematical model and obtain the basic reproductive number \(\Re_{0}\). Furthermore, the existence of equilibria is given. In Section 3, we prove the global stability of the disease-free equilibrium. Section 4 is devoted to the stability analysis of the endemic equilibria of the model. In Section 5, some numerical simulations are given to justify the theoretical analysis. Finally, we summarize this work.
2 The model and its basic properties
2.1 The structure of the model
We consider the transmission of influenza virus with disease resistance in humans. The total population is divided into four classes of individuals which are the susceptible (S), the exposed (E), the infected (I) and the recovered (R). The model is given by a system of ordinary differential equations
where A is the recruitment of susceptible, c, b are the rates at which the exposed and infectious individuals become susceptible individuals without treatment, respectively, ε is the constant rate for the exposed population becoming infectious, α is the rate at which the recovered individuals become susceptible individuals again, γ is the constant rate for recovery, and μ is the natural death rate of the human population. All parameters are assumed to be positive. Consider the epidemiological implications, we assume that \(b< c\).
Let \(N(t)=S(t)+E(t)+I(t)+R(t)\). The rate of change of \(N(t)\), which can be obtained by adding all the equations in model (2.1), is given by
Therefore, from biological consideration, we study (2.1) in the closed set
It is easy to see that the set Ω is a positively invariant set for (2.1).
2.2 Basic reproduction number
The basic reproduction number, denoted by \(\Re_{0}\), is ‘the expected number of secondary cases produced, in a completely susceptible population, by a typical infective individual’ [19]. We use the method of next generating matrix to determinate the expression for \(\Re_{0}\) [20]. Let \(x=(E,I,S,R)^{T}\), we rewrite system (2.1) in the matrix form
where
We can get
where \(S_{0}=\frac{A}{\mu}\). The next generation matrix for model (2.1) is
The spectral radius \(\rho(FV^{-1})\) is \(\frac{\beta h(S_{0})(b+\gamma+\mu +\varepsilon)}{(\gamma+b+\mu)(c+\varepsilon+\mu)}\). According to Theorem 2 in [20], the basic reproduction number of system (2.1) is \(\Re_{0}=\frac{\beta h(S_{0})(b+\gamma+\mu+\varepsilon)}{(\gamma+b+\mu )(c+\varepsilon+\mu)}\).
2.3 Existence of equilibria
Theorem 2.1
There exist at most two equilibria in Ω.
-
(i)
System (2.1) has the disease-free equilibrium \(E_{0}=(S_{0},0,0,0)=(\frac{A}{\mu},0,0,0)\).
-
(ii)
If \(\Re_{0}>1\), system (2.1) has two equilibria, the disease-free equilibrium \(E_{0}\) and the unique endemic equilibrium \(E_{c}=(S^{*},E^{*},I^{*},R^{*})\).
Proof
It is easy to see that the disease-free equilibrium \(E_{0}\) always exists.
From the second and third equations of (2.1), let their right-hand side be equal to zero, we have
It is easy to see that
If \(\Re_{0}>1\), \(F(S_{0})>0\), then Eq. (2.2) has a unique root \(S^{*}>0\). Hence, if \(\Re_{0}>1\), system (2.1) has a unique endemic equilibrium \(E_{c}=(S^{*},E^{*},I^{*},R^{*})\), where
The proof of Theorem 2.1 is completed. □
3 The stability of the disease-free equilibrium
In this section, we analyze the stability of the disease-free equilibrium.
Theorem 3.1
\(E_{0}\) is locally asymptotically stable if \(\Re_{0}<1\), whereas \(E_{0}\) is unstable if \(\Re_{0}>1\).
Proof
The Jacobian matrix at \(E_{0}\) is given by
We can obtain that the characteristic roots are \(\lambda_{1}=-\mu\), \(\lambda _{2}=-(\alpha+\mu)\) and the other two roots \(\lambda_{3}\) and \(\lambda_{4}\) are the roots of the following equation:
where
When \(\Re_{0}<1\), we have \(\beta h(S_{0})(\gamma+b+\mu+\varepsilon )<(c+\varepsilon+\mu)(\gamma+b+\mu)\) and \(\beta h(S_{0})< c+\varepsilon+\mu\). It is clear that \(a_{i}>0\); \(i=1,2\). By Vieta’s theorem, all roots of (3.2) are negative. Hence, \(E_{0}\) is locally asymptotically stable.
When \(\Re_{0}>1\), we have \(a_{2}<0\), (3.2) has a positive root, so \(E_{0}\) is unstable. □
In the following, applying LaSalle’s invariance principle and the Lyapunov direct method, we prove the global stability of the disease-free equilibrium.
Theorem 3.2
The disease-free equilibrium \(E_{0}\) is globally asymptotically stable if \(\Re_{0}<1\).
Proof
Define a Lyapunov function
When \(\Re_{0}<1\), we have \(\beta h(S_{0})< c+\varepsilon+\mu\), then \(V(t)\geq0\). The total derivative of V along the solutions of (2.1) is
From \(h'(S)>0\) and \(0< S< S_{0}\), we have \(h(S)< h(S_{0})\), and then
It is easy to see that \(V'(t)\leq0\) and \(V'(t)=0\) if and only if \(I(t)=0\). It follows from (2.1) that \(E(t)\rightarrow0\), \(R(t)\rightarrow0\), \(S(t)\rightarrow S_{0}\) as \(t\rightarrow+\infty\), and then \(E_{0}\) is the largest invariant subset of the invariant set \(\{(S,E,I,R)\in\Omega: V'(t)=0\}\). Therefore, by the LaSalle’s invariance principle, \(E_{0}\) is globally attractive in Ω. This, combined with the local stability of \(E_{0}\), completes the proof. □
4 The stability of the endemic equilibrium
In this section, we analyze the stability of the endemic equilibrium.
Theorem 4.1
If \(\Re_{0}>1\), \(\beta h(S^{*})\leq c+\varepsilon\) and \(\beta h'(S^{*})I^{*}\geq\frac {b+\gamma+\mu-\varepsilon}{b+\gamma+\mu+\varepsilon}-(\mu+b)\), then the endemic equilibrium \(E_{c}\) is locally asymptotically stable.
Proof
The Jacobian matrix at \(E_{c}\) is given by
The characteristic equation is
where
and
From Eq. (2.2), we have \((\gamma+b+\mu)(c+\varepsilon+\mu)-\beta h(S^{*})(b+\gamma+\mu+\varepsilon)=0\) and \(\beta h(S^{*})< c+\varepsilon+\mu\), so we get \(M=0\), \(F>0\).
When \(\beta h(S^{*})\geq c+\varepsilon\) and \(b< c\), we have \(\beta h(S^{*})>b+\varepsilon\). Take notice of condition \(\beta h'(S^{*})I^{*}\geq\frac{b+\gamma+\mu-\varepsilon}{b+\gamma+\mu +\varepsilon}-(\mu+b)\). It is easy to see that \(a_{i}>0\); \(i=1,2,3,4\), \(a_{1}a_{2}-a_{3}>0\) and \(a_{4}(a_{3}(a_{1}a_{2}-a_{3})-a_{1}^{2}a_{4})>0\). By the Routh-Hurwitz criterion, all roots of (4.2) have negative real parts. Hence, the endemic equilibrium \(E_{c}\) of system (2.1) is locally asymptotically stable.
The proof of Theorem 4.1 is completed. □
In the following, we use the geometric approach to discuss the global stability of the endemic equilibrium. We will expand its application to four-dimensional systems, which can also be seen in [21].
Firstly, we present some preliminaries on the geometric approach to prove global stability [22].
Definition 4.1
System (2.1) is said to be uniformly persistent in Ω if there exists a constant \(k > 0\) such that any solution \((S(t),E(t),I(t),R(t))\) of system (2.1) with the initial value \((S(0),E(0),I(0),R(0)) \in\operatorname{int}\Omega\) satisfies
Similar to [7], we can get the following.
Theorem 4.2
System (2.1) is uniformly persistent in Ω if and only if \(\Re_{0}>1\).
Remark 4.1
The uniform persistence of system (2.1) in the bounded set Ω is equivalent to the existence of a compact \(K\subset\Omega\) that is absorbing for (2.1) (see [23]). Denote
where \(\delta_{i}>0\), \(i=S,E,I,R\).
Consider the autonomous dynamical system
where \(x\rightarrow f(x)\in R^{n}\) is a \(C^{1}\) function about x in \(\Omega_{1}\subset R^{n}\).
Assume that the following hypotheses hold:
- (H1):
-
There is a compact absorbing set \(K\subset\Omega_{1}\);
- (H2):
-
Differential equation (4.3) has a unique equilibrium \(x^{*}\) in \(\Omega_{1}\).
Let \(x \rightarrow P(x)\) be a \(\bigl({\scriptsize\begin{matrix}{} n\cr 2 \end{matrix}} \bigr)\times \bigl({\scriptsize\begin{matrix}{} n\cr 2 \end{matrix}} \bigr)\) matrix-valued function that is \(C^{1}\) for \(x \in\Omega_{1}\). Assume that \(P^{-1}(x)\) exists and is continuous for \(x \in K\) and consider
where the matrix \(P_{f}\) is \((p_{ij}(x))_{f}=(\frac{\partial p_{ij}(x)}{\partial x})^{T}\cdot f(x)=\nabla p_{ij}(x)\cdot f(x)\), \(J^{[2]}\) is the second additive compound matrix of the Jacobian matrix J, i.e., \(J(x)=Df(x)\). Consider the Lozinskiĭ measure μ of Q with respect to a vector norm \(\Vert\cdot\Vert\) in \(R^{ \bigl({\scriptsize\begin{matrix}{}n\cr 2 \end{matrix}} \bigr)}\) (see [24]), that is,
A quantity q is defined as follows:
The following global stable result is proved in Theorem 3.5 of [22].
Lemma 4.1
([22])
Suppose that \(\Omega_{1}\) is simply connected and that assumptions (H1) and (H2) hold, then the unique equilibrium \(x^{*}\) of system (4.3) is globally stable in \(\Omega _{1}\) if \(q < 0\).
Now we apply Lemma 4.1 to prove the global stability of \(E_{c}\).
Theorem 4.3
If \(\Re_{0}>1\), then the endemic equilibrium \(E_{c}\) of system (2.1) is globally asymptotically stable provided that
where \(d=\min\{b,\alpha\}\).
Proof
The Jacobian matrix of system (2.1) is given by
Hence, the second additive compound matrix \(J^{[2]}\) [21] of J is given by
where
Let
then
Let
The matrix \(Q(S,E,I,R)\) can be written in the block form
where
Let \(z=(z_{1},z_{2},z_{3},z_{4},z_{5},z_{6})\) denote a vector in \(R^{6} \simeq R^{ \bigl({\scriptsize\begin{matrix}{} 4\cr2\end{matrix}} \bigr)}\), we select a norm in \(R^{6}\) as
Let \(\sigma(Q)\) be the Lozinskiĭ measure of Q with respect to the induced matrix norm \(\Vert\cdot\Vert\) in \(R^{6}\), defined by
We have the following estimate:
where
\(|Q_{ij}|\) (\(i\neq j\), \(i,j =1,2,3,4\)) are matrix norms with respect to the \(l_{1}\) vector norm, and \(\sigma_{1}\) denotes the Lozinskiĭ measure with respect to the \(l_{1}\) norm.
From the first equations of (2.1), we have
From equation (4.6), it easy to prove that there is \(t^{*}\), when \(t>t^{*}\), we have \(\beta h(S)-b>0\). In fact, if \(\beta h(S)\leq b\), from equation (4.6) and \(b< c\), we have \(S'\geq A-\mu S=\mu(\frac{A}{\mu}-S)>0\), which means that each solution starting from K must have crossed the curve \(\beta h(S)=b\) in a limited time; this is contradiction to \(\beta h(S)\leq b\). To calculate the values of \(g_{i}\), we firstly obtain that
and
where \(d=\min\{\alpha,b\}\). From \(h''(S)<0\), we have \(h'(M_{S})< h'(S)< h'(\delta_{S})\). Then we further have
Let
From condition (4.4), we have \(\bar{b}>0\) and
Along each solution \((S(t),I(t),R(t),I(t))\) of system (2.1) with the initial value \((S(0),I(0), R(0),I(0))\in K\), when \(t>t^{*}\), we have
Furthermore, we have
Therefore,
The proof of Theorem 4.1 is completed. □
5 Numerical simulation
To support our main results, we perform some numerical simulations. We choose \(h(S)=\frac{S}{1+gS}\) and consider the set of parameters:
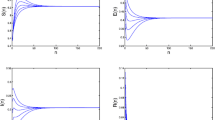
(1) \(\beta= 0.025\), \(\alpha= 0.25\), \(c = 0.15\), \(\mu= 0.15\), \(A = 0.1\), \(\gamma= 0.4\), \(g=0.8\), \(\varepsilon=0.25\), with the initial condition \((S(0),E(0),I(0),R(0))=(0.15,0.15,0.15,0.15)\), we have \(\Re _{0}=0.3403<1\). In this case, according to Theorem 3.2, the disease-free equilibrium \(E_{0}\) of system (2.1) is globally asymptotically stable (see Figure 1).

The disease-free equilibrium \(\pmb{E_{0}}\) is globally asymptotically stable.
(2) \(\beta=0.35\), \(\alpha= 0.04\), \(b = 0.04\), \(c = 0.05\), \(\mu= 0.23\), \(A = 0.23\), \(\gamma=0.008\), \(g = 0.1\), \(\varepsilon= 0.25\), with the initial condition \((S(0),E(0),I(0),R(0))=(0.5,0.1,0.1,0.1)\), we have \(\Re_{0}=1.1402>1\), and \(3\beta h(M_{S})-\beta h'(M_{S})(\delta_{E}+\delta_{I})-c-\varepsilon -b=0.4529\), \(\varepsilon+c-d-\beta h(\delta_{S})-\beta h'(M_{S})(\delta _{E}+\delta_{I})=0.0948\), \(\beta h'(\delta_{S})(M_{E}+M_{I})+2\beta h(M_{S})-c-d-\varepsilon=0.2778\), \(\varepsilon-\alpha-b-\gamma=0.1620\), so condition (4.4) is satisfied. According to Theorem 4.3, the endemic equilibrium \(E_{c}\) of system (2.1) is globally asymptotically stable (see Figure 2).

The endemic equilibrium \(\pmb{E_{c}}\) of system ( 2.1 ) is globally asymptotically stable.
6 Conclusions
In this paper, we have proposed a nonlinear mathematical model for influenza virus transmission with disease resistance; nonlinear incidence has a more general form. Through mathematical analysis we obtain the dynamic behaviors of the model. The basic reproduction number \(\Re_{0}\) is obtained. If \(\Re_{0}<1\), the disease-free equilibrium is globally asymptotically stable. It implies that the disease dies out eventually. When \(\Re_{0}>1\), the endemic equilibrium is globally asymptotically stable under some conditions. It implies that the disease persists in the population. All of these results imply that the disease resistance and nonlinear incidence can influence the dynamic behaviors of the SEIRS model. From the expression of \(\Re_{0}\), it is easy to see that when β is increased, b, c decrease and then \(\Re_{0}\) increases. \(\Re_{0}>1\) leads to the stability of the endemic equilibrium and then the prevalence of the disease. So we can get some effective strategies for controlling the disease such as reducing the contact rate β and increasing the b, c. That is to say, by taking proper isolation of the population and increasing the resistance of people, we can avoid the development of infectious diseases into endemic diseases. In reality, the exposed and infected individuals have different infection rates, which will be the focus of our future research.
References
Beretta, E, Capasso, V: On the general structure of epidemic systems, global asymptotic stability. Comput. Math. Appl. 12A, 677-694 (1986)
Buonomo, B, d’Onofrio, A, Lacitignola, D: Modeling of pseudo-rational exemption to vaccination for SEIR diseases. J. Math. Anal. Appl. 404, 385-398 (2013)
Khanh, NH: Stability analysis of an influenza virus model with disease resistance. J. Egypt. Math. Soc. 24, 193-199 (2016)
Mateus, JP, Silva, CM: Existence of periodic solutions of a periodic SEIRS model with general incidence. Nonlinear Anal., Real World Appl. 34, 379-402 (2017)
Nakata, Y, Kuniya, T: Global dynamics of a class of SEIRS epidemic models in a periodic environment. J. Math. Anal. Appl. 363, 230-237 (2010)
Qi, LX, Cui, JA: The stability of an SEIRS model with nonlinear incidence, vertical transmission and time delay. Appl. Math. Comput. 221, 360-366 (2013)
Sun, CJ, Lin, YP, Tang, SP: Global stability for a special SEIR epidemic model with nonlinear incidence rates. Chaos Solitons Fractals 33, 290-297 (2007)
Tipsri, S, Chinviriyasit, W: The effect of time delay on the dynamics of an SEIR model with nonlinear incidence. Chaos Solitons Fractals 75, 153-172 (2015)
Wang, WD: Global behavior of an SEIRS epidemic model with time delays. Appl. Math. Lett. 15, 423-428 (2002)
Zhang, TL, Teng, ZD: Global asymptotic stability of a delayed SEIRS epidemic model with saturation incidence. Chaos Solitons Fractals 37, 1456-1468 (2008)
Zhou, X, Cui, JA: Analysis of stability and bifurcation for an SEIR epidemic model with saturated recovery rate. Commun. Nonlinear Sci. Numer. Simul. 16, 4438-4450 (2011)
Dietz, K: Overall population patterns in the transmission cycle of infectious disease agents. In: Population Biology of Infectious Disease. Springer, Berlin (1982)
Heesterbeek, JAP, Metz, JAJ: The saturating contact rate in marriage and epidemic models. J. Math. Biol. 31, 529-539 (1993)
Liu, WM, Levin, SA, Iwasa, Y: Influence of nonlinear incidence rates upon the behavior of SIRS epidemiological models. J. Math. Biol. 23, 187-204 (1986)
Li, MY, Muldowney, JS: Global stability for the SEIR model in epidemiology. Math. Biosci. 125, 155-164 (1995)
Van den Hethcote, HW, Driessche, P: Some epidemiological models with nonlinear incidence. J. Math. Biol. 29, 271-287 (1991)
Mateus, JP, Silva, CM: A non-autonomous SEIRS model with general incidence rate. Appl. Math. Comput. 247, 169-189 (2014)
Korobeinikov, A: Global properties of infectious disease models with nonlinear incidence. Bull. Math. Biol. 69, 1871-1886 (2007)
Diekmann, O, Heesterbeek, JAP, Metz, JAJ: On the definition and the computation of the basic reproduction ratio \(\Re_{0}\) in models for infectious diseases in heterogeneous populations. J. Math. Biol. 28, 365-382 (1990)
Driessche, P, Watmough, J: Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 180, 29-48 (2002)
Feng, XM, Ruan, SG, Teng, Z, Wang, K: Stability and backward bifurcation in a malaria transmission model with applications to the control of malaria in China. Math. Biosci. 266, 52-64 (2015)
Li, MY, Muldowney, J: A geometric approach to global stability problems. SIAM J. Math. Anal. 27, 1070-1083 (1996)
Butler, GJ, Waltman, P: Persistence in dynamics systems. J. Differ. Equ. 63, 255-263 (1986)
Martin, RH: Logarithmic norms and projections applied to linear different differential systems. J. Math. Anal. Appl. 45, 432-454 (1974)
Kermack, WO, McKendrick, AG: Contribution to mathematical theory of epidemics. Proc. R. Soc. A 115, 700-721 (1927)
Korobeinikon, A, Wake, GC: Lyapunov functions and global stability for SIR, SIRS, and SIS epidemiological models. Appl. Math. Lett. 15, 955-960 (2002)
Li, Y, Muldowney, JS: On Bendixsons criterion. J. Differ. Equ. 106, 27-39 (1993)
Brown, GC, Hasibuan, R: Conidial discharge and transmission efficiency of neozygites floridana, an entomopathogenic fungus infecting two-spotted spider mites under laboratory conditions. J. Invertebr. Pathol. 65, 10-16 (1995)
Li, XB, Yang, LJ: Stability analysis of an SEIQV epidemic model with saturated incidence rate. Nonlinear Anal., Real World Appl. 13, 2671-2679 (2012)
Yang, LX, Yang, XF, Zhu, QY, Wen, LS: A computer virus model with graded cure rates. Nonlinear Anal., Real World Appl. 14, 414-422 (2013)
Capasso, V, Serio, G: A generalization of the Kermack-McKendrick deterministic epidemic model. Math. Biosci. 42, 43-61 (1978)
Acknowledgements
We greatly appreciate the editor and the anonymous referees’ careful reading and valuable comments, their critical comments and helpful suggestions have greatly improved the presentation of this paper.
Author information
Authors and Affiliations
Contributions
All authors contributed equally to the writing of this paper. The authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Jia, J., Xiao, J. Stability analysis of a disease resistance SEIRS model with nonlinear incidence rate. Adv Differ Equ 2018, 75 (2018). https://doi.org/10.1186/s13662-018-1494-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-018-1494-1