
Abstract
In this paper, we investigate the stochastic disease dynamics of an SEIS epidemic model with latent patients and active patients. The two parameters \(R_{0}^{s}\) and \(R_{0}^{*}\) are identified as the disease-free and endemic dynamics of the model. More specifically, we give the almost surely exponential stability of the disease-free equilibrium in terms of \(R_{0}^{s}\), and stochastic endemic dynamics in terms of \(R_{0}^{*}\). The theoretical and numerical results may be useful for studying the dynamics of disease spreading in a randomly fluctuating environment.
Similar content being viewed by others


1 Introduction
Mathematical models have been an important tool in analyzing the spread and control of infectious diseases since the pioneer work of Kermack and McKendrick [1]. Most of the research literature on these types of models assumes that the disease incubation is negligible so that, once infected, each susceptible individual (in the class S) instantaneously becomes infectious (in the class I) and later recovers (in the class R) with a permanent or temporary acquired immunity [2]. A compartmental model based on these assumptions is customarily called a SIR or SIRS model. Regarding research on the SIR or SIRS models and its generalizations, the reader can refer to [3–5].
Some diseases, however, incubate inside the hosts for a period of time before the hosts become infectious. In the case of assuming that the susceptible individual first goes through a latent period after infection before becoming infectious, the resulting models are of SEI, SEIR or SEIRS type, respectively, depending on whether the acquired immunity is permanent or otherwise. There has been a great deal of work on these types of models in the literature [6–9]. Some scholars simply considered the therapy of the active patients and neglected the importance of remedying the latent patients. However, it is very important to cure the latent patients because if some diseases such as the tuberculosis (see [10]) miss the best treatment time in latent period, the diseases will be fatal. Especially, Meng et al. [11] discussed an SEIS epidemic model in a population that is compartmentalized into three classes: the susceptible, exposed, and infectious classes, with sizes denoted by \(S(t)\), \(E(t)\), \(I(t)\), respectively. The system has the following form:
where μ is the death rate for physical disease, Λ the influx or recruitment of the susceptible and the exposed, \(\gamma_{1}\) and \(\gamma_{2}\) the treatment cure rate of latent and active disease, respectively, β the breakdown rate from latent to active condition, \(\alpha S(t)I(t)\) the bilinear incidence, p the proportion of infection instantaneously degenerating in active condition, the dynamics of latently infected population depends on the proportion of the infection that results in latent infection \((1-p)\alpha I(t)S(t)\). The basic reproduction number of model (1) is
which determines the extinction and persistence of the epidemic. According to the results in [11], one can see that
-
(a)
if \(0<\mathcal{R}_{0}<1\), the disease-free equilibrium \((\Lambda /\mu, 0, 0)\) is globally asymptotically stable, and it is unstable when \(\mathcal{R}_{0}>1\);
-
(b)
if \(\mathcal{R}_{0}>1\), the endemic equilibrium \((S^{*}, E^{*}, I^{*})\) of model (1) is globally asymptotically stable, where
$$ \begin{aligned} &S^{*}=\frac{(\beta+\gamma_{1}+\mu)(\gamma_{2}+\mu)}{\alpha(\beta +\gamma_{1}p+\mu p)}, \\ &E^{*}=\frac{\beta\Lambda\alpha(1-p)^{2}(\gamma_{2}+\mu)}{ \alpha\mu(\beta+\gamma_{1}p+\mu p)(\beta+\mu+\gamma_{1}p+\gamma_{2}(1-p))} \biggl(1-\frac{1}{\mathcal{R}_{0}} \biggr), \\ &I^{*}=\frac{(\beta+\gamma_{1}p+\mu p)E^{*} }{(1-p)(\gamma_{2}+\mu)}. \end{aligned} $$(2)
However, the deterministic approach has some limitations in the mathematical modeling transmission of an infectious disease. Stochastic differential equation (SDE) models play a significant role in various branches of applied sciences including infectious dynamics, as they provide some additional degree of realism compared to their deterministic counterpart [12]. Recently, many authors have introduced parameter perturbation into epidemic models and have studied their dynamics [13–20].
In this paper, taking account of the effect of randomly fluctuating environment, we incorporate white noise in each equation of model (1). We assume that fluctuations in the environment will manifest themselves mainly as fluctuations in parameters \(\gamma_{1}\), \(\gamma_{2}\) as follows:
where \(B_{i}(t)\) (\(i=1,2\)) is for the mutually independent standard Brownian motions with \(B_{i}(0)=0\), and \(\sigma_{i}^{2}\) (\(i = 1, 2\)), the intensities of white noise. The stochastic version corresponding to the deterministic model (1) takes the following form:
The equation for the total population \(N(t)=S(t)+ E(t) + I(t)\) size is obtained from (3) as
It follows that
By (4), we take \(S(t)=\frac{\Lambda}{\mu}-E(t)-I(t)\), and substitute it into the second and the third equations of model (3), and we can easily obtain the following limit system:
with any given initial value \((E(0), I(0)\in\mathbb{R}_{+}^{2}\). It is easy, by simple computations, to conclude that model (5) has a unique disease-free equilibrium, \(E_{0}=(0,0)\).
This paper is organized as follows. In Section 2, we give some preliminaries. In Section 3, we deduce the conditions which will cause the disease to die out. The condition for the disease to be persistent (i.e., endemic) is given in Section 4. In Section 5, we provide some numerical examples to support our analytic results. In the last section, Section 6, we provide a brief discussion and summary of main results.
2 Preliminaries
Throughout this paper, let \((\Omega, \mathcal{F}, \{\mathcal{F}_{t}\}_{t\in\mathbb{R}_{+}} \mathbb{P})\) be a complete probability space with a filtration \(\{\mathcal{F}_{t}\}_{t\in\mathbb{R}_{+}}\) satisfying the usual conditions (i.e., it is right continuous and increasing while \(\mathcal{F}_{0}\) contains all \(\mathbb{P}\)-null sets). Let
Consider the general n-dimensional stochastic differential equation
on \(t\geq0\) with initial value \(x(0)=x_{0}\), the solution is denoted by \(x(t, x_{0})\). Assume that \(f(0,t) =0\) and \(\varphi(0, t) = 0\) for all \(t\geq0\), and equation (6) has the solution \(x(t)= 0\), which is called the trivial solution.
Definition 2.1
[21]
The trivial solution \(x(t)= 0\) of equation (6) is said to be almost surely exponentially stable if for all \(x_{0}\in \mathbb{R}^{n}\),
Definition 2.2
[22]
The population \(x(t)\) is said to be strongly persistent in the mean if \(\liminf_{t\rightarrow\infty} \frac{1}{t} \int_{0}^{t}x(s)\,ds>0\).
The differential operator \(\mathcal{L}\) associated with the function displayed in equation (6) is defined for a function \(V(t,x)\in C^{1,2}(\mathbb{R}\times\mathbb{R}^{n})\) by the formula
where Trc means trace and trp denotes the transpose of a matrix.
The following lemma is quoted from [16, 23] where it was proved and applied. It plays a similar role in this paper.
Lemma 2.3
Let \(x\in C[\Omega\times[0,\infty),(0,\infty)]\). If there exist positive constants λ, μ such that
for all \(t\geq0\), where \(F \in C[\Omega\times[0,\infty),\mathbb {R}]\) and \(\lim_{t\rightarrow\infty}\frac{F(t)}{t}=0\), then
To investigate the dynamical behavior of a population model, the first concern is whether the solution of the model is positive and global. Motivated by [14], we can prove the global existence of a solution to model (5). One can obtain the following results.
Lemma 2.4
Let \((E(0),I(0))\in\Gamma\), and model (5) admits a unique solution \((E(t),I(t))\) on \(t\geq0\), which remains in Γ with probability 1. That is, the set Γ is almost surely positively invariant of model (5).
3 Stochastic disease-free dynamics
Now we present the following theorem, which gives conditions for the almost surely exponential stability of the equilibrium of model (5), which is motivated by [21, 24]. Denote \(\sigma:=\min\{\sigma_{1},\sigma_{2}\}\) and \(X(t):=(E(t),I(t))\). First of all, we give the property of the disease-free (i.e., \(I=0\)) dynamics.
Theorem 3.1
If
then the disease-free equilibrium \((0,0)\) of model (5) is almost surely exponentially stable. In other words, the disease will die out with probability one.
Proof
Let us fix any positive real number \(a_{1}\). We define the following stochastic process:
Since \(z(X(t)>0)\) for all \(t>0\), we can define a \(C^{2}\)-function \(V:\mathbb{R}_{+}^{2}\rightarrow\mathbb{R}_{+}\) by
By Itô’s formula, we can express the stochastic process \(V(X(x))\) as
where
and
Regarding the quadratic variations of the stochastic integral \(G(t)\) we have
By the strong law of large numbers for martingales [25], we have
Now we prove that
To this end, we set
It follows that
that is, the stochastic processes \(v(t)\), \(w(t)\) are bounded above by \(\max\{1/a_{1},1\}\), and non-negative. By virtue of (9) we have
This enables us to write down the expression for \(\mathcal{L}V(X(t))\) as
In view of \(a_{1}v+w=1\), we have
It follows that
where
Let \(a_{1}=\frac{\beta}{\beta+\gamma_{1}+\mu+{\sigma^{2}}/{4}}\), then \(A_{1}=0\). The condition \(R_{0}^{s} < 1\) is equivalent to the following inequality:
which implies \(A_{2}<0\). It follows from \(0< w<1\) that \(\mathcal{L}V(X(t))<0\). It finally follows from (10) by dividing t on both sides and then letting \(t\rightarrow\infty\) that
which is the required assertion. □
Remark 3.2
It is noted that \(R_{0}^{s}=\frac{\alpha\beta(1-p) \Lambda}{(\beta +\gamma_{1}+\mu+\sigma^{2}/4) (\mu\gamma_{2}+\mu^{2}+\mu\sigma^{2}/4-\alpha p\Lambda)}< R_{0}\). Therefore, if \(R_{0}<1\), no matter how the noise intensities change, we have the disease-free equilibrium to be almost surely exponentially stable. However, if \(R_{0}>1\), by increasing the values of noise intensities such that \(R_{0}^{s}<1\), the disease-free equilibrium will still be almost surely exponentially stable. That is to say, in this situation, for the deterministic model, there is an endemic equilibrium which is globally stable, but for the stochastic model, there is a stable disease-free equilibrium which means that the disease goes extinct exponentially a.s.
4 Stochastic endemic dynamics
In this section we intend to prove the stochastic endemic dynamics (i.e., persistence of E and I) of model (5) under certain parametric restrictions.
Theorem 4.1
If
then for any initial value \((E(0), I(0))\in\Gamma\), the solution \((E(t), I(t))\) of model (5) has the following property:
and
That is the solutions of model (5) are strongly persistent in the mean.
Proof
An integration of the first equation of model (5) yields
We compute that
where
Since \(E(t),I(t)<\Lambda/\mu\), by the strong law of large numbers for martingales [25], we have
Obviously, \(\lim_{t\rightarrow\infty} \varphi(t)=0\) a.s.
Applying Itô’s formula to the first equation of model (5) leads to
An integration of (19) yields
Noting that \(-\infty<\ln(\mu/\Lambda(E(t))<0\) (as \(0< E(t)<\Lambda /\mu\)), then for arbitrary \(0<\varepsilon<1\), there exist \(T=T(\omega)>0\) and a set \(\Omega_{\varepsilon}\), such that \(P(\Omega_{\varepsilon})\geq1-\varepsilon\). For all \(t\geq T(\omega)\), \(\omega\in \Omega_{\varepsilon}\),
which is equivalent to
It follows from \(0< E(t),I(t)<\Lambda/\mu\) that
Hence, applying Itô’s formula to the second equation of model (5) leads to
Integrating this from 0 to t, we have
Since
it follows from Lemma 2.3 that
Finally, according to the last equality of (20), we get
This finishes the proof. □
Remark 4.2
It is noted that \(R_{0}^{*}< R_{0}^{s}< R_{0}\). Therefore, if \(R_{0}^{*}>1\), then \(R_{0}>1\). That is to say, if for stochastic model the disease will be prevalent, for a deterministic model the disease also must be prevalent.
5 Numerical simulations
In this section, we give some numerical simulations to show the effect of noise on the dynamics of model (5) by using the Milstein method mentioned in Higham [26].
For model (5), the parameters are taken as follows:
with initial values
-
1.
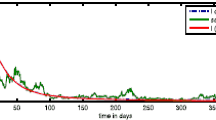
Now we note that these parameters give a value of \({R}_{0} =1.667\) to the basic reproduction number in the deterministic case (i.e., with \(\sigma_{1}=\sigma_{2}=0\)). Consequently the system eventually approaches an endemic equilibrium point \(( 0.0523, 0.18)\) (see Figure 1).
-
2.
Choose \(\sigma_{1}=0.75\), \(\sigma_{2}=0.8\), then we obtain \({R}_{0}^{s}=0.9279\). Theorem 3.1 asserts that the disease-free equilibrium is almost surely exponentially stable, and the disease will die out with probability one (see Figure 2).
-
3.
Choose \(\sigma_{1}=0.1\), \(\sigma_{2}=0.2\), then we obtain \({R}_{0}^{s}=1.6450\) and \({R}_{0}^{*}=1.5207\). Theorem 4.1 asserts that the solutions of model (5) are strongly persistent in mean (see Figure 3).
-
4.
Now change \(\sigma_{1}=0.1\), \(\sigma_{2}=0.6\), then we obtain \({R}_{0}^{s}=1.6450\) and \({R}_{0}^{*}=0.9063\). Therefore, the conditions of Theorems 3.1 and 4.1 are not satisfied. In this case, our simulations suggest that the disease will die out with probability one (see Figure 4).




6 Discussions
In this paper, we mainly focus on the SDE version of an SEIS epidemic model with latent patients and active patients. We show that the SDE model has a unique positive global solution and establish some conditions for determining the disease outbreak or extinct. The key parameters are \(R_{0}^{s}\) and \(R_{0}^{*}\), which are all less than the corresponding deterministic version of the basic reproduction number \(R_{0}\).
Theorem 3.1 shows that if \(R_{0}^{S}<1\), the disease will die out (cf. Figure 2). Theorem 4.1 shows that if \(R_{0}^{*}>1\), then the disease will persist (cf. Figure 3). By numerical simulations, we also show that if \(R_{0}^{*}<1<R_{0}^{S}\), the disease will die out (cf. Figure 4). Hence, we can make a conjecture that the behavior of the disease is determined by \(R_{0}^{*}\). It is well known that for deterministic epidemic models, the basic reproduction number \(R_{0}\) determines the prevalence or extinction of the disease. In this paper, we consider the threshold \(R_{0}^{*}\) as the basic reproduction number of model (5). Notice that \(R_{0}^{*}< R_{0}\), and it is possible that \(R_{0}^{*}<1<R_{0}\). This is the case when the deterministic model has an endemic (see Figure 1), while the stochastic model has disease extinction with probability one (see Figure 4). That is to say, in this case, noise can suppress the disease outbreak.
References
Kermack, WO, McKendrick, AG: A contribution to the mathematical theory of epidemics. Proc. R. Soc. Lond. Ser. A 115, 700-721 (1927)
Li, MY, Graef, JR, Wang, L, Karsai, J: Global dynamics of a SEIR model with varying total population size. Math. Biosci. 160(2), 191-213 (1999)
Liu, W, Levin, SA, Iwasa, Y: Influence of nonlinear incidence rates upon the behavior of SIRS epidemiological models. J. Math. Biol. 23(2), 187-204 (1986)
Durrett, R: Stochastic spatial models. SIAM Rev. 41(4), 677-718 (1999)
Hethcote, HW: The mathematics of infectious diseases. SIAM Rev. 42(4), 599-653 (2000)
Liu, W, Hethcote, HW, Levin, SA: Dynamical behavior of epidemiological models with nonlinear incidence rates. J. Math. Biol. 25(4), 359-380 (1987)
Smith, HL, Wang, L, Li, MY: Global dynamics of an SEIR epidemic model with vertical transmission. SIAM J. Appl. Math. 62(1), 58-69 (2001)
Li, G, Jin, Z: Global stability of an SEI epidemic model. Chaos Solitons Fractals 21(4), 925-931 (2004)
Zhang, T, Teng, Z: Global asymptotic stability of a delayed SEIRS epidemic model with saturation incidence. Chaos Solitons Fractals 37(5), 1456-1468 (2004)
Connell McCluskey, C: Global stability for a class of mass action systems allowing for latency in tuberculosis. J. Math. Anal. Appl. 338(1), 518-535 (2008)
Meng, X, Wu, Z, Zhang, T: The dynamics and therapeutic strategies of a SEIS epidemic model. Int. J. Biomath. 6, 1350029 (2013)
Zhao, Y, Jiang, D, Mao, X, Gray, A: The threshold of a stochastic sirs epidemic model in a population with varying size. Discrete Contin. Dyn. Syst., Ser. B 20(4), 1277-1295 (2015)
Gray, A, Greenhalgh, D, Hu, L, Mao, X, Pan, J: A stochastic differential equation SIS epidemic model. SIAM J. Appl. Math. 71(3), 876-902 (2011)
Lahrouz, A, Omari, L, Kiouach, D: Global analysis of a deterministic and stochastic nonlinear SIRS epidemic model. Nonlinear Anal., Model. Control 16(1), 59-76 (2011)
Yang, Q, Mao, X: Extinction and recurrence of multi-group SEIR epidemic models with stochastic perturbations. Nonlinear Anal., Real World Appl. 14(3), 1434-1456 (2013)
Ji, C, Jiang, D: Threshold behaviour of a stochastic SIR model. Appl. Math. Model. 38(21), 5067-5079 (2014)
Liu, M, Bai, C, Wang, K: Asymptotic stability of a two-group stochastic SEIR model with infinite delays. Commun. Nonlinear Sci. Numer. Simul. 19(10), 3444-3453 (2014)
Cai, Y, Kang, Y, Banerjee, M, Wang, W: A stochastic SIRS epidemic model with infectious force under intervention strategies. J. Differ. Equ. 259(12), 7463-7502 (2015)
Li, D, Cui, J, Liu, M, Liu, S: The evolutionary dynamics of stochastic epidemic model with nonlinear incidence rate. Bull. Math. Biol. 77(9), 1705-1743 (2015)
Cai, Y, Kang, Y, Banerjee, M, Wang, W: A stochastic epidemic model incorporating media coverage. Commun. Math. Sci. 14(4), 892-910 (2016)
Khasminskii, R: Stochastic Stability of Differential Equations, vol. 66. Springer, Berlin (2012)
Mandal, PS, Banerjee, M: Stochastic persistence and stationary distribution in a Holling-Tanner type prey-predator model. Physica A 391(4), 1216-1233 (2012)
Liu, H, Ma, Z: The threshold of survival for system of two species in a polluted environment. J. Math. Biol. 30(1), 49-61 (1991)
Witbooi, PJ: Stability of an SEIR epidemic model with independent stochastic perturbations. Physica A 392(20), 4928-4936 (2013)
Mao, X: Stochastic Differential Equations and Their Applications. Horwood, Chichester (1997)
Higham, DJ: An algorithmic introduction to numerical simulation of stochastic differential equations. SIAM Rev. 43(3), 525-546 (2001)
Acknowledgements
The author thanks the referees for their important and valuable comments. This work is supported by the Natural Science Foundation of Lanzhou University of Arts and Science.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The author declares to have no competing interests.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Yang, B. Stochastic dynamics of an SEIS epidemic model. Adv Differ Equ 2016, 226 (2016). https://doi.org/10.1186/s13662-016-0914-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-016-0914-3