
Abstract
Telomeric DNA are TTAGGG tandem repeats, which are susceptible for oxidative DNA damage and hotspot regions for formation of DNA secondary structures such as t-loop, D-loop, G-quadruplexes (G4), and R-loop. In the past two decades, unique DNA or RNA secondary structures at telomeres or some specific regions of genome have become promising therapeutic targets. G-quadruplex and R-loops at telomeres or transcribed regions of genome have been considered as the potential targets for cancer therapy. Here we discuss the potentials to target the secondary structures (G4s and R-loops) in genome as therapy approaches.
Similar content being viewed by others
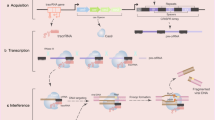


Background
Telomeres are nucleoprotein structures at the end of each chromosome, which protects the end of the chromosome from deterioration or from fusion with chromosomes, and the hotspot region for formation of many secondary nucleotide structures [1, 2]. Cancer cells get the ability to conquer the replication problem via either telomerase or Alternative Lengthening of Telomeres (ALT) pathway, which involves recombinational mechanisms to overcome incomplete replication of telomeres [3,4,5] (Fig. 1). Telomeric DNA are TTAGGG tandem repeats, which are susceptible for oxidative DNA damage and hotspot regions for formation of DNA secondary structures such as t-loop, D-loop, G-quadruplex (G4), and R-loop [6,7,8]. Single strand G-rich overhang folds back and invades into the double-stranded telomere tract to form a T-loop and D-loop structure, protecting the end of chromosome from being recognize as double strands breaks (DSBs) [9,10,11]. When single-stranded guanine-rich DNA sequences fold into stable intramolecular and intermolecular four-stranded non-B DNA structures, such a structure is so called G-quadruplexes, which may play important roles in the regulation of gene expression, DNA repair, epigenetic regulation and telomere biology [12,13,14]. R-loops are the three-stranded nucleic acid structure that contains a DNA: RNA hybrid and displaced DNA strand. We recently discovered that ROS-induced DNA damage at telomeres triggers R-loop accumulation in a TERRA and TRF2-dependent manner [8].

Targeting G-quadruplex structures in telomerase positive or ALT cancer cells. Left: unresolved G4s block the function of telomerase for telomere elongation in telomerase positive cells; Middle: stabilized G4s at some oncogene promoters lead to the repression of those oncogenes; Right: Targeting G-loop with G4s ligands or R-loop inhibitors may be a potential therapeutics for ALT cancers
Since Last century cancer chemotherapy gained the success relying on highly cytotoxic drugs, which directly or indirectly disrupt the transcription and/or replication of cellular DNA in both normal cells and tumor cells [15]. To avoid the side effects of these toxic agents, investigators have paid their attentions to design and select the more selective drugs through many strategies. In the past two decades, unique DNA or RNA structures at telomeres or some specific regions of genome have become promising therapeutic targets. The G4 and R-loop structures at telomeres or transcribed regions of genome have been considered as the potential targets for cancer therapy. To date, several strategies that target these specific structures or proteins involving maintaining these structures have been developed by a number of laboratories. Here, we summarize some recent studies that aimed at targeting the secondary structures (e.g. G4 and R-loop) as therapeutic approaches for killing cancer cells.
The distribution of G-quadruplex structures in genome
In 1962, Gellert.et al. first reported the G-tetrad structure in guanylic acid with an X-ray diffraction study [16]. G4 antibodies were made to detect those structures within genomic DNA. Schaffitzel et al. firstly used a high-affinity single-chain antibody to detect quadruplex structure at the telomere of ciliate Stylonychia lemonade [17]. Since then several G4 antibodies have been generated by a few groups to visualize those structures in cells, including Sty49 [18], BG4 [19] and 1H6 [20]. With the combination of next-generation sequencing and genomic mapping, several groups have revealed that the distribution of unique sequences that possess the potential to form stable G4 structure. The human genome contains 376,000 potential G4 structures [21, 22]. The potential G4 structures are highly occurrent in telomeres and promoter regions of oncogenes [23, 24]. Besides telomeres and promoter regions, ribosomal DNA [25], 5′untranslated region of mRNA [26], or TERRA [27] are also potent to form the G4 structures.
Targeting G-quadruplex structures of hTERT
From the genome-wide sequence analyses, potential G4 structures are enriched in promoter regions, which span 1 kb upstream of the transcription start site in humans and other vertebrates genes [24, 28]. Previous studies have paid great attentions to cancer-related genes, e.g. hTERT, c-MYC [29], BLC2 [30], KRAS [31, 32], and VEGF [33], which contain enriched G4 motifs in their promoter regions. A number of G4-targeting ligands have been developed to target the promoters of these genes as potential biomedical targets for antitumor therapy. One major obstacle impeding the clinical application of G4 ligands is the lack of selectivity. Recently, people have entered a new phase of the development of next-generation ligands that interact with G4. The goal is to improve the ligand selectivity to a particular G4 to be targeted, potentially leading to the development of molecules with high antitumor activity and bioactivity with minimal antitumor therapy side effects.
Elevated hTERT expression is observed in ~ 90% of human cancer cells, whereas it is normally silenced in most normal cells. Therefore, hTERT has been considered as the most attractive biomedical target for cancer treatment. Investigators have used two approaches to suppress the function of hTERT: downregulate the expression of hTERT or inhibit the activity of hTERT. Some unselective G4 ligands, such as telomestatin [34, 35] and substituted acridines [36, 37], may bind to a large scale of G4 structures, including hTERT, c-kit, KRAS or c-MYC promoters (Fig. 1 middle). Those unselective ligands may cause bunch of side effects along with their cancer therapeutic effects, which limited their clinic applications. Recently, Hurley and colleagues used a unique approach to address the issue of hTERT downregulation on the basis of the mutations in a G4-mediated manner. They have developed a small molecule (GTC365) that acts at an early step in the G4 folding pathway to redirect mutant promoter G-quadruplex misfolding and reduce hTERT activity through transcriptional repression. They also demonstrate the selectively therapeutic potential of this strategy in melanoma cells that overexpress hTERT [38]. In addition to hTERT, some G4 ligands that more selectivity target to particular G4s at the promoter of specific cancer-related genes (e.g. C-MYC, BLC2, KRAS, and VEGF) have been reported. Tan and colleagues report a new four-leaf-clover-like molecule, IZCZ-3, that have about eightfold preference for the c-MYC over the G4s in the promoters for other genes. More importantly, this ligand showed cytotoxicity against cancer cell lines overexpressing c-MYC but not against normal cells, suggesting reduced side effects based on G4 selectivity on c-MYC [39]. Other ligands, like Furo[2,3-d]pyridazin-4(5H)-one 9 (BLC2) [40], Indolo[3,2-c]quinolines (IQc) (KRAS) [41], and SYUIQ-FM05 (VEGF) [42], has also been reported. Those findings shed a light on the developing of the next-generation G4s ligands, which have high antitumor activity and bioactivity with minimal side effects.
Targeting G-quadruplex structures at telomeres
The regions of eukaryotic genomes with the highest concentration of potential G4 structures are telomeres [21, 23]. Telomeric G4 structures have been considered as attractive anticancer targets for many years. The investigators have successfully developed a large number of compounds that targeting telomere–G-quadruplexes [43] after the first G4s ligands (2,6-diamidoanthraquinone) [44] being reported. In telomerase positive cells, the G-overhang is extended by telomerase, a reverse transcriptase enzyme carrying its own RNA template (Fig. 1 left). G4s ligands bind to G4s tightly and block the telomerase activity through disrupting the base-pairing between G-overhang and telomerase RNA [12, 45, 46].
A lot of efforts were devoted to design more selectable G4s ligands that adopted at telomere 3′ overhang region these years. Some enantiomers, such as Ni–P, exhibit an ability to convert a monomeric antiparallel form to a monomeric hybrid form, and inhibit the cell growth via disputing the localization of TRF2 and POT1 at telomeres [47, 48]. Then, several studies found that a large variety of alternative higher-order structures derived from the canonical telomere G4 might be adopted at the 3′- overhang region. Thus, the unique structure and motif of these ligands are amenable to the gain of specificity for telomere G4s [49,50,51,52].
Telomeric R-loop formation and its relevance with G4 structure
The overabundance of R-loop as shown in a number of neurological syndromes and cancer [53, 54]. The out balance of R-loop leads to genome instability and replication stress, which is a molecular symptom of tumor cells [55]. Therefore, targeting R-loop has been considered as a potential approach to sensitize certain tumors to chemotherapeutic treatment. In 1997, Weaver et al. reported that F8-actinomycin D exhibits a unique selectivity against leukemia cells [56]. However, DNA:RNA hybrids are often formed at transcribed genome, the application of those molecules that directly bind to DNA:RNA hybrids was limited. Recently, researchers turned to pay more attentions on targeting R-loop binding proteins. Andrés Aguilera and colleagues reported that trabectedin and lurbinectedin induced DNA-RNA hybrids-dependent DNA damage in HeLa cells, impairs DNA replication and causes genome instability. The high level of R-loops increases cell sensitivity to those antitumor drugs [57]. Especially, some homologous recombination deficiency cancer cells, which showed the elevated level of R-loop formation, were hypersensitive to genotoxic drugs such as etoposide, camptothecin, trabectedin and PARP inhibitors [58,59,60]. Furthermore, several other compounds have also been reported to increase R-loops, including topoisomerase1 inhibitors [61], spliceosome inhibitors [62, 63], and RNase H2 inhibitors [64, 65].
G4 structures form in a similar genomic context as R-loops. Recent studies indicate the presence of co-existence of R-loop and G4, known as “G-loop”, which is a unique structure where G4 is formed at the displaced single-stranded of an R-loop, both in vivo and in vitro [66]. In 2004, Nancy Maizels and Colleague first described the formation of G-loop in vitro and in Escherichia coli using electron microscopy. Recently, several studies revealed that G4 ligands (PDS, CX-5461) induce R-loop-mediated DNA damage and cell death in cancer cells [67, 68]. Giovanni Capranico et al. used different G4 ligands, including pyridostatin, Braco-19, and FG, to stabilize G4 structures and simultaneously increase R-loop levels within minutes in human cancer cells. The increased level of R-loop leads to the accumulation of γH2AX foci and of G2/M cells, which are both hallmarks of genomic DSBs and DNA damage response. Importantly, overexpression of an exogenous human RNaseH1 rescued DNA damage induced by G4 ligands in BRCA2-depleted cancer cells, which indicated the G4 ligands can induce DNA damage by an R loop-dependent mechanism [67].
Is the telomeric R-loop structure a potential target for cancer therapy?
R-loops play an import role in telomere maintenance in telomerase negative cancer cells. In ALT cancer cells, R-loop facilitated telomere elongation through a recombination mechanism. Arora et al. first showed that the RNA endonuclease RNaseH1 regulates the levels of RNA–DNA hybrids between telomeric DNA and the long noncoding RNA TERRA, and is a key mediator of telomere maintenance in ALT cells. Then, several studies also confirm the function of R-loop in telomere maintenance [69, 70]. Our group recently reported that the R-loop-CSB-RAD52-POLD3 axis contributes to the repair of Reactive oxygen species (ROS) induced telomeric damage in ALT cancer [8]. Target R-loop interacting proteins at genome are also possible to enhance the cell killing effects. Therefore, targeting telomeric R-loop could be the potential treatments for ALT tumors, with more studies revealed the molecular details of ALT and the mechanisms involved in its engagement (Fig. 1 right).
There are a lot of reasons that the application of telomerase inhibitors in cancer therapy has not got too much progress. Although the non-selectivity of those inhibitors may the major limitation for their application, other reasons cannot be excluded. Two of those reasons are that the probability co-existence of telomerase and ALT pathway in some specific tumor [71,72,73,74], and ALT pathway may be activated after suppressing telomerase by inhibitors [75]. The information about the activation of ALT pathway in telomerase-positive cancer cells that treated with telomerase inhibitors is limited. Combined strategies that target both telomerase and ALT could be proved to be a powerful approach for the treatment of such tumors.
Conclusions
Both G4 and R-loops structures are potential targets for cancer therapy, however, questions including how to improve the selectivity of drugs, how to reduce the resistance and side effects, are the major obstacles for their future application. Actually, several clinical trials of G4 ligands were withdrawn due to these reasons. More ligands that have higher affinities to spatial conformations of G4 structures within unique regions of genomes are expected in the future.
Abbreviations
- G4:
-
G-quadruplexes
- ALT:
-
Alternative Lengthening of Telomeres
- DSBs:
-
Double Strands Breaks
- ROS:
-
Reactive oxygen species
References
Blackburn EH. Telomere states and cell fates. Nature. 2000;408:53.
Levy MZ, Allsopp RC, Futcher AB, Greider CW, Harley CB. Telomere end-replication problem and cell aging. J Mol Biol. 1992;225:951–60.
Dalgård C, et al. Leukocyte telomere length dynamics in women and men: menopause vs age effects. Int J Epidemiol. 2015;44:1688–95.
Maestroni L, Matmati S, Coulon S. Solving the telomere replication problem. Genes. 2017;8:55.
Verdun RE, Karlseder J. Replication and protection of telomeres. Nature. 2007;447:924.
Meyne J, Ratliff RL, MoYzIs RK. Conservation of the human telomere sequence (TTAGGG) n among vertebrates. Proc Natl Acad Sci. 1989;86:7049–53.
Moyzis RK, et al. A highly conserved repetitive DNA sequence,(TTAGGG) n, present at the telomeres of human chromosomes. Proc Natl Acad Sci. 1988;85:6622–6.
Tan J, et al. An R-loop-initiated CSB-RAD52-POLD3 pathway suppresses ROS-induced telomeric DNA breaks. Nucleic Acids Res. 2019. https://doi.org/10.1093/nar/gkz1114.
Wang RC, Smogorzewska A, De Lange T. Homologous recombination generates T-loop-sized deletions at human telomeres. Cell. 2004;119:355–68.
Stansel RM, de Lange T, Griffith JD. T-loop assembly in vitro involves binding of TRF2 near the 3′ telomeric overhang. EMBO J. 2001;20:5532–40.
Greider CW. Telomeres do D-loop–T-loop. Cell. 1999;97:419–22.
De Cian A, et al. Reevaluation of telomerase inhibition by quadruplex ligands and their mechanisms of action. Proc Natl Acad Sci. 2007;104:17347–52.
Mao S-Q, et al. DNA G-quadruplex structures mold the DNA methylome. Nat Struct Mol Biol. 2018;25:951.
Bochman ML, Paeschke K, Zakian VA. DNA secondary structures: stability and function of G-quadruplex structures. Nat Rev Genet. 2012;13:770.
Kaufmann SH, Earnshaw WC. Induction of apoptosis by cancer chemotherapy. Exp Cell Res. 2000;256:42–9.
Gellert M, Lipsett MN, Davies DR. Helix formation by guanylic acid. Proc Natl Acad Sci USA. 1962;48:2013.
Schaffitzel C, et al. In vitro generated antibodies specific for telomeric guanine-quadruplex DNA react with Stylonychia lemnae macronuclei. Proc Natl Acad Sci. 2001;98:8572–7.
Oganesian L, Bryan TM. Physiological relevance of telomeric G-quadruplex formation: a potential drug target. BioEssays. 2007;29:155–65.
Biffi G, Tannahill D, McCafferty J, Balasubramanian S. Quantitative visualization of DNA G-quadruplex structures in human cells. Nat Chem. 2013;5:182.
Henderson A, et al. Detection of G-quadruplex DNA in mammalian cells. Nucleic Acids Res. 2013;42:860–9.
Todd AK, Johnston M, Neidle S. Highly prevalent putative quadruplex sequence motifs in human DNA. Nucleic Acids Res. 2005;33:2901–7.
Hänsel-Hertsch R, Spiegel J, Marsico G, Tannahill D, Balasubramanian S. Genome-wide mapping of endogenous G-quadruplex DNA structures by chromatin immunoprecipitation and high-throughput sequencing. Nat Protoc. 2018;13:551.
Maizels N. Dynamic roles for G4 DNA in the biology of eukaryotic cells. Nat Struct Mol Biol. 2006;13:1055.
Huppert JL, Balasubramanian S. G-quadruplexes in promoters throughout the human genome. Nucleic Acids Res. 2006;35:406–13.
Wallgren M, et al. G-rich telomeric and ribosomal DNA sequences from the fission yeast genome form stable G-quadruplex DNA structures in vitro and are unwound by the Pfh1 DNA helicase. Nucleic Acids Res. 2016;44:6213–31.
Kumari S, Bugaut A, Huppert JL, Balasubramanian S. An RNA G-quadruplex in the 5′ UTR of the NRAS proto-oncogene modulates translation. Nat Chem Biol. 2007;3:218.
Xiao C-D, Shibata T, Yamamoto Y, Xu Y. An intramolecular antiparallel G-quadruplex formed by human telomere RNA. Chem Commun. 2018;54:3944–6.
Todd AK, Haider SM, Parkinson GN, Neidle S. Sequence occurrence and structural uniqueness of a G-quadruplex in the human c-kit promoter. Nucleic Acids Res. 2007;35:5799–808.
Siddiqui-Jain A, Grand CL, Bearss DJ, Hurley LH. Direct evidence for a G-quadruplex in a promoter region and its targeting with a small molecule to repress c-MYC transcription. Proc Natl Acad Sci. 2002;99:11593–8.
Onel B, et al. A new G-quadruplex with hairpin loop immediately upstream of the human BCL2 P1 promoter modulates transcription. J Am Chem Soc. 2016;138:2563–70.
Cogoi S, Paramasivam M, Membrino A, Yokoyama KK, Xodo LE. The KRAS promoter responds to Myc-associated zinc finger and poly (ADP-ribose) polymerase 1 proteins, which recognize a critical quadruplex-forming GA-element. J Biol Chem. 2010;285:22003–16.
Cogoi S, Paramasivam M, Spolaore B, Xodo LE. Structural polymorphism within a regulatory element of the human KRAS promoter: formation of G4-DNA recognized by nuclear proteins. Nucleic Acids Res. 2008;36:3765–80.
Sun D, Guo K, Rusche JJ, Hurley LH. Facilitation of a structural transition in the polypurine/polypyrimidine tract within the proximal promoter region of the human VEGF gene by the presence of potassium and G-quadruplex-interactive agents. Nucleic Acids Res. 2005;33:6070–80.
Tauchi T, et al. Telomerase inhibition with a novel G-quadruplex-interactive agent, telomestatin: in vitro and in vivo studies in acute leukemia. Oncogene. 2006;25:5719.
Saha D, et al. Epigenetic suppression of human telomerase (hTERT) is mediated by the metastasis suppressor NME2 in a G-quadruplex–dependent fashion. J Biol Chem. 2017;292:15205–15.
Carvalho J, et al. Fluorescent light-up acridine orange derivatives bind and stabilize KRAS-22RT G-quadruplex. Biochimie. 2018;144:144–52.
Guo Q-L, et al. Synthesis and evaluation of 7-substituted-5, 6-dihydrobenzo [c] acridine derivatives as new c-KIT promoter G-quadruplex binding ligands. Eur J Med Chem. 2017;130:458–71.
Kang H-J, et al. A pharmacological chaperone molecule induces cancer cell death by restoring tertiary DNA structures in mutant hTERT promoters. J Am Chem Soc. 2016;138:13673–92.
Hu M-H, et al. Discovery of a new four-leaf clover-like ligand as a potent c-MYC transcription inhibitor specifically targeting the promoter G-quadruplex. J Med Chem. 2018;61:2447–59.
Amato J, et al. Targeting the BCL2 Gene Promoter G-Quadruplex with a New Class of Furopyridazinone-Based Molecules. ChemMedChem. 2018;13:406–10.
Lavrado J, et al. KRAS oncogene repression in colon cancer cell lines by G-quadruplex binding indolo [3, 2-c] quinolines. Sci Rep. 2015;5:9696.
Wu Y, et al. Stabilization of VEGF G-quadruplex and inhibition of angiogenesis by quindoline derivatives. Biochimica et Biophysica Acta (BBA)-General Subjects. 2014;1840:2970–7.
Islam MK, Jackson PJ, Rahman KM, Thurston DE. Recent advances in targeting the telomeric G-quadruplex DNA sequence with small molecules as a strategy for anticancer therapies. Future Med Chem. 2016;8:1259–90.
Sun D, et al. Inhibition of human telomerase by a G-quadruplex-interactive compound. J Med Chem. 1997;40:2113–6.
Zahler AM, Williamson JR, Cech TR, Prescott DM. Inhibition of telomerase by G-quartet DMA structures. Nature. 1991;350:718.
Cuesta J, Read MA, Neidle S. The design of G-quadruplex ligands as telomerase inhibitors. Mini Rev Med Chem. 2003;3:11–21.
Yu H, Wang X, Fu M, Ren J, Qu X. Chiral metallo-supramolecular complexes selectively recognize human telomeric G-quadruplex DNA. Nucleic Acids Res. 2008;36:5695–703.
Qin H, Zhao C, Sun Y, Ren J, Qu X. Metallo-supramolecular complexes enantioselectively eradicate cancer stem cells in vivo. J Am Chem Soc. 2017;139:16201–9.
Mukundan VT, Phan AT. Bulges in G-quadruplexes: broadening the definition of G-quadruplex-forming sequences. J Am Chem Soc. 2013;135:5017–28.
Yue DJE, Lim KW, Phan AT. Formation of (3 + 1) G-quadruplexes with a long loop by human telomeric DNA spanning five or more repeats. J Am Chem Soc. 2011;133:11462–5.
Abraham Punnoose J, et al. Interaction of G-quadruplexes in the full-length 3′ human telomeric overhang. J Am Chem Soc. 2014;136:18062–9.
Russo Krauss I, Ramaswamy S, Neidle S, Haider S, Parkinson GN. Structural insights into the quadruplex–duplex 3′ interface formed from a telomeric repeat: a potential molecular target. J Am Chem Soc. 2016;138:1226–33.
Richard P, Manley JL. R loops and links to human disease. J Mol Biol. 2017;429:3168–80.
Skourti-Stathaki K, Proudfoot NJ. A double-edged sword: R loops as threats to genome integrity and powerful regulators of gene expression. Genes Dev. 2014;28:1384–96.
Halazonetis TD, Gorgoulis VG, Bartek J. An oncogene-induced DNA damage model for cancer development. Science. 2008;319:1352–5.
Takusagawa F, Takusagawa KT, Carlson RG, Weaver RF. Selectivity of F8-actinomycin D for RNA: DNA hybrids and its anti-leukemia activity. Bioorg Med Chem. 1997;5:1197–207.
Tumini E, et al. The antitumor drugs trabectedin and lurbinectedin induce transcription-dependent replication stress and genome instability. Mol Cancer Res. 2019;17:773–82.
Valeri A, Martínez S, Casado JA, Bueren JA. Fanconi anaemia: from a monogenic disease to sporadic cancer. Clin Transl Oncol. 2011;13:215–21.
Monk BJ, et al. Trabectedin as a chemotherapy option for patients with BRCA deficiency. Cancer Treat Rev. 2016;50:175–82.
Gorthi A, et al. EWS–FLI1 increases transcription to cause R-loops and block BRCA1 repair in Ewing sarcoma. Nature. 2018;555:387.
Marinello J, et al. Dynamic effects of topoisomerase I inhibition on R-loops and short transcripts at active promoters. PLoS ONE. 2016;11:e0147053.
Wan Y, et al. Splicing function of mitotic regulators links R-loop–mediated DNA damage to tumor cell killing. J Cell Biol. 2015;209:235–46.
Chen L, et al. The augmented R-loop is a unifying mechanism for myelodysplastic syndromes induced by high-risk splicing factor mutations. Mol Cell. 2018;69:412–25.
Flanagan JM, et al. Genomics screen in transformed stem cells reveals RNASEH2A, PPAP2C, and ADARB1 as putative anticancer drug targets. Mol Cancer Ther. 2009;8:249–60.
White R, Saxty B, Large J, Kettleborough CA, Jackson AP. Identification of small-molecule inhibitors of the ribonuclease H2 enzyme. J Biomol Screen. 2013;18:610–20.
Tan J, Wang X, Phoon L, Yang H, Lan L. Resolution of ROS-induced G-quadruplexes and R-loops at transcriptionally active sites is dependent on BLM helicase. FEBS Lett. 2020. https://doi.org/10.1002/1873-3468.13738.
De Magis A, et al. DNA damage and genome instability by G-quadruplex ligands are mediated by R loops in human cancer cells. Proc Natl Acad Sci. 2019;116:816–25.
Xu H, et al. CX-5461 is a DNA G-quadruplex stabilizer with selective lethality in BRCA1/2 deficient tumours. Nature Communications. 2017;8:14432.
Graf M, et al. Telomere length determines TERRA and R-loop regulation through the cell cycle. Cell. 2017;170:72–85.
Balk B, et al. Telomeric RNA-DNA hybrids affect telomere-length dynamics and senescence. Nat Struct Mol Biol. 2013;20:1199.
De Vitis M, Berardinelli F, Sgura A. Telomere length maintenance in cancer: at the crossroad between telomerase and alternative lengthening of telomeres (ALT). Int J Mol Sci. 2018;19:606.
Cerone MA, Londono-Vallejo JA, Bacchetti S. Telomere maintenance by telomerase and by recombination can coexist in human cells. Hum Mol Genet. 2001;10:1945–52.
Perrem K, Colgin LM, Neumann AA, Yeager TR, Reddel RR. Coexistence of alternative lengthening of telomeres and telomerase in hTERT-transfected GM847 cells. Mol Cell Biol. 2001;21:3862–75.
Grobelny JV, Kulp-McEliece M, Broccoli D. Effects of reconstitution of telomerase activity on telomere maintenance by the alternative lengthening of telomeres (ALT) pathway. Hum Mol Genet. 2001;10:1953–61.
Hu J, et al. Antitelomerase therapy provokes ALT and mitochondrial adaptive mechanisms in cancer. Cell. 2012;148:651–63.
Acknowledgements
We thank the members of Zou, Lan, and Dyson laboratories for discussions.
Funding
L.L. was supported in part by Grant from NIH GM118833.
Author information
Authors and Affiliations
Contributions
JT and LL wrote the review. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declares that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Tan, J., Lan, L. The DNA secondary structures at telomeres and genome instability. Cell Biosci 10, 47 (2020). https://doi.org/10.1186/s13578-020-00409-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13578-020-00409-z