
Abstract
Background
As a primary source of Shiga-toxin-producing Escherichia coli (STEC) infection, cattle are often targeted to develop strategies for reducing STEC contamination. Monitoring the virulence potentials of STEC isolates from cattle is important for tracing contamination sources, managing outbreaks or sporadic cases, and reducing the risks for human infection. This study aimed to investigate the prevalence of STEC in cattle farm samples in South Korea and to assess their virulence potentials.
Results
In total, 63 STEC were isolated from 496 cattle farm samples, and temperature and rainfall affected STEC prevalence (p < 0.001). The O157 serogroup was most prevalent, followed by O108, O8, O84, O15, and O119. In the stx variant test, high prevalence of stx2a and stx2c (known to be associated with high STEC virulence) were observed, and stx2g, a bovine STEC variant, was detected in STEC O15 and O109. Additionally, stx1c was detected in eae-positive STEC, suggesting genetic dynamics among the virulence genes in the STEC isolates. STEC non-O157 strains were resistant to tetracycline (17.9%), ampicillin (14.3%), and cefotaxime (3.6%), while STEC O157 was susceptible to all tested antimicrobials, except cefotaxime. The antimicrobial resistance genes, bla TEM (17.5%), tetB (6.3%), and tetC (4.8%), were only detected in STEC non-O157, whereas tetE (54.0%) was detected in STEC O157. AmpC was detected in all STEC isolates. Clustering was performed based on the virulence gene profiles, which grouped STEC O84, O108, O111, and O157 together as potentially pathogenic STEC strains. Finally, PFGE suggested the presence of a prototype STEC that continues to evolve by genetic mutation and causes within- and between-farm transmission within the Gyeonggi province.
Conclusions
Considerable numbers of STEC non-O157 were isolated from cattle farms, and the virulence and antimicrobial resistance features were different between the STEC O157 and non-O157 strains. STEC from cattle with virulence or antimicrobial resistance genes might represent a threat to public health and therefore, continual surveillance of both STEC O157 and non-O157 would be beneficial for controlling and preventing STEC-related illness.
Similar content being viewed by others

Background
Since the identification of STEC O157:H7 as a foodborne zoonotic disease in 1982 [1], human infections by STEC have been reported worldwide [2, 3]. While numerous studies have focused on STEC O157:H7, the most well-known and notorious serotype, >400 serotypes of STEC non-O157 have been implicated as etiological agents of several outbreaks and in sporadic cases of STEC infection [4]. Recently, STEC non-O157 infection cases have increased globally, highlighting the significance of investigating STEC non-O157 [3, 5, 6]. Among the STEC non-O157 serotypes, O26, O45, O103, O111, O121, and O145, were reported as the six major STEC non-O157 linked to human diseases [7, 8]. Scallen et al. reported that ~63,000 and 112,000 cases of foodborne illness caused by STEC O157 and non-O157, respectively, occur in the United States annually [2]. The progression of STEC infection varies, causing symptoms ranging from mild gastrointestinal symptoms to severe hemorrhagic colitis (HC) or hemolytic uremic syndrome (HUS) [9–11]. Predicting the risk of STEC is especially important for public health because STEC infection might develop into a life-threatening disease, and is often associated with large and multinational outbreaks [10, 12, 13].
Although the pathogenicity of STEC is not fully understood, several virulence factors have been identified [10], including Shiga toxins, intimin, and the 60-MDa plasmids (enterohemolysin or serine protease) [10]. Shiga toxins are the principal virulence factors of STEC, and two major types of Shiga toxins are known, Stx1 and 2 [14]. While the DNA sequence of stx1 is highly conserved and only a few stx1 variants have been reported (including stx1c and stx1d), the stx2 sequence shows 84–99% similarity among the stx 2 variants [10, 15]. Because the variants are related to the properties of Shiga toxin, subtyping of the stx variants is important for predicting the virulence potential of STEC in human infection [16]. Among Shiga toxin and its variants, Stx2 is most associated with severe disease [17, 18]. Stx2 is a 1000 times more toxic than Stx1 to renal microvascular endothelial cells, and Stx2 and Stx2c are more commonly reported in HUS patients [19–21]. Intimin, one of the proteins encoded by eae in the locus of enterocyte effacement, which is responsible for the formation of attaching and effacing (A/E) legions [10, 22, 23]. Several other factors also contribute to the virulence of STEC. EhxA (EHEC-enterohemolysin) disrupts the cytoplasmic membranes of mammalian cells [10, 24]. EspP (a serine protease) potentiates STEC colonization in the human gut [25, 26], and KatP (catalase peroxidase) [27], SubAB (subtilase), and Saa (STEC autoagglutinating adhesin) are associated with the virulence of STEC [14, 28, 29].
Cattle are a primary source of STEC infection and are often targeted to develop strategies for reducing contamination. Therefore, monitoring the virulence potentials of STEC isolates from cattle is important for tracing the sources of contamination, managing outbreaks or sporadic cases, and reducing the risks for human infection. This study investigated the prevalence of STEC O157 and non-O157 in cattle farm samples in South Korea and assessed the virulence potentials of STEC isolates from these samples by characterizing stx variants, antimicrobial resistance, and virulence genes. Finally, genetic analysis was performed to analyze the genetic dynamics of STEC strains isolated over a 4-year period.
Methods
Sample collection
Samples were collected from 15 cattle farms located in the Gyeonggi province in Korea during 2012–2015. Each farm was visited one to nine times during the sampling period (median = 1, average = 1.9), and cattle farm samples, including feces, ground soil, and water, were collected. Fecal samples were collected by direct rectal retrieval using disposable gloves. Environmental samples in the farm were collected using sterilized spatulas. Each sample collected had a mass of at least 5 g (or a volume of at least 5 mL for liquid samples). A total of 469 samples (419 fecal, 47 ground soil, one water, one raw milk, and one forage sample) were collected and transported immediately to the laboratory for STEC isolation.
Isolation of STEC strains
Shiga-toxin-producing Escherichia coli O157 was isolated using a combination of standard culture methods and immunomagnetic separation methods, as described previously [30]. Briefly, 1 g or 1 mL of each sample was homogenized for 1 min with 9 mL of modified EC broth (Becton, Dickinson and Company, USA) supplemented with novobiocin (20 mg/L, Oxoid, USA) using a homogenizer, and then incubated overnight at 37 °C. The enriched culture suspension was then mixed with anti-E. coli O157 antibody-coated magnetic beads (Dynal, Invitrogen, USA) and separated using a magnetic rack, as described in the manufacturer’s guidelines. The suspension was then tested for the formation of typical colonies in tellurite-added sorbitol MacConkey agar (T-SMAC; BD, USA), MacConkey agar (MAC; BD, USA), and CHROMagar O157 (CHROMagar Microbiology, France). To confirm STEC O157 isolation, the serotype of the colonies was tested using the E. coli O157 Latex Test Kit (Oxoid, UK).
To isolate STEC non-O157, multiplex polymerase chain reaction (PCR) combined with standard culture methods was used, as described previously [30]. Briefly, one loop of enriched culture was streaked onto T-SMAC and tested for the presence of Shiga toxin genes by PCR as described below in the virulence gene profiling section. The serotype of colonies harboring stx genes was then tested by conventional agglutination tests, using E. coli antisera (JoongKyeom, Ansan, Korea).
Antimicrobial-susceptibility test
A standard disk-diffusion test was performed to determine antimicrobial susceptibility for the following 14 antimicrobial drugs: ampicillin (AM, 10 µg), chloramphenicol (C, 30 µg), imipenem (IMP, 10 µg), tetracycline (TE, 30 µg), amikacin (AN, 30 µg), amoxicillin–clavulanic acid (AMC, 20/10 µg), ceftazidime (CAZ, 30 µg), gentamicin (GM, 10 µg), nalidixic acid (NA, 30 µg), trimethoprim–sulfamethoxazole (STX, 1.25/23.75 µg), ceftriaxone (CRO, 30 µg), aztreonam (ATM, 30 µg), cefotaxime (CTX, 30 µg), and cefpodoxime (CPD, 10 µg). For quality control, E. coli ATCC 25922 was used as the reference strain. Antimicrobial susceptibility was interpreted as guided by the Clinical Laboratory Standard Institute (CLSI).
ESBL-producing STEC was detected following the CLSI ESBL phenotypic confirmatory test, which is a disk-diffusion test. Briefly, STEC isolates found to be resistant to cefpodoxime, ceftazidime, aztreonam, cefotaxime, or ceftriaxone were screened in the phenotypic confirmatory test. A standard double-disk test was performed to confirm the ESBL phenotype, as described in the NCCL guidelines. Briefly, enriched STEC isolates were adjusted to 0.5 McFarland scale (McF), and 100 µL of the suspension was spread onto Mueller–Hinton agar. Antimicrobial disks containing ceftazidime + ceftazidime–clavulanic acid or cefotaxime + cefotaxime–clavulanic acid were placed on STEC-inoculated MH agar and incubated at 35 °C for 18 h. The ESBL phenotype was considered positive when the zone diameter resulting from the combination of the antimicrobial agent and clavulanic acid was > 5 mm larger than that obtained using the antimicrobial test agent alone.
Detection of stx variants, virulence genes, and antimicrobial resistance genes
The presence of virulence and antimicrobial resistance genes was determined by PCR using a MyCycler thermal cycler (Bio-Rad Laboratories, USA). For DNA preparation, a single colony of each isolate was suspended in 1 mL of normal saline and centrifuged for 3 min at 6000×g. The pellets were re-suspended with 200 µL of sterile water and boiled for 10 min. The suspension was centrifuged for 3 min at 6000×g, and the supernatant was used as the DNA template. PCR was conducted as described previously to detect Shiga toxin genes (stx1, stx1c, stx1d, stx2, stx2a, stx2c, stx2d, stx2e, stx2f, and stx2 g), virulence genes (eae, tir, espB, espD, ehxA, katP, espP, iha, subA, stcE, and saa), and antimicrobial resistance genes (ampC, tetA, tetB, tetC, tetD, tetE, tetG, cat, cml, bla OXA , bla CMY , bla TEM , and qnr). The primer sequences and reaction conditions for each gene are summarized in Table 1.
Virulence gene profiling
A phylogenetic dendrogram of the virulence profiles was constructed by using the unweighted pair group method with arithmetic mean analysis (UPGMA) for binary data using BioNumerics, version 6.6 (Applied Maths NV, Belgium).
Pulsed-field gel electrophoresis (PFGE)
Pulsed-field gel electrophoresis was performed following the CDC PulseNet protocol using CHEF MAPPER (Bio-Rad, Hercules, CA, USA). Briefly, STEC colonies were suspended in cell suspension buffer (100 mM Tris: 100 mM EDTA, pH 8.0) and then adjusted to the 4.0 McFarland scale (McF). The adjusted cell suspension (400 µL) was mixed gently with 20 µL of proteinase K (20 mg/mL) and 400 µL of 1% SeaKem Gold melted agarose gel to build a plug. The plug was soaked in a proteinase K-containing cell lysis buffer (50 mM Tris: 50 mM EDTA, pH 8.0 + 1% sarcosyl) for 2 h to lyse the cells, after which it was washed twice with sterile water for 15 min and then four times with TE buffer (10 mM Tris: 1 mM EDTA, pH 8.0) for 15 min. The plug was then digested with 50 U of XbaI restriction enzyme for 2 h. PFGE was performed with a pulse time of 2.16–54.17 s for STEC O157 and 6.76–35.38 s for STEC non-O157; S. Braenderup ATCC BAA664 was used as a size ladder marker. A Dice similarity coefficient with a UPGMA dendrogram was generated based on 1.5% tolerance windows and 1.5% optimization, using BioNumerics, version 6.6 (Applied Maths NV, Belgium).
Statistical analysis
To identify factors potentially associated with prevalence, farm-management factors, environmental factors, and animal information were collected, if present (Table 2). Farm-management factors (farm size, ground soil hygiene, and diet), and animal information (age and breed) were obtained from the veterinarian in charge of each farm. Environmental factors included average temperature on sampling date, humidity, and rainfall within 3 days prior to sampling, which were obtained from the data provided by the Meteorological Administration (http://www.kma.go.kr/weather/observation/past_table.jsp). The association between STEC prevalence and farm management and environmental factors was analyzed by using the Chi squared test, and the association between STEC prevalence and animal factors was analyzed by Fisher’s exact test. The statistical analysis was performed using SPSS, version 22.0 (SPSS IBM, New York, NY, USA), and the variables were considered to be significantly associated when the p value was <0.05.
Results
STEC prevalence in cattle farms
Of 496 samples collected from 29 visits to 15 farms, 63 STEC were isolated from 17 visits to seven farms (Table 2). Most STEC were isolated from feces (54 from adult cow and three from calves), and six STEC were isolated from ground soil. No STEC were isolated from feed, water, or milk samples. The farm prevalence of STEC varied from 0.0 to 90.0%, but no farm management factors were found to be associated with STEC prevalence. STEC prevalence was significantly higher when the average temperature was above 20 °C (p < 0.001, two tailed Chi squared test, odds ratio [OR] = 2.3), and when rain was reported within 3 days prior to sampling (p < 0.001, two tailed Chi squared test, OR = 3.5). Moreover, while the STEC prevalence was higher in calves (3/19; 15.8%) than in adults (54/405; 13.3%), the difference was not significant. By breed, STEC prevalence was significantly higher in beef cattle (4/9; 44.4%) than in dairy cattle (50/396; 12.63%; p < 0.05, two-tailed Fisher’s exact test, OR = 5.5).
Serogroup
Of the 63 STEC isolates, 61 were serogrouped. Thirty-five (55.6%) and 26 (41.3%) isolates were found to be STEC O157 and non-O157, respectively, whereas two isolates were non-typeable (NT). Among STEC non-O157, the most common serogroup was O108 (five isolates, 7.5%), followed by O8 and O84 (four isolates each, 6.0%), O15 and O119 (three isolates each, 4.5%), O109 and O111 (two isolates each, 3.0%), and O55, O169, and O185 (one isolate each, 1.5%).
Prevalence of Shiga toxin and its variant genes
The Shiga toxin gene types of the 63 STEC isolates were determined by PCR. STEC harboring both stx1 and stx2 (28/63, 41.8%) were the most common strains, followed by STEC with either stx1 (13/63, 19.4%) or stx2 (22/63, 32.8%; Table 3). STEC harboring stx1 were only detected in STEC non-O157 (O55, O84, O108, and O111), while STEC O157 harbored stx2 only or both stx1 and stx2 with a similar distribution. Of the 35 STEC harboring stx1, 34 (97.1%) carried the stx1c variant, while no stx1d variants were detected. All STEC harboring stx2 (n = 50) carried at least one stx2 variant, stx2a (45; 90.0%), stx2c (34; 68.0%), and stx2 g (5; 10.0%). Stx2d, stx2e, and stx2f were not detected. When the distribution of the stx variants was analyzed based on serotypes, stx2a was observed in O8, O119, O157, O169, and O185; stx2c in O157 (33/35, 94.3%), and O185 (1/1, 100.0%); and stx2 g in O15 (3/3, 100.0%) and O109 (2/2, 100.0%).
Antimicrobial resistance of the STEC isolates
Disc diffusion tests were conducted using 14 antimicrobial agents. While resistance was observed for TE (five isolates, 17.9%), AMP (four isolates, 14.3%), and CTX (one isolate, 3.6%) in 28 STEC non-O157 strains, resistance was not observed in 35 STEC O157 strains. Four and five STEC O157 and non-O157, respectively, showed intermediate resistance to CTX. The STEC isolates showing complete or intermediate resistance to CTX were further tested for ESBL-producing STEC, but none of them had the ESBL phenotype (Table 4).
All of the 63 STEC isolates carried more than one antimicrobial resistance genes. Of 13 tested antimicrobial resistance genes, genes tetA, tetD, tetG, cat, cml, bla OXA , bla CMY , and qnr were not detected. All STEC isolates carried ampC gene (63/63; 100.0%), and 11 isolates carried bla TEM , (11/63; 17.5%) gene. Of tetracycline resistance gene, tetE (34/63; 54.0%) was the most prevalent, followed by tetB (4/63; 6.3%) and tetC (3/63; 4.8%) (Table 4).
Virulence gene profiles
The prevalence of virulence genes was investigated. The prevalence of each virulence gene was as follows: eae (77.8%), tir (76.2%), espB (74.6%), espD (65.1%), ehxA (92.1%), katP (55.6%), espP (84.1%), iha (66.7%), subA (7.9%), stcE (65.1%), and saa (9.5%).
When clustering analysis was conducted based on the profiles of the virulence genes, including Shiga toxin and its variant genes, six clusters were generated (based on 50% similarity; Fig. 1). STEC strains in clusters 1, 2, and 6 were composed of a single serotype each, O119, O185, and O55, respectively. The strains showed 100% prevalence of eae, ehxA, stx2, and stx2c in cluster 1; espP, stx2, stx2a, and stx2c in Cluster 2; and tir, stx1, and stx1c in cluster 3. Strains in cluster 3 were characterized as having stx2 g variants, and belonged to the O15 and O109 serogroups. Cluster 4 was composed of STEC O84, O108, O111, and O157 strains with a high prevalence of eae, tir, espB, espD, ehxA, katP, espP, iha, stcE, stx2, stx2a, and stx2c. Strains in cluster 5 were characterized as having subA and saa (Table 5).

UPGMA dendrogram of STEC O157 and non-O157 based on the profiles of virulence genes. Six clusters were generated with the UPGMA method based on 50% similarity. a0806-3, 0806-5, 0806-20, 0823-4, 0823-5, 0823-8, 0909-5, 0909-9, 0909-11, 0909-14, 0909-15, 0909-16, 0909-17 (O157). b0827-1, 0827-2, 0827-3, 0827-5, 0827-6, 0827-7, 0827-8, 0827-9, 0827-10, 0814-4, 0814-5, 0814-7, 0814-8, 0814-11, 0814-20, 0814-31, 1013-19 (O157)
Genetic relatedness of the STEC isolates
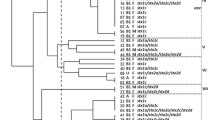
The PFGE patterns of XbaI-digested STEC O157 and non-O157 were analyzed to determine how the clonal relatedness of STEC isolates changed with temporo-spatial variation. The DNA fingerprints of 35 STEC O157 strains showed high similarity, generating 16 different PFGE profiles with 15–20 discernible fragments (mean: 17.4, median: 17). The 16 PFGE profiles clustered into four groups, based on 90% similarity; each group was composed of one to 23 isolates (Fig. 2a).

Phylogenetic analysis of STEC isolates from cattle. The UPGMA method was used with a 1.5% optimization and 1.5% tolerance window using Bionumerics software. a CP Cheongpyeong, GP Gapyeong, YP Yangpyeong. bAll STEC O157 strains were belonged to cluster 4. c AMP ampicillin, CTX cefotaxime, TE tetracycline. a STEC O157 strains were clustered into four groups (based on 90% similarity). b STEC non-O157 strains clustered into 12 groups (based on 80% similarity), STEC strains with the same serotype clustered together, except for STEC O8. The STEC O169 strains were not typeable by PFGE
Pulsed-field gel electrophoresis was performed for STEC strains belonging to 10 different serogroups, other than O157 and two strains that were non-typeable for the O serotype. Diverse PFGE patterns of XbaI-digested STEC non-O157 were observed, resulting in 20 different PFGE profiles with 14–23 discernible fragments (mean: 18.2, median: 19), except for one isolate (0911-15), which was untypeable by PFGE. A UPGMA dendrogram, generated using Dice coefficient analysis, clustered STEC non-O157 into 12 different groups, based on 80% similarity. Each group was composed of one to five strains, and STEC strains belonging to the same serotype were grouped together regardless of differences in the sampling period or geographical location of each farm (Fig. 2b).
Discussion
The prevalence of STEC in 15 different cattle farms, virulence-gene profiles, antimicrobial resistance, and genetic relatedness of STEC isolates were analyzed to investigate the virulence potentials of STEC in cattle farm.
During the sampling period, 63 STEC were isolated from 469 cattle farm samples collected from 15 cattle farm in Gyeonggi province in South Korea. Numerous studies are ongoing to identify the factors associated with STEC prevalence. In this study, high temperature and rain were found to be associated with STEC prevalence. Similarly, a previous study reported higher STEC prevalence in hot seasons than in cold seasons [31–33]. In addition, rainfall has been considered an important transmission factor for STEC. The pathogens may be transported via sediments to vast geographical regions as far away as 32 km, resulting in an increased prevalence in the environment [34, 35]. Many published reports have shown that STEC O157 prevalence is higher in calves, especially in post-weaned calves, than in adult cattle [36–38]. However, no obvious link between age and STEC non-O157 prevalence has been reported, and some investigators even observed a higher prevalence of STEC non-O157 in adult groups [37, 39, 40]. In this study, the adult group showed a higher prevalence for STEC O157 (calves: 0/19, 0.0% vs. adults: 31/405, 7.7%) and a lower prevalence for STEC non-O157 (calves: 3/19, 15.8% vs. adult: 24/405, 5.9%). This discrepancy with respect to previous data may be due to the collection of a relatively small number of calf feces samples, compared to the number of samples collected from adults. Thus, further studies may be needed to investigate the effect of age on STEC non-O157 prevalence. Here, beef cattle showed higher STEC prevalence than dairy cattle. Although only a few beef cattle were included in this study, the results are consistent with data from previous studies [31, 33].
While many studies have focused on the O157 serotype, the significance of STEC non-O157 in human infection has become clear recently [3, 7]. In this study, 11 different serotypes of STEC were identified and >40% of the STEC were non-O157, highlighting the need for active surveillance of STEC non-O157 and understanding their virulence potential in humans. Of the identified serotypes, O8, O15, O55, O84, O109, O111, O119, and O157 have been reported frequently in dairy cattle worldwide [39]. Among them, several serogroups have also been reported frequently in human clinical cases. The O111 serogroup is the second most common serogroup in human infections, and is the most common cause of HUS. Moreover, it accounts for half the STEC non-O157 outbreaks. The O15, O84, and O119 serogroups also frequently cause human illness [7, 39]. In addition, human-pathogenic STEC O8, O15, and O109 serotypes have been detected in food samples, highlighting the possible transmission of STEC via the food chain [41].
The genetic variation of Shiga toxin causes changes in its amino acid composition, which may directly influence the virulence of STEC, resulting in a change in the toxin receptor tropism or toxicity of Shiga toxin [11]. In this study, high prevalence of stx1c, stx2a, and stx2c was detected. The stx1c variants are associated with ovine-originated STEC strains [42–44], but the high prevalence of stx1c in buffaloes, cattle, and goats was reported to account for 80% of the stx1 variants, indicating a wide distribution of stx1c variants in STEC of bovine origin [45]. stx1c variants have been found as stx1c only or in combination with stx1, stx2, or stx2d. However, in this study, combinations involving stx1, stx1c, stx2, stx2a, and stx2c (16 isolates); stx1, stx1c, stx2, and stx2a (six isolates); and stx1 and stx1c (13 isolates) were newly found. In addition, stx1c-producing STEC is considered a subset of eae-negative STEC, and is responsible for asymptomatic or mild disease [42, 44, 46]. However, in this study, 29 stx1c-producing STEC harbored eae. The stx1c variants in eae-positive STEC strains might be resulted from the dynamics of virulence genes. Of the stx2 variants, stx2a, stx2c, and stx2d variants have been implicated in high STEC virulence [21, 47]. While stx2d was not detected in the current study, the high prevalence of stx2a, and stx2c suggested the wide distribution of potentially pathogenic STEC strains in cattle farms. The stx2g variant was detected from five STEC non-O157 (three O15 and two O109 STEC). Previously, the stx2g variant has been identified from various sources, including cattle, beef or beef-containing products, and humans, suggesting a possible route of exposure of these STEC types via the food chain [41, 48].
To evaluate the virulence potentials of STEC strains isolated from cattle farms, the phenotypic and genotypic antimicrobial resistance features and the prevalence of virulence genes were investigated. In this study, all STEC isolates were susceptible to all tested antimicrobials, except for AMP, TE, and CTX. Resistance to AMP and TE in diverse sources, including cattle or beef products, have commonly been reported in previous studies [28, 29, 49], but resistance to CTX is uncommon, with only one isolate (of 722) from a bovine source being reported to date [50]. CTX, a third generation cephalosporin, is used as an indicator to identify ESBL production. Although ESBL production was not identified in this study, the presence of CTX-resistant STEC indicates the need for implementing antimicrobial resistance control strategies to prevent the generation and spread of ESBL-producing STEC.
In addition, all the STEC strains that exhibited resistance to AMP, TE, and CTX were STEC non-O157 strains. Genotypic antimicrobial features also varied by its serotype. While antimicrobial resistance genes of tetB, tetC, and bla TEM were only observed in STEC non-O157 strains, tetE was detected only in STEC O157 (34/35; 97.1%). These results suggest that antimicrobial resistance is higher in STEC non-O157 than in STEC O157, consistent with previous studies [49, 50]. While the antimicrobial resistance gene ampC was amplified from all tested STEC isolates, only four STEC non-O157 strains exhibited phenotypic resistance. Since many genes and mechanisms, including efflux pumps or intrinsic resistance, are involved in the development of resistance features, genetic determinants may not represent the phenotypic resistance features [51, 52].
The prevalence of virulence genes in each serotype was either 0 or 100%, except for tir, espP, and iha, indicating the sero-specific feature of virulence genes. To estimate the virulence potentials of STEC strains that might cause a risk to public health, clustering analysis was performed based on the virulence gene profiles. Six clusters were generated, and sero-specific features were observed in each cluster. Cluster 1 was composed of O119 STEC, which has 100% prevalence of the well-known virulence factors eae and ehxA. The association between intimin (encoded by eae) and STEC virulence has been reported previously, and serogroup O119 has been detected in human infections [4, 7]. This indicates that the STEC isolates in Cluster 1 might have the potential to cause human illness. Most of the other STEC strains were grouped in Cluster 4 (47/63, 74.6%), and these strains harbored most of the virulence genes at a high frequency, except for subA and saa. The katP and stcE gene products are believed to promote STEC virulence by assisting STEC colonization in the intestines and degrading the protective layers in the intestines, respectively [27, 53]. A high prevalence of these two genes was reported for sero-pathogroups A and B, which are responsible for severe STEC illness [54]. In this study, all of the O157 and O111 serotypes, which belonged to sero-pathotypes A and B, also belonged to Cluster 4, indicating the high virulence potential of the STEC in Cluster 4. Cluster 5 was characterized by the presence of subA and saa, and consisted of O8, O169, and NT STEC. subA is purported to increase STEC virulence. Saa also increases STEC virulence by assisting in adherence to host cells in eae-negative STEC [55, 56]. On the other hand, the STEC in Clusters 2, 3, and 6 appeared to be less pathogenic to humans. High prevalence of espP, iha, and ehxA was reported regardless of sero-pathotype, suggesting the absence of a strong association between these genes and STEC virulence [54].
Pulsed-field gel electrophoresis analysis was performed to understand the clonal relatedness of STEC strains isolated from cattle farms located in different regions of the Gyeonggi province in Korea during 2012–2015. For the STEC O157 strains, those isolated from the same farm during the same sampling period had indistinguishable PFGE profiles except for a few isolates from farms 4, 7, 11, and 15, which showed one to three different bands. Considering that a single nucleotide mutation at a restriction enzyme site causes three fragment differences [57], a minor genetic variation may have occurred within the farm. In addition, STEC O157 from farm 4 showed high similarity between the 2012 and 2013 isolates. Phylogenetic analysis combined most isolates into group 3, which consisted of isolates from five farms in three different geographical locations. These results indicated the possible presence of a prototype of STEC O157 in the Gyeonggi province with a minor genetic variation, which led to within- and between-farm transmission during 2012–2013. However, STEC O157 isolates from farm 15 showed a higher degree of polymorphism; these isolates clustered in groups 1 and 2 (STEC strains isolated in 2014) and group 3 (STEC strains isolated in 2012). These results indicated that the prototype of STEC O157 in farm 15 might have changed in 2014. Because all the farms were located in the Gyeonggi province and the longest distance between farms was approximately 60 km, temporal effects may have been less important. While a high degree of genetic diversity was observed in STEC non-O157, they were grouped together for strains with the same serotype. STEC O8, O15, O84, and O111 were isolated multiple times and shared genotypic similarity over the 3-year period within the serogroup, implying that these STEC strains have endured and continue to survive, causing within-farm transmission.
Conclusions
Shiga-toxin-producing Escherichia coli prevalence differed greatly between farms, and temperature and rainfall affected the farm prevalence. A considerable number of STEC non-O157 stains were isolated, and different virulence and antimicrobial resistance features were observed between STEC O157 and non-O157 strains. While a high prevalence of virulence genes was observed in STEC O157 strains, the antimicrobial resistance rate was higher in STEC non-O157 strains. In addition, the stx1c variant was detected in eae-positive STEC, suggesting genetic dynamics among virulence genes in STEC isolates. Finally, PFGE analysis revealed the presence of a prototype STEC, which continues to evolve by genetic mutation and causes within- and between-farm transmission within the Gyeonggi province. Our results suggested that STEC from cattle have a high virulence potential and represent a threat to public health. Therefore, continual surveillance of both STEC O157 and non-O157 would be beneficial for controlling and preventing STEC illness.
Abbreviations
- AM:
-
ampicillin
- AMC:
-
amoxicillin–clavulanic acid
- AN:
-
amikacin
- ATM:
-
aztreonam
- C:
-
chloramphenicol
- CAZ:
-
ceftazidime
- CPD:
-
cefpodoxime
- CRO:
-
ceftriaxone
- CTX:
-
cefotaxime
- ESBL:
-
extended-spectrum beta-lactamase
- GM:
-
gentamicin
- IMP:
-
imipenem
- NA:
-
nalidixic acid
- PCR:
-
polymerase chain reaction
- PFGE:
-
pulsed-field gel electrophoresis
- STEC:
-
Shiga toxin-producing Escherichia coli
- STX:
-
trimethoprim–sulfamethoxazole
- TE:
-
tetracycline
- UPGMA:
-
unweighted pair group method with arithmetic mean analysis
References
Riley LW, Remis RS, Helgerson SD, McGee HB, Wells JG, Davis BR, et al. Hemorrhagic colitis associated with a rare Escherichia coli serotype. N Engl J Med. 1983;308(12):681–5.
Scallan E, Hoekstra RM, Angulo FJ, Tauxe RV, Widdowson M-A, Roy SL, et al. Foodborne illness acquired in the united states—major pathogens. Emerg Infect Dis. 2011;17(1):7–15.
Luna-Gierke R, Griffin P, Gould L, Herman K, Bopp C, Strockbine N, et al. Outbreaks of non-O157 Shiga toxin-producing Escherichia coli infection: USA. Epidemiol Infect. 2014;142(11):2270–80.
Blanco M, Blanco J, Mora A, Dahbi G, Alonso M, González E, et al. Serotypes, virulence genes, and intimin types of Shiga toxin (verotoxin)-producing Escherichia coli isolates from cattle in Spain and identification of a new intimin variant gene (eae-ξ). J Clin Microbiol. 2004;42(2):645–51.
Sep A. Ongoing multistate outbreak of Escherichia coli serotype O157: H7 infections associated with consumption of fresh spinach—United States, September 2006. Morbidity and Mortality Weekly Report 2006; 55.
Schimmer B, Nygard K, Eriksen H-M, Lassen J, Lindstedt B-A, Brandal LT, et al. Outbreak of haemolytic uraemic syndrome in Norway caused by stx 2-positive Escherichia coli O103: H25 traced to cured mutton sausages. BMC Infect Dis. 2008;8(1):1.
Brooks JT, Sowers EG, Wells JG, Greene KD, Griffin PM, Hoekstra RM, et al. Non-O157 Shiga toxin–producing Escherichia coli infections in the United States, 1983–2002. J Infect Dis. 2005;192(8):1422–9.
Pihkala N, Bauer N, Eblen D, Evans P, Johnson R, Webb J, et al. Risk profile for pathogenic non-O157 Shiga toxin-producing Escherichia coli (non-O157 STEC). US Department of Agriculture, Food Safety and Inspection Service, Office of Public Health Science, Office of Policy and Program Development 2012:1-64.
Thorpe CM. Shiga toxin-producing Escherichia coli infection. Clin Infect Dis. 2004;38(9):1298–303.
Law D. Virulence factors of Escherichia coli O157 and other Shiga toxin-producing E. coli. J Appl Microbiol. 2000;88(5):729–45.
Paton JC, Paton AW. Pathogenesis and diagnosis of Shiga toxin-producing Escherichia coli infections. Clin Microbiol Rev. 1998;11(3):450–79.
Croxen MA, Law RJ, Scholz R, Keeney KM, Wlodarska M, Finlay BB. Recent advances in understanding enteric pathogenic Escherichia coli. Clin Microbiol Rev. 2013;26(4):822–80.
Brzuszkiewicz E, Thürmer A, Schuldes J, Leimbach A, Liesegang H, Meyer F-D, et al. Genome sequence analyses of two isolates from the recent Escherichia coli outbreak in Germany reveal the emergence of a new pathotype: entero-aggregative-haemorrhagic Escherichia coli (EAHEC). Arch Microbiol. 2011;193(12):883–91.
Gyles CL. Shiga toxin-producing Escherichia coli: an overview. J Anim Sci. 2007;85(13 Suppl):E45–62.
Schmitt CK, McKEE ML, O’Brien AD. Two copies of Shiga-like toxin II-related genes common in enterohemorrhagic Escherichia coli strains are responsible for the antigenic heterogeneity of the O157: H-strain E32511. Infect Immun. 1991;59(3):1065–73.
Kumar A, Taneja N, Kumar Y, Sharma M. Detection of Shiga toxin variants among Shiga toxin–forming Escherichia coli isolates from animal stool, meat and human stool samples in India. J Appl Microbiol. 2012;113(5):1208–16.
Boerlin P, Chen S, Colbourne JK, Johnson R, De Grandis S, Gyles C. Evolution of enterohemorrhagic Escherichia coli hemolysin plasmids and the locus for enterocyte effacement in Shiga toxin-producing E. coli. Infect Immun. 1998;66(6):2553–61.
Kleanthous H, Smith H, Scotland S, Gross R, Rowe B, Taylor C, et al. Haemolytic uraemic syndromes in the British Isles, 1985-8: association with verocytotoxin producing Escherichia coli. Part 2: microbiological aspects. Arch Dis Child. 1990;65(7):722–7.
Jacewicz MS, Acheson DW, Binion DG, West GA, Lincicome LL, Fiocchi C, et al. Responses of human intestinal microvascular endothelial cells to Shiga toxins 1 and 2 and pathogenesis of hemorrhagic colitis. Infect Immun. 1999;67(3):1439–44.
Louise CB, Obrig TG. Specific interaction of Escherichia coli 0157: H7-derived Shiga-like toxin II with human renal endothelial cells. J Infect Dis. 1995;172(5):1397–401.
Eklund M, Leino K, Siitonen A. Clinical Escherichia coli strains carrying stx genes: stx variants and stx-positive virulence profiles. J Clin Microbiol. 2002;40(12):4585–93.
Kaper J, Elliott S, Sperandio V, Perna N, Mayhew G, Blattner F. Attaching and effacing intestinal histopathology and the locus of enterocyte effacement. Escherichia coli O. 1998;157:163–82.
Hicks S, Frankel G, Kaper JB, Dougan G, Phillips AD. Role of intimin and bundle-forming pili in enteropathogenic Escherichia coli adhesion to pediatric intestinal tissue in vitro. Infect Immun. 1998;66(4):1570–8.
Schmidt H, Beutin L, Karch H. Molecular analysis of the plasmid-encoded hemolysin of Escherichia coli O157: H7 strain EDL 933. Infect Immun. 1995;63(3):1055–61.
Brunder W, Schmidt H, Karch H. EspP, a novel extracellular serine protease of enterohaemorrhagic Escherichia coli O157: H7 cleaves human coagulation factor V. Mol Microbiol. 1997;24(4):767–78.
Schmidt H, Geitz C, Tarr PI, Frosch M, Karch H. Non-O157: H7 pathogenic Shiga toxin-producing Escherichia coli: phenotypic and genetic profiling of virulence traits and evidence for clonality. J Infect Dis. 1999;179(1):115–23.
Brunder W, Schmidt H, Karch H. KatP, a novel catalase-peroxidase encoded by the large plasmid of enterohaemorrhagic Escherichia coli O157: H7. Microbiology. 1996;142(11):3305–15.
Khan A, Das S, Ramamurthy T, Sikdar A, Khanam J, Yamasaki S, et al. Antibiotic resistance, virulence gene, and molecular profiles of Shiga toxin-producing Escherichia coli isolates from diverse sources in Calcutta, India. J Clin Microbiol. 2002;40(6):2009–15.
Park HJ, Yoon JW, Heo E-J, Ko E-K, Kim K-Y, Kim Y-J, et al. Antibiotic resistance and virulence potentials of shiga toxin-producing Escherichia coli isolates from raw meats of slaughterhouses and retail markets in Korea. J Microbiol Biotechnol. 2015;25(9):1460–6.
Dong HJ, Cho AR, Hahn TW, Cho S. Development of a multiplex loop-mediated isothermal amplification assay to detect shiga toxin-producing Escherichia coli in cattle. J Vet Sci. 2014;15(2):317–25.
Ekong PS, Sanderson MW, Cernicchiaro N. Prevalence and concentration of Escherichia coli O157 in different seasons and cattle types processed in North America: a systematic review and meta-analysis of published research. Prev Vet Med. 2015;121(1):74–85.
Pearce M, Evans J, McKendrick I, Smith A, Knight H, Mellor D, et al. Prevalence and virulence factors of Escherichia coli serogroups O26, O103, O111, and O145 shed by cattle in Scotland. Appl Environ Microbiol. 2006;72(1):653–9.
Venegas-Vargas C, Henderson S, Khare A, Mosci RE, Lehnert JD, Singh P, et al. Factors Associated with shiga toxin-producing Escherichia coli shedding by dairy and beef cattle. Appl Environ Microbiol. 2016;82(16):5049–56.
Cooley M, Carychao D, Crawford-Miksza L, Jay MT, Myers C, Rose C, et al. Incidence and tracking of Escherichia coli O157: H7 in a major produce production region in California. PLoS ONE. 2007;2(11):e1159.
Fremaux B, Prigent-Combaret C, Vernozy-Rozand C. Long-term survival of Shiga toxin-producing Escherichia coli in cattle effluents and environment: an updated review. Vet Microbiol. 2008;132(1):1–18.
Garber L, Wells S, Hancock D, Doyle M, Tuttle J, Shere J, et al. Risk factors for fecal shedding of Escherichia coli O157: H7 in dairy calves. J Am Vet Med Assoc. 1995;207(1):46–9.
Rugbjerg H, Nielsen EM, Andersen JS. Risk factors associated with faecal shedding of verocytotoxin-producing Escherichia coli O157 in eight known-infected Danish dairy herds. Prev Vet Med. 2003;58(3):101–13.
Cho S, Fossler CP, Diez-Gonzalez F, Wells SJ, Hedberg CW, Kaneene JB, et al. Cattle-level risk factors associated with fecal shedding of Shiga toxin-encoding bacteria on dairy farms, Minnesota, USA. Can J Vet Res. 2009;73(2):151–6.
Hussein HS, Bollinger LM. Prevalence of Shiga toxin–producing Escherichia coli in beef cattle. J Food Protect. 2005;68(10):2224–41.
Nielsen EM, Tegtmeier C, Andersen HJ, Grønbæk C, Andersen JS. Influence of age, sex and herd characteristics on the occurrence of verocytotoxin-producing Escherichia coli O157 in Danish dairy farms. Vet Microbiol. 2002;88(3):245–57.
Beutin L, Miko A, Krause G, Pries K, Haby S, Steege K, et al. Identification of human-pathogenic strains of Shiga toxin-producing Escherichia coli from food by a combination of serotyping and molecular typing of Shiga toxin genes. Appl Environ Microbiol. 2007;73(15):4769–75.
Zhang W, Bielaszewska M, Kuczius T, Karch H. Identification, characterization, and distribution of a Shiga toxin 1 gene variant (stx1c) in Escherichia coli strains isolated from humans. J Clin Microbiol. 2002;40(4):1441–6.
Friedrich AW, Borell J, Bielaszewska M, Fruth A, Tschäpe H, Karch H. Shiga toxin 1c-producing Escherichia coli strains: phenotypic and genetic characterization and association with human disease. J Clin Microbiol. 2003;41(6):2448–53.
Brett KN, Ramachandran V, Hornitzky MA, Bettelheim KA, Walker MJ, Djordjevic SP. stx1c is the most common Shiga toxin 1 subtype among Shiga toxin-producing Escherichia coli isolates from sheep but not among isolates from cattle. J Clin Microbiol. 2003;41(3):926–36.
Vu-Khac H, Cornick NA. Prevalence and genetic profiles of Shiga toxin-producing Escherichia coli strains isolated from buffaloes, cattle, and goats in central Vietnam. Vet Microbiol. 2008;126(4):356–63.
Fitzgerald A, Edrington T, Looper M, Callaway T, Genovese K, Bischoff K, et al. Antimicrobial susceptibility and factors affecting the shedding of E. coli O157: H7 and Salmonella in dairy cattle. Lett Appl Microbiol. 2003;37(5):392–8.
Ethelberg S, Olsen K, Scheutz F, Jensen C, Schiellerup P, Enberg J, et al. Virulence factors for hemolytic uremic syndrome, Denmark. Emerg Infect Dis. 2004;10(5):842–7.
Prager R, Fruth A, Busch U, Tietze E. Comparative analysis of virulence genes, genetic diversity, and phylogeny of Shiga toxin 2 g and heat-stable enterotoxin STIa encoding Escherichia coli isolates from humans, animals, and environmental sources. Int J Med Microbiol. 2011;301(3):181–91.
Sasaki Y, Usui M, Murakami M, Haruna M, Kojima A, Asai T, et al. Antimicrobial resistance in Shiga toxin-producing Escherichia coli O 157 and O 26 isolates from beef cattle. Jpn J Infect Dis. 2012;65(2):117–21.
Mora A, Blanco JsE, Blanco M, Alonso MP, Dhabi G, Echeita A. Antimicrobial resistance of Shiga toxin (verotoxin)-producing Escherichia coli O157: H7 and non-O157 strains isolated from humans, cattle, sheep and food in Spain. Res Microbiol. 2005;156(7):793–806.
Momtaz H, Dehkordi FS, Hosseini MJ, Sarshar M, Heidari M. Serogroups, virulence genes and antibiotic resistance in Shiga toxin-producing Escherichia coli isolated from diarrheic and non-diarrheic pediatric patients in Iran. Gut Pathog. 2013;5(1):39.
Iweriebor BC, Iwu CJ, Obi LC, Nwodo UU, Okoh AI. Multiple antibiotic resistances among Shiga toxin producing Escherichia coli O157 in feces of dairy cattle farms in Eastern Cape of South Africa. BMC Microbiol. 2015;15(1):213.
Bustamante A, Sanso A, Lucchesi P, Parma A. Multiplex PCR assay for the detection of five putative virulence genes encoded in verotoxigenic Escherichia coli plasmids. Curr Microbiol. 2011;62(5):1411–5.
Kobayashi N, Lee KI, Yamazaki A, Saito S, Furukawa I, Kono T. Virulence gene profiles and population genetic analysis for exploration of pathogenic serogroups of Shiga toxin-producing Escherichia coli. J Clin Microbiol. 2013;51(12):4022–8.
Khaitan A, Jandhyala D, Thorpe C, Ritchie J, Paton A. The operon encoding SubAB, a novel cytotoxin, is present in Shiga toxin-producing Escherichia coli isolates from the United States. J Clin Microbiol. 2007;45(4):1374–5.
Paton AW, Srimanote P, Woodrow MC, Paton JC. Characterization of Saa, a novel autoagglutinating adhesin produced by locus of enterocyte effacement-negative Shiga-toxigenic Escherichia coli strains that are virulent for humans. Infect Immun. 2001;69(11):6999–7009.
Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33(9):2233.
FDA. U.S. Food and Drug Administration Bacteriological analytical manual (BAM) http://www.fda.gov/food/foodscienceresearch/laboratorymethods/ucm2006949.htm/ Accessed Jul 2013.
Scheutz F, Teel LD, Beutin L, Piérard D, Buvens G, Karch H, et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J Clin Microbiol. 2012;50(9):2951–63.
Kobayashi H, Shimada J, Nakazawa M, Morozumi T, Pohjanvirta T, Pelkonen S, et al. Prevalence and characteristics of Shiga toxin-producing Escherichia coli from healthy cattle in Japan. Appl Environ Microbiol. 2001;67(1):484–9.
Schmidt H, Henkel B, Karch H. A gene cluster closely related to type II secretion pathway operons of gram-negative bacteria is located on the large plasmid of enterohemorrhagic Escherichia coli O157 strains. FEMS Microbiol Lett. 1997;148(2):265–72.
Bauer RJ, Zhang L, Foxman B, Siitonen A, Jantunen ME, Saxen H, et al. Molecular epidemiology of 3 putative virulence genes for Escherichia coli urinary tract infection–usp, iha, and iroNE. coli. J Infect Dis. 2002;185(10):1521–4.
Quiñones B, Swimley MS, Narm K-E, Patel RN, Cooley MB, Mandrell RE. O-antigen and virulence profiling of Shiga toxin-producing Escherichia coli by a rapid and cost–effective DNA microarray colorimetric method. Shiga toxin-producing Escherichia coli in human, cattle and foods Strategies for detection and control. 2014.
Ng L-K, Martin I, Alfa M, Mulvey M. Multiplex PCR for the detection of tetracycline resistant genes. Mol Cell Probes. 2001;15(4):209–15.
Féria C, Ferreira E, Correia JD, Gonçalves J, Caniça M. Patterns and mechanisms of resistance to β-lactams and β-lactamase inhibitors in uropathogenic Escherichia coli isolated from dogs in Portugal. J Antimicrob Chemother. 2002;49(1):77–85.
Keyes K, Hudson C, Maurer JJ, Thayer S, White DG, Lee MD. Detection of florfenicol resistance genes in Escherichia coli isolated from sick chickens. Antimicrob Agents Chemother. 2000;44(2):421–4.
Van TTH, Chin J, Chapman T, Tran LT, Coloe PJ. Safety of raw meat and shellfish in Vietnam: an analysis of Escherichia coli isolations for antibiotic resistance and virulence genes. Int J Food Microbiol. 2008;124(3):217–23.
Belaaouaj A, Lapoumeroulie C, Caniça MM, Vedel G, Névot P, Krishnamoorthy R, et al. Nucleotide sequences of the genes coding for the TEM-like β-lactamases IRT-1 and IRT-2 (formerly called TRI-1 and TRI-2). FEMS Microbiol Lett. 1994;120(1–2):75–80.
Authors’ contributions
HJD and SC conceived and designed the study. WK, JUA, JK, and HJD collected the samples and epidemiological information, and isolated the STEC strains. HJD and SL carried out the experiments and interpreted the data. HJD was a major contributor in writing the manuscript. All authors read and approved the final manuscript.
Acknowledgements
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Funding
This work was supported by a National Research Foundation of Korea Grant funded by the Korean Government (NRF-2015R1A1A1A05000990). The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Dong, HJ., Lee, S., Kim, W. et al. Prevalence, virulence potential, and pulsed-field gel electrophoresis profiling of Shiga toxin-producing Escherichia coli strains from cattle. Gut Pathog 9, 22 (2017). https://doi.org/10.1186/s13099-017-0169-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13099-017-0169-x