
Abstract
Background
Type 2 diabetes mellitus (T2DM) is a chronic and multifactorial disease with a rapidly rising incidence in Saudi Arabia. Various genes including zinc finger protein 1 (JAZF1) and tetraspanin 8/leucine-rich repeat-containing G protein-coupled receptor (TSPAN8/LGR5) have been previously described to be associated with T2DM. This study investigated the association of JAZF1 (rs864745) and TSPAN8 (rs7961581) with T2DM in a Saudi population.
Methods
Genomic DNA samples from 400 Saudi T2DM patients and 400 healthy controls were genotyped and analyzed using a polymerase chain reaction-restriction fragment length polymorphism method. The difference between the genotype frequencies were carried out with Chi-square test. Odds ratio, 95 % confidence intervals and p values were calculated using multinomial logistic regression. Dominant and recessive models were implemented to show the statistical significances. Analysis of variance was used to compare differences between genotypes for the various parameters.
Results
Distribution frequencies of the AA, AG, and GG genotypes of JAZF1 (rs864745) differed significantly among T2DM patients and healthy controls (p < 0.05). The AG and GG genotypes were independently and significantly associated with a T2DM risk after adjusting for factors such as age, sex, and body mass index [odds ratio (OR) 2.1 (95 % confidence interval (CI) 1.3–3.4); p = 0.002] and [OR 1.9 (95 % CI 1.2–3.1); p = 0.005], respectively. A genotype-based stratification of anthropometric and biochemical data revealed that the AG + GG genotype is associated with waist circumference (p = 0.04) and fasting blood glucose (p = 0.01) and high-density lipoprotein cholesterol levels (p = 0.02). None of the allele or genotype showed the significant association between the T2DM cases and control subjects in rs7961581 polymorphism in TSPAN8/LGR5 gene.
Conclusion
The rs864745 variant in JAZF1 gene may act as genetic risk factors for the development of T2DM in a Saudi population.
Similar content being viewed by others

Background
Type 2 diabetes mellitus (T2DM) is a multifactorial and common chronic metabolic disease that is a major public health concern in many countries including the Arab population [1]. An estimated number of 285 million people are currently diabetic worldwide, and this is expected to increase up to 438 million by the year 2030. [2–4]. Worldwide, Saudi Arabia ranks seventh in T2DM prevalence and the incidence of this disease has doubled over the last two decades in the kingdom [1]. Several risk factors such as obesity, hypertension, undesirable lipid profiles, and increased fasting glucose levels as well as a parental history of diabetes contributes the predisposition for T2DM development [5]. Besides lifestyle factors and other environmental parameters which are partially responsible for the increased T2DM incidence, also genetic risk factors were shown to contribute [6]. The increased incidence of T2DM and the importance of early detection and management have prompted investigations to identify environmental and genetic factors that increase T2DM, risk and the related complications [7].
New insights into the genetic risk for T2DM include variants of the genes juxtaposed with another zinc finger protein 1 (JAZF1) and tetraspanin 8/leucine-rich repeat-containing G protein-coupled receptor (TSPAN8/LGR5) [7–9]. Numerous replication studies have indicated that JAZF1 and TSPAN8/LGR5 affect various aspects of β-cell function [9–13].
The rs864745 variant in JAZF1 and rs7961581 variant in TSPAN8/LGR5 are significantly associated with insulin secretion and glucose-tolerance [14] and T2DM risk [9, 15]. However, there is limited information available on possible association of rs864745 (JAZF1) and rs7961581 (TSPAN8/LGR5) polymorphism with T2DM susceptibility in Saudi population. Therefore, in this case–control study, we tested whether the polymorphisms in JAZF1 (rs864745) and TSPAN8/LGR5 (rs7961581) represents an additional risk factor for T2DM.
Methods
Patient selection
A total of 800 Saudi subjects (400 T2DM patients and 400 healthy controls) belonging to the same demographic area were enrolled in this study as described previously [16]. T2DM diagnosis was based on the criteria of World Health Organization (fasting glucose level ≥126 mg/dL or >7.0 mmol/L). A generalized questionnaire was administered to all consenting subjects and included demographics and past medical history. The subjects with other medical complications such as coronary artery disease, nephropathy, thyroid, and liver or end stage renal diseases were excluded from this study. Healthy volunteers (n = 400) with normal fasting blood glucose levels were selected from the general population based on age and gender. From the total recruited subjects 88.5 % of the T2DM and 52.5 % of the controls had a family history of T2DM. The study was approved by the local ethics committee [1, 5].
Sample collection
Five milliliters (mL) of over-night fasting blood samples were obtained, 2 mL were collected in ethylenediamine tetra acetic acid (EDTA) tubes (used for molecular analysis) and the other 3 mL in serum separator tubes.
Clinical and biochemical estimations
Clinical and anthropometric parameters, including blood pressure (mmHg), weight (kg), height (m), and hip (cm) and waist (cm) circumferences were measured using standard procedures [17]. Body mass index (BMI) was calculated as weight/height2 (kg/m2). Fasting blood glucose (FBG) and lipid profile [triglyceride (TG), total cholesterol (TC), and high-density lipoprotein cholesterol (HDL-C)] were measured using a chemistry auto-analyzer and commercially available kits (Konelab, Finland). Low-density lipoprotein cholesterol (LDL-C) concentrations were calculated using Friedwald’s formula. Insulin was quantified by a solid phase enzyme amplified sensitivity immunoassay (Medgenix INS-ELISA, Biosource, Belgium). Homeostasis model assessment for insulin resistance (HOMA-IR) was calculated as follows = insulin (μU/mL) × glucose (mmol/L)/22.5. Dyslipidemia (low levels of HDL-C) was defined as an HDL-C level of <1.03 mmol/L for men and <1.29 mmol/L for women [5, 16, 17].
DNA extraction
Genomic DNA was isolated from whole blood using a standard DNA extraction kit (Norgen Biotek Corp., Canada). The quality and quantity of DNA were examined using Nanodrop and gel electrophoresis. DNA was stored at −80 °C.
Analysis of polymorphisms
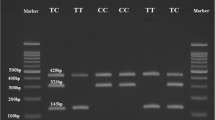
In this study, we have opted 2 genetic variants, such as JAZF1 (rs864745) and TSPAN8/LGR5 (rs7961581) for performing the molecular analysis based on previous literature studies with higher minor allele frequency (MAF) [9, 11–13]. Genotyping was carried out using the polymerase chain reaction-restriction fragment length polymorphism (PCR–RFLP) method. For rs864745 (A > G), primers 5′-GAGCCATATAAGTGATGCTCAAA-3′ (sense) and 5′-GGTTGTCAGGCTTTCCATGT-3′ (antisense) were used to amplify a 378-bp DNA fragment, whereas for rs7961581 (T > C), primers 5′-TGGCATCCAGCTTGTTATTG-3′ (sense) and 5′-TGAGAGCACTGTGTGTGTCACT-3′ (antisense) were used to amplify a 243-bp DNA fragment. PCRs were performed in a thermal cycler with initial denaturation at 95 °C for 5 min, followed by 35 cycles of denaturation at 95 °C for 30 s, annealing 58 °C for 30 s, and extension at 72 °C for 45 s. For rs864745, the PCR product (15 µL) was digested with 2U SSPI restriction endonuclease (New England Bio Labs, UK) for 16 h at 37 °C. The 378-bp (A allele) PCR product was cleaved into 338 and 40-bp fragments in the presence of a G allele (rare), but not in its absence. For rs7961581 polymorphisms, 15 µL of PCR product was digested with 4U Hpy16611 (New England Bio Labs) for 18 h at 37 °C, producing fragments of 210 and 33 bp in the case of a polymorphic variant (allele C) and T allele (243 bp) represents homozygous variant. All digested fragments were directly analyzed by polyacrylamide gel electrophoresis and visualized after silver staining [18].
Statistical analysis
Statistical analyses were performed using SPSS version 19.0 for Windows (SPSS Inc., Chicago, IL, USA). All continuous variables were presented as mean ± standard deviation. An independent sample T test was used to compare the control and T2DM groups. Significant differences were defined as p < 0.05. Genotype frequency differences between patients and controls for each single nucleotide polymorphisms (SNPs) were tested using Chi square test. Odds ratios (ORs) and 95 % confidence intervals (95 % CIs) were calculated using multinomial (multiple) logistic regression with the wild type as the reference. Variables such as age, gender and BMI were included in the multinomial logistic model. Analysis of variance (ANOVA) was used to compare differences between genotypes for the various parameters. Chi-square test was used to identify departures from Hardy–Weinberg equilibrium.
Results
Clinical and anthropometric data of subjects are shown in Table 1. Subjects with T2DM were significantly older and had higher BMI, waist and hip circumferences, systolic and diastolic blood pressure, and FBG, TG, and TC levels and lower HDL-C levels than control subjects.
Genotype distribution
Distribution of genotypes and allele frequencies of the rs864745 (JAZF1) polymorphism in patients and controls were in accordance with the Hardy–Weinberg equilibrium (p > 0.05), but not rs7961581 (TSPAN8/LGR5) (p < 0.01). The estimated powers of the study were 86 and 12.2 % (at desired significance level 0.05) for rs864745 and rs7961581, respectively.
Table 2 displays the genotype distribution for rs864745 and rs7961581 among the T2DM and control subjects. The distributions of the AA, AG, and GG genotypes (p < 0.005) and the AG + GG genotype (p < 0.0001) of rs864745 differed significantly between T2DM and control subjects. Likewise, the frequency AA genotype occurrence (rs864745) was significantly higher in the controls than the T2DM group (47.25 vs. 34.25 %), whereas the occurrence of the AG and GG genotypes was significantly higher in the T2DM group than controls (41.25 vs. 46.5 % for AG; 11.5 vs. 19.25 % for GG). No difference was observed between groups in the genotypic distribution of the rs7961581 variant.
We next determined the association of AG, GG, and AG + GG genotypes of rs864745 with T2DM risk, with AA as the (wild type) reference genotype (Table 2). Occurrence of the AG, GG, and AG + GG genotypes was associated with increased T2DM risk [ORs 1.5 (95 % CI 1.2–2.1, p = 0.004); 2.2 (95 % CI 1.4–3.4, p < 0.0001), and 1.7 (95 % CI 1.2–2.1, p < 0.0001), respectively]. No association was found for the rs7961581 variants (p > 0. 05). We next evaluated the association between JAZF1 or TSPAN8/LGR5 variants and T2DM risk using multiple logistic regression analysis for independence after controlling for other risk factors such as age, gender, and BMI. A significant association was detected for the AG, GG, and AG + GG genotypes of rs864745 (ORs 2.1 (95 % CI 1.3–3.4, p = 0.002); 1.9 (95 % CI 1.2–3.1, p = 0.005), and 2.0 (95 % CI 1.3–3.2), p = 0.001, respectively). Conversely, no effect of rs7961581 on T2DM susceptibility was observed.
Table 3 shows the stratification of the genotype distribution for the rs864745 and rs7961581 variants based on gender. The AG and AG + GG variants of JAZF1 (rs864745) were consistently associated with increased T2DM risk in both female [AG: OR 2.0 (95 % CI 1.2–3.9), p = 0.02); AG + GG: OR 1.9 (95 % CI 1.1–3.6), p = 0.03] and male subjects (AG: OR 1.8 (95 % CI 1.0–3.1), p = 0.04; AG + GG: OR 1.7 (95 % CI 1.0–2.9), p = 0.03). This effect remained significant only in males (p < 0.01) even after adjusting for age and BMI. Finally, the gender-based stratification of the genotype distribution for rs7961581 suggested that the association with T2DM is driven by male gender. The association remained significant even after adjusting for age and BMI (TC: OR 1.8 (95 % CI 1.1–2.9), p = 0.01; TC + CC: OR 1.7 (95 % CI 1.1–2.6), p = 0.01).
Association of JAZF1 and TSPAN8/LGR5 variants with metabolic parameters
To study the effects of JAZF1 and TSPAN8/ LGR5 polymorphisms on the different clinical parameters, we analyzed the distribution of these variables in relation to the various genotypes. Compared with the AA genotype, the AG + GG genotype subjects had significantly higher waist circumference (93.8 ± 20.4 vs. 89.8 ± 22.1 cm, p = 0.04), FBG (9.3 ± 5.1 vs. 8.0 ± 4.7 mmol/L, p = 0.01) and lower HDL-C concentration (0.72 ± 0.31 vs. 0.79 ± 0.34 mmol/L, p = 0.02; Table 4). No such associations were found for the rs7961581 polymorphism, for any of the variables examined Table 5.
Discussion
To the best of our knowledge, the association between the JAZF1 (rs864745) or TSPAN8/LGR5 (rs7961581) polymorphisms and T2DM has not been reported in Arab populations, particularly in Saudi Arabia, despite an increased prevalence and incidence of T2DM. The present study is the first to examine the interaction between silent substitutions with no alteration in amino acids and provides preliminary information on the association between JAZF1 or TSPAN8/LGR5 polymorphisms and T2DM risk. Additionally, the AG + GG genotype of rs864745 was significantly associated with higher waist circumference and FBG as well as lower serum HDL-C concentrations.
The JAZF1 polymorphism (rs864745) is within intron 1 of this gene, which interacts specifically with the ligand-binding domain of TR4 (known to play roles in lipid metabolism and gluconeogenesis) and functions as a TR4-selective cofactor that may be involved in mediating transcriptional repression by TR4 [14, 19, 20]. However, differences in the distribution of risk alleles in various ethnic populations have been reported [9–14]. A number of studies have found an association between certain JAZF1 (rs864745) variants and T2DM, whereas others have found no association [21]. A Framingham Heart Study in whites found that variants of JAZF1 (rs864745) are associated with increased risk for T2DM [22], but a meta-analysis in a Japanese population identified a nominal association between JAZF1 (rs864745) variants and T2DM [23]. Certain variants in the JAZF1 locus are associated abnormal pancreatic β-cell function, which plays a role in the pathogenesis of T2DM and are significantly associated with impaired glucose regulation as well as diabetic nephropathy [14, 19, 20, 24]. Our results are similar to those of the above studies showing that JAZF1 (rs864745) variants are significantly associated with the risk of T2DM [9, 11, 13]. A previous study in the same local Saudi population showed that other JAZF1 variants (rs849134) are associated with increased BMI and T2DM risk [1, 25]. In contrast, a study in a Japanese population reported no significant association between JAZF1 polymorphisms (rs864745) and T2DM [26]. McCarthy et al. [27] have reported that T2DM-associated variants, including those in JAZF1, in healthy populations have a predominant effect on insulin secretion, whereas a study in nondiabetic Finnish men failed to confirm such an association with JAZF1 [28].
The SNP associated with TSPAN8 (rs7961581) have been mapped to a region 110-kb upstream of this gene, which encodes a widely expressed cell surface glycoprotein known to form complexes with integrins. The binding of 6-integrin to laminin negatively affects pancreatic β-cell mass maintenance [20], and SNPs in TSPAN8 may influence pancreatic β-cell function [14, 19, 20]. Using the results from oral glucose tolerance tests, several studies are described an association between certain TSPAN8/LGR5 variants (rs7961581) and insulin release, reflecting altered β-cell function in T2DM [9, 29, 30]. A significant association between TSPAN8/LGR5 rs7961581 and T2DM was found in East Asians and European populations [9, 23]. By contrast, analysis of different TSPAN8 variants (rs7961581) in the present study and rs4760790 from a previous study [31] in the same population failed to show such an association. The discrepancy may be explained by the ethnic and allele frequency differences in various population. The insufficient study power (12.2 %) could also be a probable reason for lacking the association between rs7961581 and T2DM in Saudi population.
We examined various clinical and anthropometric characteristics in the study subjects and correlated them with the various JAZF1 and TSPAN8/LGR5 genotypes (i.e., AG + GG vs. AA and TC + CC vs. TT). Only the AG + GG versus AA genotype comparison revealed an association with waist circumference, FBG, and HDL-C (p < 0.05). T2DM is associated with an increase in total TG and a decrease in HDL-C concentrations. Several population-based studies have shown that visceral fat accumulation, high levels of TGs, and low levels of HDL-C predict the development of T2DM [28, 32]. Indeed, the results from our study suggest that subjects with T2DM have increased waist circumference and TG concentrations and decreased HDL-C levels. To derive further support for the role of JAZF1 (rs864745) and TSPAN8/LGR5 (rs7961581) variants in T2DM development, we analyzed the distribution of confounding factors in relation to possible genotypes. We found the AG + GG genotypes of rs864745 to be significantly associated with increased waist circumference, elevated FBG levels, and reduced levels of serum HDL-C, suggesting the possible participation of rs864745 polymorphism in T2DM etiology. The analysis by Hotta et al. [32] revealed no association of either polymorphism with visceral fat accumulation [19]. There is evidence to indicate an association between JAZF1 variants (rs864745) and IGR risk, 2-h glucose concentrations, or both [33]. Furthermore, An et al. [22] have reported that JAZF1, independently or interactively with other genes, is associated with increased T2DM risk and TG/HDL-C ratio change across age groups. A study in Finnish men showed that JAZF1 variants (rs864745) were nominally associated with an increase in blood lipoprotein levels [28]. Results from another study show that JAZF1 variants are also associated with arteriolosclerosis [34].
The present study has certain limitations that need to be taken into account. A major limitation lies in the unmatched design of controls and T2DM cases. Control subjects were younger and leaner than T2DM patients.
Conclusion
To our knowledge, the current study is the first to determine the distribution and frequency of rs864745 (JAZF1) polymorphisms and their association with T2DM risk, increased waist circumference, and decreased HDL-C concentrations in a Saudi population. This finding, along with those of a previous report [1] emphasize that JAZF1 is a genetic susceptibility locus for the development of T2DM in Saudi populations. Functional analysis are necessary to identify the molecular basis underlying these genetic predispositions and phenotype application may provide useful strategies for predicting T2DM risk in this population.
Abbreviations
- T2DM:
-
type 2 diabetes mellitus
- JAZF1 :
-
juxtaposed with another zinc finger protein 1
- TSPAN8 :
-
tetraspanin 8/leucine-rich repeat-containing G protein-coupled receptor
- EDTA:
-
ethylenediaminetetraaceticacid
- BMI:
-
body mass index
- PCR:
-
polymerase chain reaction
- RFLP:
-
restriction fragment length polymorphism
References
Al-Daghri NM, Alkharfy KM, Alokail MS, Alenad AM, Al-Attas OS, Mohammed AK, Sabico S, Albagha OM. Assessing the contribution of 38 genetic loci to the risk of type 2 diabetes in the Saudi Arabian Population. Clin Endocrinol (Oxf). 2013;80(Suppl 4):532–7.
Tota K, Rayabarapu N, Moosa S, Talla V, Bhyravbhatla B, Rao. InDiaMed: a comprehensive database of Indian medicinal plants for diabetes. Bioinformation. 2013;9(Suppl 7):378–80.
Raza ST, Abbas S, Ahmed F, Fatima J, Zaidi ZH, Mahdi F. Association of MTHFR and PPARγ2 gene polymorphisms in relation to type 2 diabetes mellitus cases among north Indian population. Gene. 2012;511(Suppl 2):375–9.
Al-Daghri NM, Al-Attas OS, Alokail MS, Alkharfy KM, Hussain T, Yakout S, Vinodson B, Sabico S. Adiponectin gene polymorphisms (T45G and G276T), adiponectin levels and risk for metabolic diseases in an Arab population. Gene. 2012;493(Suppl 1):142–7.
Alharbi KK, Khan IA, Syed R. Circulating C5L2 gene polymorphism is associated with type 2 diabetes mellitus in Saudi population. Mol Biol Rep. 2013;40(Suppl 11):6323–7.
Alharbi KK, Hussain T, Alharbi FK, Tabassum SN, Mohammed AA, Gambhir D, Khan IA. Apolipoprotein C3 gene variants and risk of developing type 2 diabetes in Saudi subjects. Metab Syndr Relat Disord. 2015;13(Suppl 7):298–303.
Jeong JH, Yook S, Lee H, Ahn CH, Lee DY, Byun Y. Effects of surface camouflaged islet transplantation on pathophysiological progression in a db/db type 2 diabetic mouse model. Biochem Biophys Res Commun. 2013;433(Suppl 4):513–8.
Goodarzi MO, Guo X, Cui J, Jones MR, Haritunians T, Xiang AH, Chen YD, Taylor KD, Buchanan TA, Hsueh WA, Raffel LJ, Rotter JI. Systematic evaluation of validated type 2 diabetes and glycaemic trait loci for association with insulin clearance. Diabetologia. 2013;56(Suppl 6):1282–90.
Zhou DZ, Liu Y, Zhang D, Liu SM, Yu L, Yang YF, Zhao T, Chen Z, Kan MY, Zhang ZF, Feng GY, Xu H, He L. Variations in/nearby genes coding for JAZF1, TSPAN8/LGR5 and HHEX-IDE and risk of type 2 diabetes in Han Chinese. J Hum Genet. 2010;55(Suppl 12):810–5.
Zeggini E, Scott LJ, Saxena R, et al. Meta-analysis of genome-wide association data and large-scale replication identifies additional susceptibility loci for type 2 diabetes. Nat Genet. 2008;40(Suppl 5):638–45.
Chang YC, Liu PH, Yu YH, Kuo SS, Chang TJ, Jiang YD, Nong JY, Hwang JJ, Chuang LM. Validation of type 2 diabetes risk variants identified by genome-wide association studies in Han Chinese population: a replication study and meta-analysis. PLoS ONE. 2014;9(Suppl4):e95045.
Gamboa-Melendez MA, Huerta-Chagoya A, Moreno-Macias H, Vazquez-Cardenas P, Ordonez-Sanchez ML, Rodriguez-Guillen R, Riba L, Rodriguez-Torres M, Guerra-Garcia MT, Guillen-Pineda LE, Choudhry S, Del Bosque-Plata L, Canizales-Quinteros S, Perez-Ortiz G, Escobedo-Aguirre F, Parra A, Lerman-Garber I, Aguilar-Salinas CA, Tusie-Luna MT. Contribution of common genetic variation to the risk of type 2 diabetes in the Mexican Mestizo population. Diabetes. 2012;61(Suppl 12):3314–21.
Tam CH, Ho JS, Wang Y, Lam VK, Lee HM, Jiang G, Lau ES, Kong AP, Fan X, Woo JL, Tsui SK, Ng MC, So WY, Chan JC, Ma RC. Use of net reclassification improvement (NRI) method confirms the utility of combined genetic risk score to predict type 2 diabetes. PLoS One. 2013;8(Suppl 12):e83093.
Grarup N, Andersen G, Krarup NT, Albrechtsen A, Schmitz O, Jorgensen T, Borch-Johnsen K, Hansen T, Pedersen O. Association testing of novel type 2 diabetes risk alleles in the JAZF1, CDC123/CAMK1D, TSPAN8, THADA, ADAMTS9, and NOTCH2 loci with insulin release, insulin sensitivity, and obesity in a population-based sample of 4516 glucose-tolerant middle-aged Danes. Diabetes. 2008;57(Suppl 9):2534–40.
Cheng I, Caberto CP, Lum-Jones A, Seifried A, Wilkens LR, Schumacher FR, Monroe KR, Lim U, Tiirikainem M, Kolonel LM, Henderson BE, Stram DO, Haiman CA, Le Marchand L. Type 2 diabetes risk variants and colorectal cancer risk: the multiethnic cohort and PAGE studies. Gut. 2011;60(Suppl 12):1703–11.
Alharbi KK, Khan IA, Al-Daghri NM, Munshi A, Sharma V, Mohammed AK, Wani KA, Al-Sheikh YA, Al-Nbaheen MS, Ansari MG, Syed R. ABCA1 C69T gene polymorphism and risk of type 2 diabetes mellitus in a Saudi population. J Biosci. 2013;38(Suppl 5):893–7.
Alharbi KK, Khan IA, Munshi A, Alharbi FK, Al-Sheikh Y, Alnbaheen MS. Association of the genetic variants of insulin receptor substrate 1 (IRS-1) with type 2 diabetes mellitus in a Saudi population. Endocrine. 2014;47(Suppl 2):472–7.
Al-Hakeem MM, Abotalib Z, Alharbi KK, Khan IA. Relationship between the paraoxonase 1 gene glutamine 192 to arginine polymorphism and gestational diabetes mellitus in Saudi women. Clin Biochem. 2014;47(Suppl 15):122–5.
Jang WY, Bae KB, Kim SH, Yu DH, Kim HJ, Ji YR, Park SJ, Park SJ, Kang MC, Jeong JI, Park SJ, Lee SG, Lee I, Kim MO, Yoon D, Ryoo ZY. Overexpression of Jazf1 reduces body weight gain and regulates lipid metabolism in high fat diet. Biochem Biophys Res Commun. 2013;444(Suppl 3):296–301.
Kilkenny DM, Rocheleau JV. Fibroblast growth factor receptor-1 signaling in pancreatic islet beta-cells is modulated by the extracellular matrix. Mol Endocrinol. 2008;22(Suppl 1):196–205.
Haiman CA, Fesinmeyer MD, Spencer KL, Buzkova P, Voruganti VS, Wan P, Haessler J, Franceschini N, Monroe KR, Howard BV, Jackson RD, Florez JC, Kolonel LN, Buyske S, Goodloe RJ, Liu S, Manson JE, Meigs JB, Waters K, Mukamal KJ, Pendergrass SA, Shrader P, Wilkens LR, Hindorff LA, Ambite JL, North KE, Peters U, Crawford DC, Le Marchand L, Pankow JS. Consistent directions of effect for established type 2 diabetes risk variants across populations: the population architecture using Genomics and Epidemiology (PAGE) Consortium. Diabetes. 2012;61(Suppl 6):1642–7.
An P, Feitosa M, Ketkar S, Adelman A, Lin S, Borecki I, Province M. Epistatic interactions of CDKN2B-TCF7L2 for risk of type 2 diabetes and of CDKN2B-JAZF1 for triglyceride/high-density lipoprotein ratio longitudinal change: evidence from the Framingham Heart Study. BMC Proc. 2009;3(Suppl 7):S71.
Omori S, Tanaka Y, Horikoshi M, Takahashi A, Hara K, Hirose H, Kashiwagi A, Kaku K, Kawamori R, Kadowaki T, Nakamura Y, Maeda S. Replication study for the association of new meta-analysis derived risk loci with susceptibility to the type 2 diabetes in 6244 Japanese individuals. Diabetologia. 2009;52(Suppl 8):1554–60.
Chen G, Xu Y, Lin Y, Lai X, Yao J, Huang B, Chen Z, Huang H, Fu X, Lin L, Lai S, Wen J. Association study of genetic variants of 17 diabetes-related genes/loci and cardiovascular risk and diabetic nephropathy in the Chinese She population. J Diabetes. 2013;5(Suppl 2):136–45.
Al-Daghri NM, Alkharfy KM, Al-Attas OS, Krishnaswamy S, Mohammed AK, Albagha OM, Alenad AM, Chrousos GP, Alokail MS. Association between type 2 diabetes mellitus-related SNP variants and obesity traits in a Saudi population. Mol Bio Rep. 2014;41(Suppl 3):1731–40.
Takeuchi F, Serizawa M, Yamamoto K, Fujisawa T, Nakashima E, Ohnaka K, Ikegami H, Sugiyama T, Katsuya T, Miyagishi M, Nakashima N, Nawata H, Nakamura J, Kono S, Takayanagi R, Kato N. Confirmation of multiple risk Loci and genetic impacts by a genome-wide association study of type 2 diabetes in the Japanese population. Diabetes. 2012;58(Suppl 7):1690–9.
McCarthy MI, Zeggini E. Genome-wide association studies in type 2 diabetes. Curr Diab rep. 2009;9(Suppl 2):164–71.
Stancakova A, Kuulasmaa T, Paananen J, Jackson AU, Bonnycastle LL, Collins FS, Boehnke M, Kuusisto J, Laakso M. Association of 18 confirmed susceptibility loci for type 2 diabetes with indices of insulin release, proinsulin conversion, and insulin sensitivity in 5327 nondiabetic Finnish men. Diabetes. 2009;58(Suppl 9):2129–36.
Boesgaard TW, Gjesing AP, Grarup N, Rutanen J, Jansson PA, Hribal ML, Sesti G, Fritsche A, Stefan N, Staiger H, Haring H, Smith U, Laakso M, Pedersen O, Hansen T, EUGENE2 Consortium. Variant near ADAMTS9 known to be associate with type 2 diabetes is related to insulin resistance in offspring of type 2 diabetes patients—EUGENE2 study. PLoS One. 2009;4(Suppl 9):e7236.
Villegas R, Goodloe RJ, McClellan BE Jr, Boston J, Crawford DC. Gene-carbohydrate and gene-fiber interactions and type 2 diabetes in diverse populations from the National Health and Nutrition Examination Surveys (NHANES) as part of the Epidemiologic Architecture for Genes Linked to Environment (EAGLE) study. BMC Genet. 2014;15(Suppl 69):1–8.
Al-Daghri NM, Clerici M, Al-Attas O, Forni D, Alokail MS, Alkharfy KM, Sabico S, Mohammed AK, Cagliani R, Sironi M. A nonsense polymorphism (R392X) in TLR5 protects from obesity but predisposes to diabetes. J Immunol. 2013;190(Suppl 7):3716–20.
Hotta K, Kitamoto A, Kitamoto T, Mizusawa S, Teranishi H, So R, Matsuo T, Nakata Y, Hyogo H, Ochi H, Nakamura T, Kamohara S, Miyatake N, Kotani K, Komatsu R, Itoh N, Mineo I, Wada J, Yoneda M, Nakajima A, Funahashi T, Miyazaki S, Tokunaga K, Masuzaki H, Ueno T, Chayama K, Hamaguchi K, Yamada K, Hanafusa T, Oikawa S, Yoshimatsu H, Sakata T, Tanaka K, Matsuzawa Y, Nakao K, Sekine A. Association between type 2 diabetes genetic susceptibility loci and visceral and subcutaneous fat area as determined by computed tomography. J Hum Genet. 2012;57(Suppl 5):305–10.
Brito EC, Lyssenko V, Renstrom F, Berglund G, Nilsson PM, Groop L, Franks PW. Previously associated type 2 diabetes variants may interact with physical activity to modify the risk of impaired glucose regulation and type 2 diabetes: a study of 16,003 Swedish adults. Diabetes. 2009;58(Suppl 6):1411–8.
Chou SH, Shulman JM, Keenan BT, Secor EA, Buchman AS, Schneider J, Bennett DA, De Jager PL. Genetic susceptibility for ischemic infarction and atherosclerosis based on neuropathologic evaluations. Cerebrovasc Dis. 2013;36(Suppl 3):181–8.
Sanghera DK, Been L, Ortega L, Wander GS, Mehra NK, Aston CE, Mulvihill JJ, Ralhan S. Testing the association of novel meta-analysis-derived diabetes risk genes with type II diabetes and related metabolic traits in Asian Indian Sikhs. J Hum Genet. 2009;54(Suppl 3):162–8.
Authors’ contributions
AKK: acquisition, analysis and interpretation data. KIA: corresponding author, planned the experiment, and helped in drafting the manuscript. MAK: drafted the manuscript. VB: performed the statistical analysis. AFK: helped VB for preparing the excels in the statistical analysis and interpretation data. ANM: selection of cases and controls, analysis, interpretation data and finalized the draft. All authors read and approved the final manuscript.
Acknowledgements
The authors would like to extend their sincere appreciation to the Deanship of Scientific Research at King Saud University for its funding of this research through the Research Group Project no RGP-VPP-244.
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Alharbi, K.K., Ali Khan, I., Syed, R. et al. Association of JAZF1 and TSPAN8/LGR5 variants in relation to type 2 diabetes mellitus in a Saudi population. Diabetol Metab Syndr 7, 92 (2015). https://doi.org/10.1186/s13098-015-0091-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13098-015-0091-7