
Abstract
Background
Glucoamylase is an important industrial enzyme in the saccharification of starch into glucose. However, its poor thermostability and low catalytic efficiency limit its industrial saccharification applications. Therefore, improving these properties of glucoamylase is of great significance for saccharification in the starch industry.
Results
In this study, a novel glucoamylase-encoding gene TlGa15B from the thermophilic fungus Talaromyces leycettanus JCM12802 was cloned and expressed in Pichia pastoris. The optimal temperature and pH of recombinant TlGa15B were 65 ℃ and 4.5, respectively. TlGa15B exhibited excellent thermostability at 60 ℃. To further improve thermostability without losing catalytic efficiency, TlGa15B-GA1 and TlGa15B-GA2 were designed by introducing disulfide bonds and optimizing residual charge–charge interactions in a region distant from the catalytic center. Compared with TlGa15B, mutants showed improved optimal temperature, melting temperature, specific activity, and catalytic efficiency. The mechanism underlying these improvements was elucidated through molecular dynamics simulation and dynamics cross-correlation matrices analysis. Besides, the performance of TlGa15B-GA2 was the same as that of the commercial glucoamylase during saccharification.
Conclusions
We provide an effective strategy to simultaneously improve both thermostability and catalytic efficiency of glucoamylase. The excellent thermostability and high catalytic efficiency of TlGa15B-GA2 make it a good candidate for industrial saccharification applications.
Similar content being viewed by others


Background
Glucoamylase (1,4-α-d-glucan glucohydrolase, EC 3.2.1.3) is one of the most important and widely used industrial enzymes, exhibiting great application potential in sugar, ethanol, bread, beer, textile, and pharmaceutical industries [1]. Glucoamylase could catalyze the hydrolysis of α-1,4 and α-1,6 glycosidic linkages in starch and related oligosaccharides from non-reducing ends to release β-d-glucose [2]. Currently, commercial glucoamylases are mainly obtained from filamentous fungi, such as Aspergillus niger, Rhizopus niveus, and R. delemar, displaying moderate thermostability and slow catalytic activities [3, 4]. However, the industrial conversion of starch to glucose consists of a liquefaction process at 95–105 ℃ for 2 h and a saccharification process at 60 ℃ for 48–72 h. Switching from the liquefaction to the saccharification process requires additional specialized equipment for cooling to ensure the catalytic integrity of glucoamylase [5]. The poor thermostability of current glucoamylases results in great energy consumption and process inconvenience to undergo the saccharification process in starch industries.
Generally speaking, enzyme discovery and engineering are the major approaches for acquiring enzymes with higher thermostability and catalytic efficiency. Moreover, the search for thermostable glucoamylases derived from thermophilic microorganisms has attracted immense attention over recent years. So far, several thermostable glucoamylases have been identified from A. wentii, A. oryzae, A. flavus, Fomitopsis palustris, Chaetomium thermophilum, Rhizomucor pusillus, Thermoanaerobacter tengcongensis, and Sulfolobus solfataricus, all of which exhibit optimal temperature or thermostability above 60 ℃ [3, 6,7,8,9,10,11,12]. Nevertheless, further research is needed pertaining to the industrial aspect of the saccharification process. Reconstructing enzymes by protein engineering is another efficient way to obtain glucoamylases with high thermostability. Based on previous research, directed evolution and rational design, such as in the construction of disulfide bonds, result in significant thermostability improvement of glucoamylases. [13,14,15,16]. However, due to the activity-stability trade-off, an increase in enzyme thermostability is accompanied by a decrease in catalytic efficiency [17, 18].
Recently, it has been reported that increasing the overall rigidity of protein while minimizing influence on the active site could render improved thermostability either without affecting catalytic activity or ideally with positively promoting catalytic activity [19]. Moreover, many previous studies on phytase, β-glucanase, and xylanase thermostability engineering support this conclusion [20,21,22]. Such studies indicate that selecting mutant sites in a region distant from the catalytic center may be feasible to design enzymes with increased thermostability and catalytic efficiency.
Talaromyces leycettanus JCM12802 is a typical thermophilic fungus with an optimum temperature for growth at 40 ℃, producing a wide range of thermostable glycoside hydrolases, such as cellulase, β-glucosidase, and mannanase [23,24,25]. In this study, a novel thermostable glucoamylase TlGa15B from T. leycettanus JCM12802 was expressed and characterized. In order to further improve thermostability, TlGa15B mutants were rationally designed by introducing disulfide bonds and optimizing residual charge–charge interactions in a region distant from the catalytic center. Moreover, the application potential of the best mutant was evaluated by comparison with a commercial glucoamylase commonly used in the starch industry for saccharification.
Results and discussion
Cloning and sequence analysis of TlGa15B
In the present study, a novel glucoamylase-encoding gene TlGa15B was identified in the genome of thermophilic fungus T. leycettanus JCM12802, and the corresponding cDNA sequences were successfully obtained from the fungus grown in PDB medium. The TlGa15B was interrupted by 4 introns, resulting in an open reading frame of 1842 bp. Deduced TlGa15B contained 614 amino acid residues and harbored a signal peptide of 20 residues based on predictions established by the SignalP 5.0 program [26]. The isoelectric point and calculated molecular weight of glucoamylase TlGa15B were 4.76 and 63.3 kDa, respectively. TlGa15B shared the highest identity (69.85%) with a previously reported glucoamylase derived from Rasamsonia emersonii [27]. The sequence and structure analysis showed that TlGa15B was a typical glucoamylase, containing a catalytic domain of GH15 and a carbohydrate-binding domain of CBM20. Glu211, and Glu433 was predicted to be the catalytic residues that served as the proton donor and acceptor, respectively. Moreover, five highly conserved residues associated with substrate binding in the GH15 family were identified in the catalytic domain of TlGa15B, including Arg86, Asp87, Leu209, Trp210, Glu212, and Arg338 [28]. In addition, the modeled structure of TlGa15B was constructed using the glucoamylase from Penicillium oxalicum (PDB: 6FHV, 58% sequence identity and 94% coverage) as template with the help of SWISS-MODEL. Meanwhile, the Ramachandran plot was used to evaluate the quality of the structural model. There were only two amino acid residues observed in disallowed region (Additional file 1), indicating that the modeled structure of TlGa15B was credible.
Expression and purification of recombinant TlGa15B
Pichia pastoris, the microbial expression system most widely used for the large-scale production of commercial enzymes, was used as the recombinant expression host to express glucoamylase TlGa15B [29]. The cDNA fragment coding for the mature TlGa15B was inserted into PIC9 and successfully expressed in Pichia pastoris GS115. After methanol induction, the glucoamylase activity of the culture supernatant reached 5000 U/L at 48 h. After purification by anion exchange, the recombinant TlGa15B had an apparent molecular weight of ~65 kDa on the SDS-PAGE (Additional file 2), which was close to the theoretical molecular weight.
Biochemical characterization of purified recombinant TlGa15B
The optimal pH of purified recombinant TlGa15B was pH 4.5 (Fig. 1a). Similar pH optima for glucoamylase activities had been reported from C. thermophilum pH 4.5–5.0 [11], A. tritici pH 4.0–5.0 [30], and Bispora sp. 4.0 [31]. Meanwhile, TlGa15B exhibited high stability in pH values ranging from 2.0 to 12.0 (Fig. 1b). The residual hydrolytic activity of TlGa15B after 1 h of incubation was over 75% of the original activity prior to incubation. These results suggested that TlGa15B was an acidic glucoamylase with broad-range pH stability.

Effect of pH and temperature on the activity and stability of purified recombinant TlGa15B and mutants TlGa15B-GA1 and TlGa15B-GA2. a Effect of pH on the activity at 37 ℃. b Effect of pH on the stability at 37 ℃. c Effect of temperature (20–90 ℃) on the activity. d Effect of temperature (75 ℃) on the stability
As shown in Fig. 1c, the hydrolytic activity of purified recombinant TlGa15B increased with increasing temperature, reaching a maximum at 65 ℃. The optimal temperature was the same as the most widely used glucoamylases from A. niger [1, 32]. In terms of thermostability, TlGa15B from T. leycettanus JCM12802 exhibited excellent thermostability. TlGa15B retained 79.5%, 54.8%, and 36.7% of hydrolytic activity after 1 h of incubation at 55 ℃, 60 ℃, and 65 ℃, respectively (Additional file 3). When the temperature increased to above 75 ℃, the thermostability of TlGa15B significantly decreased (Fig. 1d). In contrast, the thermostability of TlGa15B is superior to that of most reported glucoamylase, which are stable at temperatures below 55 ℃ [1].
In addition, the kinetic parameters of purified TlGa15B with respect to soluble starch were determined at 55 ℃. The Km and Vmax values of purified recombinant TlGa15B were 0.77 mg/mL and 719.2 μmol/min/mg, respectively (Table 1). Meanwhile, the specific enzyme activities of purified recombinant TlGa15B were also evaluated using various substrates, including soluble starch, amylose, glycogen, α-cyclodextrin, β-cyclodextrin, and γ-cyclodextrin. The results revealed that TlGa15B was active toward soluble starch, amylose, and glycogen, but not other substrates, such as α-cyclodextrin, β-cyclodextrin, and γ-cyclodextrin. The specific activity toward glycogen was higher than toward soluble starch and amylose. The specific activities of the purified recombinant TlGa15B to soluble starch, amylose, and glycogen were 496.2, 221.1, and 658.3 U/mg, respectively.
Selection of the mutagenesis sites in TlGa15B
It was reported that the construction of disulfide bonds and the optimization of residual charge–charge interactions were efficient strategies to improve the thermostability of industrial enzymes [33]. In this study, these protein engineering approaches were applied to enhance the thermostability of TlGa15B. Based on multiple sequence alignment and structure analysis, S132, Y492, L548, and A562 were chosen as targets to construct two disulfide bonds. The mutant TlGa15B-GA1 (S132C-Y492C/L548C-A562C) was constructed via PCR-mediated site-directed mutagenesis. As shown in Additional file 4, two disulfide-linked peptides SNPSGGLCT/SASGPCA (132C/492C) and PLWYCIV/SAIPCSA (548C/562C) were observed in the LC–MS/MS analysis, indicating that TlGa15B mutants formed disulfide bonds in the target sites. In addition, the enzyme thermal stability system (ETSS) was used to calculate the total interaction energy (Eij) between charged amino acids of TlGa15B [34]. 6 non-conserved amino acid residues (C94/E98/Q108/D289 /D296/E473) in a region distant from the catalytic center but in the catalytic domain of glucoamylase were selected for further analysis by ETSS (Fig. 2; Additional file 5). On account of the ETSS analysis, Q108 (total Eij = 27.3 kJ/mol) was mutated to a neutral amino acid alanine (total Eij = 2.3 kJ/mol), exhibiting the most significant decline in the Eij value. Subsequently, the mutant TlGa15B-GA2 (S132C-Y492C/L548C-A562C/Q108E) was generated via site-directed mutagenesis using TlGa15B-GA2 as the template.

Total interaction energy of each chargeable residue in TlGa15B by ETSS analysis (a). The modeled structure of TlGa15B (b). Six candidate mutated sites away from the catalytic center were indicated in blue balls. The sites for introduction of disulfide bonds were shown as red sticks
Comparison of the enzymatic properties of TlGa15B and mutants
As shown in Fig. 1a, b, there were no significant differences in pH optima and stability between TlGa15B and its mutants TlGa15B-GA1 and TlGa15B-GA2. By comparison, the optimal temperature and thermostability of mutants both significantly improved. The optimal temperatures of TlGa15B-GA1 and TlGa15B-GA2 were 70 ℃ and 75 ℃, which were 5 ℃ and 10 ℃ higher than that of TlGa15B, respectively (Fig. 1c). The TlGa15B incubated at 75 ℃ for 2 min only had 13.8% residual enzyme activity, while the mutant TlGa15B-GA1 still contained 60.6% and 24.6% of enzyme activity after incubation at 75 ℃ for 10 min and 30 min, respectively. Furthermore, the mutant TlGa15B-GA2 was extremely stable at 75 ℃, and 40% of enzyme activity remained after 1 h of incubation (Fig. 1d). Meanwhile, the DSC results supported the conclusion that both TlGa15B-GA1 and TlGa15B-GA2 exhibited higher thermostability than TlGa15B (Fig. 3). Compared with the Tm value of TlGa15B (72 ℃), the Tm values of TlGa15B-GA1 and TlGa15B-GA2 showed a significant increase of 8 °C (80 ℃) and 9 °C (81 ℃), respectively.

Thermograms of TlGa15B and mutants TlGa15B-GA1 and TlGa15B-GA2 detected using the DSC
Remarkably, TlGa15B-GA1 and TlGa15B-GA2 also exhibited higher catalytic activity and efficiency than TlGa15B (Table 1; Additional file 6). The specific activities of TlGa15B-GA1 and TlGa15B-GA2 were approximately 1.6-fold and 2.2-fold higher than that of TlGa15B, respectively. In contrast to TlGa15B, the catalytic efficiencies of TlGa15B-GA1 and TlGa15B-GA2 increased by 55.7% and 305.4%, respectively. Usually, thermostability improvement of enzymes is accompanied by a decrease in catalytic activity due to the activity-stability trade-off [17, 18]. However, these results indicate that the activity-stability trade-off could be overcome by carefully selecting mutation sites distant from the catalytic center.
MD simulation and DCCM analysis of TlGa15B and mutants
In order to elucidate the mechanism underlying the thermostability of mutants TlGa15B-GA1 and TlGa15B-GA2, MD simulations were carried out at 300 K for 20 ns. The root mean square deviation (RMSD) was an important parameter to reflect the rigidity of the protein structure, which correlated with protein thermostability [35]. The RMSD values of TlGa15B-GA1 and TlGa15B-GA2 significantly decreased compared with that of TlGa15B, indicating that the overall structures of the mutants were more thermostable (Fig. 4a). Moreover, based on the root mean square fluctuation (RMSF) analysis, residues Q108, S132, Y492, L548, and A562 of TlGa15B were more flexible than the corresponding ones of the mutants TlGa15B-GA1 and TlGa15B-GA2 (Fig. 4b). Besides, the flexibilities of amino acid residues in the CBM domain of thermostable mutants were markedly decreased. The lower RMSF values of these amino acid residues rendered the CBM domain more stable, which might result in improved catalytic efficiency of TlGa15B-GA1 and TlGa15B-GA2.

MD simulation analysis of TlGa15B and mutants TlGa15B-GA1 and TlGa15B-GA2. Comparison of RMSD values between WT and mutants calculated from 20-ns MD simulations at 300 K (a). Comparison of normalized RMSF between WT and mutants (b). DCCM analysis of TlGa15B (c) and TlGa15B-GA2 (d) based on the MD simulation trajectories
In addition, to understand the complex interaction between the selected mutated amino acid residues and the CBM domain, the DCCMs of TlGa15B and TlGa15B-GA2 were calculated using the coordinates of the Cα atoms from the trajectories. As shown in Fig. 4c, d, the negative correlation between the amino acid residues (100–120) and the CBM domain (residues 490–613) was weakened in the mutant TlGa15B-GA2, thereby indicating a marked reduction in the movement of the negative correlation between them. These results were consistent with the conclusions of the RMSF analysis.
Starch saccharification using TlGa15B-GA2 in combination with pullulanase
In order to evaluate the performance of industrial saccharification applications, the best mutant TlGa15B-GA2 was chosen for enzymatic saccharification of liquefied starch. As shown in Fig. 5, the maximum glucose productions for TlGa15B-GA2 and the commercial glucoamylase GA-LD were obtained after 30 h of incubation at 60 °C. The DX value (glucose content) for TlGa15B-GA2 and GA-LD reached 96.4% and 96.7%, respectively, indicating no significant difference between TlGa15B-GA2 and the commercial glucoamylase GA-LD. Moreover, the DX value of TlGa15B-GA2 was in accordance with the industrial requirement for the production of glucose with a DX value above 96%. These results suggested that TlGa15B-GA2 had the same saccharification effect as industrial glucoamylases for the enzymatic saccharification process of starch.

Comparison of the starch saccharification effect between TlGa15B-GA2 and the commercial glucoamylase GA-LD in glucose production at 60 °C
Conclusions
TlGa15B, derived from thermophilic fungus T. leycettanus JCM12802, exhibited high optimal temperature and excellent thermostability above 60 ℃. By introducing disulfide bonds and optimizing residual charge–charge interactions in a region distant from the catalytic center, TlGa15B-GA1 and TlGa15B-GA2 showed improvements in optimal temperature, melting temperature, specific activity, and catalytic efficiency. The mechanism underlying the improved thermostability and catalytic efficiency of glucoamylase was elucidated by MD simulation and DCCM analysis. The performance of TlGa15B-GA2 was similar to that of the commercial glucoamylase during the saccharification process of starch. These properties of TlGa15B-GA2 make it a promising candidate for industrial saccharification applications.
Methods
Strain and substrates
The thermophilic fungus T. leycettanus JCM12802 was purchased from the Japan Collection of Microorganisms RIKEN BioResource Center (Tsukuba, Japan) and was maintained at 4 ℃ on a potato dextrose agar plate. Soluble starch, amylose, glycogen, α-cyclodextrin, β-cyclodextrin, and γ-cyclodextrin were purchased from Sigma-Aldrich. All other chemicals were of analytical grade and commercially available.
Cloning of TlGa15B and mutants
T. leycettanus JCM12802 was cultured in the potato dextrose broth (PDB) medium at 40 ℃ for 3 days. Total RNA was extracted using the TRIZOL reagent (Invitrogen, Waltham, MA, USA) according to the operation manual. The first strand cDNA was synthesized from the total RNA using the FastKing RT Kit with gDNase (Tiangen, Beijing). Based on the 5′ and 3′-end sequences of the TlGa15B structural gene, the TlGa15B-encoding gene devoid of the signal peptide sequence was amplified with gene-specific primers (as shown in Additional file 7). The PCR product was cloned into the plasmid pPIC9 to construct a recombinant plasmid pPIC9-TlGa15B. Site-directed mutants pPIC9-TlGa15B-GA1 and pPIC9-TlGa15B-GA2 were constructed using the Fast Mutagenesis System Kit (TransGen, Beijing) with the plasmid pPIC9-TlGa15B as the template.
Expression and purification of TlGa15B and mutants
The recombinant plasmid PIC9-TlGa15B, pPIC9-TlGa15B-GA1, and pPIC9-TlGa15B-GA2 were linearized with BglII and transformed into P. pastoris GS115 competent cells, respectively. The positive transformant with the highest glucoamylase activity was inoculated into YPD medium at 30 °C for 2 days with shaking at 200 rpm and used as the inoculum of 300 mL BMGY medium. The cultures were grown at 30 °C for 2 days, and cells were then harvested and resuspended in 200 mL BMMY medium for 2 days growth at 30 °C. The crude enzyme was collected by centrifugation at 12,000g for 10 min at 4 °C, followed by concentration through a 10 kDa cut-off centrifuge filter. The crude enzyme was dialyzed in citrate–phosphate buffer (20 mM, pH 6.3) at 4 °C overnight using a 3 kDa dialysis tube (Vivascience, Hannova, Germany), and run through a HiTrapTM Q Sepharose XL anion exchange column (GE Healthcare, Munich, Germany) for purification. The purified recombinant TlGa15B and mutants were verified by SDS-PAGE using a 10% polyacrylamide gel.
Glucoamylase activity assay
The glucoamylase activity of TlGa15B and mutants was determined using soluble starch as the substrate. Reactions containing 900 μL of 1% soluble starch in citrate–phosphate buffer (pH 4.5) and 100 μL of appropriately diluted enzyme solution were incubated at 60 °C for 30 min, using the 3,5-dinitrosalicylic acid (DNS) method to detect the amount of reducing sugar in the reaction. One unit (U) of enzyme activity was defined as the amount of enzyme that released 1 μmol of glucose per minute.
Biochemical characterization of purified TlGa15B and mutants
The effects of pH on the activity of purified TlGa15B and mutants were determined in glycine–HCl (pH 1–3), citric acid-Na2HPO4 (pH 3–8), and glycine–NaOH (pH 8–12) buffers at 37 ℃. To determine pH stability, the purified TlGa15B and mutants were incubated in different pH (1–12) for 1 h at 37 ℃. The effect of temperature on activity was measured in citric acid-Na2HPO4 buffer (pH 4.5) at 20–90 ℃. To evaluate thermostability, the purified TlGa15B and mutants were incubated at 75 ℃ for different time periods (0–60 min).
Kinetic studies were performed in a citric acid-Na2HPO4 buffer (pH 4.5) at 55 ℃ using 1.0–10.0 mg/mL soluble starch as the substrates. The Lineweaver–Burk plot method was used to calculate the Km and Vmax values of the purified TlGa15B and mutants. The substrate specificities of TlGa15B were studied in terms of the hydrolysis of six different substrates, including soluble starch, amylose, glycogen, α-cyclodextrin, β-cyclodextrin, and γ-cyclodextrin in a citric acid-Na2HPO4 buffer (pH 4.5).
Differential scanning calorimetry (DSC) analysis
The melting temperatures (Tm) of TlGa15B and mutants were determined using MicroCal™ VP-Capillary DSC (GE Healthcare). The purified TlGa15B and mutants were dissolved in a 10 mM phosphate buffer (pH 7.4) as an infusion, and the protein concentration was controlled at 0.50 mg/mL. The temperature program was run in the range of 50–100 ℃, with a rising rate of 1 ℃/min and a scanning rate of 1℃/min.
Molecular dynamics (MD) simulation and dynamics cross-correlation matrix (DCCM) analysis
MD simulations were performed using the AMBER14 package at a temperature of 300 K for 20 ns with ff99SB force field. The closest distance between periodic box and atom was set 12 Å, and the time step was set to 2 fs. Before MD simulation, hydrogen atoms were added and any water molecules that did not interact with the protein were removed, and 20 mM sodium chloride was added to neutralize the charge in the system. The conjugate gradient method with α-carbon atoms restriction was used for energy minimization. The energy was minimized again without limiting the atoms, and finally the temperature was raised from 0 to 300 K. The MD trajectory data were analyzed using the CPPTRAJ software [36]. In addition, the last 10 ns trajectories from MD simulations were used to perform DCCM analysis using the Bio3D packages [37, 38].
Application of recombinant TlGa15B-GA2 in starch saccharification
To evaluate the industrial application potential of TlGa15B-GA2 in starch saccharification, TlGa15B-GA2 was compared with the commercial glucoamylase GA-LD from Shandong Longda Bio-products Co Ltd. The saccharification process of starch was carried out at 60 °C for 30, 40, 50, and 60 h, containing 50 U/g TlGa15B or GA-LD, 0.6 U/g pullulanase, and 200 g of liquefied starch. The enzymatic saccharification effect of starch was measured by monitoring the DX value (glucose content) of the saccharification solution.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- ETSS:
-
Enzyme thermal stability system
- RMSD:
-
Root mean square deviation
- RMSF:
-
Root mean square fluctuation
- PDB:
-
Potato dextrose broth
- DNS:
-
3,5-Dinitrosalicylic acid
- MD:
-
Molecular dynamics
- DCCM:
-
Dynamics cross-correlation matrix
References
Kumar P, Satyanarayana T. Microbial glucoamylases: characteristics and applications. Crit Rev Biotechnol. 2009;29:225.
Sauer J, Sigurskjold BW, Christensen U, Frandsen TP, Mirgorodskaya E, Harrison M, Roepstorff P, Svensson B. Glucoamylase: structure/function relationships, and protein engineering. Biochim et Biophys Acta (BBA) Protein Struct Mol Enzymol. 2000;1543:275–93.
Wang C, Yang L, Luo L, Tang S, Wang Q. Purification and characterization of glucoamylase of Aspergillus oryzae from Luzhou-flavour Daqu. Biotech Lett. 2020;42:2345–55.
Carrasco M, Alcaíno J, Cifuentes V, Baeza M. Purification and characterization of a novel cold adapted fungal glucoamylase. Microb Cell Fact. 2017;16:75.
Parashar D, Satyanarayana T: An insight into ameliorating production, catalytic efficiency, thermostability and starch saccharification of acid-stable α-amylases from acidophiles. Front Bioeng Biotechnol. 2018;6.
Lago MC, dos Santos FC, Bueno PSA, de Oliveira MAS, Barbosa-Tessmann IP. The glucoamylase from Aspergillus wentii: purification and characterization. J Basic Microbiol. 2021;61:443–58.
Karim KMR, Husaini A, Sing NN, Tasnim T, Mohd Sinang F, Hussain H, Hossain MA, Roslan H. Characterization and expression in Pichia pastoris of a raw starch degrading glucoamylase (GA2) derived from Aspergillus flavus NSH9. Protein Exp Purif. 2019;164:105462.
Tanaka Y, Konno N, Suzuki T, Habu N. Starch-degrading enzymes from the brown-rot fungus Fomitopsis palustris. Protein Exp Purif. 2020;170:105609.
Zheng Y, Xue Y, Zhang Y, Zhou C, Schwaneberg U, Ma Y. Cloning, expression, and characterization of a thermostable glucoamylase from Thermoanaerobacter tengcongensis MB4. Appl Microbiol Biotechnol. 2010;87:225–33.
He Z, Zhang L, Mao Y, Gu J, Pan Q, Zhou S, Gao B, Wei D. Cloning of a novel thermostable glucoamylase from thermophilic fungus Rhizomucor pusillus and high-level co-expression with α-amylase in Pichia pastoris. BMC Biotechnol. 2014;14:114.
Chen J, Zhang YQ, Zhao CQ, Li AN, Zhou QX, Li DC. Cloning of a gene encoding thermostable glucoamylase from Chaetomium thermophilum and its expression in Pichia pastoris. J Appl Microbiol. 2007;103:2277–84.
Kim M, Park J, Kim Y, Lee H, Nyawira R, Shin H, Park C, Yoo S, Kim Y, Moon T, Park K. Properties of a novel thermostable glucoamylase from the hyperthermophilic archaeon Sulfolobus solfataricus in relation to starch processing. Appl Environ Microbiol. 2004;70:3933.
Schmidt A, Shvetsov A, Soboleva E, Kil Y, Sergeev V, Surzhik M. Thermostability improvement of Aspergillus awamori glucoamylase via directed evolution of its gene located on episomal expression vector in Pichia pastoris cells. Protein Eng Des Sel. 2019;32:251–9.
Liu H, Wang W. Protein engineering to improve the thermostability of glucoamylase from Aspergillus awamori based on molecular dynamics simulations. Protein Eng Des Sel. 2003;16:19–25.
McDaniel A, Fuchs E, Liu Y, Ford C. Directed evolution of Aspergillus niger glucoamylase to increase thermostability. Microb Biotechnol. 2008;1:523–31.
Wang Y, Fuchs E, Silva RD, Mc Da Niel A, Seibel J, Ford C. Improvement of Aspergillus niger glucoamylase thermostability by directed evolution. Starch Strike. 2010;58:501–8.
Kurahashi R, Tanaka S, Takano K. Activity-stability trade-off in random mutant proteins. J Biosci Bioeng. 2019;128:405–9.
Siddiqui KS. Defying the activity–stability trade-off in enzymes: taking advantage of entropy to enhance activity and thermostability. Crit Rev Biotechnol. 2017;37:309–22.
Contreras F, Thiele MJ, Pramanik S, Rozhkova AM, Schwaneberg U. KnowVolution of a GH5 cellulase from Penicillium verruculosum to improve thermal stability for biomass degradation. ACS Sustain Chem Eng. 2020;8:12388–99.
Shivange AV, Roccatano D, Schwaneberg U. Iterative key-residues interrogation of a phytase with thermostability increasing substitutions identified in directed evolution. Appl Microbiol Biotechnol. 2016;100:227–42.
You S, Tu T, Zhang L, Wang Y, Huang H, Ma R, Shi P, Bai Y, Su X, Lin Z, et al. Improvement of the thermostability and catalytic efficiency of a highly active β-glucanase from Talaromyces leycettanus JCM12802 by optimizing residual charge–charge interactions. Biotechnol Biofuels. 2016;9:124.
Wang X, Ma R, Xie X, Liu W, Tu T, Zheng F, You S, Ge J, Xie H, Yao B, Luo H. Thermostability improvement of a Talaromyces leycettanus xylanase by rational protein engineering. Sci Rep. 2017;7:15287.
Gu Y, Zheng F, Wang Y, Su X, Bai Y, Yao B, Huang H, Luo H. Characterization of two thermophilic cellulases from Talaromyces leycettanus JCM12802 and their synergistic action on cellulose hydrolysis. PLoS ONE. 2019;14:e0224803–e0224803.
Li X, Xia W, Bai Y, Ma R, Yang H, Luo H, Shi P. A novel thermostable GH3 β-glucosidase from Talaromyce leycettanus with broad substrate specificity and significant soybean isoflavone glycosides-hydrolyzing capability. Biomed Res Int. 2018;2018:4794690.
Wang C, Luo H, Niu C, Shi P, Huang H, Meng K, Bai Y, Wang K, Hua H, Yao B. Biochemical characterization of a thermophilic β-mannanase from Talaromyces leycettanus JCM12802 with high specific activity. Appl Microbiol Biotechnol. 2015;99:1217–28.
Almagro Armenteros JJ, Tsirigos KD, Sønderby CK, Petersen TN, Winther O, Brunak S, von Heijne G, Nielsen H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat Biotechnol. 2019;37:420–3.
Nielsen BR, Lehmbeck J, Frandsen TP. Cloning, heterologous expression, and enzymatic characterization of a thermostable glucoamylase from Talaromyces emersonii. Protein Expr Purif. 2002;26:1–8.
Lee J, Paetzel M. Structure of the catalytic domain of glucoamylase from Aspergillus niger. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2011;67:188–92.
MacauleyPatrick S, Fazenda ML, McNeil B, Harvey LM. Heterologous protein production using the Pichia pastoris expression system. Yeast. 2005;22:249–70.
Xian L, Feng J. Purification and biochemical characterization of a novel mesophilic glucoamylase from Aspergillus tritici WZ99. Int J Biol Macromol. 2018;107:1122–30.
Hua H, Luo H, Bai Y, Wang K, Niu C, Huang H, Shi P, Wang C, Yang P, Yao B. A thermostable glucoamylase from Bispora sp. MEY-1 with stability over a broad pH range and significant starch hydrolysis capacity. PLoS ONE. 2014;9:e113581.
Karim KMR, Husaini A, Hossain MA, Sing NN, Mohd Sinang F, Hussain MHM, Roslan HA. Heterologous, expression, and characterization of thermostable glucoamylase derived from Aspergillus flavus NSH9 in Pichia pastoris. Biomed Res Int. 2016;2016:5962028–5962028.
Zhu B, Wang D, Wei N. Enzyme discovery and engineering for sustainable plastic recycling. Trends Biotechnol. 2021.
Zhang L, Tang X, Cui D, Yao Z, Gao B, Jiang S, Yin B, Yuan YA, Wei D. A method to rationally increase protein stability based on the charge–charge interaction, with application to lipase LipK107. Protein Sci. 2014;23:110–6.
Wang R, Wang S, Xu Y, Yu X. Enhancing the thermostability of Rhizopus chinensis lipase by rational design and MD simulations. Int J Biol Macromol. 2020;160:1189–200.
Roe DR, Cheatham TE. PTRAJ and CPPTRAJ: software for processing and analysis of molecular dynamics trajectory data. J Chem Theory Comput. 2013;9:3084–95.
Skjærven L, Yao X, Scarabelli G, Grant BJ. Integrating protein structural dynamics and evolutionary analysis with Bio3D. BMC Bioinformat. 2014;15:399.
Grant BJ, Rodrigues APC, ElSawy KM, McCammon JA, Caves LSD. Bio3d: an R package for the comparative analysis of protein structures. Bioinformatics. 2006;22:2695–6.
Acknowledgements
Not applicable.
Funding
This research was supported by the National Key Research and Development Program of China (2021YFC2100400), the State Key Laboratory of Animal Nutrition Project (2004DA125184G2101), and the China Agriculture Research System of MOF and MARA (CARS-41).
Author information
Authors and Affiliations
Contributions
BY and HYL conceived and designed the experiments. LGT, JZ, and XW performed the experiments. XLW, HQH, HMY, and TT analyzed the data. YW and YGB contributed analytic tools. LGT and XQ wrote the manuscript. HYL and XQ reviewed and revised the manuscript. All the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1
: The Ramachandran plot of homology modeling for TlGa15B.
Additional file 2
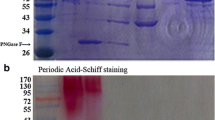
: SDS-PAGE analysis of the recombinant TlGa15B and mutants. Lane 1, 3, 5, the culture supernatant of transformants TlGa15B, TlGa15-GA1, and TlGa15-GA2; lane 2, 4, 6, the purified TlGa15B, TlGa15-GA1, and TlGa15-GA2.
Additional file 3
: The thermostability of the purified recombinant TlGa15B.
Additional file 4
LC-MS/MS analysis of the formation of disulfide bonds in the TlGa15B mutants. a: the disulfide-linked peptide SNPSGGLCT/SASGPCA (132C/492C); b: the disulfide-linked peptide PLWYCIV/SAIPCSA (548C/562C).
Additional file 5
: The modeled structure of TlGa15B-GA2. Six candidate mutated sites away from the catalytic center were indicated in blue balls. Two disulfide bonds were indicated as yellow sticks.
Additional file 6
: The specific activity of TlGa15B and its mutants in the temperature range from 20 to 90 ℃.
Additional file 7
: Primers used in this study.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Tong, L., Zheng, J., Wang, X. et al. Improvement of thermostability and catalytic efficiency of glucoamylase from Talaromyces leycettanus JCM12802 via site-directed mutagenesis to enhance industrial saccharification applications. Biotechnol Biofuels 14, 202 (2021). https://doi.org/10.1186/s13068-021-02052-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13068-021-02052-3