
Abstract
Choroid plexus (ChP) epithelial cells are crucial for the function of the blood-cerebrospinal fluid barrier (BCSFB) in the developing and mature brain. The ChP is considered the primary source and regulator of CSF, secreting many important factors that nourish the brain. It also performs CSF clearance functions including removing Amyloid beta and potassium. As such, the ChP is a promising target for gene and drug therapy for neurodevelopmental and neurological disorders in the central nervous system (CNS). This review describes the current successful and emerging experimental approaches for targeting ChP epithelial cells. We highlight methodological strategies to specifically target these cells for gain or loss of function in vivo. We cover both genetic models and viral gene delivery systems. Additionally, several lines of reporters to access the ChP epithelia are reviewed. Finally, we discuss exciting new approaches, such as chemical activation and transplantation of engineered ChP epithelial cells. We elaborate on fundamental functions of the ChP in secretion and clearance and outline experimental approaches paving the way to clinical applications.
Similar content being viewed by others


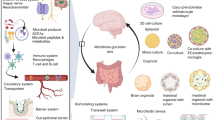
Introduction
The choroid plexus (ChP) comprises a set of epithelial sheets sandwiching stromal cells and vasculature located in the lateral (LV), third (3V), and fourth (4V) ventricles of the brain. The ChP has several vital roles in the central nervous system (CNS). First, the ChP forms the blood-cerebrospinal fluid barrier (BCSFB), which is important for protecting the CNS from peripheral challenges including inflammation, pathogens, toxins [1,2,3,4]. Second, the ChP produces cerebrospinal fluid (CSF) into which it secretes nutrients and signaling molecules [5,6,7,8,9,10]. Third, it can perform CSF clearance functions including the removal of amyloid-beta [11] and ions (e.g. K+) under certain conditions [12]. In these capacities, the ChP-CSF system is an active player in brain development and lifelong brain health. ChP epithelial cell dysfunction is associated with several neurodegenerative conditions [1, 13,14,15,16]. Therefore, manipulating gene expression and functions of the ChP is appealing from basic science as well as therapeutic perspectives. As relatively long-lived cells in the CNS, ChP epithelial cells provide an attractive platform for gene therapy, scalable delivery of health-promoting factors for the brain, tissue engineering, and transplantation [5, 17, 18].
Many techniques have been applied to target ChP epithelial cells. For example, gene knockdown using small interfering RNAs (siRNA) can be used to silence gene expression in primary cultures of ChP epithelial cells and in vivo in mice [19,20,21]. In utero electroporation provides a powerful and rapid approach to deliver plasmid expression vectors for overexpression or knockdown studies in target cells and has been broadly used in the developing brain (e.g., cerebral cortex) [22]. Since the ChP is uniquely positioned in each brain ventricle and electroporation is directional [22], only a select population of ChP cells can be targeted by this approach at any time. However, electroporation of the embryonic 4V ChP has proven remarkably successful [19]. To effectively reach all ChP tissues in all ventricles, viral vectors and transgenic approaches have emerged as preferred strategies. Vectors including adeno-associated virus (AAV), adeno- and lentiviruses are commonly employed as carriers for gene delivery into the brain via intracerebroventricular (I.C.V.), intrathecal (I.T.), or intravenous (I.V.) routes. These techniques lend helpful temporal and spatial control to investigating ChP functions. Transgenic mice can be readily leveraged to delete or overexpress genes of interest in ChP epithelial cells.
Here, we review current experimental approaches available for targeting ChP epithelial cells, with a focus on in vivo strategies. We discuss the use of genetic reporter lines to visualize the targeted ChP, as well as the viruses used to target these cells for manipulation. We touch on the application of chemogenetic approaches using genetically engineered receptors to modulate activity of ChP epithelial cells. Finally, we provide a brief summary of ChP cell-targeted grafts and transplantation studies and their application to repair the damaged CNS.
Genetic tools for targeting ChP
This section covers genetic approaches including traditional transgenic mouse models, inducible systems, cell-specific systems for loss or gain of gene function in ChP epithelial cells, and genome editing tools.
Reporter mouse lines
Reporter mice can facilitate tracing cell lineage and performing cell morphological analyses. They may also prove to be useful in the emerging ChP field of transplantation to evaluate cell integration and survival (see below). Reporter lines that enable tracking of labeled ChP epithelial cells have been established by leveraging the most highly expressed ChP signature gene, Transthyretin (Ttr) [23, 24]. TTR is secreted by ChP epithelial cells into the CSF, where it functions as a carrier protein for thyroid hormone (thyroxine4) [25]. An early Ttr::RFP reporter transgenic mouse was created with red fluorescent protein (mRFP1) [26]. Although broadly expressed across ChP epithelial cells during early embryonic development, Ttr::RFP transgene expression is lost in a fraction of ChP cells during later embryonic stages in this line. Labeling takes on a mosaic pattern in ChP, and expression is sparse postnatally [24, 27]. To achieve more complete postnatal expression, a human TTR BAC (bacterical artificial chromosome)-tdTomato mouse line was developed in which Ttr itself is replaced by tdTomato. In this line, robust tdTomato expression appears during early embryogenesis and remains stable well into adult life (e.g., up to 10 months) [27].
The transcription factor Forkhead box J1 (FoxJ1) can also be leveraged to target ChP cells. FoxJ1 is expressed by multi-ciliated cells throughout the body [28]. In the pre-natal mouse brain, ChP epithelial cells are the sole multi-ciliated cells contacting the ventricles. However, they lose this privileged status soon thereafter when multi-ciliated ependymal cells differentiate from radial glia and mature in the week following birth [29, 30]. Thus, depending on the experimental question, FOXJ1-Cre transgenic mice, in which the human FOXJ1 promoter drives Cre recombinase expression, can be an excellent tool for studying ChP epithelial cells [28] (Table 1). When expressed, Cre catalyzes site specific recombination between two loxP (locus of crossing over in bacteriophage P1) sites [31,32,33]. Thus, Cre will excise DNA at two loxP sites, deleting the intervening DNA sequence [34]. FOXJ1-Cre mice can be crossed with a double-knock-in line in which all cells express a membrane bound form of tdTomato (mT) at baseline, and in the presence of Cre recombinase, the mT cassette is excised, enabling expression of the mG (GFP) cassette [35]. In this way, mT/mG mice crossed with FOXJ1-Cre transgenic mice reveal GFP-positive ChP epithelial cells (Fig. 1) [24].

Representative GFP expression in LV ChP cells of wild type and mT/mG;FOXJ1-Cre mice. A mT/mG;FOXJ1-Cre reporter mice show GFP positive ChP epithelial cells immunostained with anti-GFP (green) antibodies and stained with Hoechst (blue) to mark nuclei. Scale bar, 10 μm [24]. Flow cytometry analyses of LV ChP cells of wild type (control, B) and mT/mG;FOXJ1-Cre adult mice C indicate that more than 40% of cells are mG-positive in mT/mG;FOXJ1-Cre adult mice. Experiment conducted as in [35], where recombination pattern of this reporter mouse was examined in lymphoid organs, thymus, and spleen
The FOXJ1-Cre transgenic system has also been used to trigger gene expression in embryonic ChP epithelial cells to facilitate a variety of experimental paradigms. For example, EGFP tagged ribosomal subunit L10a can be expressed in ChP epithelial cells by crossing FOXJ1-Cre mice with EGFP:L10a BAC mice [12]; combined with tissue dissection, this approach enables translating ribosome affinity purification (TRAP) studies [36]. In another example, ChP epithelial cells can be targeted by crossing FOXJ1-Cre mice with Ai95D mice [37], a calcium reporter GCaMP6f line, to observe calcium activity in ChP epithelial cells [38].
Reporter zebrafish lines
Due to its large size and semi-transparent embryos, Danio rerio (zebrafish) provides another powerful vertebrate model for studying the brain’s ventricular system and ChP development [39]. Several brain pathologies associated with human diseases can also be modeled in zebrafish (e.g., autism spectrum disorder, depression, and Parkinson's disease) [40, 41]. A better understanding of ChP involvement may shed light on the molecular mechanisms of different neurological disorders and establish further tools for drug development [40,41,42]. In contrast to mammals, the zebrafish ChP is located close to the dorsal surface of the brain and is therefore experimentally more accessible. Thus, non-invasive in vivo live imaging is relatively straight-forward, making it possible to observe ChP development [43]. Early studies reported enhancer trap transgenic lines that express GFP in the zebrafish ChP [43,44,45], enabling the study of signaling molecules during development. One of these lines, the Et(cp:EGFP)sj2 line, which expresses GFP under the control of the epithelia-specific keratin4 promoter, results in targeted GFP expression in diencephalic and myelencephalic ChP epithelial cells that correspond to 3V and 4V ChP in the mammalian brain, respectively. GFP expression in this line begins at the larval stage and is retained at least until 1 month of age [46]. Barrier proteins such as Claudins can also be harnessed to mark the ChP. While expressed more broadly, claudin5-GFP zebrafish provide another good marker of the ChP barrier [47]. Because the ChP has a very distinctive, fenestrated vasculature, using Plvap, a gene required for formation of fenestrated capillaries [(Tg(plvap:EGFP) lines)], or the Tg(kdrl:EGFP) endothelial cell-specific reporter line, have also proven effective for visualizing the tissue for imaging studies [48].
Genetic gain- and loss-of-function in mouse ChP
In studies using transgenic mice, various promoters can be used to disrupt gene expression in ChP epithelial cells, and the most common ones are summarized in Table 1. The FOXJ1-Cre line [28] represents one of the most widely used approaches to date. For example, the circadian rhythm of ChP epithelial cells was tested by deleting the essential circadian clock gene Bmal1 in ChP (and all other multi-ciliated cells in the body) by crossing FOXJ1-Cre transgenic mice with floxed Bmal1 mice (Bmal1fl/fl) [49]. Conditional knockout strategies can enable temporal and spatial control, which is necessary to determine the role(s) of specific target genes in certain locations during development. For example, Wnt1-Cre lines can be used to target 4V ChP where Wnt1-Cre deletes Sonic hedgehog (Shh) in the Shhfl/fl line [50, 51]. However, due to findings that Wnt1-Cre transgenic mice show ectopic upregulation of Wnt1 expression and unintended midbrain phenotypes, a newer, Wnt1-Cre2 has been generated [52]. This new Wnt1-Cre2 line has been used to delete Meis1 in 4V ChP in a Meis1fl/fl line [53]. Other cases of a 4V ChP conditional knockout of Sox9 can be achieved with Pax2-Cre mice [19]. In addition, the lymphotropic papovavirus control region (LPVcr) allows generation of ChP conditional knockout by the Cre-recombinase/loxP system (e.g., LPV-Cre.0607 transgenic line) [54].
Cre-based approaches can also be harnessed for gain-of-function studies and to model diseases of the CNS including ChP carcinoma [55,56,57,58]. While ChP epithelial cells differentiate from the roof plate and neuroectodermal lineage along the neural tube [59], they are not typically considered part of the Nestin lineage [60, 61]. Accordingly, Nestin-Cre/RosamTmG reporter mice show limited recombination (GFP-positive cells) in ChP epithelium during embryonic brain development [56]. However, following birth, unexpected recombination occurs, first in ventral regions of the 4V ChP, followed by recombination throughout each ChP in each ventricle of the brain [56]. By the end of the first postnatal week, Cre recombination recognized by RosamTmG occurs throughout each ChP in this line [56]. This postnatal ChP Nestin-Cre expression has been paired with StopFLMYC mice [62], in which a floxed stop cassette [loxP-stop-loxP] is located between a promoter and the gene of interest [63], the tumor oncogene MYC. Cre-mediated excision of the stop cassette drives human c-MYC overexpression in ChP epithelial cells, resulting in devastating ChP tumors, matching WHO Grade III carcinoma classification used in the clinical setting. These tumors form predominantly in the posterior domain of the LV ChP and 4V ChP [56]. ChP tumor models can also be generated when pairing MYC overexpression with Trp53 deletion under the Atoh1-Cre line [57].
Tamoxifen inducible knock-in approaches can further refine temporal dynamics of Cre-based strategies. In this approach, Cre recombinase is fused to a mutant form of the estrogen receptor ligand binding domain (Cre-ERT2) and localizes in the cytoplasm [64]. Upon tamoxifen binding, Cre translocates to the nucleus where it catalyzes recombination [65]. In the ChP, the mouse Foxj1 promoter-driven CreERT2 system (Foxj1CreERT2::GFP) [66] induced by tamoxifen at E10.5-E12.5 can be used for conditional ablation of gene expression during the early stages of ChP development, as was done with Wnt5a in 4V ChP epithelial cells [53, 67], resulting in altered morphogenesis of 4V ChP. To elucidate the critical role of transcription factor Otx2 in the development and maintenance of the ChP, Otx2-CreERT2 mice were generated by replacing the genomic region spanning the Otx2 coding sequence, and then crossed with Otx2fl/fl mice [68, 69].
Another elegant loss-of-function technique entails I.C.V. delivery of TAT-Cre, a recombinant fusion protein of Cre and the cell-permeable TAT sequence, that is readily taken up by mammalian cells and catalyzes recombination [70]. When TAT-Cre is delivered into adult ventricles, it is taken up by ChP epithelial cells. In Otx2fl/fl mice, TAT-Cre results in ChP-Otx2 deletion, resulting in altered cortical plasticity and neurogenesis [71,72,73]. Similarly, TAT-Cre injection into adult Appfl/fl mice induces App knockdown in ChP, with downstream consequences on adult neurogenesis [74].
Tetracycline-controlled gene expression systems (e.g., Tet-ON) are commonly used to regulate gene expression and can be applied to the ChP [75]. In this approach, a gene of interest is placed under the regulatory control of the transcriptional activator rtTA (reverse tetracycline-controlled transactivator) and its Tet response element (TRE). Tetracyline (or a derivative such as Doxycycline, Dox) binding to rtTA induces a conformational change that allows rtTA to bind the TRE and induce expression of the target gene of interest [76]. To investigate Shh overexpression in ChP, transgenic lines overexpressing rtTA were generated under the Ttr and Otx2 promoters (pTtr-rtTA and pFuguOtx2-rtTA) [75]. These mice were then crossed with pTRE-mShh/d2EGFP transgenic mice [77], in which Shh and GFP were expressed in the presence of Dox. While ChP-Shh expression is typically restricted to the 4V ChP in mice [23, 24], Dox administration to pregnant dams resulted in robust ChP-Shh mRNA expression in each ventricle’s ChP and expanded ChP growth [75]. ChP-Shh expression was also accompanied by Gli1 and Gli2 expression in cerebral cortical progenitors lining the CSF-filled ventricles, enlarged ventricles, and disrupted cerebral cortical development [75]. One limitation of the pTtr-rtTA line is that exhibits progressively diminished rtTA expression postnatally [75], similar to the Ttr::RFP line (see above [24]). However, pFuguOtx2-rtTA transgenic mice provide a suitable alternative with sustained rtTA expression postnatally, resulting in increased neural stem cell proliferation in the subventricular zone [75].
In addition to gain- and loss-of-function gene expression studies, mouse genetics approaches can be harnessed for ChP ablation. For example, Gdf7 lineage cells are localized to the anterior domain of the LV ChP. Therefore, Gdf7-driven diphtheria toxin A chain expression can be used to ablate the anterior domain of the LV ChP [78]. Intriguingly, because the posterior domain of the LV ChP forms in a contiguous field with the anterior domain, the posterior domain also fails to form following anterior endotoxin expression [78]. Some mouse lines also may fail to develop a ChP, as is the case in the extra-toesJ (XtJ), which carries a Gli3 deletion and fails to develop a LV ChP [79] as well as Otx2-deficient mice [68], where all ChP were affected when gene deletion was induced at E9.
New tools for genetic manipulation: CRISPR/Cas9
While most of the mouse lines discussed above were generated by traditional transgenic approaches, the advent of CRISPR/Cas9 methods has revolutionized mouse transgenesis [80, 81]. These newer strategies can relatively quickly introduce gene disruptions in mammalian cells. For example, use of the Easi-CRISPR (Efficient additions with ssDNA inserts-CRISPR) targeting strategy enables progress from “concept” to “F1 founder mouse” in as little as three months [82]. It succeeds by leveraging injection of long single-stranded DNA donors with pre-assembled ribonucleoprotein complexes with two guide RNAs into mouse zygotes [82]. The CRISPR/Cas9 approach was used to study serotonin receptor signaling in ChP epithelial cells [38]. Antibodies for the 5-HT2C receptor are often unreliable, and hence, the addition of a fluorescent mRuby tag to the receptor in the Htr2cmRuby line provides an opportunity to visualize the subcellular localization of this receptor on the apical and basal surfaces of ChP epithelial cells [38]. The ease of CRISPR/Cas9 approaches has the potential to generate large-scale genetics resources for the field. Applications now range from zebrafish, rats, and pigs to primates, opening avenues to unprecedented opportunities for CSF research [83, 84].
Limitations of mouse genetic model systems
Genetic model systems provide powerful and enduring tools for ChP targeting but are not flawless. One of the inherent challenges to using transgenic mice in the study of the ChP is that tissue and cell-type specificity are dependent on promoter specificity. While it has been desirable to find ChP-specific Cre drivers for selective targeting of ChP epithelial cells (e.g., LPV-Cre.0607 [54]), single cell transcriptomics studies suggest no single gene is likely to be expressed only in ChP epithelial cells compared to all other cells throughout the body [23]. Foxj1-Cre transgenic lines are extensively employed for transgene expression in multi-ciliated cells including ChP epithelial cells. However, as mentioned above, the FoxJ1 promoter also drives expression in other ciliated epithelial cells in the brain and body (e.g., ependymal cells) [28]. Thus, CreERT2-mediated Otx2 silencing occurs not only in the ChP but also in other parts of the brain [68]. Workarounds to achieve greater specificity include intersectional Cre approaches and direct I.C.V. delivery of Cre-expressing viruses such as AAV-Cre (see below). Limitations notwithstanding, genetic model systems provide highly valuable resources that enhance the study of the development of ChP epithelial cells and will continue to be optimized for investigating fundamental ChP biology.
Viral approaches for targeting ChP
The ability to manipulate genes with viral vectors has introduced a new flexibility for the study of the ChP. Long-term breeding can be avoided, and target gene expression in ChP epithelial cells can be obtained as soon as 24–48 h following vector introduction, making it possible to study acute effects of target gene manipulation in, for example, embryonic mice. Vector-based gene manipulation is particularly powerful when combined with transgenic technology as evidenced primarily by Cre-Lox experiments. Here, we review the DNA- or RNA-based viral vectors (e.g., AAV, adeno- and lentiviral vector systems) typically used for targeting ChP epithelial cells and then discuss the most used routes of viral administration at embryonic and adult ages (Fig. 2).

Commonly used viral delivery approaches for targeting mouse developing and adult ChP. Schematic of in utero I.C.V. A and ultrasound-guided in utero I.C.V. B delivery strategies for pre-natal mice. Schematic of adult stereotactic I.C.V. delivery C and various I.V. delivery routes in neonatal and adult D mice. Figure created with BioRender.com
Adeno-associated virus (AAV)
AAV vectors have cell type, tissue, and organ tropism depending on their serotype. In addition to tropism, viral titer, delivery route, animal genetics, and age also influence AAV gene expression. AAV2 is widely used to target the CNS but has not been noted to successfully transduce the ChP [85, 86]. By contrast, the hybrid serotype AAV2/5, which incorporates the genome of AAV2 and the capsid of AAV5, has proven tropism for ChP epithelial cells and produces robust expression (Table 2). Embryonic I.C.V. delivery of AAV2/5 is followed by ChP transduction in as soon as two days ([87] and Fig. 3), and gene expression can be sustained for months [88]. AAV2/5-mediated ChP gene expression in adult mice has been reported to occur within weeks of injection [89] and persists up to 12 months [74]. Other serotypes have been tested in several conditions with variable outcomes (see Table 2 and [89]); none have been as successful as AAV2/5. Importantly, genes introduced with AAV2/5 encode functional and demonstrably therapeutic proteins. For example, overexpression of the protein ATP7A with an AAV2/5 vector (AAV2/5-rsATP7A) mitigates the Menkes disease phenotype involving copper metabolism in the Atp7a knockout mouse model [90]. Overexpression of the Na+-K+-Cl− co-transporter, NKCC1 [91], in developing ChP can modulate CSF-K+ levels and have long-term impact on ventricular size [12].

GFP expression in LV ChP following in utero I.C.V. delivery of AAV2/5-GFP. AAV2/5 delivered at E13.5 and immunostained 5 days later at E18.5 with anti-GFP (green) and anti-AQP1 (red) antibodies. Tissue stained with Hoechst (blue) to mark nuclei. Inset (dashed line box) highlights detection of AAV2/5-GFP and AQP1 expression in the ChP epithelium. Experiment conducted as in [53]
As previously mentioned, mouse age and virus titer also influence AAV transduction. AAV2/5 delivered to neonates can transduce not only the ChP but also newborn ependymal cells lining the ventricles [92]. I.T. AAV2/5 delivery in adult mice leads to robust ChP transduction in each ventricle with the 4 V ChP having the highest expression, followed by the 3V ChP, and finally the LV ChP. This effect is likely related to a concentration gradient of virus exposure [93]. Consistent with this idea, the epithelial cells at the tips of the villi of the 3V and 4V ChP that are bathed by the CSF show higher rates of transduction, compared to epithelial cells at the base of the villi (see Supp Fig. 7D in [53]). While I.C.V. delivery of AAV2/5 shows tropism for ChP epithelial cells, I.T. injections reach several areas throughout the brain, with labeling including cerebellum, hippocampus, midbrain and olfactory bulb [93].
AAVs are widely used to deliver shRNA for knockdown studies or Cre recombinase to transgenic mice engineered with loxP sites for conditional gene deletion. For example, Chd4 deletion in ChP epithelial cells can be achieved by in utero I.C.V. delivery of AAV2/5-Cre into Chd4fl/fl mice [12]. Similarly, I.C.V. delivery of AAV2/5-Cre into adult Klothofl/fl mice deletes Klotho expression in ChP [94]. Remarkably, when delivered I.C.V., conditional gene deletion in ChP epithelial cells can be obtained without disrupting the same gene’s expression in nearby hippocampus [74]. AAV2/5-harboring Otx2 shRNA injected I.C.V. into adult mice can also be used for Otx2 knockdown in the ChP for at least three weeks following injection [73]. ChP-specific inhibition of miR-204 was achieved using AAV2/5 following adult mouse I.C.V. delivery [95].
AAV transduction can also be combined with CRISPR/Cas9 genome editing technology, as has been done with in utero I.C.V. injection of AAV2/9-PHP.eB-expressing gRNAs [96]. This approach achieves widespread, efficient gene targeting in the developing brain (e.g., Fig. 1C in [96]), which can be leveraged for gene deletion. Combining experimental strategies in this manner opens avenues for generating rapid knockout of ChP genes of interest and also helps address cell type specificity issues common to Cre-mediated approaches throughout the body.
Recent successes of AAV-based approaches that target neurons in brain regions affected by CNS diseases have ushered in an era of new hope that extends to the ChP and CSF [97]. Efforts are being taken to generate AAV variants with higher efficiency to cross the blood–brain barrier (BBB) and BCSFB so that they may reach the brain through the bloodstream. This strategy would also enable less invasive I.V. rather than I.C.V. injections for delivering genes to the ChP. The Cre recombination-based AAV targeted evolution (CREATE) strategy enables the development of AAV capsids that more efficiently transduce defined Cre-expressing cell populations in vivo [98,99,100]. For example, the AAV-PHP.B variant, generated using CREATE, transfers genes to target cells throughout the CNS with remarkably improved efficiency [98]. However, strain- and species-specific differences must always be considered. AAVs use multiple cellular receptors for attachment, internalization, and intracellular trafficking [101]. The AAV-PHP.B variant [98, 102] requires expression of the Ly6a receptor [102], revealing one host factor that underlies tissue tropism for this AAV variant. Because Ly6a is differentially expressed at the BBB of various mouse strains (e.g., present in C57BL/6 J but absent in BALB/cJ mice), its expression can pre-determine if peripherally injected AAV-PHP.B will cross the BBB. Accordingly, next-generation AAVs aim to demonstrate peripheral-to-CNS transfer and tropism to target cells of interest, so that systemic delivery routes can be used to lessen procedure invasiveness.
AAV vector safety and clinical efficacy have been demonstrated by promising Phase I/II/III clinical trials in various human disease settings, including lysosomal storage disorders (LSD) [103] and spinal muscular atrophy (SMA) [104,105,106]. Nonetheless, AAVs are known to trigger immune responses. Thus, off-target inflammatory effects should be considered in each experiment. Clinically, corticosteroids are often used to suppress unintended immune responses. In the case of AAVs, the genome of the vector can activate Toll-like receptor 9 (TLR9), which recognizes foreign DNA [107]. AAV vectors that are intrinsically less immunogenic have consequently been designed; these vectors incorporate short DNA oligonucleotides that antagonize TLR9 activation. This approach reduces innate immune and T-cell responses and has been tested in tissues including liver, muscle, and retina but may not be adequate for all immune responses of concern. A recent study also suggests that AAV transduction may be damaging to hippocampal stem cells residing in the dentate gyrus [108]. To our knowledge, hippocampal toxicity has not been reported with the AAV strategies used for targeting the ChP. However, all these points are important considerations for broadening therapeutic windows for AAV therapies and other DNA-based gene transfer methods.
Adenovirus
Similar to AAV vectors, adenoviral approaches confer notable advantages of long-term gene expression and reduced toxicity. However, adenoviruses tend to transduce a broader population of cells; for instance, adenoviral vectors delivered into adult rat ventricles revealed transduction in the ependymal cell layer and cervical spinal cord [109]. Moreover, while helper-dependent adenoviral (HDAd) vector injected into adult mice intrathecally exhibits ChP transduction as early as two days following injection, brain ependyma and other regions are also transduced and express the transgene [110].
Lentivirus
Lentiviral vectors offer an alternative and robust strategy for introducing proteins or peptides into the CSF via the ChP. One example comes from I.C.V. delivery of the vesicular stomatitis virus (VSV-G) vector into neonates, which results in transduction and long-term gene expression in ChP and ependymal cells [92]. Another valuable example comes from studies of Klotho, a transmembrane protein expressed by ChP epithelial cells and implicated in aging [111]. Sustained Klotho expression (for several months) was achieved with I.C.V. injected lentivirus in mouse models of Alzheimer's disease and cerebral ischemia [111,112,113]. In contrast to overexpression studies, lentiviral vectors can also produce loss-of-function by delivering shRNA. For example klotho shRNA-harboring lentivirus delivered I.C.V. into adult rats caused klotho knockdown in the ChP [111]. ChP-Megalin levels can also be modulated by lentiviral overexpression and RNA interference [21]. However, lentivirus ChP transduction is reported to be somewhat variable in adult mice [89].
To achieve greater cell-type specificity, two complementary lentiviral vectors can be used to achieve overexpression in ChP epithelial cells in a Tet-ON approach (similar to mouse genetic approach, see above). In this case, the effector virus contains a promoter that drives expression of rtTA and the GFP reporter [114]. The target construct includes the TRE DNA sequence, upstream of the gene of interest, followed by the RFP reporter. Expression of the target transgene is induced in the presence of Dox, which is known to cross the BBB (see Fig. 1 in [114]). Following I.C.V. delivery, replication-defective lentiviruses containing the effector and target integrate stably into the ChP epithelial cell genome for long-term expression. Mice provided with Dox in their drinking water then have expression of the target gene of interest. A more ChP-specific promoter for the brain (albeit with expression in other tissues throughout the body) was generated by isolating the 5’ flanking region of the corticotropin releasing factor receptor type 2 beta gene (CRFR2β) [114]. This lentiviral approach can result in expression of biologically active neuropeptides (e.g., corticotropin-releasing factor and gonadotropin-releasing factor) in the ChP, which are secreted into the CSF to great effect [114]. Gene expression can be induced as soon as two hours following Dox administration. The use of different ChP-brain-specific promoters may provide opportunities for fine-tuning transgene expression levels.
Embryonic in utero I.C.V. injection
Various injection ages during development are reported, depending on the purpose of studies. In utero I.C.V. injection is widely used for delivering genes to target tissue. Following laparotomy, the virus is injected into the LV or 4V with a glass capillary tube or Hamilton syringe driven by a microinjector. The virus rapidly distributes throughout the ventricular system, transducing epithelial cells that contact CSF in each ventricle’s ChP. This approach is typically performed after the ChP has developed into a clearly visible anatomical structure (~ E12–E16) in order to provide sufficient numbers of epithelial cells for transduction [12, 53, 87, 88]. This time frame also corresponds to peak neurogenesis in the cerebral cortex [115]. However, tradeoffs need to be evaluated for each experimental age. At younger ages closer to E12, the injection procedure is easier owing to larger ventricles and limited surrounding brain tissue. In contrast, more ChP is available for transduction at E16, but the injection procedure becomes increasingly challenging due to the increased growth of the brain, skull, and more restricted access into the ventricles. Pairing I.C.V. injection with ultrasound imaging enables targeted viral delivery into even younger embryonic ventricles (E10-E11) to target ChP cells as they emerge. This age is more challenging to successfully target by eye due to the small size of the embryos and the opacity of the extraembryonic membranes, including the decidua. Ultrasound-guided viral delivery was recently optimized for E10.5 studies of early stages of ChP development [53].
Adult I.C.V. delivery
I.C.V. or I.T. administration of gene vehicles can also effectively deliver viruses throughout the adult ventricular system. A stereotactic injection method is used for I.C.V. delivery for the adult ChP. Following head stabilization using a stereotactic frame, the injection site is determined by a designated 3-dimensional coordinate system of anterior–posterior (AP), medial–lateral (ML), and dorsal–ventral (DV) axes that aims for the lateral ventricle or cisterna magna. Example coordinates used for an adult (4–16 weeks) C57BL/6 mouse lateral ventricle are: 0.4 mm from bregma (AP), 1.0 mm (ML) and 2.0 mm (DV) from the brain surface [89, 116]. However, it is important to note that these coordinates may differ based on postnatal mouse age and strain. This approach has been successfully used in mice ranging from 8 to 16 weeks of age [74, 89], with enduring target gene expression up to and beyond 12 months [74].
Intraventricular delivery can be achieved by cisterna magna injection, lumbar puncture, or intrathecal catheter insertion. In mice, cisterna magna injection is the most commonly used intrathecal route. For example, helper-dependent adenoviral (HDAd) vector can be injected into the cisterna magna of 8–12-week-old C57BL/6 mice, and transgene expression can be observed in the ChP two days post-injection [110]. Alternatively, direct lumbar puncture of AAV into C57BL/6 mice shows clear transduction of each ChP throughout the ventricles along with various additional brain regions [93]. In general, the transduction pattern of CNS regions by I.T. injection reveals widespread and varying distribution that follows a caudal to rostral gradient of transduction.
The above procedures are all highly invasive. In an attempt to target other regions of the brain using less invasive approaches, both intranasal and I.V. approaches have been developed [97, 117,118,119,120]. However, these approaches require larger volumes of virus. New, more selective and potent viruses (such as the AAV-PHP.B as mentioned above) are being developed to circumvent these issues and will need to be tested on the ChP.
Other approaches for targeting the ChP
Bioluminescence for studying the ChP in vivo
Understanding the functions of the ChP in vivo requires the ability to experimentally modulate its constituent cells, ideally by non-invasive means. Pharmacological and chemogenetic manipulations may provide innovative new strategies, taking advantage of unique direct chemical access to this brain structure through circulating blood and CSF [121].
The BioLuminescent-OptoGenetic (‘BL-OG’) [122, 123] method takes advantage of light that is produced by a chemical reaction within a cell when an enzyme (luciferase) oxidizes a small molecule (luciferin). While this approach is now established in neurons [122, 123], it has recently been adapted to the ChP [121]. In this approach, a bioluminescent reporter is expressed in ChP epithelial cells. A reporter molecule that expresses an optogenetic element is tethered to the bioluminescent luciferase enzyme. For example, the molecule LuMinOpsin3 (LMO3), a fusion of a bright and small luciferase enzyme (Gaussia; GLuc) and the light-activated Volvox channelrhodopsin (VChR1) [123,124,125,126], can be used. When GLuc binds its small molecule driver, the exogenously injected luciferin coelenterazine (CTZ), photons are released. The light is in turn absorbed by the VChR1, leading to a conformational change and allowing ions to diffuse down their concentration gradients. In neurons, this would lead to depolarization and firing of action potentials. Depending on the strength of the activation, the resting membrane potential of ChP epithelial cells can presumably be influenced transiently by this same approach, possibly with downstream effects on calcium-dependent cellular processes.
In ChP epithelial cells, luciferase expression can be driven by crossing a Cre line of choice (e.g., the FOXJ1-Cre mouse [28] with the LSL-Lmo3-eYFP [127] reporter mouse). These crosses strongly express LMO3 in the ChP, and bioluminescent signals can be imaged using a CCD camera via an implanted cannula. Luciferin cofactors can be administered by I.C.V., I.V., or intraperitoneal (I.P.) injection routes, providing readouts of the distinct pharmacokinetics associated with each delivery route. This approach therefore also sets the stage for future studies involving drug delivery.
The positioning of the ChP deep within the brain ventricles makes commonly used optogenetic methods with external light challenging [128]. The BL-OG approach provides an appealing workaround to that issue. It also provides optical confirmation, through bioluminescent light, of chemogenetic engagement on the target cell. In addition, the use of bioluminescence rather than fluorescence removes issues related to fluorophore bleaching and autofluorescence, thereby ensuring accuracy of estimates and providing a signal that is potentially detectable using non-invasive imaging methods.
ChP tissue engineering and transplantation
Gene therapy paired with cell and tissue transplantation may provide a viable approach for future biological and clinical applications. Indeed, prior cell transplantation studies have suggested the therapeutic potential of ChP epithelial cells for brain injury and disease [129, 130]. For example, early studies hinted that transplanted ChP cells may have neuroprotective properties, especially in neurodegenerative disease models [131,132,133]. Human and mouse ChP epithelial cells can be derived from embryonic stem cells in response to bone morphogenetic protein 4 (BMP4), and these cells have self-assembling, secretory properties. Moreover, these cells can integrate into host mouse ChP epithelium [134], inviting the exciting possibility of harnessing this approach for intraventricular injections, transplants, and other interventions. In contrast to other body epithelia, mature ChP epithelial cells undergo surprisingly little proliferation or turnover under baseline conditions [135,136,137]. Recent attempts have succeeded in expanding cultured ChP epithelial cells in response to growth factors insulin-like growth factor-1 (IGF-1) and epidermal growth factor (EGF) [138].
Expanding on this foundation, a new methodology created 3-dimensional cultures of human ChP-like organoids [139] by treating human telencephalic organoids [140] with brief pulses of ChP-inducing signaling molecules (e.g., BMPs and WNTs) [134]. These predominantly epithelial cell organoids are well-developed and grow in vitro over extended periods of time, despite lacking vasculature and immune cells. Long-term culture of these organoids (e.g., 68–146 days) results in secretion of CSF-like fluid that approximates native human CSF. Because AAV-mediated approaches can be used for robust, long-term expression and secretion of factors of choice into the CSF [87], the future combination of viral and induced ChP tools can open doors to new therapies for a range of CNS disorders. Cultured ChP cells can also be transfected by polyamine-based transfection reagents to modulate gene expression [20, 141]. Collectively, these tools undoubtedly provide powerful platforms for drug screening. Leveraging these technologies for clinical use will require improved understanding of the survival of transplanted cells in vivo, endurance of viral transduction, and conflicting immune responses that may ensue.
Conclusion
Here, we highlighted the current tools for manipulating the ChP, including transgenic mouse systems, viral vectors, pharmacological and chemogenetic manipulations, and ChP tissue engineering. These various approaches will contribute to understanding the function of ChP and further elucidate the ChP-CSF system. The spatial and temporal resolution offered by genetic approaches can better illuminate the many vital roles of ChP during development. Targeting the expression of critical genes in the ChP through gain- or loss-of-function studies will reveal their contributions to the growth and health of the CNS. In addition, targeting the ChP by specific AAV serotypes offers potential therapeutic strategies for neurodevelopmental and neurological disorders.
Even as current tools broaden our knowledge of in vivo ChP physiology, specificity remains a challenge. Combinatorial genetics and inducible Cre/LoxP systems and AAV vectors hold the most promise for enabling spatial and temporal control of gene manipulation. Inducible or cell-/tissue-specific promoters may also be applied to achieve specificity, and recent advances in optogenetics have welcomed a new era of photo-activatable Cre options [142, 143]. Although the I.C.V. administration route is well established and widely used to deliver viral vectors to the ChP mice of all ages, intranasal and I.V. routes are less invasive and technically easier—intranasal routes can bypass the BBB, and viral vectors that more selectively enter the brain are being developed (e.g., following I.V. delivery) [118, 144].
While we focus primarily on in vivo studies in this review, concerted efforts across many laboratories are aimed at improving in vitro approaches for investigating the ChP that should be amenable to the genetic targeting tools discussed in this review. These in vitro approaches include: primary ChP epithelial cells cultured in dishes and transwells [19, 145,146,147], cell lines such as immortalized mouse ChP epithelial cells (Z310 cells) [148] and porcine ChP epithelial cells (PCP-R) [149] and induced ChP cells [134]. Recently, ChP organoids [139] and ChP explants [38] were developed for investigating ChP functions and pre-clinical treatment strategies.
Recent advances that are beyond the scope of this review and were not covered include peptide- and ligand-mediated targeting of the ChP [150, 151]. Recombinant proenzymes and antibodies can be delivered by I.C.V. infusion in the clinical setting to target the ChP and treat neurologic diseases [150]. Early studies of ligand-mediated approaches also demonstrate that EGF-targeted phages can target the ChP epithelium and transduce genes ex vivo and in vivo [151]. Taken together with the aforementioned techniques, the next level of tools for targeting the ChP will have to be optimized properly; once they are, they will certainly contribute to a better understanding of genetic and molecular mechanisms of the ChP-CSF system and provide important insights to treat CNS disorders.
Availability of data and materials
Not applicable.
Abbreviations
- ChP:
-
Choroid plexus
- CSF:
-
Cerebrospinal fluid
- BBB:
-
Blood–brain barrier
- CNS:
-
Central nervous system
- BCSFB:
-
Blood-cerebrospinal fluid barrier
- AAV:
-
Adeno-associated virus
- I.C.V.:
-
Intracerebroventricular
- I.T.:
-
Intrathecal
- I.V.:
-
Intravenous
- Ttr:
-
Transthyretin
- LV:
-
Lateral ventricle
- 3V:
-
Third ventricle
- 4V:
-
Fourth ventricle
- gRNA:
-
Guide RNA
- CRISPR/Cas9:
-
Clustered regulatory interspaced short palindromic repeats associated RNA-guided Cas9
- LSD:
-
Lysosomal storage disorders
- SMA:
-
Spinal muscular atrophy
References
Solar P, Zamani A, Kubickova L, Dubovy P, Joukal M. Choroid plexus and the blood-cerebrospinal fluid barrier in disease. Fluids Barriers CNS. 2020;17(1):35.
Johansson PA, Dziegielewska KM, Liddelow SA, Saunders NR. The blood-CSF barrier explained: when development is not immaturity. BioEssays. 2008;30(3):237–48.
Ghersi-Egea JF, Strazielle N, Catala M, Silva-Vargas V, Doetsch F, Engelhardt B. Molecular anatomy and functions of the choroidal blood-cerebrospinal fluid barrier in health and disease. Acta Neuropathol. 2018;135(3):337–61.
Rao VV, Dahlheimer JL, Bardgett ME, Snyder AZ, Finch RA, Sartorelli AC, et al. Choroid plexus epithelial expression of MDR1 P glycoprotein and multidrug resistance-associated protein contribute to the blood-cerebrospinal-fluid drug-permeability barrier. Proc Natl Acad Sci U S A. 1999;96(7):3900–5.
Fame RM, Lehtinen MK. Emergence and developmental roles of the cerebrospinal fluid system. Dev Cell. 2020;52(3):261–75.
Lehtinen MK, Zappaterra MW, Chen X, Yang YJ, Hill AD, Lun M, et al. The cerebrospinal fluid provides a proliferative niche for neural progenitor cells. Neuron. 2011;69(5):893–905.
Cserr HF. Physiology of the choroid plexus. Physiol Rev. 1971;51(2):273–311.
Thouvenot E, Lafon-Cazal M, Demettre E, Jouin P, Bockaert J, Marin P. The proteomic analysis of mouse choroid plexus secretome reveals a high protein secretion capacity of choroidal epithelial cells. Proteomics. 2006;6(22):5941–52.
Damkier HH, Brown PD, Praetorius J. Cerebrospinal fluid secretion by the choroid plexus. Physiol Rev. 2013;93(4):1847–92.
Lun MP, Monuki ES, Lehtinen MK. Development and functions of the choroid plexus-cerebrospinal fluid system. Nat Rev Neurosci. 2015;16(8):445–57.
Alvira-Botero X, Carro EM. Clearance of amyloid-beta peptide across the choroid plexus in Alzheimer’s disease. Curr Aging Sci. 2010;3(3):219–29.
Xu H, Fame RM, Sadegh C, Sutin J, Naranjo C, Della S, et al. Choroid plexus NKCC1 mediates cerebrospinal fluid clearance during mouse early postnatal development. Nat Commun. 2021;12(1):447.
Serot JM, Bene MC, Faure GC. Choroid plexus, aging of the brain, and Alzheimer’s disease. Front Biosci. 2003;8:s515–21.
Sousa JC, Cardoso I, Marques F, Saraiva MJ, Palha JA. Transthyretin and Alzheimer’s disease: where in the brain? Neurobiol Aging. 2007;28(5):713–8.
Weller RO. Pathology of cerebrospinal fluid and interstitial fluid of the CNS: significance for Alzheimer disease, prion disorders and multiple sclerosis. J Neuropathol Exp Neurol. 1998;57(10):885–94.
Vercellino M, Votta B, Condello C, Piacentino C, Romagnolo A, Merola A, et al. Involvement of the choroid plexus in multiple sclerosis autoimmune inflammation: a neuropathological study. J Neuroimmunol. 2008;199(1–2):133–41.
Johanson CE, Duncan JA, Stopa EG, Baird A. Enhanced prospects for drug delivery and brain targeting by the choroid plexus-CSF route. Pharm Res. 2005;22(7):1011–37.
Gonzalez AM, Leadbeater WE, Burg M, Sims K, Terasaki T, Johanson CE, et al. Targeting choroid plexus epithelia and ventricular ependyma for drug delivery to the central nervous system. BMC Neurosci. 2011;12:4.
Vong KI, Ma TC, Li B, Leung TCN, Nong W, Ngai SM, et al. SOX9-COL9A3-dependent regulation of choroid plexus epithelial polarity governs blood-cerebrospinal fluid barrier integrity. Proc Natl Acad Sci U S A. 2021. https://doi.org/10.1073/pnas.2009568118.
Boassa D, Stamer WD, Yool AJ. Ion channel function of aquaporin-1 natively expressed in choroid plexus. J Neurosci. 2006;26(30):7811–9.
Carro E, Spuch C, Trejo JL, Antequera D, Torres-Aleman I. Choroid plexus megalin is involved in neuroprotection by serum insulin-like growth factor I. J Neurosci. 2005;25(47):10884–93.
LoTurco J, Manent JB, Sidiqi F. New and improved tools for in utero electroporation studies of developing cerebral cortex. Cereb Cortex. 2009;19(Suppl 1):i120–5.
Dani N, Herbst RH, McCabe C, Green GS, Kaiser K, Head JP, et al. A cellular and spatial map of the choroid plexus across brain ventricles and ages. Cell. 2021;184(11):3056-74 e21.
Lun MP, Johnson MB, Broadbelt KG, Watanabe M, Kang YJ, Chau KF, et al. Spatially heterogeneous choroid plexus transcriptomes encode positional identity and contribute to regional CSF production. J Neurosci. 2015;35(12):4903–16.
Chanoine JP, Braverman LE. The role of transthyretin in the transport of thyroid hormone to cerebrospinal fluid and brain. Acta Med Austriaca. 1992;19(Suppl 1):25–8.
Kwon GS, Hadjantonakis AK. Transthyretin mouse transgenes direct RFP expression or Cre-mediated recombination throughout the visceral endoderm. Genesis. 2009;47(7):447–55.
Johnson BA, Coutts M, Vo HM, Hao X, Fatima N, Rivera MJ, et al. Accurate, strong, and stable reporting of choroid plexus epithelial cells in transgenic mice using a human transthyretin BAC. Fluids Barriers CNS. 2018;15(1):22.
Zhang Y, Huang G, Shornick LP, Roswit WT, Shipley JM, Brody SL, et al. A transgenic FOXJ1-Cre system for gene inactivation in ciliated epithelial cells. Am J Respir Cell Mol Biol. 2007;36(5):515–9.
Spassky N, Meunier A. The development and functions of multiciliated epithelia. Nat Rev Mol Cell Biol. 2017;18(7):423–36.
Delgehyr N, Meunier A, Faucourt M, Bosch Grau M, Strehl L, Janke C, et al. Ependymal cell differentiation, from monociliated to multiciliated cells. Methods Cell Biol. 2015;127:19–35.
Sternberg N, Hamilton D. Bacteriophage P1 site-specific recombination. I. Recombination between loxP sites. J Mol Biol. 1981;150(4):467–86.
Thyagarajan B, Guimaraes MJ, Groth AC, Calos MP. Mammalian genomes contain active recombinase recognition sites. Gene. 2000;244(1–2):47–54.
Semprini S, Troup TJ, Kotelevtseva N, King K, Davis JR, Mullins LJ, et al. Cryptic loxP sites in mammalian genomes: genome-wide distribution and relevance for the efficiency of BAC/PAC recombineering techniques. Nucleic Acids Res. 2007;35(5):1402–10.
Nagy A. Cre recombinase: the universal reagent for genome tailoring. Genesis. 2000;26(2):99–109.
Muzumdar MD, Tasic B, Miyamichi K, Li L, Luo L. A global double-fluorescent Cre reporter mouse. Genesis. 2007;45(9):593–605.
Heiman M, Schaefer A, Gong S, Peterson JD, Day M, Ramsey KE, et al. A translational profiling approach for the molecular characterization of CNS cell types. Cell. 2008;135(4):738–48.
Madisen L, Garner AR, Shimaoka D, Chuong AS, Klapoetke NC, Li L, et al. Transgenic mice for intersectional targeting of neural sensors and effectors with high specificity and performance. Neuron. 2015;85(5):942–58.
Shipley FB, Dani N, Xu H, Deister C, Cui J, Head JP, et al. Tracking calcium dynamics and immune surveillance at the choroid plexus blood-cerebrospinal fluid interface. Neuron. 2020;108(4):623-39 e10.
Korzh V. Development of brain ventricular system. Cell Mol Life Sci. 2018;75(3):375–83.
Kalueff AV, Stewart AM, Gerlai R. Zebrafish as an emerging model for studying complex brain disorders. Trends Pharmacol Sci. 2014;35(2):63–75.
Najib NHM, Nies YH, Abd Halim SAS, Yahaya MF, Das S, Lim WL, et al. Modeling Parkinson’s disease in zebrafish. CNS Neurol Disord Drug Targets. 2020;19(5):386–99.
MacRae CA, Peterson RT. Zebrafish as tools for drug discovery. Nat Rev Drug Discov. 2015;14(10):721–31.
Garcia-Lecea M, Kondrychyn I, Fong SH, Ye ZR, Korzh V. In vivo analysis of choroid plexus morphogenesis in zebrafish. PLoS ONE. 2008;3(9): e3090.
Bill BR, Balciunas D, McCarra JA, Young ED, Xiong T, Spahn AM, et al. Development and Notch signaling requirements of the zebrafish choroid plexus. PLoS ONE. 2008;3(9): e3114.
Bill BR, Korzh V. Choroid plexus in developmental and evolutionary perspective. Front Neurosci. 2014;8:363.
Henson HE, Parupalli C, Ju B, Taylor MR. Functional and genetic analysis of choroid plexus development in zebrafish. Front Neurosci. 2014;8:364.
van Leeuwen LM, Evans RJ, Jim KK, Verboom T, Fang X, Bojarczuk A, et al. A transgenic zebrafish model for the in vivo study of the blood and choroid plexus brain barriers using claudin 5. Biol Open. 2018. https://doi.org/10.1242/bio.030494.
Parab S, Quick RE, Matsuoka RL. Endothelial cell-type-specific molecular requirements for angiogenesis drive fenestrated vessel development in the brain. Elife. 2021. https://doi.org/10.7554/eLife.64295.
Myung J, Schmal C, Hong S, Tsukizawa Y, Rose P, Zhang Y, et al. The choroid plexus is an important circadian clock component. Nat Commun. 2018;9(1):1062.
Huang X, Ketova T, Fleming JT, Wang H, Dey SK, Litingtung Y, et al. Sonic hedgehog signaling regulates a novel epithelial progenitor domain of the hindbrain choroid plexus. Development. 2009;136(15):2535–43.
Lewis PM, Dunn MP, McMahon JA, Logan M, Martin JF, St-Jacques B, et al. Cholesterol modification of sonic hedgehog is required for long-range signaling activity and effective modulation of signaling by Ptc1. Cell. 2001;105(5):599–612.
Lewis AE, Vasudevan HN, O’Neill AK, Soriano P, Bush JO. The widely used Wnt1-Cre transgene causes developmental phenotypes by ectopic activation of Wnt signaling. Dev Biol. 2013;379(2):229–34.
Kaiser K, Jang A, Kompanikova P, Lun MP, Prochazka J, Machon O, et al. MEIS-WNT5A axis regulates development of fourth ventricle choroid plexus. Development. 2021. https://doi.org/10.1242/dev.192054.
Crouthamel MH, Kelly EJ, Ho RJ. Development and characterization of transgenic mouse models for conditional gene knockout in the blood-brain and blood-CSF barriers. Transgenic Res. 2012;21(1):113–30.
Safaee M, Oh MC, Bloch O, Sun MZ, Kaur G, Auguste KI, et al. Choroid plexus papillomas: advances in molecular biology and understanding of tumorigenesis. Neuro Oncol. 2013;15(3):255–67.
Shannon ML, Fame RM, Chau KF, Dani N, Calicchio ML, Geleoc GS, et al. Mice expressing Myc in neural precursors develop choroid plexus and ciliary body tumors. Am J Pathol. 2018;188(6):1334–44.
Wang J, Merino DM, Light N, Murphy BL, Wang YD, Guo X, et al. Myc and loss of p53 cooperate to drive formation of choroid plexus carcinoma. Cancer Res. 2019;79(9):2208–19.
Dang L, Fan X, Chaudhry A, Wang M, Gaiano N, Eberhart CG. Notch3 signaling initiates choroid plexus tumor formation. Oncogene. 2006;25(3):487–91.
Awatramani R, Soriano P, Rodriguez C, Mai JJ, Dymecki SM. Cryptic boundaries in roof plate and choroid plexus identified by intersectional gene activation. Nat Genet. 2003;35(1):70–5.
Liang H, Hippenmeyer S, Ghashghaei HT. A Nestin-cre transgenic mouse is insufficient for recombination in early embryonic neural progenitors. Biol Open. 2012;1(12):1200–3.
Tronche F, Kellendonk C, Kretz O, Gass P, Anlag K, Orban PC, et al. Disruption of the glucocorticoid receptor gene in the nervous system results in reduced anxiety. Nat Genet. 1999;23(1):99–103.
Calado DP, Sasaki Y, Godinho SA, Pellerin A, Kochert K, Sleckman BP, et al. The cell-cycle regulator c-Myc is essential for the formation and maintenance of germinal centers. Nat Immunol. 2012;13(11):1092–100.
Lakso M, Sauer B, Mosinger B Jr, Lee EJ, Manning RW, Yu SH, et al. Targeted oncogene activation by site-specific recombination in transgenic mice. Proc Natl Acad Sci U S A. 1992;89(14):6232–6.
Indra AK, Warot X, Brocard J, Bornert JM, Xiao JH, Chambon P, et al. Temporally-controlled site-specific mutagenesis in the basal layer of the epidermis: comparison of the recombinase activity of the tamoxifen-inducible Cre-ER(T) and Cre-ER(T2) recombinases. Nucleic Acids Res. 1999;27(22):4324–7.
Sauer B. Inducible gene targeting in mice using the Cre/lox system. Methods. 1998;14(4):381–92.
Muthusamy N, Vijayakumar A, Cheng G Jr, Ghashghaei HT. A knock-in Foxj1(CreERT2::GFP) mouse for recombination in epithelial cells with motile cilia. Genesis. 2014;52(4):350–8.
Kaiser K, Gyllborg D, Prochazka J, Salasova A, Kompanikova P, Molina FL, et al. WNT5A is transported via lipoprotein particles in the cerebrospinal fluid to regulate hindbrain morphogenesis. Nat Commun. 2019;10(1):1498.
Johansson PA, Irmler M, Acampora D, Beckers J, Simeone A, Gotz M. The transcription factor Otx2 regulates choroid plexus development and function. Development. 2013;140(5):1055–66.
Puelles E, Acampora D, Lacroix E, Signore M, Annino A, Tuorto F, et al. Otx dose-dependent integrated control of antero-posterior and dorso-ventral patterning of midbrain. Nat Neurosci. 2003;6(5):453–60.
Peitz M, Pfannkuche K, Rajewsky K, Edenhofer F. Ability of the hydrophobic FGF and basic TAT peptides to promote cellular uptake of recombinant Cre recombinase: a tool for efficient genetic engineering of mammalian genomes. Proc Natl Acad Sci U S A. 2002;99(7):4489–94.
Spatazza J, Lee HH, Di Nardo AA, Tibaldi L, Joliot A, Hensch TK, et al. Choroid-plexus-derived Otx2 homeoprotein constrains adult cortical plasticity. Cell Rep. 2013;3(6):1815–23.
Planques A, Oliveira Moreira V, Dubreuil C, Prochiantz A, Di Nardo AA. OTX2 signals from the choroid plexus to regulate adult neurogenesis. eNeuro. 2019. https://doi.org/10.1523/ENEURO.0262-18.2019.
Planques A, Oliveira Moreira V, Benacom D, Bernard C, Jourdren L, Blugeon C, et al. OTX2 homeoprotein functions in adult choroid plexus. Int J Mol Sci. 2021;22(16):8951.
Arnaud K, Oliveira Moreira V, Vincent J, Dallerac G, Dubreuil C, Dupont E, et al. Choroid plexus APP regulates adult brain proliferation and animal behavior. Life Sci Alliance. 2021;4(11):e202000703.
Kinoshita A, Shqirat M, Kageyama R, Ohtsuka T. Modification of gene expression and soluble factor secretion in the lateral ventricle choroid plexus: Analysis of the impacts on the neocortical development. Neurosci Res. 2021;177:38–51.
Das AT, Tenenbaum L, Berkhout B. Tet-on systems for doxycycline-inducible gene expression. Curr Gene Ther. 2016;16(3):156–67.
Shqirat M, Kinoshita A, Kageyama R, Ohtsuka T. Sonic hedgehog expands neural stem cells in the neocortical region leading to an expanded and wrinkled neocortical surface. Genes Cells. 2021;26(6):399–410.
Currle DS, Cheng X, Hsu CM, Monuki ES. Direct and indirect roles of CNS dorsal midline cells in choroid plexus epithelia formation. Development. 2005;132(15):3549–59.
Grove EA, Tole S, Limon J, Yip L, Ragsdale CW. The hem of the embryonic cerebral cortex is defined by the expression of multiple Wnt genes and is compromised in Gli3-deficient mice. Development. 1998;125(12):2315–25.
Wang D, Zhang F, Gao G. CRISPR-based therapeutic genome editing: strategies and in vivo delivery by AAV vectors. Cell. 2020;181(1):136–50.
Karimian A, Gorjizadeh N, Alemi F, Asemi Z, Azizian K, Soleimanpour J, et al. CRISPR/Cas9 novel therapeutic road for the treatment of neurodegenerative diseases. Life Sci. 2020;259: 118165.
Quadros RM, Miura H, Harms DW, Akatsuka H, Sato T, Aida T, et al. Easi-CRISPR: a robust method for one-step generation of mice carrying conditional and insertion alleles using long ssDNA donors and CRISPR ribonucleoproteins. Genome Biol. 2017;18(1):92.
Doudna JA. The promise and challenge of therapeutic genome editing. Nature. 2020;578(7794):229–36.
Maynard LH, Humbert O, Peterson CW, Kiem HP. Genome editing in large animal models. Mol Ther. 2021;29(11):3140–52.
Passini MA, Watson DJ, Vite CH, Landsburg DJ, Feigenbaum AL, Wolfe JH. Intraventricular brain injection of adeno-associated virus type 1 (AAV1) in neonatal mice results in complementary patterns of neuronal transduction to AAV2 and total long-term correction of storage lesions in the brains of beta-glucuronidase-deficient mice. J Virol. 2003;77(12):7034–40.
Passini MA, Wolfe JH. Widespread gene delivery and structure-specific patterns of expression in the brain after intraventricular injections of neonatal mice with an adeno-associated virus vector. J Virol. 2001;75(24):12382–92.
Cui J, Shipley FB, Shannon ML, Alturkistani O, Dani N, Webb MD, et al. Inflammation of the embryonic choroid plexus barrier following maternal immune activation. Dev Cell. 2020;55(5):617–28.
Haddad MR, Donsante A, Zerfas P, Kaler SG. Fetal brain-directed AAV gene therapy results in rapid, robust, and persistent transduction of mouse choroid plexus epithelia. Mol Ther Nucleic Acids. 2013;2: e101.
Chen X, He Y, Tian Y, Wang Y, Wu Z, Lan T, et al. Different serotypes of adeno-associated virus vector- and lentivirus-mediated tropism in choroid plexus by intracerebroventricular delivery. Hum Gene Ther. 2020;31(7–8):440–7.
Donsante A, Yi L, Zerfas PM, Brinster LR, Sullivan P, Goldstein DS, et al. ATP7A gene addition to the choroid plexus results in long-term rescue of the lethal copper transport defect in a Menkes disease mouse model. Mol Ther. 2011;19(12):2114–23.
Steffensen AB, Oernbo EK, Stoica A, Gerkau NJ, Barbuskaite D, Tritsaris K, et al. Cotransporter-mediated water transport underlying cerebrospinal fluid formation. Nat Commun. 2018;9(1):2167.
Watson DJ, Passini MA, Wolfe JH. Transduction of the choroid plexus and ependyma in neonatal mouse brain by vesicular stomatitis virus glycoprotein-pseudotyped lentivirus and adeno-associated virus type 5 vectors. Hum Gene Ther. 2005;16(1):49–56.
Schuster DJ, Belur LR, Riedl MS, Schnell SA, Podetz-Pedersen KM, Kitto KF, et al. Supraspinal gene transfer by intrathecal adeno-associated virus serotype 5. Front Neuroanat. 2014;8:66.
Zhu L, Stein LR, Kim D, Ho K, Yu GQ, Zhan L, et al. Klotho controls the brain-immune system interface in the choroid plexus. Proc Natl Acad Sci U S A. 2018;115(48):E11388–96.
Lepko T, Pusch M, Muller T, Schulte D, Ehses J, Kiebler M, et al. Choroid plexus-derived miR-204 regulates the number of quiescent neural stem cells in the adult brain. EMBO J. 2019;38(17): e100481.
Hu S, Yang T, Wang Y. Widespread labeling and genomic editing of the fetal central nervous system by in utero CRISPR AAV9-PHP.eB administration. Development. 2021. https://doi.org/10.1242/dev.195586.
Hudry E, Vandenberghe LH. Therapeutic AAV gene transfer to the nervous system: a clinical reality. Neuron. 2019;102(1):263.
Deverman BE, Pravdo PL, Simpson BP, Kumar SR, Chan KY, Banerjee A, et al. Cre-dependent selection yields AAV variants for widespread gene transfer to the adult brain. Nat Biotechnol. 2016;34(2):204–9.
Goertsen D, Flytzanis NC, Goeden N, Chuapoco MR, Cummins A, Chen Y, et al. AAV capsid variants with brain-wide transgene expression and decreased liver targeting after intravenous delivery in mouse and marmoset. Nat Neurosci. 2022;25(1):106–15.
Ravindra Kumar S, Miles TF, Chen X, Brown D, Dobreva T, Huang Q, et al. Multiplexed Cre-dependent selection yields systemic AAVs for targeting distinct brain cell types. Nat Methods. 2020;17(5):541–50.
Huang LY, Halder S, Agbandje-McKenna M. Parvovirus glycan interactions. Curr Opin Virol. 2014;7:108–18.
Huang Q, Chan KY, Tobey IG, Chan YA, Poterba T, Boutros CL, et al. Delivering genes across the blood-brain barrier: LY6A, a novel cellular receptor for AAV-PHP.B capsids. PLoS ONE. 2019;14(11):e0225206.
Massaro G, Geard AF, Liu W, Coombe-Tennant O, Waddington SN, Baruteau J, et al. Gene therapy for lysosomal storage disorders: ongoing studies and clinical development. Biomolecules. 2021;11(4):611.
Mendell JR, Al-Zaidy SA, Rodino-Klapac LR, Goodspeed K, Gray SJ, Kay CN, et al. Current clinical applications of in vivo gene therapy with AAVs. Mol Ther. 2021;29(2):464–88.
Al-Zaidy SA, Mendell JR. From clinical trials to clinical practice: practical considerations for gene replacement therapy in SMA type 1. Pediatr Neurol. 2019;100:3–11.
Chen TH. New and developing therapies in spinal muscular atrophy: from genotype to phenotype to treatment and where do we stand? Int J Mol Sci. 2020;21(9):3297.
Chan YK, Wang SK, Chu CJ, Copland DA, Letizia AJ, Costa Verdera H, et al. Engineering adeno-associated viral vectors to evade innate immune and inflammatory responses. Sci Transl Med. 2021. https://doi.org/10.1126/scitranslmed.abd3438.
Johnston S, Parylak SL, Kim S, Mac N, Lim C, Gallina I, et al. AAV ablates neurogenesis in the adult murine hippocampus. Elife. 2021. https://doi.org/10.7554/eLife.59291.
Nishida F, Morel GR, Herenu CB, Schwerdt JI, Goya RG, Portiansky EL. Restorative effect of intracerebroventricular insulin-like growth factor-I gene therapy on motor performance in aging rats. Neuroscience. 2011;177:195–206.
Dindot S, Piccolo P, Grove N, Palmer D, Brunetti-Pierri N. Intrathecal injection of helper-dependent adenoviral vectors results in long-term transgene expression in neuroependymal cells and neurons. Hum Gene Ther. 2011;22(6):745–51.
Zhou HJ, Li H, Shi MQ, Mao XN, Liu DL, Chang YR, et al. Protective effect of klotho against ischemic brain injury is associated with inhibition of RIG-I/NF-kappaB signaling. Front Pharmacol. 2017;8:950.
Zeng CY, Yang TT, Zhou HJ, Zhao Y, Kuang X, Duan W, et al. Lentiviral vector-mediated overexpression of Klotho in the brain improves Alzheimer’s disease-like pathology and cognitive deficits in mice. Neurobiol Aging. 2019;78:18–28.
Zhao Y, Zeng CY, Li XH, Yang TT, Kuang X, Du JR. Klotho overexpression improves amyloid-beta clearance and cognition in the APP/PS1 mouse model of Alzheimer’s disease. Aging Cell. 2020;19(10):e13239.
Regev L, Ezrielev E, Gershon E, Gil S, Chen A. Genetic approach for intracerebroventricular delivery. Proc Natl Acad Sci U S A. 2010;107(9):4424–9.
Kriegstein A, Alvarez-Buylla A. The glial nature of embryonic and adult neural stem cells. Annu Rev Neurosci. 2009;32:149–84.
Liu G, Martins IH, Chiorini JA, Davidson BL. Adeno-associated virus type 4 (AAV4) targets ependyma and astrocytes in the subventricular zone and RMS. Gene Ther. 2005;12(20):1503–8.
Lochhead JJ, Thorne RG. Intranasal delivery of biologics to the central nervous system. Adv Drug Deliv Rev. 2012;64(7):614–28.
Saraiva J, Nobre RJ, Pereira de Almeida L. Gene therapy for the CNS using AAVs: the impact of systemic delivery by AAV9. J Control Release. 2016;241:94–109.
Belur LR, Temme A, Podetz-Pedersen KM, Riedl M, Vulchanova L, Robinson N, et al. Intranasal adeno-associated virus mediated gene delivery and expression of human iduronidase in the central nervous system: a noninvasive and effective approach for prevention of neurologic disease in mucopolysaccharidosis type I. Hum Gene Ther. 2017;28(7):576–87.
Wang D, Tai PWL, Gao G. Adeno-associated virus vector as a platform for gene therapy delivery. Nat Rev Drug Discov. 2019;18(5):358–78.
Moore CI. A systems approach to the choroid plexus. 2019 Keystone symposia.
Moore CI, Berglund K. BL-OG: BioLuminescent-OptoGenetics. J Neurosci Res. 2020;98(3):469–70.
Gomez-Ramirez M, More AI, Friedman NG, Hochgeschwender U, Moore CI. The BioLuminescent-OptoGenetic in vivo response to coelenterazine is proportional, sensitive, and specific in neocortex. J Neurosci Res. 2020;98(3):471–80.
Berglund K, Birkner E, Augustine GJ, Hochgeschwender U. Light-emitting channelrhodopsins for combined optogenetic and chemical-genetic control of neurons. PLoS ONE. 2013;8(3): e59759.
Berglund K, Clissold K, Li HE, Wen L, Park SY, Gleixner J, et al. Luminopsins integrate opto- and chemogenetics by using physical and biological light sources for opsin activation. Proc Natl Acad Sci U S A. 2016;113(3):E358–67.
Berglund K, Tung JK, Higashikubo B, Gross RE, Moore CI, Hochgeschwender U. Combined optogenetic and chemogenetic control of neurons. Methods Mol Biol. 2016;1408:207–25.
Medendorp WE, Bjorefeldt A, Crespo EL, Prakash M, Pal A, Waddell ML, et al. Selective postnatal excitation of neocortical pyramidal neurons results in distinctive behavioral and circuit deficits in adulthood. iScience. 2021;24(3):102157.
Kim CK, Adhikari A, Deisseroth K. Integration of optogenetics with complementary methodologies in systems neuroscience. Nat Rev Neurosci. 2017;18(4):222–35.
Thanos CG, Bintz B, Emerich DF. Microencapsulated choroid plexus epithelial cell transplants for repair of the brain. Adv Exp Med Biol. 2010;670:80–91.
Skinner SJ, Geaney MS, Lin H, Muzina M, Anal AK, Elliott RB, et al. Encapsulated living choroid plexus cells: potential long-term treatments for central nervous system disease and trauma. J Neural Eng. 2009;6(6): 065001.
Matsumoto N, Taguchi A, Kitayama H, Watanabe Y, Ohta M, Yoshihara T, et al. Transplantation of cultured choroid plexus epithelial cells via cerebrospinal fluid shows prominent neuroprotective effects against acute ischemic brain injury in the rat. Neurosci Lett. 2010;469(3):283–8.
Borlongan CV, Thanos CG, Skinner SJ, Geaney M, Emerich DF. Transplants of encapsulated rat choroid plexus cells exert neuroprotection in a rodent model of Huntington’s disease. Cell Transplant. 2008;16(10):987–92.
Emerich DF, Thanos CG, Goddard M, Skinner SJ, Geany MS, Bell WJ, et al. Extensive neuroprotection by choroid plexus transplants in excitotoxin lesioned monkeys. Neurobiol Dis. 2006;23(2):471–80.
Watanabe M, Kang YJ, Davies LM, Meghpara S, Lau K, Chung CY, et al. BMP4 sufficiency to induce choroid plexus epithelial fate from embryonic stem cell-derived neuroepithelial progenitors. J Neurosci. 2012;32(45):15934–45.
Chauhan AN, Lewis PD. A quantitative study of cell proliferation in ependyma and choroid plexus in the postnatal rat brain. Neuropathol Appl Neurobiol. 1979;5(4):303–9.
Kaplan MS. Proliferation of epithelial cells in the adult primate choroid plexus. Anat Rec. 1980;197(4):495–502.
Korzhevskii DE. Proliferative zones in the epithelium of the choroid plexuses of the human embryo brain. Neurosci Behav Physiol. 2000;30(5):509–12.
Barkho BZ, Monuki ES. Proliferation of cultured mouse choroid plexus epithelial cells. PLoS ONE. 2015;10(3): e0121738.
Pellegrini L, Bonfio C, Chadwick J, Begum F, Skehel M, Lancaster MA. Human CNS barrier-forming organoids with cerebrospinal fluid production. Science. 2020. https://doi.org/10.1126/science.aaz5626.
Lancaster MA, Renner M, Martin CA, Wenzel D, Bicknell LS, Hurles ME, et al. Cerebral organoids model human brain development and microcephaly. Nature. 2013;501(7467):373–9.
Dana H, Chalbatani GM, Mahmoodzadeh H, Karimloo R, Rezaiean O, Moradzadeh A, et al. Molecular mechanisms and biological functions of siRNA. Int J Biomed Sci. 2017;13(2):48–57.
Morikawa K, Furuhashi K, de Sena-Tomas C, Garcia-Garcia AL, Bekdash R, Klein AD, et al. Photoactivatable Cre recombinase 3.0 for in vivo mouse applications. Nat Commun. 2020;11(1):2141.
Kawano F, Okazaki R, Yazawa M, Sato M. A photoactivatable Cre-loxP recombination system for optogenetic genome engineering. Nat Chem Biol. 2016;12(12):1059–64.
Erdo F, Bors LA, Farkas D, Bajza A, Gizurarson S. Evaluation of intranasal delivery route of drug administration for brain targeting. Brain Res Bull. 2018;143:155–70.
Monnot AD, Zheng W. Culture of choroid plexus epithelial cells and in vitro model of blood-CSF barrier. Methods Mol Biol. 2013;945:13–29.
Lallai V, Ahmed A, Fowler CD. Method for primary epithelial cell culture from the rat choroid plexus. Bio Protoc. 2020;10(4): e3532.
Strazielle N, Ghersi-Egea JF. Demonstration of a coupled metabolism-efflux process at the choroid plexus as a mechanism of brain protection toward xenobiotics. J Neurosci. 1999;19(15):6275–89.
Zheng W, Zhao Q. Establishment and characterization of an immortalized Z310 choroidal epithelial cell line from murine choroid plexus. Brain Res. 2002;958(2):371–80.
Schroten M, Hanisch FG, Quednau N, Stump C, Riebe R, Lenk M, et al. A novel porcine in vitro model of the blood-cerebrospinal fluid barrier with strong barrier function. PLoS ONE. 2012;7(6): e39835.
Bryniarski MA, Ren T, Rizvi AR, Snyder AM, Morris ME. Targeting the choroid plexuses for protein drug delivery. Pharmaceutics. 2020;12(10):963.
Gonzalez AM, Leadbeater W, Podvin S, Borboa A, Burg M, Sawada R, et al. Epidermal growth factor targeting of bacteriophage to the choroid plexus for gene delivery to the central nervous system via cerebrospinal fluid. Brain Res. 2010;1359:1–13.
Karolewski BA, Wolfe JH. Genetic correction of the fetal brain increases the lifespan of mice with the severe multisystemic disease mucopolysaccharidosis type VII. Mol Ther. 2006;14(1):14–24.
Broekman ML, Comer LA, Hyman BT, Sena-Esteves M. Adeno-associated virus vectors serotyped with AAV8 capsid are more efficient than AAV-1 or -2 serotypes for widespread gene delivery to the neonatal mouse brain. Neuroscience. 2006;138(2):501–10.
Yamazaki Y, Hirai Y, Miyake K, Shimada T. Targeted gene transfer into ependymal cells through intraventricular injection of AAV1 vector and long-term enzyme replacement via the CSF. Sci Rep. 2014;4:5506.
Liu G, Martins I, Wemmie JA, Chiorini JA, Davidson BL. Functional correction of CNS phenotypes in a lysosomal storage disease model using adeno-associated virus type 4 vectors. J Neurosci. 2005;25(41):9321–7.
Dodge JC, Treleaven CM, Fidler JA, Hester M, Haidet A, Handy C, et al. AAV4-mediated expression of IGF-1 and VEGF within cellular components of the ventricular system improves survival outcome in familial ALS mice. Mol Ther. 2010;18(12):2075–84.
Acknowledgements
We thank N. Chamberlin and members of the Lehtinen lab for helpful discussions and reading of this manuscript. Schematics in Figure 2 created with BioRender.com. Maria K. Lehtinen is a New York Stem Cell Foundation – Robertson Investigator.
Funding
This work was supported by NIH R01 NS088566 and the New York Stem Cell Foundation (M.K.L.).
Author information
Authors and Affiliations
Contributions
AJ and MKL wrote the manuscript. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publications
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Jang, A., Lehtinen, M.K. Experimental approaches for manipulating choroid plexus epithelial cells. Fluids Barriers CNS 19, 36 (2022). https://doi.org/10.1186/s12987-022-00330-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12987-022-00330-2