
Abstract
Background
Nonnucleoside reverse transcriptase inhibitors (NNRTIs) are a class of antiretroviral compounds that bind in an allosteric binding pocket in HIV-1 RT, located about 10 Å from the polymerase active site. Binding of an NNRTI causes structural changes that perturb the alignment of the primer terminus and polymerase active site, preventing viral DNA synthesis. Rilpivirine (RPV) is the most recent NNRTI approved by the FDA, but like all other HIV-1 drugs, suboptimal treatment can lead to the development of resistance. To generate better compounds that could be added to the current HIV-1 drug armamentarium, we have developed several RPV analogs to combat viral variants that are resistant to the available NNRTIs.
Results
Using a single-round infection assay, we identified several RPV analogs that potently inhibited a broad panel of NNRTI resistant mutants. Additionally, we determined that several resistant mutants selected by either RPV or Doravirine (DOR) caused only a small increase in susceptibility to the most promising RPV analogs.
Conclusions
The antiviral data suggested that there are RPV analogs that could be candidates for further development as NNRTIs, and one of the most promising compounds was modeled in the NNRTI binding pocket. This model can be used to explain why this compound is broadly effective against the panel of NNRTI resistance mutants.
Similar content being viewed by others


Background
HIV-1 reverse transcriptase (RT) is a target for many drugs used in highly active antiretroviral therapy (HAART) to treat HIV-1 infections [1]. RT has two enzymatic activities: (1) a DNA polymerase that can copy either an RNA or a DNA template and (2) an RNase H that degrades RNA if and only if the RNA is part of a RNA: DNA hybrid. Although there are, as yet, no drugs that target the RNase H of HIV-1 RT, there are two classes of drugs that target the DNA polymerase. The first class consists of inhibitors that are analogs of normal nucleosides used to synthesize DNA (nucleoside reverse transcriptase inhibitors, NRTIs). All of the FDA-approved NRTIs lack the 3′ –OH present on the deoxyribose of normal nucleosides. When an NRTI is incorporated into the growing viral DNA strand, it acts as a chain terminator. The second class consists of inhibitors that bind in a small hydrophobic pocket ~10 Å from the polymerase active site (non-nucleoside reverse transcriptase inhibitors, NNRTIs) [2–4]. The NNRTI binding pocket is closed in the absence of a bound NNRTI [5]. The NNRTI binding pocket is formed by the following residues: L100, K101, K103, V106, V179, Y181, Y188, G190, F227, W229, L234, P236, and Y318. The binding pocket lies underneath the bound double-stranded nucleic acid substrate. Binding of an NNRTI distorts RT, which affects the alignment of the primer terminus and the polymerase active site, blocking the chemical step of viral DNA synthesis [6–8]. There currently are sixteen drugs that target RT that have been approved by the FDA for the treatment of HIV-1 infections; eleven are NRTIs and five are NNRTIs. The five approved NNRTI drugs are: nevirapine (NVP, Viramune), delavirdine (DLV, Rescriptor), efavirenz (EFV, Sustiva), etravirine (ETR, Intelence), and rilpivirine (RPV, Endurant,). While NNRTIs effectively block the replication of WT HIV-1, NNRTI-resistant mutants can emerge during treatment, many of which cause cross-resistance among the approved NNRTIs.
There are two primary factors that contribute to the emergence of resistance to NNRTIs: (1) HIV-1 RT can tolerate a wide range of sequences in and around the NNRTI binding pocket and (2) there is extensive HIV-1 genetic variation [9, 10]. Although most HIV-infected individuals are initially infected with a single virion, HIV-1 variants arise rapidly due to high viral loads in HIV-1 infected patients, which leads to the infection of large numbers of cells, the rapid turnover of these infected cells, and to errors made during HIV-1 replication [10–12]. Ultimately, error prone replication creates the mutations that enable HIV-1 to develop resistance against antiretroviral drugs.
A number of drug resistant mutants were selected in HIV-infected individuals by the first generation NNRTIs (NVP, EFV, and DLV). Resistance mutations are commonly seen in the residues that surround the NNRTI binding pocket, including: L100I, K103N, V106A, Y181C, Y188L, and H221Y. These mutations alter the geometry of the NNRTI binding pocket in ways that interfere with the binding of NNRTIs. The first generation inhibitors were rigid and bulky compounds that were particularly vulnerable to the effects of resistance mutations. The second generation NNRTIs, ETR and RPV, were designed to be less bulky and more flexible, and are better able to adapt to the changes in the NNRTI binding pocket caused by resistance mutations. This allows these newer NNRTIs to effectively inhibit both WT HIV-1 and a number of drug resistant variants [13, 14]. However, when these drugs were tested in clinical trials, patients who were on an ETR containing regimen were found to have a combination of mutations including: V90I, A98G, L100I, K101E/P, V106I, V179D/F, Y181C/I/V, and G190A/S [15]. A variety of mutations including K101E/P, E138A/G/K/Q/R, Y181C/I/V, and M230L have been shown to be associated with a decrease in susceptibility to RPV and have been detected in clinical samples from HIV-1-infected individuals treated with RPV [16]. Individuals who failed in a clinical trial involving therapy with RPV and NRTIs had mutations at positions E138K and M184I/V; the M184V mutation was likely selected by FTC, an NRTI often included in HAART regimens [17, 18].
Because development of resistance can occur with all of the available NNRTIs, there are ongoing efforts to develop NNRTIs that extend and improve the effectiveness of the previously identified compounds. For example, doravirine (DOR) is a new NNRTI in late stage clinical trials [19]. Although it selects for resistance, the mutations that reduce the potency of DOR do not, for the most part, overlap with mutations that reduce the potency of RPV [20]. As an alternative approach, we are testing whether RPV analogs have better resistance profiles than the parent compound.
Several of our RPV analogs are broadly effective against HIV-1 mutants. One compound (11) was particularly effective in inhibiting the replication of mutants that have reduced susceptibility to RPV and DOR, suggesting that it has the potential to become a clinically useful NNRTI.
Results and discussion
Susceptibility of mutant HIV-1 to RPV analogs
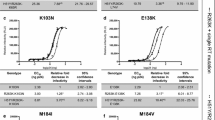
We previously determined the ability of several of our RPV analogs to inhibit infection of WT HIV-1 or mutants that contain several well-known NNRTI-resistant mutations (L100I, K103N, E138K, Y181C, Y188L, H221Y, and K103N/Y181C) using a single-round infection luciferase assay [21]. As shown in Figs. 3 and 4 of that aforementioned study, RPV and several analogs (6, 7, 8, 9, 11, 12, 13, 14, 15, 16, 17, 21, and 27) had potent antiviral activities against both WT HIV-1 and several of the resistant mutants L100I, K103N, Y181C, and H221Y (all IC50s < 7 nM). The E138K mutant caused only a small drop in the IC50s for most of the compounds (<5 nM); however there was a greater loss in susceptibility to 12 (15 nM ± 1.2). The resistant mutant Y188L showed modest drop in susceptibility to 8, 9, 11, 12, 14, 15, 16, and 17 (IC50s ranged from 12 to 64 nM), whereas compounds 6, 7, 13, 21, and 27 showed a modest loss of potency against this resistant mutant (all <10 nM). The NNRTI resistant double mutant K103N/Y181C caused a substantial decrease in susceptibility to RPV analogs 6, 8, 9, 13, 21, and 27 (ranged from 25 to 100 nM), whereas this mutant caused a minor loss in susceptibility to RPV, 7, 11, and 13 (<5 nM).
Because several of our RPV analogs were active against the first set of NNRTI resistant mutants, we tested the antiviral activities of RPV and the most promising analogs 6, 7, 8, 9, 11, 12, 13, 14, 15, 16, 17, 21, 26, and 27 against a much broader panel of resistant mutants. These compounds are structural homologs of RPV (Fig. 1) that have the following differences: the addition of an exocyclic moiety at position 5 (6, 7) or the central pyrimidine moiety is in a “flipped” conformation relative to RPV (8,9), a substitution of the central pyrimidine ring with a 2,6-purine ring system (11) or this same 2,6-purine ring system in the “flipped” conformation with or without a protecting group (12, 13) or this same 2,6-purine ring system with additional modifications on the benzyl moiety (14, 15, 16, 17, 21), or a substitution of central pyrimidine ring with a 2,9-purine ring system (27) or this ring system in a “flipped” orientation (26).

Chemical Structures of the RPV Analogs. Chemical structures of RPV and the RPV analogs. The parts of the analogs that differ from RPV are indicated in red
To determine the ability of these RPV analogs to inhibit a wider range of NNRTI mutants, we tested their antiviral activity in a single-round infectivity assay against: G190A, G190S, M230L, P236L, L100I/K103N, K103N/P225H, and V106A/G190A/F227L (Fig. 2; Additional file 1: Table S1). Compound 26, which has a central 2,9-purine ring system in the “flipped” orientation, had poor activities in the initial study and was included with the more promising compounds to see if there was obvious complementarity in terms of which mutants the compounds would inhibit. RPV and the analogs 6, 7, 8, 9, 11, 12, 13, 14, 15, 16, 17, 21, 26, and 27 all potently inhibited the replication of WT HIV-1. The IC50 values in the single round assay were, for the most part, low nanomolar against the WT (all <2.2 nM; except for 26, which was 7.7 nM ± 2.3), and against several of the RT mutants: G190A (all <1.6 nM; except for 26, which was 2.2 nM ± 0.71), G190S (all <1 nM; except for 26, which was 2.1 nM ± 0.2), P236L RT (all <1.7 nM; except for 26, which was 2.91 nM ± 0.007), and K103N/P225H (all <4.6 nM; except for 26, which was 12.9 nM ± 3.3). In the crystal structure of HIV-1 RT bound to RPV, position 190 is located at the back of the NNRTI binding pocket and is >6 Å away from RPV. When we modeled the binding of the RPV analogs in the NNRTI binding pocket, the distances of the compounds to position 190 were similar to RPV (Figs. 6, 7), which explains why mutations at 190 do not interfere with the binding of this family of inhibitors. The models show that the RPV analogs are 3–4 Å from amino acids 225 and 236, which reside in the depths of the NNRTI binding pocket (Figs. 6, 7). HIV-1 containing the M230L mutation was susceptible to RPV and to the analogs 6, 7, 11, 13, 14, 16, 21, and 27 (all IC50s < 6.1 nM), but showed a modest decrease in susceptibility to analogs 8 (11.9 nM ± 1.1), 9 (29.6 nM ± 1.1), 12 (13.8 nM ± 7.1), 15 (16.8 nM ± 1.4), and 17 (21.1 nM ± 8.8), while compound 26 was not effective in our assay (IC50 > 100 nM, above the limit of detection in our assay). The antiviral results suggest that most of the RPV analogs with a central purine scaffold and a benzonitrile moiety identical to the one on RPV, or a similar moiety that lacks the fluoride modification, can successfully inhibit viruses with the M230L mutation in RT. RPV analogs with modifications at position 5 on the pyrimidine scaffold, with the scaffold in the same orientation as RPV, are also capable of inhibiting the M230L variant. The NNRTI resistant double mutant L100I/K103N posed a problem for some of the RPV analogs. Both compounds 14 (12.6 nM ± 1.6) and 15 (15.5 nM ± .6) showed minor reductions in sensitivity to L100I/K103N, while compounds 9 (44.1 nM ± 17.6), 17 (63.4 nM ± 12.6), 21 (37.6 nM ± 0.14), and 27 (39.8 nM ± 9.2) had a significant drop in sensitivity and 26 was beyond the limits of detection using our assay (>100 nM). However, RPV and analogs 6, 7, 8, 11, 12, 13, and 16, showed strong antiviral activities (all <5.4 nM) against L100I/K103N. These data illustrate the importance of preserving the benzonitrile group as one of the arylamines. The problems associated with the K103N mutation might restrict the range of functional groups on the inhibitors that can be used at this position [22].

Antiviral activities of RPV and the RPV analogs against well-characterized NNRTI resistant mutants. The IC50 values of RPV and the RPV analogs were determined using a vector that carries WT RT and several well-known NNRTI resistant mutants in a single round infection assay. Error bars represent thestandard deviations of independent experiments, n = 4. The IC50 values of the graph are at a maximum value of 100 nM and the IC50 value of 26 against the M230L and L100I/K103N resistant mutants was beyond the point of detection in our single round infection assay, >100 nM. To better illustrate the lower IC50 values, the Y-axis is broken from 20 to 40 nM and also shown in the corresponding bars
Susceptibility of mutants selected by DOR to inhibition by RPV analogs
DOR is a new NNRTI currently undergoing evaluation in Phase III clinical trials [19]. DOR was able to inhibit the replication of several well-known NNRTI-resistant mutants but readily selected resistant mutants in in vitro experiments [20]. V106A was the major variant selected in vitro and it was often accompanied by additional mutations, such as F227L, L234I, and F227L/L234I. We screened a larger panel of NNRTI-resistant mutants for their susceptibility to DOR and RPV (unpublished observations). DOR and RPV appear to differ in their susceptibility to NNRTI resistance mutations. We screened the RPV analogs against the mutants that were selected by DOR: V106A, 234I, V106A/F227L, V106/L234I, and V106A/F227L/L234I (Fig. 3; Additional file 1: Table S2). All 5 of these mutants were effectively inhibited by RPV and 6, 7, 8, 9, 11, 12, 13, 14, 15, 16, 17, 21, and 27 (all of the tested compounds had IC50s < 5 nM except 26; 7.7 ± 2.3).

Antiviral Activities of RPV and RPV analogs against mutants selected by DOR in cell culture. RPV and the RPV analogs were tested against several mutants selected by DOR in cell culture. The IC50 values for RPV and the RPV analogs were measured using a single round infection assay. Error bars represent the standard deviations of independent experiments, n = 4. The IC50 values of the graph are at a maximum value of 100 nM. To better illustrate the lower IC50 values, the Y-axis is broken from 10 to 20 nM and also shown in the corresponding bars
Susceptibility of viruses carrying RPV-resistance mutations to the RPV analogs
In individuals who fail RPV-containing regimens, the E138K mutation is one of the most common NNRTI-resistance mutations. The E138K mutation is frequently associated with M184V or M184I, which are presumably selected by FTC or 3TC, one of which was also present in the regimen [23, 24]. K101E is a resistance mutation that is commonly selected by RPV and ETR, an NNRTI that is related to RPV. In addition, we selected for mutants in cell culture by passaging replication competent HIV-1 in the presence of 11 and obtained viruses that carry additional mutations: E40K, D67E, V111A, E138K, Y181C, and M230I. The selection of viruses carrying mutations at E138K and Y181C was expected because of the contacts made between 11 and the residues of the NNRTI binding pocket; M230I is near the NNRTI binding pocket. Thus, these mutations may interfere with the binding of 11 into the NNRTI binding pocket. The V111A mutation is close in proximity to the triad of aspartic acids (D110, D185, and D186) that comprise the polymerase active site; it is not clear how this mutation could directly affect NNRTI binding. Similarly, the mutations at E40K and D67E are in the fingers subdomain of RT, distant from the NNRTI binding pocket and should not affect NNRTI binding. As discussed earlier, we previously measured the effects of the Y181C and M230L (which should be similar to M230I) mutations on the susceptibility of the vector to these compounds [21]. The RT residues E138 in the RT p51 subunit and K101 of the RT p66 subunit are important because they form a salt bridge at the rim of the NNRTI binding pocket and act as “gate-keepers” that inhibit the entry of NNRTIs into the pocket [25, 26]. The amine linkers of RPV make hydrogen bond contacts with the main chain carbonyls of K101 and E138, which help to compensate for the loss of the interaction between the acidic side chains of these two amino acids when RPV is bound in the NNRTI binding pocket [27]. To determine if our RPV analogs could inhibit the replication of mutants selected in the presence of RPV, we tested RPV and the RPV analogs 6, 7, 8, 9, 11, 12, 13, 14, 15, 16, 17, 21, 26, and 27 for their ability to inhibit the replication of: E40K, D67E, K101E, V111A, E138K, M184I, M184V, K101E/M184I, K101E/M184V, E138K/M184I, and E138K/M184V (Fig. 4; Additional file 1: Table S3). Although the M184I/V mutations should not directly affect the binding of an NNRTI, some of the viruses selected in patients in trials that included RPV and an NRTI had one of these mutations, and we wanted to make sure that the M184I/V mutations did not affect the impact of the K101E or E138K mutations on the susceptibility of the vector to any of the RPV analogs. RPV and the analogs 6, 7, 11, 13, 14, 16, 21, and 27 potently inhibited the replication (<2.5 nM) of E40K, D67E, K101E, V111A, E138K, M184I, M184V, K101E/M184I, K101E/M184V, E138K/M184I, and E138K/M184V. The RPV analogs 8, 9, and 15 were almost as potent (<6 nM) against these mutants, except K101E, which was less susceptible to 8 (10.1 nM ± 4), and D67E, which caused a reduction in susceptibility to both 8 and 15 (12.7 nM ± 0.2 and 13.1 nM ± 1, respectively). RPV and the analogs 6, 13, 14, and 27 had comparable antiviral activities against this set of mutants while 11 showed a modestly enhanced susceptibility profile. Analog 17 showed consistently lower potencies against E40K (6.4 nM ± 0.8), D67E (26 nM ± 0.07), K101E (8.9 nM ± 0.07), E138K (13.5 nM ± 4), K101E/M184I (15 nM ± 4.9), and K101E/M184V (8.1 nM ± 2.5). The D67E mutation caused a decrease in susceptibility to the some of the RPV analogs 8, 12, 15, and 17, which is surprising, because, as mentioned earlier, this mutation is not in or near the NNRTI binding pocket. Analog 26 was relatively ineffective against this set of mutants. Overall, it seems that either a pyrimidine or purine is acceptable as a central scaffold as long as amine linkers are present that can interact with K101 of the p66 subunit and E138 of the p51 subunit.

Antiviral activities of RPV and RPV analogs against mutants selected by RPV. The IC50 values of RPV and the RPV analogs were measured against mutants found in RPV-treated patients (K101E, E138K, M184I, M184V, K101E/M184I, K101E/M184V, E138K/M184I, and E138K/M184V) or selected by RPV in cell culture (E40K, D67E, and V111A) using a single round infection assay. Error bars represent the standard deviations of independent experiments, n = 4. The IC50 values of the graph are at a maximum value of 100 nM. To better illustrate the lower IC50 values, the Y-axis is broken from 10 to 20 nM and also shown in the corresponding bars
In addition to the E138K resistance pathway, there is another RPV resistance pathway based on K101P [18, 20]. We tested RPV and the RPV analogs 6, 7, 8, 9, 11, 12, 13, 14, 15, 16, 17, 21, 26, and 27 against the mutants K101P, Y181I, and K101P/V179I (Fig. 5; Additional file 1: Table S4). As has already been discussed, we previously tested these compounds against the Y181C mutant [21]. The K101P mutant showed a modest decease in susceptibility to RPV (6.2 nM ± 1.6) and the analogs 6 (2.5 nM ± 0.92), 7 (9 nM ± 2.3), 8 (9.9 nM ± 2.5), 11 (2.4 nM ± .14), 13 (18.1 nM ± 1.6), and 27 (4.9 nM ± 2.2) and decreased the susceptibility above the limits of detection in our assay (>100 nM) for the analogs 9, 12, 14, 15, 16, 21, and 26. The Y181I mutant showed a small reduction in sensitivity to RPV (8.8 nM ± 0.12), 11 (2.4 nM ± 0.6), and 13 (1.2 nM ± .06) and showed a more significant loss in susceptibility to 6 (55.3 nM ± 1), 7 (19 nM ± 2.6), 8 (74.8 nM ± 15.5), 15 (50.2 nM ± 0.85), 16 (63.2 nM ± 9.1), 17 (78.9 nM ± 8.8), and 27 (23.1 nM ± 4.5) and a greater loss of susceptibility (>100 nM) for 9, 12, 14, 21, and 26. This shows, in terms of susceptibility of the mutants to these compounds, that the impact of the Y181I mutation was considerably greater than the effect of the Y181C mutation [21]. The K101P/Y179I double mutant caused a minor reduction in susceptibility to 6 (12.6 nM ± 2.4), 11 (2.5 nM ± 0.95), and 27 (16.6 nM ± 0.5), a greater decrease in susceptibility to RPV, 7, and 8, and a large loss of susceptibility (>100 nM) to 9, 12, 13, 14, 15, 16, 17, 21, and 26. As had been true with previous sets of NNRTI-resistant mutants, analog 11 was the most broadly effective in inhibiting the replication of this set of mutants. The K101P/Y179I double mutant was the first mutant we have tested that caused a substantial decrease in susceptibility to RPV; this highlights the significance of the fact that this double mutant does not significantly reduce the potency of 11. The analogs 6 and 27 were also broadly effective against this panel of resistant mutants.

Antiviral activities of RPV and the RPV analogs against mutants selected by RPV. The IC50 values for RPV and the RPV analogs against mutants selected by RPV were measured by using a single round infection assay. Error bars represent the standard deviations of independent experiments, n = 4. The IC50 values of the graph are at a maximum value of 100 nM. The IC50 value of several of the RPV analogs against the RPV resistant mutants were beyond the point of detection in our single round infection assay, >100 nM
Modeling the binding of 11 to HIV-1 RT
Because of the improved antiviral profile of 11 versus the NNRTI-resistant mutants that we tested, we wanted to understand how it binds to HIV-1 RT, and the contacts it makes in the NNRTI binding pocket (Fig. 6). We docked 11 in the NNRTI binding pocket using the crystal structure of RPV bound to HIV-1 RT as the template to model the binding of 11 [28]. The interactions of 11 with the NNRTI binding pocket are similar to the interactions of RPV and the pocket, except that the 2,6-purine ring system of 11 resides deeper in the pocket (>1 Å). The benzonitrile linker amine is positioned similarly to the corresponding element of RPV and hydrogen bonds with the carbonyl of K101, while the dimethylphenyl linker amine resides in the general vicinity as that of RPV (~1 Å away) and most likely interacts with water to form a hydrogen bond network with E138 of the RT p51 subunit. The cyanovinyl that is attached to the dimethylphenyl moiety of 11 is ~0.5 Å deeper in the binding pocket than that of RPV (Fig. 6, purple circle), while the benzonitrile moiety of 11 is deeper (~1 Å) in the NNRTI binding pocket compared to RPV (Fig. 6, red circle). When RPV is modeled into the NNRTI binding pocket of RT with the mutations K101P and Y179I (data not shown), there is steric clash between pyrimidine ring of RPV and P101 and I179. It appears that 11 avoids this clash by a slight bend in the central purine ring and by extending the cyanovinyl constituent deeper in the hydrophobic tunnel as well as bending, turning, and extending the benzonitrile moiety towards the back of the NNRTI binding pocket (~2 Å).

RPV and compound 11 in the NNRTI binding pocket. RPV (maroon) and compound 11 (green) are shown bound in the HIV-1 RT NNRTI binding pocket; 11 was docked onto the RPV crystal structure. The residues shown in blue are important contacts for the binding of RPV and denote residues where mutations are selected by the drug. The residues shown in orange are important for the interaction of DOR with the NNRTI binding pocket; DOR selects for mutations at these positions. The red dashes represent hydrogen bonds between K101 and 11 and RPV and the hydrogen bond network between with water and E138 and 11 and RPV. The purple circle denotes the difference in the cyanovinyl binding depth between 11 and RPV, and the red circle shows the difference in the benzonitrile binding depth between 11 and RPV
RPV is the most recent NNRTI to be FDA-approved for the treatment of HIV-1 infection. It was designed and developed to effectively inhibit the replication of a number of common NNRTI-resistant mutants. RPV is comprised of three linked rings, and it is smaller than the largest compounds that can bind in the NNRTI binding pocket. As a consequence, it has the ability, when bound to mutant RTs, to undergo conformational changes and shifts in its binding position that allow it to bind tightly to both the WT and mutant forms of RT, altering its conformation to adapt to the changes in the geometry of the binding pocket. However, recent data suggest that RPV is susceptible to some NNRTI binding pocket mutations [16–18]. We have made additional RPV analogs to determine whether structural changes would allow the new derivatives to be more effective against mutants that reduce the efficacy of RPV.
To evaluate the potential of our RPV analogs relative to RPV, the related drug ETR, and DOR, we examined antiviral efficacies of the compounds against a broad panel of NNRTI-resistant HIV-1 mutants. Most of the RPV analogs exhibited excellent antiviral activities against mutants that were selected by first generation NNRTIs and the new compound DOR, which is in late stage clinical trials [19]. In particular, most of the RPV analogs displayed strong antiviral activities against mutants that contain E138K. More importantly, a double mutant, K101P/Y179I, which showed a significant drop in susceptibility to RPV, remained susceptible to analog 11, demonstrating that it has an antiviral profile that is broader that any of the NNRTIs we have tested thus far.
Different mutation can arise at position Y181, including Y181I and Y181C. Y181C is susceptible to RPV and RPV analogs as previously described [21]. However, Y181I caused a substantial decrease in susceptibility to the RPV analogs, except for 11 and 13 (2.4 nM ± .6 and 1.2 nM ± .06, respectfully); a minor drop in susceptibility to RPV (8.8 nM ± .12) was observed. The isoleucine side chain at position 181 could cause a steric clash with RPV and most of the RPV analogs that would prevent an interaction with Y183 and the cyanovinyl modification. This interaction is thought to compensate for the disruption of the π-π stacking interaction between the aromatic side chain of Y181 and the phenyl moiety of RPV (and most of the RPV analogs). The interaction with Y183 can still occur with the Y181C mutant [28].
11 has a structure that is distinct from the other RPV analogs. The central 2,6-purine ring system and the aryl amines of 11 are in a similar conformation to the central pyrimidine ring of the parent compound RPV, as opposed to 12, 13, 14, 15, 16, 17, and 21, which are in a “flipped” conformation. In this arrangement, the cyanovinyl functionality can extend deeper into the hydrophobic tunnel and the benzonitrile moiety can move more towards the back of the NNRTI binding pocket. In addition to the deeper binding of the aryl moieties of 11, relative to RPV, the overall structure and size of 11 could allow it (like RPV) to use its torsional flexibility in the NNRTI binding pocket in response to mutations in the NNRTI binding pocket [14, 28].
We also investigated why some of the compounds in the series are much less effective than RPV and 11. Docking 26 into the NNRTI binding pocket (Fig. 7) and comparing its binding to the binding of RPV, points to the importance of a hydrogen bond between the amine linker of the RPV analogs and the main chain carbonyl of K101 as well as hydrogen bond network between the water molecule, E138 of the RT p51 subunit, and amine linker of RPV analogs. 27 is a promising analog that effectively inhibits the double mutant K101P/V179I, which has a reduced susceptibility to RPV. It is possible that 27 is effective because of the flexibility of its 2,9-purine ring system and the similarities in the orientation of the central ring to RPV and 11. These two compounds, 11 and 27, could be used as leads to generate additional modifications that could improve contacts with the NNRTI binding pocket of WT HIV-1 RT and, more importantly, with mutant forms of RT.

Binding of RPV and compound 26 in the NNRTI binding pocket. Using the crystal structure of RPV (maroon) in the NNRTI binding pocket as a template, 26 (green) was docked into the HIV-1 RT NNRTI binding pocket. The residues shown in blue are important contacts for the binding of RPV; RPV selects for mutations at these residues. The residues shown in orange are important for the interaction of DOR in the NNRTI binding pocket; DOR selects for mutations at these positions. The red circle shows where amine linker of RPV resides, which is a structural feature 26 lacks. Thus 26 fails to interact with main chain carbonyl of K101, suggesting that this interaction, made by RPV, is important. The purple circle highlights a hydrogen bond interaction between the pyrimidine of RPV and the main chain amide of K101, which 26 fails to make, presumably due to its lack of the amine linker. The red dashes also depict a hydrogen bond network between water and E138 and RPV and 11 of the RT p51 subunit
Conclusions
The wide range of NNRTI-resistant mutants that have been identified shows how daunting the challenges are for designing new NNRTIs for the treatment of HIV-1 infection. More and better compounds should be developed to achieve the goal of suppressing the emerging resistant HIV-1 mutants. One possible approach is to identify combinations of two NNRTIs that have non-overlapping resistance profiles and then use them in combination (unpublished observations), rather than trying to prepare a single compound that is effective against the growing array of NNRTI-resistant mutants [29]. Preventing the emergence HIV-1 resistance is a much better and safer treatment strategy than attempting to deal with resistant mutants after they have been selected. However, that strategy does not mean that we should abandon the search for more effective compounds. NNRTIs offer better toxicity profiles than NRTIs and protease inhibitors (PIs) [30, 31]. 11 and 27 represent a promising step in what should be an ongoing process for the development of future NNRTIs. It also appears, based on what is now known, that DOR, or perhaps some improved DOR derivative, could be useful if it was given in combination with these RPV analogs.
Methods
Cell-based assays
The human osteosarcoma cell line, HOS, was obtained from Dr. Richard Schwartz (Michigan State University, East Lansing, MI) and grown in Dulbecco’s modified Eagle’s medium (Invitrogen, Carlsbad, CA) supplemented with 5 % (v/v) fetal bovine serum, 5 % newborn calf serum, and penicillin (50 units/ml) plus streptomycin (50 µg/ml; Quality Biological, Gaithersburg, MD). Virion production and single-round infectivity assays were used to determine antiviral activity (IC50 values) of the compounds as previously described [32].
Selection of NNRTI mutations in HIV-1
HuT-CCR5 cells were maintained in RPMI 1640 medium (Life Technologies), 10 % fetal bovine serum (Atlanta Biologicals), 100 U/ml penicillin (Life Technologies), 100 μg/ml streptomycin (Life Technologies), 0.292 mg/ml l-glutamine (Life Technologies), 0.5 μg/ml puromycin (EMD Millipore), and 100 μg/ml G418 (Life Technologies). HuT-CCR5 cells were infected with HIV-1LAI for 2 h at a multiplicity of infection of 0.1–0.01. Cells were cultured in the presence or absence of RPV, analog 7, or analog 11. Viral RNA from the supernatant was isolated and used as template in RT-PCR assays with primers spanning the RT coding region. PCR product was sequenced to identify any RT mutations.
Vector constructs
pNLNgoMIVR-ΔEnv.LUC has been described previously [33]. The RT open reading frame was removed from pNLNgoMIVR-ΔEnv.LUC (digestion with SpeI and SalI) and inserted between the SpeI and SalI sites of pBluescript II KS+. Using that construct as the wild-type template, we prepared the following HIV-1 RT mutants using the QuikChange II XL (Stratagene, La Jolla, CA) site-directed mutagenesis protocol kit: L100I, K103N, Y181C, Y188L, H221Y, K103N/Y181C, G190A, G190S, M230L, P236L, L100I/K103N, K103N/P225H, V106A/G190A/F227L, V106A, L234I, V106A/F227L, V106A/L234I, V106A/F227L/L234I, E40K, D67E, K101E, V111A, E138K, M184I, M184V, K101E/M184I, K101E/M184V, E138K/M184I, E138K/M184V, K101P, Y181I, and K101P/V179I. The following sense oligonucleotides were used with matching cognate antisense oligonucleotides (not shown) (Integrated DNA Technologies, Coralville, IA) in the mutagenesis: L100I, 5′-CATCCCGCAGGGATAAAAAAGAAAAAATCA-3′; L100I 2-103, 5′-ATACCACATCCCGCAGGGATTAAAAAGAATAAATCAGTA-3′; K103N, 5′-GCAGGGTTAAAAAAGAATAAATCAGTAACAGTA-3′; Y181C, 5′-CCAGACATAGTTATCTGTCAATACATGGATGAT-3′; Y188L, 5′-TACATGGATGATTTGCTAGTAGGATCTGACTTA-3′; H221Y, 5′-ACACCAGACAAAAAATATCAGAAAGAACCTCCA-3′; G190A, 5′-ATGGATGATTTGTATGTAGCATCTGACTTAGAAATAGGG-3′; G190S, 5′-ATGGATGATTTGTATGTAAGTTCTGACTTAGAAATAGGG-3′; P225H, 5′-AAAAAACATCAGAAAGAACATCCATTCCTTTGGATGGGT-3′; F227, 5′-CATCAGAAAGAACCTCCATTACTTTGGATGGGTTATGAA-3′; M230L, 5′-GAACCTCCATTCCTTTGGCTGGGTTATGAACTCCATCCT-3′; P236L, 5′-ATGGGTTATGAACTGCATCTCGATAAATGGACAGTACAG-3′; V106A, 5′-AAAAAGAAAAAATCAGCAACAGTACTGGATGTG-3′; L234I, 5′-TTCCTTTGGATGGGTTATGAAATCCATCCTGATAAATGGACAGTA-3′; E40K, 5′-GAAATTTGTACAAAAATGGAAAAGGAAGGG-3′; D67E, 5′-GCCATAAAGAAAAAAGAAAGTACTAAATGGAGA-3′; K101E, 5′-CATCCCGCAGGGTTAGAAAAGAAAAAATCAGTAACA-3′; V111A, 5′-GTAACAGTACTGGATGTAGGTGATGCATATTTTTCA-3′; E138K, 5′-CCTAGTATAAACAATAAGACACCAGGGATTAGA-3′; M184I, 5′-GTTATCTATCAATACATAGATGATTTGTATGTA-3′; M184V, 5′-GTTATCTATCAATACGTTGATGATTTGTATGTA-3′; K101P, 5′-CCACATCCCGCAGGGTTACCAAAGAAAAAATCAGTAACA-3′; Y181I, 5′-AATCCAGACATAGTTATCATTCAATACATGGATGATTTG-3′; K101P/V179I, 5′-AAACAAAATCCAGACATAATCATCTATCAATACATGGAT-3′. The double mutants K103N/Y181C, K103N/P225H, and L100I/K103N were made using the previously generated K103N mutant and the appropriate oligonucleotides to add the second mutation, Y181C, P225, and L100I respectively. The double mutants V106A/F227L and V106A/L234I were prepared using the previously generated V106A mutant and the appropriate oligonucleotides to add the second mutation, F227L and L234I, respectively. The double mutants K101E/M184I and K101E/M184V were constructed using the previously generated K101E mutant and the appropriate oligonucleotides to add the second mutation, M184I and M184V, respectively. The double mutants E138K/M184I and E138K/M184V were constructed using the previously generated E138K mutant and the appropriate oligonucleotides for the second mutation, M184I and M184V, respectively. The triple mutant V106A/F227L/L234I was made using the previously generated V106A/F227L double mutant and the appropriate oligonucleotides for the third mutation, L234I. The triple mutant V106A/G190A/F227L was constructed in a series of steps using the previously generated V106A mutant and the appropriate oligonucleotides for the second mutation, G190A, to generate the double mutant V106A/G190A, and then the triple mutant was made using double mutant V106A/G190A and oligonucleotides for the third mutation, F227L. The DNA sequence of each construct was verified by DNA sequence determination. The sequences encoding the mutant RTs were then subcloned into pNLNgoMIVR-ΔEnv.LUC (between the KpnI and SalI sites) to produce the mutant HIV-1 constructs. These DNA sequences were also checked independently by DNA sequence determination.
Computer modeling
All computer modeling was performed using MOE2014.09 (Chemical Computing Group, Montreal, Quebec, Canada). The previously reported crystal structure of WT HIV-1 RT/RPV complex (PDB ID: 2ZD1; [28]) was used in the docking stimulation to model the RPV analogs in the NNRTI binding pocket. The docking simulation used a rigid receptor protocol and refinement to predict the pose of the RPV analogs in the NNRTI binding pocket. We also used an induced fit docking protocol and refinement that permits the sidechains of the NNRTI binding pocket to move to determine whether this would affect predicted binding of the RPV analogs in the NNRTI binding pocket. No new poses were detected.
References
Hartman TL, Buckheit RW Jr. The continuing evolution of HIV-1 therapy: identification and development of novel antiretroviral agents targeting viral and cellular targets. Mol Biol Int. 2012;2012:401965.
Kohlstaedt LA, Wang J, Friedman JM, Rice PA, Steitz TA. Crystal structure at 3.5 A resolution of HIV-1 reverse transcriptase complexed with an inhibitor. Science. 1992;256:1783–90.
Esnouf R, Ren J, Ross C, Jones Y, Stammers D, Stuart D. Mechanism of inhibition of HIV-1 reverse transcriptase by non-nucleoside inhibitors. Nat Struct Biol. 1995;2:303–8.
Das K, Ding J, Hsiou Y, Clark AD Jr, Moereels H, Koymans L, Andries K, Pauwels R, Janssen PA, Boyer PL, Clark P, Smith RH Jr, Kroeger Smith MB, Michejda CJ, Hughes SH, Arnold E. Crystal structures of 8-Cl and 9-Cl TIBO complexed with wild-type HIV-1 RT and 8-Cl TIBO complexed with the Tyr181Cys HIV-1 RT drug-resistant mutant. J Mol Biol. 1996;264:1085–100.
Hsiou Y, Ding J, Das K, Clark AD Jr, Hughes SH, Arnold E. Structure of unliganded HIV-1 reverse transcriptase at 2.7 A resolution: implications of conformational changes for polymerization and inhibition mechanisms. Structure. 1996;4:853–60.
Das K, Martinez SE, Bauman JD, Arnold E. HIV-1 reverse transcriptase complex with DNA and nevirapine reveals non-nucleoside inhibition mechanism. Nat Struct Mol Biol. 2012;19:253–9.
Rittinger K, Divita G, Goody RS. Human immunodeficiency virus reverse transcriptase substrate-induced conformational changes and the mechanism of inhibition by nonnucleoside inhibitors. Proc Natl Acad Sci USA. 1995;92:8046–9.
Spence RA, Kati WM, Anderson KS, Johnson KA. Mechanism of inhibition of HIV-1 reverse transcriptase by nonnucleoside inhibitors. Science. 1995;267:988–93.
Das K, Arnold E. HIV-1 reverse transcriptase and antiviral drug resistance. Part 1. Curr Opin Virol. 2013;3:111–8.
Coffin JM. HIV population dynamics in vivo: implications for genetic variation, pathogenesis, and therapy. Science. 1995;267:483–9.
Mansky LM, Temin HM. Lower in vivo mutation rate of human immunodeficiency virus type 1 than that predicted from the fidelity of purified reverse transcriptase. J Virol. 1995;69:5087–94.
Abram ME, Ferris AL, Shao W, Alvord WG, Hughes SH. Nature, position, and frequency of mutations made in a single cycle of HIV-1 replication. J Virol. 2010;84:9864–78.
Das K, Lewi PJ, Hughes SH, Arnold E. Crystallography and the design of anti-AIDS drugs: conformational flexibility and positional adaptability are important in the design of non-nucleoside HIV-1 reverse transcriptase inhibitors. Prog Biophys Mol Biol. 2005;88:209–31.
Das K, Clark AD Jr, Lewi PJ, Heeres J, De Jonge MR, Koymans LM, Vinkers HM, Daeyaert F, Ludovici DW, Kukla MJ, De Corte B, Kavash RW, Ho CY, Ye H, Lichtenstein MA, Andries K, Pauwels R, De Bethune MP, Boyer PL, Clark P, Hughes SH, Janssen PA, Arnold E. Roles of conformational and positional adaptability in structure-based design of TMC125-R165335 (etravirine) and related non-nucleoside reverse transcriptase inhibitors that are highly potent and effective against wild-type and drug-resistant HIV-1 variants. J Med Chem. 2004;47:2550–60.
Lazzarin A, Campbell T, Clotet B, Johnson M, Katlama C, Moll A, Towner W, Trottier B, Peeters M, Vingerhoets J, de Smedt G, Baeten B, Beets G, Sinha R, Woodfall B, Group D-S. Efficacy and safety of TMC125 (etravirine) in treatment-experienced HIV-1-infected patients in DUET-2: 24-week results from a randomised, double-blind, placebo-controlled trial. Lancet. 2007;370:39–48.
Picchio GR, Rimsky LT, Van Eygen V, Haddad M, Napolitano LA, Vingerhoets J. Prevalence in the USA of rilpivirine resistance-associated mutations in clinical samples and effects on phenotypic susceptibility to rilpivirine and etravirine. Antivir Ther. 2014;19:819–23.
Cohen CJ, Molina JM, Cahn P, Clotet B, Fourie J, Grinsztejn B, Wu H, Johnson MA, Saag M, Supparatpinyo K, Crauwels H, Lefebvre E, Rimsky LT, Vanveggel S, Williams P, Boven K, Group ES, Group TS. Efficacy and safety of rilpivirine (TMC278) versus efavirenz at 48 weeks in treatment-naive HIV-1-infected patients: pooled results from the phase 3 double-blind randomized ECHO and THRIVE trials. J Acquir Immune Defic Syndr. 2012;60:33–42.
Rimsky L, Vingerhoets J, Van Eygen V, Eron J, Clotet B, Hoogstoel A, Boven K, Picchio G. Genotypic and phenotypic characterization of HIV-1 isolates obtained from patients on rilpivirine therapy experiencing virologic failure in the phase 3 ECHO and THRIVE studies: 48-week analysis. J Acquir Immune Defic Syndr. 2012;59:39–46.
Lai MT, Feng M, Falgueyret JP, Tawa P, Witmer M, DiStefano D, Li Y, Burch J, Sachs N, Lu M, Cauchon E, Campeau LC, Grobler J, Yan Y, Ducharme Y, Cote B, Asante-Appiah E, Hazuda DJ, Miller MD. In vitro characterization of MK-1439, a novel HIV-1 nonnucleoside reverse transcriptase inhibitor. Antimicrob Agents Chemother. 2014;58:1652–63.
Feng M, Wang D, Grobler JA, Hazuda DJ, Miller MD, Lai MT. In vitro resistance selection with doravirine (MK-1439), a novel nonnucleoside reverse transcriptase inhibitor with distinct mutation development pathways. Antimicrob Agents Chemother. 2015;59:590–8.
Johnson BC, Pauly GT, Rai G, Patel D, Bauman JD, Baker HL, Das K, Schneider JP, Maloney DJ, Arnold E, Thomas CJ, Hughes SH. A comparison of the ability of rilpivirine (TMC278) and selected analogues to inhibit clinically relevant HIV-1 reverse transcriptase mutants. Retrovirology. 2012;9:99.
Hsiou Y, Ding J, Das K, Clark AD Jr, Boyer PL, Lewi P, Janssen PA, Kleim JP, Rosner M, Hughes SH, Arnold E. The Lys103Asn mutation of HIV-1 RT: a novel mechanism of drug resistance. J Mol Biol. 2001;309:437–45.
Sarafianos SG, Das K, Clark AD Jr, Ding J, Boyer PL, Hughes SH, Arnold E. Lamivudine (3TC) resistance in HIV-1 reverse transcriptase involves steric hindrance with beta-branched amino acids. Proc Natl Acad Sci USA. 1999;96:10027–32.
Gao HQ, Boyer PL, Sarafianos SG, Arnold E, Hughes SH. The role of steric hindrance in 3TC resistance of human immunodeficiency virus type-1 reverse transcriptase. J Mol Biol. 2000;300:403–18.
Das K, Bauman JD, Rim AS, Dharia C, Clark AD Jr, Camarasa MJ, Balzarini J, Arnold E. Crystal structure of tert-butyldimethylsilyl-spiroaminooxathioledioxide-thymine (TSAO-T) in complex with HIV-1 reverse transcriptase (RT) redefines the elastic limits of the non-nucleoside inhibitor-binding pocket. J Med Chem. 2011;54:2727–37.
Singh K, Marchand B, Rai DK, Sharma B, Michailidis E, Ryan EM, Matzek KB, Leslie MD, Hagedorn AN, Li Z, Norden PR, Hachiya A, Parniak MA, Xu HT, Wainberg MA, Sarafianos SG. Biochemical mechanism of HIV-1 resistance to rilpivirine. J Biol Chem. 2012;287:38110–23.
Das K, Arnold E. HIV-1 reverse transcriptase and antiviral drug resistance. Part 2. Curr Opin Virol. 2013;3:119–28.
Das K, Bauman JD, Clark AD Jr, Frenkel YV, Lewi PJ, Shatkin AJ, Hughes SH, Arnold E. High-resolution structures of HIV-1 reverse transcriptase/TMC278 complexes: strategic flexibility explains potency against resistance mutations. Proc Natl Acad Sci USA. 2008;105:1466–71.
Li JZ, Paredes R, Ribaudo HJ, Svarovskaia ES, Metzner KJ, Kozal MJ, Hullsiek KH, Balduin M, Jakobsen MR, Geretti AM, Thiebaut R, Ostergaard L, Masquelier B, Johnson JA, Miller MD, Kuritzkes DR. Low-frequency HIV-1 drug resistance mutations and risk of NNRTI-based antiretroviral treatment failure: a systematic review and pooled analysis. JAMA. 2011;305:1327–35.
Margolis AM, Heverling H, Pham PA, Stolbach A. A review of the toxicity of HIV medications. J Med Toxicol. 2014;10:26–39.
Stolbach A, Paziana K, Heverling H, Pham P. A review of the toxicity of HIV medications II: interactions with drugs and complementary and alternative medicine products. J Med Toxicol. 2015;11:326–41.
Smith SJ, Hughes SH. Rapid screening of HIV reverse transcriptase and integrase inhibitors. J Vis Exp. 2014;. doi:10.3791/51400.
Zhao XZ, Semenova EA, Vu BC, Maddali K, Marchand C, Hughes SH, Pommier Y, Burke TR Jr. 2,3-Dihydro-6,7-dihydroxy-1H-isoindol-1-one-based HIV-1 integrase inhibitors. J Med Chem. 2008;51:251–9.
Authors’ contributions
SS performed the infectivity assays and conducted the molecular modeling. SS and AA made the vector constructs and DNA preparations. KM conducted NNRTI selection experiments. GP, CT, GR, DM, and JS contributed to synthesis of compounds. SS, ZA, and SH designed the experiments. SS and SH drafted the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The authors would like to thank Teresa Burdette for help in preparing the manuscript and Joseph Meyer for help preparing the figures. This research was supported by the Intramural Research Programs of the National Cancer Institute, the National Human Genome Research Institute, the National Center for Advancing Translational Sciences, the Intramural AIDS Targeted Antiviral Program (IATAP), and NIH Grants R01 AI080290 (ZA) and T32 AI065380 (KM).
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Additional file
12977_2016_244_MOESM1_ESM.pdf
Additional file 1: Table S1–S4. The IC50 values (nM) of RPV and the RPV analogs were determined against the NNRTI resistant mutants were measured by using a single round infection assay, n = 4. The concentrations (nM) are measured by reductions in luciferase reporter activity followed by the standard deviations. In parenthesis is the fold-change that describes the difference between the IC50 value of the NNRTI resistant mutant relative to WT.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Smith, S.J., Pauly, G.T., Akram, A. et al. Rilpivirine analogs potently inhibit drug-resistant HIV-1 mutants. Retrovirology 13, 11 (2016). https://doi.org/10.1186/s12977-016-0244-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12977-016-0244-2