
Abstract
NF-E2-related factor 2 (Nrf2) protein is a basic-region leucine zipper transcription factor that defends against endogenous or exogenous stressors. By inducing several cytoprotective and detoxifying gene expressions, Nrf2 can increase the sensitivity of the cells to oxidants and electrophiles. Transient Nrf2 activation, by its specific activators, has protective roles against carcinogenesis and cancer development. However, permanent activation of Nrf2 promotes various cancer properties, comprising malignant progression, chemo/radio resistance, and poor patient prognosis. Taken together, these findings suggest that reaching an optimal balance between paradoxical functions of Nrf2 in malignancy may render a selective improvement to identify therapeutic strategies in cancer treatment. In this review, we describe lately discovered Nrf2 inducers and inhibitors, and their chemopreventive and/or anticancer activities.
Plain english summary
The Nrf2 pathway signifies one of the most significant cell defense procedures against exogenous or endogenous stressors. Certainly, by increasing the expression of several cytoprotective genes, the transcription factor Nrf2 can shelter cells and tissues from multiple sources of damage including electrophilic, xenobiotic, metabolic, and oxidative stress. Notably, the aberrant activation or accumulation of Nrf2, a common event in many tumors, confers a selective advantage to cancer cells and is connected to malignant progression, therapy resistance, and poor prognosis. Therefore, lately, Nrf2 has arisen as a hopeful target in treatment of cancer, and many struggles have been made to detect therapeutic strategies intended at disrupting its pro-oncogenic role. By summarizing the outcomes from past and recent studies, this review provided an overview concerning the Nrf2 pathway and the molecular mechanisms causing Nrf2 hyperactivation in cancer cells. Finally, this paper also described some of the most promising therapeutic approaches that have been successfully employed to counteract Nrf2 activity in tumors, with a particular emphasis on the development of natural compounds and the adoption of drug repurposing strategies.
Video abstract
Similar content being viewed by others


Background
Despite the numerous efforts that researchers have carried out to improve the cancer outcomes, the overall cancer mortality rate has not significantly diminished over the past 30 years [1,2,3]. Reactive oxygen species (ROS) and reactive nitrogen species (RNS) are produced in the cells as the result of normal physiological procedures such as during inflammatory responses and mitochondrial aerobic respiration [4]. The elevated amounts of ROS and RNS in cancer cells can be responsible for triggering oxidative stress, which leads to DNA damage, alterations in tumor-suppressor genes and eventually initiation, development, and progression of cancer [4,5,6]. One of the key systems that confronts oxidative stress and xenobiotics is the Nrf2 signaling system.
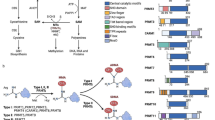
Nuclear factor erythroid 2 [NF-E2]–related factor 2 (also called NFE2L2 or Nrf2) is a basic-region leucine zipper (bZIP) transcription protein and a member of CNC (cap ‘‘n’’ collar) family chiefly localized in the cytoplasm [7]. Kelch-like ECH-associated protein 1 (Keap1) or inhibitor of Nrf2 (INrf2) is a redox-regulated substrate adaptor protein for a Cullin-3/Rbx-1 ubiquitin ligase complex [8]. Under unstressed situations, Nrf2 protein is ubiquitinated by Keap1-Cullin3 E3 ubiquitin ligase enzymes and subsequently degraded by the proteasomal machinery [4, 5]. Upon exposure to electrophiles or oxidative stresses, the interaction between Nrf2 and that complex is interrupted, followed by decreased Nrf2 degradation and increased its translocation into the nucleus where heterodimerized with the small MAF (musculoaponeurotic fibrosarcoma) proteins (MafF, MafK, and MafG) (Fig. 1). These proteins are necessary for the Nrf2-related upregulation of antioxidant response element (ARE)-dependent target genes [5]. In response to the oxidative stress, about 200 cytoprotective genes are regulated by Nrf2 [9,10,11,12]. There are a lot of synthetic or plant-derived chemopreventive compounds, which exert their cancer-preventive roles by involving Nrf2-related defense responses [13,14,15,16,17,18]. Transient activation of Nrf2 is beneficial in countering carcinogens and mutagens and has protective roles versus tumor initiation in normal cells [19,20,21,22]. Howevere, in several of pathological conditions, comprising inflammation and cancer, some of the changes, such as somatic mutations in Nrf2,Keap1,and Cul3, Keap1 modification by metabolic mediators, epigenetically silencing of Keap1, Nrf2 transcriptional activation via oncogene-mediated signaling, and unusual accumulation of the proteins that disrupting the Keap1-Nrf2 interactions, lead to prolonged activation of Nrf2 [23,24,25,26,27,28,29]. Several studies have provided evidence that preventing the permanent activity of Nrf2 by its inhibitors renders cancer cells susceptible to apoptosis and enhances the efficacy of chemotherapeutics [30,31,32,33]. Therefore, Nrf2 is involved not only in biological defense against carcinogenesis and cancer development but also in cancer development and resistance to chemo-and radiotherapy. In the present review, we describe Nrf2 activators and inhibitors to contribute to obtaining an optimal balance between the tumor-preventive or -promoting activities of Nrf2 (Table 1).

Nrf2 pathway under homeostasis and stress conditions. Under unstressed situations, most of Nrf2 protein is ubiquitinated by Keap1-Cullin3 E3 ubiquitin ligase enzymes and subsequently degraded by the proteasomal machinery. Upon exposure to oxidative stresses, the interaction between Nrf2 and that complex is interrupted, which resulted in decreased Nrf2 degradation and increased its translocation into the nucleus where heterodimerized with the small MAF proteins
Molecular mechanisms of Nrf2 regulation in cancer
Somatic mutations in Keap1, Nrf2 and Cullin-3
Initially, somatic mutations of Keap1 were recognized in lung tumor tissues and cell lines [34], which are the second most common and significant genetic modifications in lung cancer [35]. However, Keap1 mutations have also been reported in other human cancers, for example, ovary (19%), gastric (11%), liver (9%), colon (8%), prostate (8%), and breast cancer (2%) [36,37,38]. These mutations were reported in several Keap1 domains, which result in inactivation of Keap1 and accumulation of Nrf2 in the nucleus of cancer cells [39,40,41,42]. Besides the Keap1 mutations, gain of function mutations of Nrf2 have also been identified in human cancers for example esophageal carcinoma, lung, head and neck cancer [23, 27]. Interestingly, almost all Nrf2 mutations occur specifically within either the ETGE (57%) or DLG (43%) motifs [24]. When the mutations occur at the ETGE motif, the high-affinity interaction between the keap1 and Nrf2 is destroyed [26], while DLG motif mutations lead to destruction of low-affinity interaction [27] (Fig. 2). Recently, Ooiet al. [43] identified somatic mutations of Cul3 in sporadic papillary renal cell carcinoma type-2 (PRCC2). They showed that Cul3 mutation can be the trigger of Nrf2 activation in some of the sporadic PRCC2.

Somatic mutations of Keap1 and/or Nrf2 can result in nuclear accumulation of Nrf2
Epigenetic silencing of Keap1
Epigenetic changes are the most common causes of Keap1 gene silencing in solid tumors and are obviously involved in the complicated regulation of the Keap1-Nrf2 system [44]. Multiple studies have indicated that epigenetic alterations in the Keap1 gene donate a growth advantage to cancer cells and are correlated with poor clinical prognosis in cancer patients [23, 45, 46]. The promoter hypermethylation of Keap1 that can lead to reduction of Keap1 expression and Nrf2 accumulation in the nucleus has been identified in malignant glioma [47], lung [48, 49], prostate [38], colorectal [50], gastric [51] and breast cancers [52]. Besides the effects of hypermethylation, the Keap1 expression can be affected by several microRNAs that act as oncogenes or tumor suppressor-microRNAs in tumor cells [53]. For example, miR-7,miR-200a and miR-141 that are overexpressed in neuroblastoma, breast and ovarian cancer, respectively, down-regulate Keap1 expression through binding to the 3′-untranslated region (3′-UTR) of Keap1 mRNA [54,55,56], while inhibition of miR-200a results in up-regulation of Keap1 and subsequently reduction in Nrf2 activation [54].
The role of proteins that disrupting the Nrf2-Keap1 interactions
p21, an inhibitor of cyclin-dependent kinase, is one of the non-electrophilic activators of Nrf2 that has been reported to relate to the DLG motif of Nrf2. Indeed, p21 disrupts two-site binding of Nrf2 to the Keap1 and results in stabilization of Nrf2 protein. Therefore, in the presence of p21, the expression of Nrf2 and its cytoprotective genes is elevated, which followed by promoted cell survival in response to oxidative stress [57, 58].
Another protein involved in the disruption of Keap1-Nrf2 interactions is p62, which links to the autophagy-mediated degradation and contains a Keap1-interacting region (KIR) domain. Interestingly, the STGE (Ser-Thr-Gly-Glu) motif in KIR domain is similar to the Nrf2 ETGE motif and therefore is responsible for the direct interaction between Keap1 and p62 proteins [59,60,61]. It has been suggested that STGE-binding motif in p62 is bound to the Kelch domain of Keap1 with lower affinity than ETGE motif [62]. However, serine phosphorylation (S351) of the p62 STGE motif can remarkably increase this affinity and facilitate p62-dependent autophagic degradation of Keap1 which leading to subsequent activation of Nrf2 [63]. Significantly, the unusual accumulation of p62 has been identified in certain cancers such as hepatocellular carcinoma [64], lung [65, 66], gastric [67, 68], breast [69, 70], and colon cancers [67, 71], which might increase the malignant behavior of these tumors through improving Nrf2 activity.
Oncogene signaling mediated-Nrf2 upregulation
Although the Nrf2 protein level primarily is regulated by the mutation/degradation process, a different study has been conducted on the control of the Nrf2 gene transcription. DeNicola et al. [72] showed that the transcriptional start site of Nrf2 has Jun and Myc binding sites and therefore, the expression of Nrf2 and its downstream genes can be increased remarkably by activating the oncogenic alleles of C-MYC, BRAF, and KRAS (C-MYCERT12, BRAFV619E, and KRASG12D) which followed by more reduction in the intracellular redox environment. In another study, promoter analysis of Nrf2 showed that in regulatory region in exon 1 of Nrf2, a 2-O-tetradecanoylphorbol-13-acetate (TPA) response element (TRE) was activated by KRAS in human non-small cell lung cancer (NSCLC) cells [73]. The oncogenic KRAS can induce antioxidant program through MAPK-mediated Nrf2 activation in pancreatic cancer. Furthermore, KRAS silencing or obstruction of MAP kinase signaling pathway efficiently decrease Nrf2 level and elevate ROS formation [74].
Nrf2 activators
There are many synthetic or extracted substances that function as Nrf2 activators (Fig. 3), which frequently are extracted from plants. Some examples of natural Nrf2 activators include curcumin, sulforaphane (SF), kahweol, resveratrol, garlic oganosulfur compounds, zerumbone, epigallocatechin-3-gallate, carnosol, cinnamonyl-based compounds, lycopene, and cafestol [75,76,77]. Magesh et al. [78] have categorized about 90 kinds of these synthetic or natural activators of Nrf2 in several groups: (1) isothiocyanates and sulfoxythiocarbamates; (2) oxidizable phenols and quinones; (3) Michael acceptors; (4) vicinal dimercaptans; (5) trivalent arsenicals; (6) dithiolethiones and diallyl sulfides; (7) heavy metals and metal complexes; (8) miscellaneous inducers; (9) selenium-based compounds; (10) polyenes; and (11) hydroxyl peroxides. By inducing the Nrf2-mediated defense response, these chemopreventive agents can activate the antioxidants, phase II detoxification factors, and transducers, and protect the cells from carcinogenic exposure [23].

The common activators and inhibitors of Nrf2
Sulforaphane [1-isothiocyanato-4-(methylsulfonyl)-butane] (SFN)
SFN is an effective dietary isothiocyanate, which found in cruciferous plants like Brussels sprouts and broccoli. SFN has anti-genotoxicity, anti-cancer and antioxidant activity as well as chemotherapeutic effect [79,80,81]. Besides enhancing cellular capacity in defense against oxidants, and electrophiles, it has been shown that sulforaphane is able to provoke apoptosis and reduce angiogenesis and cell cycle progression [82,83,84]. Interesting preclinical investigations show that sulforaphane prevents mice from forming carcinogen-mediated mammary carcinogenesis, lung, and gastric cancer, as well as colonic crypt foci [7, 85]. Previously, SFN was recognized as the most powerful activator of NQO1, but later it has been proved that the upregulation of NQO1 by SFN, indeed is regulated by the Nrf2-Keap1 signaling [86]. According to the in vivo experiments, site-directed mutagenesis and mass spectrometry analysis, it was evidenced that SFN can directly modify critical Keap1 cysteine 151 which followed by activation of the Nrf2-Keap1-ARE signaling [87, 88]. Thiols from Keap1 Kelch domain and isothiocyanate from SFN are covalently bound together which lead to the releasing of Nrf2 from Keap1 and finally inducing phase II metabolic enzymes [89, 90]. In another study, Kobayashi et al. categorized SFN as class 1 ARE inducers. Moreover, they found that the induction of ARE-regulated genes in zebrafish via sulforaphane is highly dependent on keap1 Cys151 [88]. It has been shown that SFN has a protective role against cancer development in different kinds of transgenic and carcinogen-induced tumor models (reviewed [91, 92]). In an animal study, Kalpana et al. revealed the inhibitory effect of SFN on benzo(a)pyrene (B(a)P)-induced lung cancer in the mouse, with emphasis on the effect of SFN on Nrf2 signal pathway [93]. Besides, in some clinical trial studies, the effect of SFN, as an Nrf2 activator, was evaluated in patients with colon, prostate, breast, and pancreatic cancers [79,80,81,82]. SFN can concomitantly upregulate Nrf2 and its downstream target genes, including HO1, NQOs, GSTs, and UGTs, rapidly in less than 30 min [94]. Another study demonstrated that sulforaphane epigenetically restored Nrf2 mRNA expression through the demethylation of its promoter CpGs in TRAMP-C1 and JB6 cells [95, 96].
Oltipraz (4-methyl-5-[2-pyrazinyl]-1,2-dithiole- 3-thione) (OPZ)
OPZ, also known as a dithiolthione substitute, is one of the synthetic Nrf2 inducers [97]. Previous studies showed that OPZ can induce a number of Phase I and II antioxidant enzymes, especially that ones that increase glutathione levels, such as glucose-6-phosphate dehydrogenase, glutathione S-transferase (GST) and glutathione reductase [98,99,100,101]. The induction of GST and other cytoprotective enzymes has been termed the “antioxidant response”, which implies the possible impact of OPZ in cancer prevention [102, 103]. Moreover, OPZ is effective against many different kinds of common carcinogens such as 2-amino-1-methyl-6- phenylimidazo [4,5-b] pyridine (PhIP), aflatoxin B1 (AFB1) and benzo[a]pyrene (B[a]P) [104]. In another study, Sharma et al. demonstrated that oltipraz inhalation spray could inhibit B(a)P-mediated lung carcinoma in mice [105].
OPZ also may enhance the binding activity of Nrf2 to the ARE [100, 106]. Jia et al. [107] showed that dithiolethiones potentially have the ability to Keap1 cysteine modification. For example, 3H-1,2-dithiole-3-thione, which is structurally similar to OPZ, induces intermolecular disulfide cross-linking between two Keap1 monomers by targeting Cys273 and Cys288 [108]. Moreover, another study proved that Nrf2 is responsible for the chemopreventive effects of OPZ against bladder cancer [109].
Epigallocatechin-3-gallate (EGCG)
EGCG is known to be a potent inducer of Nrf2 among several polyphenols found in green tea extract [110]. EGCG has anti-oxidative stress and anti-inflammatory activities through the downregulation of cyclooxygenase-2 and nitric oxide synthase [111]. According to the previous studies, EGCG has protective activity against experimentally induced prostate [112], fore-stomach [113], lung [114], skin [115], breast [116], and colon cancer [117]. Khan et al. [118] reported that EGCG could inhibit the cell adhesion function and downregulate the expression of matrix metalloproteinases that results in a reduction in angiogenesis, metastasis, and invasion of cancer cells. In addition to its ability to function as an anti-angiogenic agent, EGCG can induce apoptosis in numerous types of cancers by stabilizing the tumor suppressor p53 [119] and inactivating some transcription factors [119, 120].
It seems that EGCG by oxidizing or modifying Keap1 cysteine residues accelerates disassociation of Nrf2-Keap1 complex [121]. Moreover, EGCG is reported to activate Nrf2 via induction of upstream signalings, ERK and PI3K, which led to phosphorylation of Nrf2 serine/threonine residue in human mammary epithelial cells [122]. Other studies have described that EGCG inhibits the expression of Bach1, an Nrf2 competitor for binding to ARE sites (INrf2), in cultured A549 cells [123, 124].
Dimethyl fumarate (tecfidera or DMF)
DMF, a methyl ester of fumaric acid (FA), is a chemical potent activator of Nrf2 [125]. The molecular mechanisms which DMF exerts its effects on Nrf2 are not understood completely but there are some suggested mechanisms. DMF, by activating the Nrf2-dependent anti-oxidant response pathway, stimulates the anti-inflammatory and cytoprotective responses [126, 127]. It also inhibits NF-κB-driven processes [128], and as an α, β carboxylic acid ester is able to bind to the thiol groups and modulate glutathione availability and production [127, 129, 130]. Ahuja et al. [131] demonstrated that DMF and DMF metabolite monomethylfumarate (MMF) activate the Nrf2 signaling through alkylation of Keap1 Cys residues and via induction of Bach1 exclusion from the nucleus. It has been demonstrated that Nrf2 activation by MMF is dose and time-dependent [132]. Furthermore, pre-treatment of neuronal cells with a low concentration of dimethylfumarate, by promoting cellular GSH, could protect them from oxidative glutamate toxicity [133, 134].
DMF-related activation of Nrf2 exerts cytoprotective actions in different cell types, for example, splenocytes, embryonic primary cortical cells, astrocytes, microglial cells, renal fibroblasts, and mesangial cells. Moreover, a growing body of evidence suggests that DMF suppresses proliferation, invasion, and angiogenesis and promotes apoptosis of various cancer cells [135,136,137]. An animal study showed that DMF has pro-apoptotic and anti-proliferative activities in melanoma cells and postpones progression and metastasis of melanoma [138].
Diallyl trisulfide (DATS)
DATS is one of the well-known kinds of isothiocyanates (ITC) found in garlic oil and in a diversity of edible cruciferous vegetables, for example, cabbage, broccoli, and watercress [139]. In ex and in vivo experiments, DATS by modification of the Keap1, Cys288, is able to activate Nrf2 and promote NQO1 and HO-1 expression [140, 141].
In cardiomyoblast cells, the Nrf2 level and its nuclear accumulation, as well as the expression of its target antioxidant enzymes, were significantly higher in the cells treated with DATS. In addition, the Keap1 and GSK3β (enhancer of Nrf2 degradation) protein levels were significantly lower in those cells. Further analysis showed that DATS activates Nrf2 signaling by a MAPK-independent pathway which followed by suppression of hypoglycemia-induced apoptosis [142]. Moreover, it has been demonstrated that DATS can regulate drug metabolism by activation of Nrf2/ARE signaling. Because DATS-mediated detoxifying gene expression was detected in a wild-type mouse, but not in Nrf2 null mouse [143].
Curcumin (CUR)
CUR, as a classical activator of Nrf2, is one of the well-investigated natural chemopreventive agents extracted from turmeric [144] (an Indian spice). CUR has a variety of therapeutic properties comprise of anti-oxidant [145], anti-inflammatory [146, 147] and anti-cancer activities [148, 149]. It also has been shown that CUR, via affecting the expression of AR, NQO1, GST, and HO-1, is able to activate a xenobiotic response in the cells [150, 151]. Subsequent analyses suggested that chemopreventive/therapeutic effects of CUR may be exerted by epigenetic alteration [152]. CUR, at lower concentrations, had demethylating effects on the promoter region of Nrf2, which resulted in elevated expressions of Nrf2 and its target genes [153]. Besides epigenetic modifications in Nrf2, curcumin may indirectly phosphorylate Nrf2 at serine- and/or threonine-rich regions and facilitate the nuclear transition of Nrf2. In addition, it can directly interact with sensor cysteine thiol(s) of Keap1 and diminish its inhibitory effect on Nrf2 [154]. Interestingly, CUR is able to play radiation and chemotherapy sensitizer role in some of the human cancers such as prostate [155], colorectal [156, 157] and ovarian cancer [158]. Some of the clinical trial studies indicated that CUR is quite safe and probably has therapeutic applicability in cancer treatment. Curcumin consumption for 3 months could improve the pre-cancerous lesions of patients with resected uterine cervical intraepithelial neoplasia, intestinal metaplasia, oral leukoplakia, and bladder cancer [159]. Despite the activatory effect of CUR on Nrf2 signaling, it is able to exert inhibitory effects on some other signalings, such as Notch1 [160], NF-kappa B [158] and mitochondrial signaling pathways [161].
2-cyano-3,12-dioxooleana-1,9(11)-dien-28-oic acid(CDDO)
CDDO is a synthetic triterpenoid analog, which applied in two types CDDO-Imidazolide (CDDO-Im) and CDDO-methyl ester (CDDO-Me), and conjugated with electron-withdrawing groups by covalent connection. At low concentrations (in picomolar range), CDDO-Im and CDDO-Me are the most effective known Nrf2 activators that have shown anti-inflammatory, pro-apoptotic, anti-proliferative and cytoprotective properties [162, 163]. Cleasby et al. [19] indicated that CDDO-Im can disrupt the BTB-Cul3 interface via covalent interaction with reactive cysteine 151 in the BTB domain of Keap1. Besides Keap1, CDDO-Im interacts with several targets such as PTEN, JAK1/STAT3, ErbB2, NF-kB, PPAR, mTOR and results in alterations in down-stream events [164, 165]. Yates et al. [163] showed that CDDO-Im may be an effective chemopreventive agent against cancers-mediated by electrophilic carcinogens or -associated with obesity. Another study showed chemopreventive potency of CDDO-Im against aflatoxin-induced hepatic tumorigenesis in an Nrf2-dependent manner [17]. Moreover, a knockout mouse study showed that CDDO-Me selectively induces the Nrf2/Keap1 signaling [166]. Significant alterations in the expression of 43 proteins were detected in the wild-type mice following intraperitoneal injection of CDDO-Me. However, among these altered proteins, only two proteins were similarly affected in Nrf2 null mice, indicating that almost all of these protein alterations were mediated through the Nrf2/Keap1 system. High doses of CDDO-Me (micromolar range) can suppress cancer cell proliferation and provoke apoptosis and cancer cell death in various tumor types [167, 168]. Furthermore, CDDO-Me has a higher potential to reduce lung tumor size in the mouse model in compare with CDDO-Im. Despite less efficacy in this lung cancer model, CDDO-Im has an effective role in the prevention and treatment of many other tumors including breast [169], prostate [170], and liver carcinogenesis [17].
Apigenin (4,5,7-trihydroxyavone (Api))
Api is a kind of flavonoids that plentifully exists in plant-derived beverages and vegetables including orange, onion, parsley, tea, wheat sprouts and chamomile [171]. According to the previous studies Api has different pharmacological properties, for example, antivirus [172], anti-inflammatory [173], anti-oxidant [174] and anticancer activity [175]. In addition, Api, because of low bioactivity and slow pharmacokinetics, effectively accumulates in cells/tissues [176, 177]. Api potently increases the transcription of Nrf2 and leads to the elevated level of phase II detoxification proteins in t-BHP (tert-Butyl hydroperoxide)-treated ARPE cells (Retinal pigment epithelium cells). It is noteworthy that the endogenous mRNA and protein expressions of Nrf2 and its downstream gene, hemeoxygenase-1 (HO-1), considerably are elevated by Api, which followed by cellular protection against oxidative condition [178,179,180]. While knock down or knock out of the Nrf2 by CRISPER/Cas9 system or specific shRNA lead to decreasing the protective effects of Api in oxidative stress conditions [178]. Frequently, it was reported that Api by CpG site demethylation along with attenuated activities of DNA methyltransferase and histone deacetylases capable to restore the silenced status of Nrf2 in skin epidermal JB6 P + cell line [179]. Nevertheless, in contrast with the previous study, it has been reported that Api, via down-regulating of PI3K/Akt pathway, diminishes the expression of Nrf2 at both protein and mRNA levels which leads to a reduced expression of Nrf2-target genes in BEL-7402 cells (human hepatocellular carcinoma cells). Moreover, apigenin, in combination with chrysin, directly inhibits the PI3K/Akt pathway, which is associated with the survival of cancer cells [181,182,183].
Resveratrol (3,5,4-trihydroxystilbene (RES))
Resveratrol, a naturally non-flavonoid polyphenol compound, is found in various food and plants for example: cranberry, mulberry, peanut, and the skin of red grape with different concentrations [184, 185]. Various environmental stress and stimulators, for examples excessive sunlight, UV irradiation, microbial and fungal infection, and mechanical injury, are capable of paving the way for producing the resveratrol by many plants species [184, 186]. Originally, there are two isomeric forms of resveratrol, trans-resveratrol and cis-resveratrol. Although both isomers are biologically active, the early one is more stable isomer and serves as a dominant form in the vast majority of resveratrol’s biological functions [186]. Numerous in vitro and in vivo studies have been illustrated that resveratrol occupies an axial role in modulating the signaling pathways associated with cellular growth and division, apoptosis, angiogenesis, invasion, and metastasis. Furthermore, resveratrol possesses anti-diabetic, anticancer, antioxidant, and anti-inflammatory effects [184,185,186,187]. In breast cancer, resveratrol through increasing the expression of Nrf2 and UGT1A8, an enzyme that can metabolize the catechol estrogen, contributes to the degradation of catechol estrogen [188, 189]. Zhang et al. found that resveratrol were capable of elevating the expression levels of Nrf2, HO-1 and reducing the level of ROS production and Keap1. Additionally, they reported that cell treatment with resveratrol, suppressed cell proliferation and Bcl-2 protein expression and stimulated expression of Bax protein and apoptosis [190]. According to the previous studies, it is remarkable to note that resveratrol is capable of activating the Nrf2/ARE signaling pathway and exerts antioxidant protective effects by regulating the expression of phase II detoxification enzymes [184, 185, 188].
Nrf2 inhibitors
In contrast with the several agents that function as Nrf2 inducers, very few molecular components have been recognized as Nrf2 inhibitors. Since Nrf2 has multifaceted roles in cancer cells, Nrf2 inhibitors can be applied as anticancer agents [27, 77, 191]. Indirectly, Nrf2 inhibitors down-regulate drug detoxifying and eliminating enzymes and sensitize cancer cells to chemotherapeutics [76, 176]. According to the Nrf2 deactivation mechanisms and their potential applications in cancer treatment, several small molecules have been characterized as Nrf2 pathway inhibitors (Fig. 3).
Brusatol (BRU)
Brusatol is a quassinoid which is extracted from Brucea Javanica (Simaroubaceae), an evergreen shrub grown in Northern Australia and Southeast Asia [27]. Initially, it was recognized as an anticancer agent against leukemia [192]. Brusatol changes the Nrf2 protein levels without changing in the Keap1 level [193]. Moreover, brusatol sensitizes various types of cancer cells, including HeLa cells, MDA-MB-231, and A549 cell lines, to some chemotherapeutics such as 5-fluorouracil, carboplatin, paclitaxel as well as etoposide [193]. This Nrf2 inhibitor also reduces tumor burden and improves survival in the A549 xenograft mice model [194]. However, there are several obstacles in the application of the brusatol as a therapeutic agent including toxicity, drug delivery and its effect reversibility that is necessary to be resolved [176]. Although it has been indicated that brusatol has transient and rapid post-transcriptional inhibitory effect on Nrf2 [195], the specificity and precise anticancer mechanism of brusatol have not yet been fully understood.
Luteolin (3′,4′,5,7-tetrahydroxyflavone (LUT))
Luteolin is a natural polyphenolic flavonoid which obtained from various kinds of plants for example broccoli, celery, parsley, perilla leaf, and peppers, and characterized as one of the Nrf2 inhibitors [193, 196]. Luteolin has a wide range of biological effects such as antibacterial, antioxidant, anti-inflammatory and anticancer as well as cytoprotective activities [197]. Luteolin considerably enhances the anticancer efficiency of chemotherapeutic drugs, such as bleomycin, oxaliplatin, and doxorubicin, on A549 cell lines. When applying as an Nrf2 inhibitor, Luteolin reverses the sensitivity of colorectal cancer cells to the chemotherapy agents [198]. Additionally, luteolin is capable to suppress the cell cycle promotion and also acts as an apoptosis-inducer and anti-proliferative agent in several cancers, for example, gastric, prostate and pancreatic cancer, hepatoma, melanoma, leukemia, and epidermoid carcinoma [196]. It should be noted that luteolin in some studies has been categorized as an Nrf2 activator, on the other [180, 199, 200].
Trigonelline (TRG)
Trigonelline, a heterocyclic compound, is widely existing in plants, coffee and fenugreek seed that in comparison with chemicals is less toxic to humans [201]. Recently it has been identified that Trg has anti-diabetic, hypocholesterolemic, antimigraine, anticancer, as well as, Nrf2-inhibitory effects [196, 201] which blocks the Nrf2-dependent expression of proteasomal genes [193]. In another study, Trg reversed resistance to ferroptotic cell death in head and neck cancer by blocking the Nrf2 pathway, both in vitro and in vivo [202]. It has been recognized that using a combination of etoposide and trigonelline can lead to the enhancement of anticancer efficacy of etoposide and reduction in tumor size [196], especially in the tumors have high-level activity of Nrf2 [193].
Ascorbic acid (vitamin C, L-ascorbic acid, AscA, AA)
Ascorbic acid which generally known as an antioxidant agent [176], suppresses the Nrf2/DNA complex [78] and through inhibition of the nucleus translocation of Nrf2, reduces the cellular level of peroxides [176]. Additionally, AA, because of abilities in the hydrogen peroxide generation, is categorized as a pro-oxidant which sensitizes tumor cells to the therapeutics, but sometimes has been observed the opposite effects [176, 203]. Based on S.R. Kim et al. [204] results, AA through involving the Cys151Ser in the Keap1, leading to activation of PI3K/Nrf-2 and finally inducing the HO-1 which has antioxidant effects by increasing the level of reduced glutathione [205]. In another study,Vineetha RC et al. [206] reported AA by elevating the level of oxidative stress leading to the up regulation of Nrf2 and Bcl2 expression.
Retinoic acid (RA)
Retinoic Acid (RA), also known as All-trans-retinoic acid (ATRA), is a metabolite of vitamin A [207]. RA by ARE-inducing elements, for example, tBHQ, decreases the capability of Nrf2 to mediate the induction of ARE-regulated genes in both in vivo and ex vivo conditions [78]. RA has biological functions such as regulating cell differentiation, proliferation, and morphogenesis. RA by stimulation of cellular differentiation and suppression of cell growth inhibits tumorigenesis [208] and also is able to enhance the apoptosis [209] which this property may contribute to the anticancer role of RA [210]. RA exerts its effects by retinoid X receptors (RXR-a, b, and g) and retinoic acid receptors (RAR-a, b, and g), which are specific nuclear receptors. RARs, RXRs and/or other hormone receptors of the nucleus, create heterodimers and act as transcriptional regulators [210], which through binding to the transcription factors like Nrf2, prevent the binding of this transcription factors to the ARE [176]. Furthermore, the interaction between RAR/RXR heterodimers and other transcription factors like estrogen receptor, AP-1 and NF-B leads to the RA activation or repression and subsequently gene expression variations [211]. Although RA has been identified as Nrf2 inhibitor, in some studies it has been reported as an Nrf2 activator [23]. Therefore, prior to using RA as an anticancer agent, it is necessary to carry out additional studies on the specificity and mechanism of its actions.
Chrysin (5,7-dihydroxy-2-phenyl-4H-chromen-4-one (CHR))
Chrysin, a natural flavonoid, is found in many plant extracts including honey, propolis, mushroom, blue passion flower, vegetables, and fruits [212]. According to the literature reports, the most reliable pharmacological activities of chrysin are antioxidant, anti-inflammatory, anti-diabetic, hepatoprotective, neuroprotective, anti-aging, and anticancer effects [213,214,215]. In many studies, it has been reported that chrysin occupies an axial role in many biological process for example suppression the pro-inflammatory cytokines expression, down-regulation of nuclear factor kappa B (NF-kB), tumor necrosis factor α (TNF-α), and interleukin 1β (IL-1β), up-regulation of apoptotic pathways, and inhibition of angiogenesis and metastasis formation [216,217,218]. Gao and colleagues have been reported a higher level of Nrf2 expression in BEL-7402/ADM cells. They showed that chrysin, by quenching ERK and PI3K-Akt pathway, makes a contribution to inhibition of Nrf2 and its downstream target genes as well as AKR1B10, HO-1, and MRP5 expression [219]. Additionally, Zeng et al. [220] disclosed that chrysin treatment promotes the expression of osteogenesis genes in preosteoblast MC3T3-E1 cell lines by activation of ERK/MAPK signaling pathway. In another study, it has been revealed that chrysin reduces the mRNA expression of Nrf2, MRP1, NQO-1, and HO-1 in breast cancer MCF7 cell lines [221]. It is completely obvious that chrysin decrease both mRNA and protein expression levels of Nrf2, however, it should be emphasized that the effects of chrysin on the Nrf2-ARE signaling pathway appears to be cell type-specific, concentration dependent, and may vary depending on the nature of Nrf2 regulation.
Conclusion
Since Nrf2 has paradoxical roles in cancer biology, it is essential to understand the molecular mechanisms leading to tumor suppressor or oncogenic effects of Nrf2. Furthermore, to pave the way for identifying therapeutic strategies based on Nrf2 signaling in malignancy treatment, it has to be considered when the specific Nrf2 inducer or inhibitor is appropriate. However, further studies should be conducted to find cancer targeting drug candidates with good pharmacodynamic/pharmacokinetic parameters for human cancer.
Availability of data and materials
The authors confirm that the data supporting the findings of this study is available within the article.
Abbreviations
- Nrf2:
-
NF-E2-related factor 2
- ROS:
-
Reactive oxygen species
- RNS:
-
Reactive nitrogen species
- bZIP:
-
Basic-region leucine zipper
- MAF:
-
Musculoaponeurotic fibrosarcoma
- SF:
-
Sulforaphane
- GST:
-
Glutathione S-transferase
- EGCG:
-
Epigallocatechin-3-gallate
- MMF:
-
Metabolite monomethyl fumarate
References
Guan X: Cancer metastases: challenges and opportunities. Acta Pharm Sin B. 2015;5:402–418.
Adlravan E, Nejati K, Karimi MA, Mousazadeh H, Abbasi A, Dadashpour M. Potential activity of free and PLGA/PEG nanoencapsulated nasturtium officinale extract in inducing cytotoxicity and apoptosis in human lung carcinoma A549 cells. J Drug Deliv Sci Technol. 2021;61: 102256.
Samadzadeh S, Mousazadeh H, Ghareghomi S, Dadashpour M, Babazadeh M, Zarghami N. In vitro anticancer efficacy of Metformin-loaded PLGA nanofibers towards the post-surgical therapy of lung cancer. J Drug Deliv Sci Technol. 2021;61: 102318.
Abed DA, Goldstein M, Albanyan H, Jin H, Hu L. Discovery of direct inhibitors of Keap1–Nrf2 protein–protein interaction as potential therapeutic and preventive agents. Acta Pharm Sin B. 2015;5:285–99.
Waris G, Ahsan H. Reactive oxygen species: role in the development of cancer and various chronic conditions. J Carcinog. 2006;5:14–14.
Gupta SC, Hevia D, Patchva S, Park B, Koh W, Aggarwal BB. Upsides and downsides of reactive oxygen species for cancer: the roles of reactive oxygen species in tumorigenesis, prevention, and therapy. Antioxid Redox Signal. 2012;16:1295–322.
Moon EJ, Giaccia A. Dual roles of NRF2 in tumor prevention and progression: possible implications in cancer treatment. Free Radical Biol Med. 2015;79:292–9.
Namani A, Li Y, Wang XJ, Tang X. Modulation of NRF2 signaling pathway by nuclear receptors: implications for cancer. Biochim et Biophys Acta (BBA) Mol Cell Res. 2014;1843:1875–85.
Sadeghi MR, Jeddi F, Soozangar N, Somi MH, Shirmohamadi M, Khaze V, Samadi N. Nrf2/P–glycoprotein axis is associated with clinicopathological characteristics in colorectal cancer. Biomed Pharmacother. 2018;104:458–64.
Hayes JD, Dinkova-Kostova AT. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem Sci. 2014;39:199–218.
Lee SB, Sellers BN, DeNicola GM. The regulation of NRF2 by nutrient-responsive signaling and its role in anabolic cancer metabolism. Antioxid Redox Signal. 2018;29:1774–91.
Hammad A, Namani A, Elshaer M, Wang XJ, Tang X. “NRF2 addiction” in lung cancer cells and its impact on cancer therapy. Cancer Lett. 2019;467:40–9.
Kou X, Kirberger M, Yang Y, Chen N. Natural products for cancer prevention associated with Nrf2–ARE pathway. Food Sci Human Wellness. 2013;2:22–8.
Tian Y, Liu Q, He X, Yuan X, Chen Y, Chu Q, Wu K. Emerging roles of Nrf2 signal in non-small cell lung cancer. J Hematol Oncol. 2016;9:14.
Geismann C, Arlt A, Sebens S, Schäfer H. Cytoprotection “gone astray”: Nrf2 and its role in cancer. Onco Targets Ther. 2014;7:1497–518.
Ramos-Gomez M, Dolan PM, Itoh K, Yamamoto M, Kensler TW. Interactive effects of nrf2 genotype and oltipraz on benzo[a]pyrene–DNA adducts and tumor yield in mice. Carcinogenesis. 2003;24:461–7.
Yates MS, Kwak M-K, Egner PA, Groopman JD, Bodreddigari S, Sutter TR, Baumgartner KJ, Roebuck B, Liby KT, Yore MM. Potent protection against aflatoxin-induced tumorigenesis through induction of Nrf2-regulated pathways by the triterpenoid 1-[2-cyano-3-, 12-dioxooleana-1, 9 (11)-dien-28-oyl] imidazole. Cancer Res. 2006;66:2488–94.
Javan N, Khadem Ansari MH, Dadashpour M, Khojastehfard M, Bastami M, Rahmati-Yamchi M, Zarghami N. Synergistic antiproliferative effects of co-nanoencapsulated curcumin and chrysin on mda-mb-231 breast cancer cells through upregulating mir-132 and mir-502c. Nutr Cancer. 2019;71:1201–13.
Cleasby A, Yon J, Day PJ, Richardson C, Tickle IJ, Williams PA, Callahan JF, Carr R, Concha N, Kerns JK. Structure of the BTB domain of Keap1 and its interaction with the triterpenoid antagonist CDDO. PLoS ONE. 2014;9: e98896.
Das L, Vinayak M. Long term effect of curcumin in restoration of tumour suppressor p53 and phase-II antioxidant enzymes via activation of Nrf2 signalling and modulation of inflammation in prevention of cancer. PLoS ONE. 2015;10: e0124000.
Tsai P-Y, Ka S-M, Chang J-M, Chen H-C, Shui H-A, Li C-Y, Hua K-F, Chang W-L, Huang J-J, Yang S-S. Epigallocatechin-3-gallate prevents lupus nephritis development in mice via enhancing the Nrf2 antioxidant pathway and inhibiting NLRP3 inflammasome activation. Free Radical Biol Med. 2011;51:744–54.
Balstad TR, Carlsen H, Myhrstad MC, Kolberg M, Reiersen H, Gilen L, Ebihara K, Paur I, Blomhoff R. Coffee, broccoli and spices are strong inducers of electrophile response element-dependent transcription in vitro and in vivo–Studies in electrophile response element transgenic mice. Mol Nutr Food Res. 2011;55:185–97.
Jaramillo MC, Zhang DD. The emerging role of the Nrf2–Keap1 signaling pathway in cancer. Genes Dev. 2013;27:2179–91.
Govindan R, Hammerman PS, Hayes DN, Wilkerson MD, Baylin S, Meyerson M. Comprehensive genomic characterization of squamous cell carcinoma of the lung. Am Soc Clin Oncol. 2012;30:7006.
Kim YR, Oh JE, Kim MS, Kang MR, Park SW, Han JY, Eom HS, Yoo NJ, Lee SH. Oncogenic NRF2 mutations in squamous cell carcinomas of oesophagus and skin. J Pathol. 2010;220:446–51.
Shibata T, Ohta T, Tong KI, Kokubu A, Odogawa R, Tsuta K, Asamura H, Yamamoto M, Hirohashi S. Cancer related mutations in NRF2 impair its recognition by Keap1-Cul3 E3 ligase and promote malignancy. Proc Natl Acad Sci. 2008;105:13568–73.
Mitsuishi Y, Motohashi H, Yamamoto M. The Keap1–Nrf2 system in cancers: stress response and anabolic metabolism. Front Oncol. 2012;2:200.
Jeddi F, Soozangar N, Sadeghi MR, Somi MH, Shirmohamadi M, Eftekhar-Sadat A-T, Samadi N. Nrf2 overexpression is associated with P-glycoprotein upregulation in gastric cancer. Biomed Pharmacother. 2018;97:286–92.
de la Vega MR, Chapman E, Zhang DD. NRF2 and the hallmarks of cancer. Cancer Cell. 2018;34:21–43.
Torrente L, Sanchez C, Moreno R, Chowdhry S, Cabello P, Isono K, Koseki H, Honda T, Hayes JD, Dinkova-Kostova AT, de la Vega L. Crosstalk between NRF2 and HIPK2 shapes cytoprotective responses. Oncogene. 2017;36:6204.
Bao L, Wu J, Dodson M, Rojo De La Vega EM, Ning Y, Zhang Z, Yao M, Zhang DD, Xu C, Yi X. ABCF2, an Nrf2 target gene, contributes to cisplatin resistance in ovarian cancer cells. Mol Carcinog. 2017;56:1543–53.
Telkoparan-Akillilar P, Suzen S, Saso L. Pharmacological applications of Nrf2 inhibitors as potential antineoplastic drugs. Int J Mol Sci. 2025;2019:20.
Panieri E, Buha A, Telkoparan-Akillilar P, Cevik D, Kouretas D, Veskoukis A, Skaperda Z, Tsatsakis A, Wallace D, Suzen S. Potential applications of NRF2 modulators in cancer therapy. Antioxidants. 2020;9:193.
Singh A, Misra V, Thimmulappa RK, Lee H, Ames S, Hoque MO, Herman JG, Baylin SB, Sidransky D, Gabrielson E. Dysfunctional KEAP1–NRF2 interaction in non-small-cell lung cancer. PLoS Med. 2006;3: e420.
Lawrence MS, Stojanov P, Mermel CH, Robinson JT, Garraway LA, Golub TR, Meyerson M, Gabriel SB, Lander ES, Getz G. Discovery and saturation analysis of cancer genes across 21 tumour types. Nature. 2014;505:495–501.
Yoo NJ, Kim HR, Kim YR, An CH, Lee SH. Somatic mutations of the KEAP1 gene in common solid cancers. Histopathology. 2012;60:943–52.
Konstantinopoulos PA, Spentzos D, Fountzilas E, Francoeur N, Sanisetty S, Grammatikos AP, Hecht JL, Cannistra SA. Keap1 mutations and Nrf2 pathway activation in epithelial ovarian cancer. Cancer Res. 2011;71:5081–9.
Zhang P, Singh A, Yegnasubramanian S, Esopi D, Kombairaju P, Bodas M, Wu H, Bova SG, Biswal S. Loss of Kelch-like ECH-associated protein 1 function in prostate cancer cells causes chemoresistance and radioresistance and promotes tumor growth. Mol Cancer Ther. 2010;9:336–46.
Nioi P, Nguyen T. A mutation of Keap1 found in breast cancer impairs its ability to repress Nrf2 activity. Biochem Biophys Res Commun. 2007;362:816–21.
Shibata T, Kokubu A, Gotoh M, Ojima H, Ohta T, Yamamoto M, Hirohashi S. Genetic alteration of Keap1 confers constitutive Nrf2 activation and resistance to chemotherapy in gallbladder cancer. Gastroenterology. 2008;135(1358–1368):1368.e1351-1354.
Singh A, Misra V, Thimmulappa RK, Lee H, Ames S, Hoque MO, Herman JG, Baylin SB, Sidransky D, Gabrielson E, et al. Dysfunctional KEAP1-NRF2 interaction in non-small-cell lung cancer. PLoS Med. 2006;3: e420.
Takahashi T, Sonobe M, Menju T, Nakayama E, Mino N, Iwakiri S, Nagai S, Sato K, Miyahara R, Okubo K, et al. Mutations in Keap1 are a potential prognostic factor in resected non-small cell lung cancer. J Surg Oncol. 2010;101:500–6.
Ooi A, Dykema K, Ansari A, Petillo D, Snider J, Kahnoski R, Anema J, Craig D, Carpten J, Teh B-T. CUL3 and NRF2 mutations confer an NRF2 activation phenotype in a sporadic form of papillary renal cell carcinoma. Cancer Res. 2013;73:2044–51.
Copple IM. The Keap1–Nrf2 cell defense pathway–a promising therapeutic target? Adv pharmacol. 2012;63:43–79.
Taguchi K, Motohashi H, Yamamoto M. Molecular mechanisms of the Keap1–Nrf2 pathway in stress response and cancer evolution. Genes Cells. 2011;16:123–40.
Brigelius-Flohé R, Müller M, Lippmann D, Kipp AP. The yin and yang of nrf2-regulated selenoproteins in carcinogenesis. Int J Cell Biol. 2012;2012:7–8.
Muscarella LA, Barbano R, D’Angelo V, Copetti M, Coco M, Balsamo T, la Torre A, Notarangelo A, Troiano M, Parisi S. Regulation of KEAP1 expression by promoter methylation in malignant gliomas and association with patient’s outcome. Epigenetics. 2011;6:317–25.
Wang R, An J, Ji F, Jiao H, Sun H, Zhou D. Hypermethylation of the Keap1 gene in human lung cancer cell lines and lung cancer tissues. Biochem Biophys Res Commun. 2008;373:151–4.
Muscarella LA, Parrella P, D’Alessandro V, la Torre A, Barbano R, Fontana A, Tancredi A, Guarnieri V, Balsamo T, Coco M. Frequent epigenetics inactivation of KEAP1 gene in non-small cell lung cancer. Epigenetics. 2011;6:710–9.
Hanada N, Takahata T, Zhou Q, Ye X, Sun R, Itoh J, Ishiguro A, Kijima H, Mimura J, Itoh K. Methylation of the KEAP1 gene promoter region in human colorectal cancer. BMC Cancer. 2012;12:66.
Soozangar N, Sadeghi MR, Jeddi F, Samadi N, Hejazi MS, Azad M, Shirmohamadi M, Somi MH. Decreased expression of the Keap1 gene and its clinicopathological significance in gastric cancer: correlation with promoter DNA methylation. Gazz Med Ital Arch Sci Med. 2019;178:292–300.
Barbano R, Muscarella LA, Pasculli B, Valori VM, Fontana A, Coco M, la Torre A, Balsamo T, Poeta ML, Marangi GF. Aberrant Keap1 methylation in breast cancer and association with clinicopathological features. Epigenetics. 2013;8:105–12.
Jeddi F, Alipour S, Najafzadeh N, Dadashpour M, Pouremamali F, Sadeghi MR, Samadi N, Soozangar N, Khamaneh AM. Reduced levels of miR–28 and miR–200a act as predictor biomarkers of aggressive clinicopathological characteristics in gastric cancer patients. Galen Med J. 2019;8:1–9.
Eades G, Yang M, Yao Y, Zhang Y, Zhou Q. miR-200a regulates Nrf2 activation by targeting Keap1 mRNA in breast cancer cells. J Biol Chem. 2011;286:40725–33.
Van Jaarsveld M, Helleman J, Boersma A, Van Kuijk P, Van Ijcken W, Despierre E, Vergote I, Mathijssen R, Berns E, Verweij J. miR-141 regulates KEAP1 and modulates cisplatin sensitivity in ovarian cancer cells. Oncogene. 2013;32:4284–93.
Kabaria S, Choi DC, Chaudhuri AD, Jain MR, Li H, Junn E. MicroRNA-7 activates Nrf2 pathway by targeting Keap1 expression. Free Radical Biol Med. 2015;89:548–56.
Chen W, Sun Z, Wang XJ, Jiang T, Huang Z, Fang D, Zhang DD. Direct interaction between Nrf2 and p21(Cip1/WAF1) upregulates the Nrf2-mediated antioxidant response. Mol Cell. 2009;34:663–73.
O’Reilly MA. Redox activation of p21Cip1/WAF1/Sdi1: a multifunctional regulator of cell survival and death. Antioxid Redox Signal. 2005;7:108–18.
Komatsu M, Kurokawa H, Waguri S, Taguchi K, Kobayashi A, Ichimura Y, Sou YS, Ueno I, Sakamoto A, Tong KI, et al. The selective autophagy substrate p62 activates the stress responsive transcription factor Nrf2 through inactivation of Keap1. Nat Cell Biol. 2010;12:213–23.
Lau A, Wang XJ, Zhao F, Villeneuve NF, Wu T, Jiang T, Sun Z, White E, Zhang DD. A noncanonical mechanism of Nrf2 activation by autophagy deficiency: direct interaction between Keap1 and p62. Mol Cell Biol. 2010;30:3275–85.
Jain A, Lamark T, Sjottem E, Larsen KB, Awuh JA, Overvatn A, McMahon M, Hayes JD, Johansen T. p62/SQSTM1 is a target gene for transcription factor NRF2 and creates a positive feedback loop by inducing antioxidant response element-driven gene transcription. J Biol Chem. 2010;285:22576–91.
Hancock R, Bertrand HC, Tsujita T, Naz S, El-Bakry A, Laoruchupong J, Hayes JD, Wells G. Peptide inhibitors of the Keap1–Nrf2 protein–protein interaction. Free Radical Biol Med. 2012;52:444–51.
Bae SH, Sung SH, Oh SY, Lim JM, Lee SK, Park YN, Lee HE, Kang D, Rhee SG. Sestrins activate Nrf2 by promoting p62-dependent autophagic degradation of Keap1 and prevent oxidative liver damage. Cell Metab. 2013;17:73–84.
Inami Y, Waguri S, Sakamoto A, Kouno T, Nakada K, Hino O, Watanabe S, Ando J, Iwadate M, Yamamoto M. Persistent activation of Nrf2 through p62 in hepatocellular carcinoma cells. J Cell Biol. 2011;193:275.
Inoue D, Suzuki T, Mitsuishi Y, Miki Y, Suzuki S, Sugawara S, Watanabe M, Sakurada A, Endo C, Uruno A, et al. Accumulation of p62/SQSTM1 is associated with poor prognosis in patients with lung adenocarcinoma. Cancer Sci. 2012;103:760–6.
Schläfli AM, Adams O, Galván JA, Gugger M, Savic S, Bubendorf L, Schmid RA, Becker K-F, Tschan MP, Langer R. Prognostic value of the autophagy markers LC3 and p62/SQSTM1 in early-stage non-small cell lung cancer. Oncotarget. 2016;7:39544.
Mohamed A, Ayman A, Deniece J, Wang T, Kovach C, Siddiqui MT, Cohen C. P62/Ubiquitin IHC expression correlated with clinicopathologic parameters and outcome in gastrointestinal carcinomas. Front Oncol. 2015;5:70.
Masuda GO, Yashiro M, Kitayama K, Miki Y, Kasashima H, Kinoshita H, Morisaki T, Fukuoka T, Hasegawa T, Sakurai K, et al. Clinicopathological correlations of autophagy-related proteins LC3, Beclin 1 and p62 in gastric cancer. Anticancer Res. 2016;36:129–36.
Luo RZ, Yuan ZY, Li M, Xi SY, Fu J, He J. Accumulation of p62 is associated with poor prognosis in patients with triple-negative breast cancer. Onco Targets Ther. 2013;6:883–8.
Choi J, Jung W, Koo JS. Expression of autophagy-related markers beclin-1, light chain 3A, light chain 3B and p62 according to the molecular subtype of breast cancer. Histopathology. 2013;62:275–86.
Park JM, Huang S, Wu TT, Foster NR, Sinicrope FA. Prognostic impact of Beclin 1, p62/sequestosome 1 and LC3 protein expression in colon carcinomas from patients receiving 5-fluorouracil as adjuvant chemotherapy. Cancer Biol Ther. 2013;14:100–7.
DeNicola GM, Karreth FA, Humpton TJ, Gopinathan A, Wei C, Frese K, Mangal D, Kenneth HY, Yeo CJ, Calhoun ES. Oncogene-induced Nrf2 transcription promotes ROS detoxification and tumorigenesis. Nature. 2011;475:106–9.
Tao S, Wang S, Moghaddam SJ, Ooi A, Chapman E, Wong PK, Zhang DD. Oncogenic KRAS confers chemoresistance by upregulating NRF2. Cancer Res. 2014;74:7430–41.
Kong B, Qia C, Erkan M, Kleeff J, Michalski CW. Overview on how oncogenic Kras promotes pancreatic carcinogenesis by inducing low intracellular ROS levels. Front Physiol. 2013;4:246.
Milkovic L, Zarkovic N, Saso L. Controversy about pharmacological modulation of Nrf2 for cancer therapy. Redox Biol. 2017;12:727–32.
Robledinos-Antón N, Fernández-Ginés R, Manda G, Cuadrado A. Activators and inhibitors of NRF2: a review of their potential for clinical development. Oxid Med Cell Longev. 2019;2019:1–20.
Sova M, Saso L. Design and development of Nrf2 modulators for cancer chemoprevention and therapy: a review. Drug Des Dev Ther. 2018;12:3181.
Magesh S, Chen Y, Hu L. Small molecule modulators of Keap1-Nrf2-ARE pathway as potential preventive and therapeutic agents. Med Res Rev. 2012;32:687–726.
Houghton CA, Fassett RG, Coombes JS. Sulforaphane: translational research from laboratory bench to clinic. Nutr Rev. 2013;71:709–26.
Juge N, Mithen R, Traka M. Molecular basis for chemoprevention by sulforaphane: a comprehensive review. Cell Mol Life Sci. 2007;64:1105.
Wang X, de Rivero Vaccari JP, Wang H, Diaz P, German R, Marcillo AE, Keane RW. Activation of the nuclear factor E2-related factor 2/antioxidant response element pathway is neuroprotective after spinal cord injury. J Neurotrauma. 2012;29:936–45.
Kensler TW, Egner PA, Agyeman AS, Visvanathan K, Groopman JD, Chen J-G, Chen T-Y, Fahey JW, Talalay P. Keap1–nrf2 signaling: a target for cancer prevention by sulforaphane. Nat Prod Cancer Prev Ther. 2012. https://doi.org/10.1007/128_2012_339.
Higdon JV, Delage B, Williams DE, Dashwood RH. Cruciferous vegetables and human cancer risk: epidemiologic evidence and mechanistic basis. Pharmacol Res. 2007;55:224–36.
Zhang Y, Tang L. Discovery and development of sulforaphane as a cancer chemopreventive phytochemical. Acta Pharmacol Sin. 2007;28:1343–54.
Conaway CC, Wang C-X, Pittman B, Yang Y-M, Schwartz JE, Tian D, McIntee EJ, Hecht SS, Chung F-L. Phenethyl isothiocyanate and sulforaphane and their <em>N</em>-acetylcysteine conjugates inhibit malignant progression of lung adenomas induced by tobacco carcinogens in A/J mice. Cancer Res. 2005;65:8548–57.
Paul N, McMahon M, Yamamoto M, Hayes JD. Identification of a novel Nrf2-regulated antioxidant response element (ARE) in the mouse NAD (P) H: quinone oxidoreductase 1 gene: reassessment of the ARE consensus sequence. Biochem J. 2003;374:337–48.
Hu C, Eggler AL, Mesecar AD, van Breemen RB. Modification of keap1 cysteine residues by sulforaphane. Chem Res Toxicol. 2011;24:515–21.
Kobayashi M, Li L, Iwamoto N, Nakajima-Takagi Y, Kaneko H, Nakayama Y, Eguchi M, Wada Y, Kumagai Y, Yamamoto M. The antioxidant defense system Keap1-Nrf2 comprises a multiple sensing mechanism for responding to a wide range of chemical compounds. Mol Cell Biol. 2009;29:493–502.
Zhang DD, Hannink M. Distinct cysteine residues in Keap1 are required for Keap1-dependent ubiquitination of Nrf2 and for stabilization of Nrf2 by chemopreventive agents and oxidative stress. Mol Cell Biol. 2003;23:8137–51.
Russo M, Spagnuolo C, Russo GL, Skalicka-Woźniak K, Daglia M, Sobarzo-Sánchez E, Nabavi SF, Nabavi SM. Nrf2 targeting by sulforaphane: a potential therapy for cancer treatment. Crit Rev Food Sci Nutr. 2018;58:1391–405.
Hayes JD, McMahon M, Chowdhry S, Dinkova-Kostova AT. Cancer chemoprevention mechanisms mediated through the Keap1-Nrf2 pathway. Antioxid Redox Signal. 2010;13:1713–48.
Kensler TW, Egner PA, Agyeman AS, Visvanathan K, Groopman JD, Chen JG, Chen TY, Fahey JW, Talalay P. Keap1-nrf2 signaling: a target for cancer prevention by sulforaphane. Top Curr Chem. 2013;329:163–77.
Priya DKD, Gayathri R, Sakthisekaran D. Role of sulforaphane in the anti-initiating mechanism of lung carcinogenesis in vivo by modulating the metabolic activation and detoxification of benzo(a)pyrene. Biomed Pharmacother. 2011;65:9–16.
Thimmulappa RK, Mai KH, Srisuma S, Kensler TW, Yamamoto M, Biswal S. Identification of Nrf2-regulated genes induced by the chemopreventive agent sulforaphane by oligonucleotide microarray. Cancer Res. 2002;62:5196–203.
Zhang C, Su ZY, Khor TO, Shu L, Kong AN. Sulforaphane enhances Nrf2 expression in prostate cancer TRAMP C1 cells through epigenetic regulation. Biochem Pharmacol. 2013;85:1398–404.
Su ZY, Zhang C, Lee JH, Shu L, Wu TY, Khor TO, Conney AH, Lu YP, Kong AN. Requirement and epigenetics reprogramming of Nrf2 in suppression of tumor promoter TPA-induced mouse skin cell transformation by sulforaphane. Cancer Prev Res (Phila). 2014;7:319–29.
Yu Z, Shao W, Chiang Y, Foltz W, Zhang Z, Ling W, Fantus IG, Jin T. Oltipraz upregulates the nuclear factor (erythroid-derived 2)-like 2 [corrected](NRF2) antioxidant system and prevents insulin resistance and obesity induced by a high-fat diet in C57BL/6J mice. Diabetologia. 2011;54:922–34.
Weerachayaphorn J, Luo Y, Mennone A, Soroka CJ, Harry K, Boyer JL. Deleterious effect of oltipraz on extrahepatic cholestasis in bile duct-ligated mice. J Hepatol. 2014;60:160–6.
Eba S, Hoshikawa Y, Moriguchi T, Mitsuishi Y, Satoh H, Ishida K, Watanabe T, Shimizu T, Shimokawa H, Okada Y, et al. The nuclear factor erythroid 2-related factor 2 activator oltipraz attenuates chronic hypoxia-induced cardiopulmonary alterations in mice. Am J Respir Cell Mol Biol. 2013;49:324–33.
Yu Z, Shao W, Chiang Y, Foltz W, Zhang Z, Ling W, Fantus IG, Jin T. Oltipraz upregulates the nuclear respiratory factor 2 alpha subunit (NRF2) antioxidant system and prevents insulin resistance and obesity induced by a high-fat diet in C57BL/6J mice. Diabetologia. 2011;54:922–34.
Piton A, Rauch C, Langouet S, Guillouzo A, Morel F. Involvement of pregnane X receptor in the regulation of CYP2B6 gene expression by oltipraz in human hepatocytes. Toxicol In Vitro. 2010;24:452–9.
Jaiswal AK. Regulation of antioxidant response element-dependent induction of detoxifying enzyme synthesis. Methods Enzymol. 2004;378:221–38.
Merrell MD, Jackson JP, Augustine LM, Fisher CD, Slitt AL, Maher JM, Huang W, Moore DD, Zhang Y, Klaassen CD. The Nrf2 activator oltipraz also activates the constitutive androstane receptor. Drug Metab Dispos Biol Fate Chem. 2008;36:1716.
Roebuck BD, Curphey TJ, Li Y, Baumgartner KJ, Bodreddigari S, Yan J, Gange SJ, Kensler TW, Sutter TR. Evaluation of the cancer chemopreventive potency of dithiolethione analogs of oltipraz. Carcinogenesis. 2003;24:1919–28.
Sharma S, Gao P, Steele VE. The chemopreventive efficacy of inhaled oltipraz particulates in the B[a]P-induced A/J mouse lung adenoma model. Carcinogenesis. 2006;27:1721–7.
Choi SH, Kim YM, Lee JM, Kim SG. Antioxidant and mitochondrial protective effects of oxidized metabolites of oltipraz. Expert Opin Drug Metab Toxicol. 2010;6:213–24.
Jia Z, Zhu H, Trush MA, Misra HP, Li Y. Generation of superoxide from reaction of 3H–1,2-dithiole-3-thione with thiols: implications for dithiolethione chemoprotection. Mol Cell Biochem. 2008;307:185–91.
Wakabayashi N, Dinkova-Kostova AT, Holtzclaw WD, Kang MI, Kobayashi A, Yamamoto M, Kensler TW, Talalay P. Protection against electrophile and oxidant stress by induction of the phase 2 response: fate of cysteines of the Keap1 sensor modified by inducers. Proc Natl Acad Sci USA. 2004;101:2040–5.
Iida K, Itoh K, Kumagai Y, Oyasu R, Hattori K, Kawai K, Shimazui T, Akaza H, Yamamoto M. Nrf2 is essential for the chemopreventive efficacy of oltipraz against urinary bladder carcinogenesis. Cancer Res. 2004;64:6424–31.
Sun W, Liu X, Zhang H, Song Y, Li T, Liu X, Liu Y, Guo L, Wang F, Yang T. Epigallocatechin gallate upregulates NRF2 to prevent diabetic nephropathy via disabling KEAP1. Free Radical Biol Med. 2017;108:840–57.
Lee S-J, Lee I-S, Mar W. Inhibition of inducible nitric oxide synthase and cyclooxygenase-2 activity by 1, 2, 3, 4, 6-penta-O-galloyl-β-D-glucose in murine macrophage cells. Arch Pharmacal Res. 2003;26:832–9.
Stuart EC, Scandlyn MJ, Rosengren RJ. Role of epigallocatechin gallate (EGCG) in the treatment of breast and prostate cancer. Life Sci. 2006;79:2329–36.
Kuroiwa Y, Ishii Y, Umemura T, Kanki K, Mitsumori K, Nishikawa A, Nakazawa H, Hirose M. Combined treatment with green tea catechins and sodium nitrite selectively promotes rat forestomach carcinogenesis after initiation with N-methyl-N′-nitro-N-nitrosoguanidine. Cancer Sci. 2007;98:949–57.
Fujiki H, Watanabe T, Sueoka E, Rawangkan A, Suganuma M. Cancer prevention with green tea and its principal constituent, EGCG: from early investigations to current focus on human cancer stem cells. Mol Cells. 2018;41:73.
Katiyar SK. Green tea prevents non-melanoma skin cancer by enhancing DNA repair. Arch Biochem Biophys. 2011;508:152–8.
Zan L, Chen Q, Zhang L, Li X. Epigallocatechin gallate (EGCG) suppresses growth and tumorigenicity in breast cancer cells by downregulation of miR-25. Bioengineered. 2019;10:374–82.
Kumar N, Shibata D, Helm J, Coppola D, Malafa M. Green tea polyphenols in the prevention of colon cancer. Front Biosci. 2007;12:309.
Khan N, Mukhtar H. Multitargeted therapy of cancer by green tea polyphenols. Cancer Lett. 2008;269:269–80.
Hastak K, Gupta S, Ahmad N, Agarwal MK, Agarwal ML, Mukhtar H. Role of p53 and NF-κB in epigallocatechin-3-gallate-induced apoptosis of LNCaP cells. Oncogene. 2003;22:4851.
Bhattacharya S, Ray RM, Johnson LR. STAT3-mediated transcription of Bcl-2, Mcl-1 and c-IAP2 prevents apoptosis in polyamine-depleted cells. Biochem J. 2005;392:335–44.
Na H-K, Surh Y-J. Modulation of Nrf2-mediated antioxidant and detoxifying enzyme induction by the green tea polyphenol EGCG. Food Chem Toxicol. 2008;46:1271–8.
Shin J-H, Jeon H-J, Park J, Chang M-S. Epigallocatechin-3-gallate prevents oxidative stress-induced cellular senescence in human mesenchymal stem cells via Nrf2. Int J Mol Med. 2016;38:1075–82.
Dhakshinamoorthy S, Jain AK, Bloom DA, Jaiswal AK. Bach1 competes with Nrf2 leading to negative regulation of the antioxidant response element (ARE)-mediated NAD(P)H: quinone oxidoreductase 1 gene expression and induction in response to antioxidants. J Biol Chem. 2005;280:16891–900.
Kweon M-H, Adhami VM, Lee J-S, Mukhtar H. Constitutive overexpression of Nrf2-dependent heme oxygenase-1 in A549 cells contributes to resistance to apoptosis induced by epigallocatechin 3-gallate. J Biol Chem. 2006;281:33761–72.
Keum Y-S, Choi BY. Molecular and chemical regulation of the Keap1-Nrf2 signaling pathway. Molecules. 2014;19:10074–89.
Gold R, Linker RA, Stangel M. Fumaric acid and its esters: an emerging treatment for multiple sclerosis with antioxidative mechanism of action. Clin Immunol. 2012;142:44–8.
Scannevin RH, Chollate S, Jung M-Y, Shackett M, Patel H, Bista P, Zeng W, Ryan S, Yamamoto M, Lukashev M, Rhodes KJ. Fumarates promote cytoprotection of central nervous system cells against oxidative stress via the nuclear factor (Erythroid-Derived 2)-like 2 pathway. J Pharmacol Exp Ther. 2012;341:274–84.
Peng H, Guerau-de-Arellano M, Mehta VB, Yang Y, Huss DJ, Papenfuss TL, Lovett-Racke AE, Racke MK. Dimethyl fumarate inhibits dendritic cell maturation via nuclear factor κB (NF-κB) and extracellular signal-regulated kinase 1 and 2 (ERK1/2) and mitogen stress-activated kinase 1 (MSK1) signaling. J Biol Chem. 2012;287:28017–26.
Schmidt MM, Dringen R. Fumaric acid diesters deprive cultured primary astrocytes rapidly of glutathione. Neurochem Int. 2010;57:460–7.
Dibbert S, Clement B, Skak-Nielsen T, Mrowietz U, Rostami-Yazdi M. Detection of fumarate–glutathione adducts in the portal vein blood of rats: evidence for rapid dimethylfumarate metabolism. Arch Dermatol Res. 2013;305:447–51.
Ahuja M, Kaidery NA, Yang L, Calingasan N, Smirnova N, Gaisin A, Gaisina IN, Gazaryan I, Hushpulian DM, Kaddour-Djebbar I. Distinct Nrf2 signaling mechanisms of fumaric acid esters and their role in neuroprotection against 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine-induced experimental Parkinson’s-like disease. J Neurosci. 2016;36:6332–51.
Liu X, Zhou W, Zhang X, Lu P, Du Q, Tao L, Ding Y, Wang Y, Hu R. Dimethyl fumarate ameliorates dextran sulfate sodium-induced murine experimental colitis by activating Nrf2 and suppressing NLRP3 inflammasome activation. Biochem Pharmacol. 2016;112:37–49.
Lin-Holderer J, Li L, Gruneberg D, Marti HH, Kunze R. Fumaric acid esters promote neuronal survival upon ischemic stress through activation of the Nrf2 but not HIF-1 signaling pathway. Neuropharmacology. 2016;105:228–40.
Albrecht P, Bouchachia I, Goebels N, Henke N, Hofstetter HH, Issberner A, Kovacs Z, Lewerenz J, Lisak D, Maher P. Effects of dimethyl fumarate on neuroprotection and immunomodulation. J Neuroinflamm. 2012;9:163.
Boivin A, Hanot M, Malesys C, Maalouf M, Rousson R, Rodriguez-Lafrasse C, Ardail D. Transient alteration of cellular redox buffering before irradiation triggers apoptosis in head and neck carcinoma stem and non-stem cells. PLoS ONE. 2011;6: e14558.
García-Caballero M, Marí-Beffa M, Medina MA, Quesada AR. Dimethylfumarate inhibits angiogenesis in vitro and in vivo: a possible role for its antipsoriatic effect? J Investig Dermatol. 2011;131:1347–55.
To C, Ringelberg CS, Royce DB, Williams CR, Risingsong R, Sporn MB, Liby KT. Dimethyl fumarate and the oleanane triterpenoids, CDDO-imidazolide and CDDO-methyl ester, both activate the Nrf2 pathway but have opposite effects in the A/J model of lung carcinogenesis. Carcinogenesis. 2015;36:769–81.
Loewe R, Valero T, Kremling S, Pratscher B, Kunstfeld R, Pehamberger H, Petzelbauer P. Dimethylfumarate impairs melanoma growth and metastasis. Cancer Res. 2006;66:11888–96.
Hwang JS, Lee YY, Lee D-H, Kwon KH. DATS sensitizes glioma cells to TRAIL-mediated apoptosis by up-regulation of death receptor 5 via ROS. Food Chem Toxicol. 2017;106:514–21.
Huang Y, Li W, Su Z-Y, Kong A-NT. The complexity of the Nrf2 pathway: beyond the antioxidant response. J Nutr Biochem. 2015;26:1401–13.
Kim S, Lee H-G, Park S-A, Kundu JK, Keum Y-S, Cha Y-N, Na H-K, Surh Y-J. Keap1 cysteine 288 as a potential target for diallyl trisulfide-induced Nrf2 activation. PLoS ONE. 2014;9: e85984.
Tsai C-Y, Wang C-C, Lai T-Y, Tsu H-N, Wang C-H, Liang H-Y, Kuo W-W. Antioxidant effects of diallyl trisulfide on high glucose-induced apoptosis are mediated by the PI3K/Akt-dependent activation of Nrf2 in cardiomyocytes. Int J Cardiol. 2013;168:1286–97.
Fisher CD, Augustine LM, Maher JM, Nelson DM, Slitt AL, Klaassen CD, Lehman-McKeeman LD, Cherrington NJ. Induction of drug-metabolizing enzymes by garlic and allyl sulfide compounds via activation of constitutive androstane receptor and nuclear factor E2-related factor 2. Drug Metab Dispos. 2007;35:995–1000.
Zhou N, Liu C, Lv S, Sun D, Qiao Q, Zhang R, Liu Y, Xiao J, Sun G. Degradation prediction model and stem cell growth of gelatin-PEG composite hydrogel. J Biomed Mater Res A. 2016;104:3149–56.
Serati-Nouri H, Rasoulpoor S, Pourpirali R, Sadeghi-Soureh S, Esmaeilizadeh N, Dadashpour M, Roshangar L, Zarghami N. In vitro expansion of human adipose-derived stem cells with delayed senescence through dual stage release of curcumin from mesoporous silica nanoparticles/electrospun nanofibers. Life Sci. 2021;285: 119947.
Lotfi-Attari J, Pilehvar-Soltanahmadi Y, Dadashpour M, Alipour S, Farajzadeh R, Javidfar S, Zarghami N. Co-delivery of curcumin and chrysin by polymeric nanoparticles inhibit synergistically growth and hTERT gene expression in human colorectal cancer cells. Nutr Cancer. 2017;69:1290–9.
Rasouli S, Montazeri M, Mashayekhi S, Sadeghi-Soureh S, Dadashpour M, Mousazadeh H, Nobakht A, Zarghami N, Pilehvar-Soltanahmadi Y. Synergistic anticancer effects of electrospun nanofiber-mediated codelivery of curcumin and chrysin: possible application in prevention of breast cancer local recurrence. J Drug Deliv Sci Technol. 2020;55: 101402.
Farajzadeh R, Pilehvar-Soltanahmadi Y, Dadashpour M, Javidfar S, Lotfi-Attari J, Sadeghzadeh H, Shafiei-Irannejad V, Zarghami N. Nano-encapsulated metformin-curcumin in PLGA/PEG inhibits synergistically growth and hTERT gene expression in human breast cancer cells. Artif Cells Nanomed Biotechnol. 2018;46:917–25.
Khazei K, Jamali M, Sarhadi S, Dadashpour M, Shokrollahzade S, Zarghami N. Transcriptome profiling of curcumin-treated T47D human breast cancer cells by a system-based approach. Gene Rep. 2022;27:101556.
Balamurugan A, Akhov L, Selvaraj G, Pugazhenthi S. Induction of antioxidant enzymes by curcumin and its analogues in human islets: implications in transplantation. Pancreas. 2009;38:454–60.
Wu J, Li Q, Wang X, Yu S, Li L, Wu X, Chen Y, Zhao J, Zhao Y. Neuroprotection by curcumin in ischemic brain injury involves the Akt/Nrf2 pathway. PLoS ONE. 2013;8: e59843.
Aggarwal BB, Sung B. Pharmacological basis for the role of curcumin in chronic diseases: an age-old spice with modern targets. Trends Pharmacol Sci. 2009;30:85–94.
Khor TO, Huang Y, Wu T-Y, Shu L, Lee J. Kong A-NT: Pharmacodynamics of curcumin as DNA hypomethylation agent in restoring the expression of Nrf2 via promoter CpGs demethylation. Biochem Pharmacol. 2011;82:1073–8.
Hong F, Freeman ML, Liebler DC. Identification of sensor cysteines in human Keap1 modified by the cancer chemopreventive agent sulforaphane. Chem Res Toxicol. 2005;18:1917–26.
Li M, Zhang Z, Hill DL, Wang H, Zhang R. Curcumin, a dietary component, has anticancer, chemosensitization, and radiosensitization effects by down-regulating the MDM2 oncogene through the PI3K/mTOR/ETS2 pathway. Cancer Res. 2007;67:1988–96.
Li L, Ahmed B, Mehta K, Kurzrock R. Liposomal curcumin with and without oxaliplatin: effects on cell growth, apoptosis, and angiogenesis in colorectal cancer. Mol Cancer Ther. 2007;6:1276–82.
Kunnumakkara AB, Diagaradjane P, Guha S, Deorukhkar A, Shentu S, Aggarwal BB, Krishnan S. Curcumin sensitizes human colorectal cancer xenografts in nude mice to γ-radiation by targeting nuclear factor-κB–regulated gene products. Clin Cancer Res. 2008;14:2128–36.
Lin YG, Kunnumakkara AB, Nair A, Merritt WM, Han LY, Armaiz-Pena GN, Kamat AA, Spannuth WA, Gershenson DM, Lutgendorf SK. Curcumin inhibits tumor growth and angiogenesis in ovarian carcinoma by targeting the nuclear factor-κB pathway. Clin Cancer Res. 2007;13:3423–30.
Strimpakos AS, Sharma RA. Curcumin: preventive and therapeutic properties in laboratory studies and clinical trials. Antioxid Redox Signal. 2008;10:511–46.
Li Y, Zhang J, Ma D, Zhang L, Si M, Yin H, Li J. Curcumin inhibits proliferation and invasion of osteosarcoma cells through inactivation of Notch-1 signaling. FEBS J. 2012;279:2247–59.
Wu S-H, Hang L-W, Yang J-S, Chen H-Y, Lin H-Y, Chiang J-H, Lu C-C, Yang J-L, Lai T-Y, Ko Y-C. Curcumin induces apoptosis in human non-small cell lung cancer NCI-H460 cells through ER stress and caspase cascade-and mitochondria-dependent pathways. Anticancer Res. 2010;30:2125–33.
Cao M, Onyango EO, Williams CR, Royce DB, Gribble GW, Sporn MB, Liby KT. Novel synthetic pyridyl analogues of CDDO-Imidazolide are useful new tools in cancer prevention. Pharmacol Res. 2015;100:135–47.
Yates MS, Tran QT, Dolan PM, Osburn WO, Shin S, McCulloch CC, Silkworth JB, Taguchi K, Yamamoto M, Williams CR. Genetic versus chemoprotective activation of Nrf2 signaling: overlapping yet distinct gene expression profiles between Keap1 knockout and triterpenoid-treated mice. Carcinogenesis. 2009;30:1024–31.
Liby KT, Sporn MB. Synthetic oleanane triterpenoids: multifunctional drugs with a broad range of applications for prevention and treatment of chronic disease. Pharmacol Rev. 2012;64:972–1003.
Yore MM, Kettenbach AN, Sporn MB, Gerber SA, Liby KT. Proteomic analysis shows synthetic oleanane triterpenoid binds to mTOR. PLoS ONE. 2011;6: e22862.
Walsh J, Jenkins RE, Wong M, Olayanju A, Powell H, Copple I, O’Neill PM, Goldring CE, Kitteringham NR, Park BK. Identification and quantification of the basal and inducible Nrf2-dependent proteomes in mouse liver: biochemical, pharmacological and toxicological implications. J Proteomics. 2014;108:171–87.
Wang Y-Y, Zhe H, Zhao R. Preclinical evidences toward the use of triterpenoid CDDO-Me for solid cancer prevention and treatment. Mol Cancer. 2014;13:30.
Shanmugam MK, Dai X, Kumar AP, Tan BK, Sethi G, Bishayee A. Oleanolic acid and its synthetic derivatives for the prevention and therapy of cancer: preclinical and clinical evidence. Cancer Lett. 2014;346:206–16.
Bishayee A, Ahmed S, Brankov N, Perloff M. Triterpenoids as potential agents for the chemoprevention and therapy of breast cancer. Front Biosci. 2011;16:980.
Hyer ML, Shi R, Krajewska M, Meyer C, Lebedeva IV, Fisher PB, Reed JC. Apoptotic activity and mechanism of 2-cyano-3, 12-dioxoolean-1, 9-dien-28-oic-acid and related synthetic triterpenoids in prostate cancer. Cancer Res. 2008;68:2927–33.
Xu X, Li M, Chen W, Yu H, Yang Y, Hang L. Apigenin attenuates oxidative injury in ARPE-19 cells thorough activation of Nrf2 pathway. Oxid Med Cell Longev. 2016.
Zhang W, Qiao H, Lv Y, Wang J, Chen X, Hou Y, Tan R, Li E. Apigenin inhibits enterovirus-71 infection by disrupting viral RNA association with trans-acting factors. PLoS ONE. 2014;9: e110429.
Patil RH, Babu R, Kumar MN, Kumar KK, Hegde SM, Nagesh R, Ramesh GT, Sharma SC. Anti-inflammatory effect of apigenin on LPS-induced pro-inflammatory mediators and AP-1 factors in human lung epithelial cells. Inflammation. 2016;39:138–47.
Andueza A, García-Garzón A, de Galarreta MR, Ansorena E, Iraburu MJ, López-Zabalza MJ, Martínez-Irujo JJ. Oxidation pathways underlying the pro-oxidant effects of apigenin. Free Radical Biol Med. 2015;87:169–80.
Tseng TH, Chien MH, Lin WL, Wen YC, Chow JM, Chen CK, Kuo TC, Lee WJ. Inhibition of MDA-MB-231 breast cancer cell proliferation and tumor growth by apigenin through induction of G2/M arrest and histone H3 acetylation-mediated p21WAF1/CIP1 expression. Environ Toxicol. 2017;32:434–44.
Zhu J, Wang H, Chen F, Fu J, Xu Y, Hou Y, Kou HH, Zhai C, Nelson MB, Zhang Q. An overview of chemical inhibitors of the Nrf2-ARE signaling pathway and their potential applications in cancer therapy. Free Radical Biol Med. 2016;99:544–56.
Patel D, Shukla S, Gupta S. Apigenin and cancer chemoprevention: progress, potential and promise (review). Int J Oncol. 2007;30:233–46.
Feng X, Yu W, Li X, Zhou F, Zhang W, Shen Q, Li J, Zhang C, Shen P. Apigenin, a modulator of PPARγ, attenuates HFD-induced NAFLD by regulating hepatocyte lipid metabolism and oxidative stress via Nrf2 activation. Biochem Pharmacol. 2017;136:136–49.
Paredes-Gonzalez X, Fuentes F, Su Z-Y. Kong A-NT: apigenin reactivates Nrf2 anti-oxidative stress signaling in mouse skin epidermal JB6 P + cells through epigenetics modifications. AAPS J. 2014;16:727–35.
Paredes-Gonzalez X, Fuentes F, Jeffery S, Saw CLL, Shu L, Su ZY, Kong ANT. Induction of NRF2-mediated gene expression by dietary phytochemical flavones apigenin and luteolin. Biopharm Drug Dispos. 2015;36:440–51.
Weldon CB, McKee A, Collins-Burow BM, Melnik LI, Scandurro AB, McLachlan JA, Burow ME, Beckman BS. PKC-mediated survival signaling in breast carcinoma cells: a role for MEK1-AP1 signaling. Int J Oncol. 2005;26:763–8.
Lee SJ, Yoon J-H, Song KS. Chrysin inhibited stem cell factor (SCF)/c-Kit complex-induced cell proliferation in human myeloid leukemia cells. Biochem Pharmacol. 2007;74:215–25.
Gao A-M, Ke Z-P, Wang J-N, Yang J-Y, Chen S-Y, Chen H. Apigenin sensitizes doxorubicin-resistant hepatocellular carcinoma BEL-7402/ADM cells to doxorubicin via inhibiting PI3K/Akt/Nrf2 pathway. Carcinogenesis. 2013;34:1806–14.
Tian Y, Song W, Li D, Cai L, Zhao Y. Resveratrol as a natural regulator of autophagy for prevention and treatment of cancer. Onco Targets Ther. 2019;12:8601.
Whitlock NC, Baek SJ. The anticancer effects of resveratrol: modulation of transcription factors. Nutr Cancer. 2012;64:493–502.
Saldanha JF, Leal VdO, Stenvinkel P, Carraro-Eduardo JC, Mafra D. Resveratrol: why is it a promising therapy for chronic kidney disease patients? Oxid Med Cell Longev. 2013;2013:1–6.
Jiménez-Osorio AS, Gonzalez-Reyes S, Pedraza-Chaverri J. Natural Nrf2 activators in diabetes. Clin Chim Acta. 2015;448:182–92.
Alavi M, Farkhondeh T, Aschner M, Samarghandian S. Resveratrol mediates its anti-cancer effects by Nrf2 signaling pathway activation. Cancer Cell Int. 2021;21:1–9.
Zhou X, Zhao Y, Wang J, Wang X, Chen C, Yin D, Zhao F, Yin J, Guo M, Zhang L. Resveratrol represses estrogen-induced mammary carcinogenesis through NRF2-UGT1A8-estrogen metabolic axis activation. Biochem Pharmacol. 2018;155:252–63.
Zhang Y, Wang G, Wang T, Cao W, Zhang L, Chen X. Nrf2–Keap1 pathway–mediated effects of resveratrol on oxidative stress and apoptosis in hydrogen peroxide–treated rheumatoid arthritis fibroblast-like synoviocytes. Ann N Y Acad Sci. 2019;1457:166–78.
Telkoparan-Akillilar P, Panieri E, Cevik D, Suzen S, Saso L. Therapeutic targeting of the NRF2 signaling pathway in cancer. Molecules. 2021;26:1417.
Hall IH, Lee KH, Eigebaly SA, Imakura Y, Sumida Y, Wu RY. Antitumor agents. XXXIV: mechanism of action of bruceoside A and brusatol on nucleic acid metabolism of P-388 lymphocytic leukemia cells. J Pharm Sci. 1979;68:883–7.
No JH, Kim Y-B, Song YS. Targeting nrf2 signaling to combat chemoresistance. J Cancer Prev. 2014;19:111.
Ren D, Villeneuve NF, Jiang T, Wu T, Lau A, Toppin HA, Zhang DD. Brusatol enhances the efficacy of chemotherapy by inhibiting the Nrf2-mediated defense mechanism. Proc Natl Acad Sci USA. 2011;108:1433–8.
Olayanju A, Copple IM, Bryan HK, Edge GT, Sison RL, Wong MW, Lai Z-Q, Lin Z-X, Dunn K, Sanderson CM. Brusatol provokes a rapid and transient inhibition of Nrf2 signaling and sensitizes mammalian cells to chemical toxicity—implications for therapeutic targeting of Nrf2. Free Radical Biol Med. 2015;78:202–12.
Tong Y-H, Zhang B, Fan Y, Lin N-M. Keap1–Nrf2 pathway: a promising target towards lung cancer prevention and therapeutics. Chronic Dis Transl Med. 2015;1:175–86.
Pandurangan AK, Esa NM. Luteolin, a bioflavonoid inhibits colorectal cancer through modulation of multiple signaling pathways: a review. Asian Pac J Cancer Prev. 2014;15:5501–8.
Chian S, Li Y-Y, Wang X-J, Tang X-W. Luteolin sensitizes two oxaliplatin-resistant colorectal cancer cell lines to chemotherapeutic drugs via inhibition of the Nrf2 pathway. Asian Pac J Cancer Prev APJCP. 2014;15:2911–6.
Yang D, Tan X, Lv Z, Liu B, Baiyun R, Lu J, Zhang Z. Regulation of Sirt1/Nrf2/TNF-α signaling pathway by luteolin is critical to attenuate acute mercuric chloride exposure induced hepatotoxicity. Sci Rep. 2016;6:37157.
Esmaeili MA. Combination of siRNA-directed gene silencing with epigallocatechin-3-gallate (EGCG) reverses drug resistance in human breast cancer cells. J Chem Biol. 2016;9:41–52.
Qin W, Guan D, Ma R, Yang R, Xing G, Shi H, Tang G, Li J, Lv H, Jiang Y. Effects of trigonelline inhibition of the Nrf2 transcription factor in vitro on Echinococcus granulosus. Acta Biochim Biophys Sin. 2017;49:696–705.
Shin D, Kim EH, Lee J, Roh J-L. Nrf2 inhibition reverses resistance to GPX4 inhibitor-induced ferroptosis in head and neck cancer. Free Radical Biol Med. 2018;129:454–62.
Pallauf K, Duckstein N, Hasler M, Klotz L-O, Rimbach G. Flavonoids as putative inducers of the transcription factors Nrf2, FoxO, and PPARγ. Oxid Med Cell Longev. 2017;2017:4397340–4397340.
Kim SR, Ha YM, Kim YM, Park EJ, Kim JW, Park SW, Kim HJ, Chung HT, Chang KC. Ascorbic acid reduces HMGB1 secretion in lipopolysaccharide-activated RAW 264.7 cells and improves survival rate in septic mice by activation of Nrf2/HO-1 signals. Biochem Pharmacol. 2015;95:279–89.
Turkseven S, Kruger A, Mingone CJ, Kaminski P, Inaba M, Rodella LF, Ikehara S, Wolin MS, Abraham NG. Antioxidant mechanism of heme oxygenase-1 involves an increase in superoxide dismutase and catalase in experimental diabetes. Am J Physiol Heart Circ Physiol. 2005;289:H701-707.
Vineetha RC, Binu P, Arathi P, Nair RH. L-ascorbic acid and α-tocopherol attenuate arsenic trioxide-induced toxicity in H9c2 cardiomyocytes by the activation of Nrf2 and Bcl2 transcription factors. Toxicol Mech Methods. 2018;28:353–60.
Bushue N, Wan Y-JY. Retinoid pathway and cancer therapeutics. Adv Drug Deliv Rev. 2010;62:1285–98.
Soprano DR, Qin P, Soprano KJ. Retinoic acid receptors and cancers. Annu Rev Nutr. 2004;24:201–21.
Mei D, Lv B, Chen B, Xiao S, Jiang J, Xie Y, Jiang L. All-trans retinoic acid suppresses malignant characteristics of CD133-positive thyroid cancer stem cells and induces apoptosis. PLoS ONE. 2017;12: e0182835.
Wang XJ, Hayes JD, Henderson CJ, Wolf CR. Identification of retinoic acid as an inhibitor of transcription factor Nrf2 through activation of retinoic acid receptor alpha. Proc Natl Acad Sci. 2007;104:19589–94.
Shaulian E, Karin M. AP-1 as a regulator of cell life and death. Nat Cell Biol. 2002;4:E131–6.
Khoshravan Azar L, Dadashpour M, Hashemi M, Zarghami N. Design and development of nanostructured Co delivery of artemisinin and chrysin for targeting hTERT gene expression in breast cancer cell line: possible clinical application in cancer treatment. Asian Pac J Cancer Prev. 2022;23:919–27.
Deldar Y, Pilehvar-Soltanahmadi Y, Dadashpour M, Montazer Saheb S, Rahmati-Yamchi M, Zarghami N. An in vitro examination of the antioxidant, cytoprotective and anti-inflammatory properties of chrysin-loaded nanofibrous mats for potential wound healing applications. Artif Cells Nanomed Biotechnol. 2018;46:706–16.
Firouzi-Amandi A, Dadashpour M, Nouri M, Zarghami N, Serati-Nouri H, Jafari-Gharabaghlou D, Karzar BH, Mellatyar H, Aghebati-Maleki L, Babaloo Z. Chrysin-nanoencapsulated PLGA-PEG for macrophage repolarization: Possible application in tissue regeneration. Biomed Pharmacother. 2018;105:773–80.
Maasomi ZJ, Soltanahmadi YP, Dadashpour M, Alipour S, Abolhasani S, Zarghami N. Synergistic anticancer effects of silibinin and chrysin in T47D breast cancer cells. Asian Pac J Cancer Prev APJCP. 2017;18:1283.
Li Y, Wang X. Chrysin attenuates high glucose-induced BMSC dysfunction via the activation of the PI3K/AKT/Nrf2 signaling pathway. Drug Des Dev Ther. 2022;16:165.
Kasala ER, Bodduluru LN, Madana RM, Gogoi R, Barua CC. Chemopreventive and therapeutic potential of chrysin in cancer: mechanistic perspectives. Toxicol Lett. 2015;233:214–25.
Talebi M, Talebi M, Farkhondeh T, Simal-Gandara J, Kopustinskiene DM, Bernatoniene J, Samarghandian S. Emerging cellular and molecular mechanisms underlying anticancer indications of chrysin. Cancer Cell Int. 2021;21:1–20.
Gao A-M, Ke Z-P, Shi F, Sun G-C, Chen H. Chrysin enhances sensitivity of BEL-7402/ADM cells to doxorubicin by suppressing PI3K/Akt/Nrf2 and ERK/Nrf2 pathway. Chem Biol Interact. 2013;206:100–8.
Zeng W, Yan Y, Zhang F, Zhang C, Liang W. Chrysin promotes osteogenic differentiation via ERK/MAPK activation. Protein Cell. 2013;4:539–47.
Suraweera LT, Rupasinghe H, Dellaire G, Xu Z. Regulation of Nrf2/ARE pathway by dietary flavonoids: a friend or foe for cancer management? Antioxidants. 2020;9:973.
Acknowledgements
We thank the current and former members of our laboratories and collaborators for their contributions to the publications cited in this review article. The research field in Keap1 and Nrf2 is rapidly growing, and we apologize for not being able to cite all the recent publications, due to space limitation.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
FP and AP were principle investigators of the study and led writing of this manuscript. MD provided advice for fieldwork, helped write drafts of the manuscript, and shared in figure development. NS and FJ conceived and planed the study, drafted the manuscript and supervised the project. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This research was supported by the Ardabil University of Medical Sciences (Grant No.: IR.ARUMS.REC.1399.162).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Pouremamali, F., Pouremamali, A., Dadashpour, M. et al. An update of Nrf2 activators and inhibitors in cancer prevention/promotion. Cell Commun Signal 20, 100 (2022). https://doi.org/10.1186/s12964-022-00906-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12964-022-00906-3