
Abstract
Background
Chronic inflammation and oxidant/antioxidant imbalance are two main pathological features associated with lipopolysaccharide (LPS)-induced acute lung injury (ALI). The following study investigated the protective role of hydrogen (H2), a gaseous molecule without known toxicity, in LPS-induced lung injury in mice and explored its potential molecular mechanisms.
Methods
Mice were randomly divided into three groups: H2 control group, LPS group, and LPS + H2 group. The mice were euthanized at the indicated time points, and the specimens were collected. The 72 h survival rates, cytokines contents, pathological changes, expression of Toll-like receptor 4 (TLR4), and oxidative stress indicators were analyzed. Moreover, under different culture conditions, RAW 264.7 mouse macrophages were used to investigate the potential molecular mechanisms of H2 in vitro. Cells were divided into the following groups: PBS group, LPS group, and LPS + H2 group. The cell viability, intracellular ROS, cytokines, and expression of TLR4 and nuclear factor kappa-B (NF-κB) were observed.
Results
Hydrogen inhalation increased the survival rate to 80%, reduced LPS-induced lung damage, and decreased inflammatory cytokine release in LPS mice. Besides, H2 showed remarked anti-oxidative activity to reduce the MDA and NO contents in the lung. In vitro data further indicated that H2 down-regulates the levels of ROS, NO, TNF-α, IL-6, and IL-1β in LPS-stimulated macrophages and inhibits the expression of TLR4 and the activation of nuclear factor kappa-B (NF-κB).
Conclusion
Hydrogen gas alleviates lipopolysaccharide-induced acute lung injury and inflammatory response most probably through the TLR4-NF-κB pathway.
Similar content being viewed by others
Introduction
Acute lung injury (ALI) is a severe disease syndrome with high morbidity and high mortality [1,2,3]. Its pathogenesis is complex and characterized by inflammatory processes, oxidative stress, apoptosis, and increased autophagy [4]. Treatment of ALI is based on both ventilatory and nonventilatory strategies. Yet, the efficacy of standard therapies such as prone positioning and administration of neuromuscular blocking agents, glucocorticoids and surfactants stem cells injection, and lung-protective mechanical ventilation is limited [5,6,7]. Thus, there is an urgent need to develop novel therapeutic strategies and agents for ALI.
Lipopolysaccharide (LPS) is an outer membrane of Gram-negative bacteria and a common inducing agent of ALI [8]. LPS activates the macrophages and inflammatory cells, which then release uncontrolled inflammatory cytokines [9]. Studies have suggested that LPS-activated Toll-like receptors 4 (TLR4) can generate inflammatory cytokines by activating the NF-κB signaling pathway [10,11,12,13]. These cytokines exert critical roles in ALI development by amplifying inflammatory responses [3]. Thus, targeted inhibition of TLR4 and downstream NF-κB signaling pathways, breaking the cascade of inflammation, may be a promising strategy for ALI.
Inflammatory lung diseases are characterized by inflammation development and oxidant/antioxidant imbalance. The balance between the production and elimination of ROS maintains the functional integrity of redox-sensitive signaling [14]. ROS-activated redox-sensitive transcription factors, such as NF-κB, which regulate the chromatin remodeling and gene expression of pro-inflammatory mediators, can enhance inflammatory responses and tissue injury [15,16,17]. In this study, we hypothesized that inhibition of the downstream NF-κB signaling pathway might be a useful strategy for scavenging ROS and treating ALI.
Hydrogen (H2) is a gaseous molecule without known toxicity, which reacts with hydroxyl radical to remove the reactive oxygen species (ROS). Recent studies have revealed that hydrogen is an important physiological regulator with antioxidant, anti-inflammatory, and anti-apoptotic effects on cells and organs [18,19,20,21]. Moreover, studies showed that hydrogen in the human body acts as a regulator of signal transduction like other gaseous signaling molecules and thus has been proposed as ‘the fourth signal gas molecule’ [22, 23]. However, whether H2 improves the specific anti-inflammatory mechanism of LPS-induced ALI is still unclear.
This study aimed to investigate the protective role of H2 in LPS-induced lung injury and explore its potential molecular mechanisms.
Material and methods
In vivo experimental design
Animals and experimental design
A total of 105 female C57BL/6 (6–8 weeks of age and weighing 19–22 g) mice were assessed in the present study. All the animals were housed in an environment with a temperature of 22 ± 1 ºC, relative humidity of 50 ± 1%, and a light/dark cycle of 12/12 h and were given water and food ad libitum. All animal studies (including the mice euthanasia procedure) were done in compliance with the regulations and guidelines of Tongji University School of Medicine in Shanghai institutional animal care and conducted according to the AAALAC and the IACUC guidelines (Shanghai) 2017–0005).
All mice were randomly divided into 3 groups (n = 35): 42% hydrogen gas inhalation group (H2 group), LPS-induced ALI group (LPS group, LPS derived from Escherichia coli (serotype O111:B4)), and ALI with 42% hydrogen inhalation group (LPS + H2 group).
Ten mice were randomly drawn from each group to determine the 72 h survival statistics, which were used to assess the protective effect of H2. The other 75 mice were used for collecting other samples. The two ALI groups were induced by intraperitoneal injection of 10 mg/kg LPS. Treatments with inhalation of 42% hydrogen gas for 72 h were administered after the injection of LPS or saline. Hydrogen was produced by a hydrogen–oxygen nebulizer (license No: AMS-H-03, Shanghai Asclepius Meditec Co., Ltd., Shanghai, China) that generates 3 L/min hydrogen gas by water electrolysis. As measured by gas chromatography, the gas generated consisted of 67% hydrogen and 33% oxygen. To keep the oxygen content at 21%, a certain amount of nitrogen was passed in. Thus, the hydrogen mixed gas contained 42% hydrogen, 21% oxygen, and 37% nitrogen (endotoxin-free, purity > 99.9%) in the air.
Cytokine measurements
To measure the cytokines in the serum of mice, blood samples were collected from the eyeball at 3, 6, 12, 24, 48, and 72 h after LPS injection. The serum was separated by centrifugation at 3000 g for 15 min (4℃) and then stored at -80℃. The corresponding enzyme-linked immunosorbent kit (CUSABIO BioTECH., Ltd., China) was used to detect the concentrations of IL-1β, TNF-α, IL-6, and IL-10. All samples were measured in triplicate.
Measurement of MDA and NO levels in lung tissues
Malondialdehyde (MDA) and nitric oxide (NO) are commonly used to represent local or systemic oxidative stress. The lung tissues were collected at 6, 12, and 24 h after LPS injection. Then, 10% of lung tissue homogenates were separated from the supernatant by 3000 g centrifugation for 20 min at 4 °C. Subsequently, MDA and NO content in the supernatants was measured using an MDA assay kit and NO assay kit (Jiancheng Bioengineering Institute, Nanjing, China).
Histological examinations
Lung tissue was collected at 6, 12, 24, and 48 h after LPS injection and fixed in 10% neutral buffered formalin for 24 h and then embedded in paraffin. All specimens were cut into 5 μm thick sections and stained with hematoxylin and eosin (HE). Samples were photographed and examined immediately by Leica DM Microscopes (DM 2500B, Germany, × 200).
TLR4 expressions in lung tissues
Twelve and 24 h after LPS injection, lung specimens were cut into 5 μm thick sections, then immersed in 3% H2O2 for 25 min and rinsed with PBS. They were blocked with 10% normal rabbit serum for 30 min at room temperature and stained with anti-TLR4 primary antibody (1:500; Abcam) overnight at 4 °C, followed by horseradish peroxidase or fluorescein isothiocyanate-conjugated goat anti-mouse immunoglobulin G antibody (1:5000; zhongshan bio.) for 50 min at room temperature. Sections were observed under a microscope (Eclipse Ti-SR; Nikon, Japan, × 200).
In vitro experimental design
Cell Culture and groups
The mouse RAW 264.7 macrophage cells were purchased from American Type Culture Collection [ATCC], USA. Cells were cultured in a humidified incubator containing 5% CO2 with Dulbecco’s modified Eagle’s medium (DMEM) medium (Gibco, Grand Island, NY) at 37 °C, containing 100-IU/mL penicillin G, 100-IU/ mL streptomycin, and 10% heat-inactivated fetal bovine serum.
The experiment included three groups: PBS group, LPS group, and LPS + H2 group. The PBS group was cultured with a normal DMEM medium and stimulated with PBS in a humidified atmosphere containing 5%CO2/95% air at 37ºC; the LPS group was cultured with a normal DMEM medium and stimulated with LPS in a humidified atmosphere containing 5%CO2/95% air at 37ºC; the LPS + H2 group was cultured with normal DMEM medium + 60% H2 and stimulated with LPS in a humidified atmosphere containing hydrogen mixed gas (60% hydrogen, 21% oxygen, 5% carbon dioxide, and 14% nitrogen (endotoxin-free, purity > 99.9%) in air. Hydrogen was produced by a hydrogen–oxygen nebulizer (license No: AMS-H-03, Shanghai Asclepius Meditec Co., Ltd., Shanghai, China).
Nitric oxide measurement
RAW 264.7 mouse macrophages (1 × 105cell/ml) were plated in 96-well plates. On day 2, the LPS + H2 group and LPS group were pretreated with H2 (60% H2 + normal cell medium) or vehicle (normal cell medium) for 24 h, respectively, and then all cells were incubated with LPS (200 ng/mL) for another 24 h. The concentration of nitric oxide (NO) in media was determined using the NO assay kit (Jiancheng Bioengineering Institute, Nanjing, China).
Cell viability assay
Cell viability was assessed by the Cell Counting Kit (Beyotime, China). In 96-well culture dishes, 1 × 105 macrophages in each well and cell were pretreated with 60% H2 for 24 h and then stimulated with LPS (200 ng/ml) or PBS for another 24 h. Then, a 20 μl CCK-8 solution was added to each well and incubated at 37 °C for another 4 h. The cell viability was measured by a microplate spectrophotometer (Thermo, USA) at OD450 nm.
Detection of intracellular ROS
Intracellular ROS production was monitored by a ROS assay kit (Solarbio, China). RAW 264.7 mouse macrophages (1 × 105cell/ml) were seeded into 6-well. Cells were pretreated with 60% H2 for 24 h and then stimulated with LPS (1 μg/ml) or PBS for another 24 h. Then, cells were exposed to a serum-free medium containing 10-μM 2', 7'-Dichlorodihydrofluorescein diacetate (DCFDA). After 20 min of incubation in darkness, the cells were washed three times with a blank DMEM medium. At last, cells were observed under the fluorescence microscope (OLYMPUS, IX71).
Real-time quantitative PCR
In 6-well culture dishes, 1 × 105 macrophages were placed in each well and pretreated with 60% H2 for 24 h and then stimulated with LPS (1 μg/ml) or PBS for another 24 h. Total RNA was extracted by Trizol reagent (Invitrogen, USA) and reverse-transcribed using Thermo Scientific RevertAid cDNA Synthesis Kit (Thermo, USA) to produce cDNA. The quantitative real-time PCR was performed with the ABI-7500 machine using SYBR Green PCR Kit (Thermo, USA).Glyceraldehydes-3-phosphate dehydrogenase (GAPDH) mRNA was used as an internal control. The quantitative PCR program used was as follows: predenaturation (94 °C, 10 min), denaturation (94 °C, 20 s), annealing (55 °C, 20 s), and extension (72 °C, 20 s), using primers specific for GAPDH, IL-6, IL‐1β, IL‐10, and TNF-α.
Each sample was conducted in triplicate, and the gene expression levels were calculated relative to the amount of GAPDH using the 2−ΔΔCT method. The primer sequences for the tested genes are listed in Table 1.
Western blot
All cells were collected and lysed in RIPA buffer (Beyotime, China), and the whole proteins, nucleoproteins, and cytoplasmic proteins were extracted as required. Protein concentrations were determined by BCA protein assay kit (KeyGEN BioTECH, China). Equal quantities of protein were separated on 12% SDS–PAGE, electrophoretically transferred to polyvinylidene fluoride membranes (Millipore, USA), and then blocked with 5% non-fat milk in TBST buffer for 2 h at room temperature. The membranes were then incubated with the corresponding antibodies overnight at 4 °C. The corresponding antibodies were: anti-IκB, anti-pIκB, anti-TLR4, and anti-NF-κB(Abcam, Cambridge, UK). Samples were then washed three times and incubated with the horseradish peroxidase-conjugated secondary anti-rabbit/mouse antibody for 1 h at room temperature. The proteins were visualized using an enhanced ECL detection kit (Dingguo changsheng biotechnology CO., Ltd., China) and scanned with a Clinx ChemiScope chemiluminescence imaging system (ChemiScope 5300 Pro). The relative optical densities of specific proteins were estimated utilizing a ChemiScope analysis program.
Statistical analysis
Data were reported as the mean ± SEM. All statistical analyses were performed using Prism 5.0 (GraphPad Software, USA). The nonparametric Mann–Whitney U test was employed to compare the value of all indicators between pairwise. A significance level of 0.05 was considered to be significant for all calculations. #p or *p represents p-value < 0.05. ##p or **p represents p-value < 0.01. ###p or ***p represents p-value < 0.001.
Result
Hydrogen inhalation improved the survival rate of ALI-mice
The 72 h survival rates were recorded after experiments and analyzed by Log-rank (Mantel-Cox) test. As shown in Fig. 1A, the survival of mice in the H2 group was 100%, 60% in the LPS group, and 80% in the H2 treatment group (all p < 0.05).

Effects of H2 on the survival rate of mice. Results are expressed as percent survival, n = 10. *P < 0.05 values of LPS mice compared with the LPS + H2 mice
Hydrogen inhalation reduced the oxidative stress of ALI-mice
MDA and NO concentration was an index of lipid peroxidation. The levels of oxidative product (NO and MDA) in lung tissues were measured at 6, 12, and 24 h after LPS administration. As shown in Fig. 2, NO and MDA were significantly decreased in mice treated with H2 (all p < 0.01).

Effects of H2 on MDA levels and NO content. H2 treatment significantly reduced the increased NO and MDA levels induced by LPS. (A) NO levels; (B) MDA levels. *P < 0.05 and **P < 0.01 values of LPS mice compared with the LPS + H2 mice
Hydrogen inhalation ameliorated inflammatory responses of ALI-mice
Cytokines that promote inflammation like TNF-α, IL-1β, IL-6, and adjusted inflammation like IL-10 were chosen to represent the systematic inflammation level. Figure 3A, B, and C represent TNF-α, IL-1β, and IL-6 levels in serum, respectively. The results showed the LPS induced TNF-α and IL-1β; contrary, these expressions were markedly reduced in the H2 treatment group. However, the level of IL-6 did not change after treatment. Furthermore, Fig. 4D showed that IL-10 level was significantly higher than the control group in a short time (< 24 h).

Effect of H2 on inflammatory cytokines in the serum of mice after LPS stimulation. (A) TNF-a; (B) IL-1β; (C) IL-6; (D) IL-10. *P < 0.05, **P < 0.01 and ***P < 0.001 values of LPS mice compared with the LPS + H2 mice

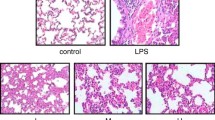
Effects of H2 treatment on histological changes in lung of LPS-induced ALI mice (× 200)
Hydrogen inhalation reduced the lung injury of ALI-mice
Histological observation using H&E stain is shown in Fig. 4. Alveolar collapse, alveolar septal thickening, and a inflammatory cell infiltration were obvious in the lung tissue of LPS mice (Marked with yellow, black and red arrows respectively). This histological examination showed that LPS induced oxidative stress and inflammations, which resulted in tissue injury, and was consistent with the lipid peroxidation and cytokines results. Contrary, H2 inhalation significantly prevented the histopathological changes caused by LPS.
Hydrogen inhalation reduced the expression of TLR4
Immunohistochemistry was performed to explore the effects of H2 on the expression of TLR4 in lung tissues. As shown in Fig. 5, H2-treated mice showed lower TLR4 expression compared to LPS mice.

The expression of TLR4 in mice lung tissues analyzed by immunohistochemical staining in the LPS and LPS + H2 groups (× 200)
Hydrogen reduced the release of the cytokine in LPS-stimulated macrophages
To assess the inhibitory role of H2 on cytokine production, we used the quantitative real-time PCR assay to detect the TNF-α, IL-6, IL-1β, and IL-10 mRNA expression in macrophages after LPS stimulation. Compared with the basal level without LPS stimulation, the contents of the four cytokines were significantly increased. At the same time, the TNF-α, IL-6, and IL-1β mRNA levels were all inhibited by H2 (Fig. 6). These findings indicate that H2 could neutralize the pro-inflammatory mediator effect of LPS, showing an anti-inflammatory effect.

Effect of H2 (pretreated with H2 for 24 h) on inflammatory cytokines. (A) TNF-a; (B) IL-1β; (C) IL-6; (D) IL-10. **P < 0.01values of LPS + H2 and LPS group compared with the PBS group. ##P < 0.01values of LPS + H2 group compared with the LPS group
Hydrogen increased cell viability and reduced the NO level in RAW 264.7 Cells
After LPS stimulation, cell proliferation was significantly reduced compared with the PBS and LPS + H2 groups (Fig. 7). Also, the NO content increased significantly (116.632 ± 7.729 μmol/L) compared to PBS (23.464 ± 8.615 μmol/L) and LPS + H2 groups (48.999 ± 8.661 μmol/L).

Cell viability and NO level of different groups (pretreated with H2 for 24 h). (A) Cell viability; (B) NO levels. **P < 0.01values of LPS + H2 and LPS group compared with the PBS group. ##P < 0.01values of LPS + H2 group compared with the LPS group
Hydrogen reduced the ROS production in LPS-stimulated RAW 264.7 cells
The RAW 264.7 + PBS group showed good cell status and low intracellular ROS levels. After LPS stimulation, the cell status deteriorated, and ROS levels increased significantly. When H2 treatment was performed before LPS stimulation, intracellular ROS levels were significantly reduced (Fig. 8).

The ROS level of RAW 264.7 + PBS group, RAW 264.7 + LPS group, and RAW 264.7 + LPS + H2 group (pretreated with H2 for 24 h)
Hydrogen reduced the TLR4 expression and suppressed NF-κB activation in RAW 264.7 cells caused by LPS
Compared with the control group, the expression level of IκB was significantly decreased after stimulation with LPS at 1 μg/ml, while the expression levels of phosphorylated IκB, TLR4, and NF-κB (cytoplasm and nucleus) were significantly increased (Fig. 9). After 60% H2 treatment, the expression level of IκB was significantly increased, while the phosphorylated IκB, TLR4, and NF-κB (cytoplasmic and nuclear) expression levels were significantly reduced compared to the LPS stimulation group.

H2 inhibits the TLR4 expression and NF-κB activation (pretreated with H2 for 24 h). Expression of GAPDH (Proteintech, USA) and Lamin B (Abcam, UK) were internal controls. The values presented are the means ± SEM of three independent experiments. ##p < 0.01 compared to PBS group; *p < 0.05 and **p < 0.01 compared to LPS group
Discussion
Acute lung injury (ALI) is a serious inflammatory disease [1,2,3]. LPS is a key mediator of organ dysfunction and death associated with severe gram-negative infections [8] and the most important pathogen causing ALI [24, 25]. In this study, we used LPS (i.p., 10 mg/kg) to establish the ALI model mice.
Recent studies have shown that hydrogen gas (H2) is an important physiological regulatory factor with antioxidant, anti-inflammatory, and anti-apoptotic properties [26]. In the present study, we investigated the protective effects of H2 treatment in LPS-induced ALI mice. We hypothesized that its protective effect might be related to its ability to ameliorate the extent of oxidative stress and prevent the release of pro-inflammatory molecules [14,15,16,17]. H2 inhalation can reduce lung injury caused by the ventilator, transplantation, hyperoxia, irradiation, and sepsis [27,28,29,30]. The histopathologic evaluation shows that the lung in the LPS group showed a thickened alveolar wall, edema and hemorrhage, less alveolar space, and inflammatory cell infiltration after LPS stimulation, which, however, were not observed in LPS mice treated with H2 inhalation. The H2 treatment ameliorates LPS-induced lung neutrophil infiltration and inflammation.
To verify the success of the ALI model, we observed the pathological and inflammatory changes at different time points after LPS injection in mice. The critical feature of ALI is the lung parenchyma injury and acute inflammatory process, including the release of inflammatory mediators such as TNF-α, IL-1β, and IL-6 [31]. Pro-inflammatory cytokines appear in the early stages of inflammation, indicating the severity of ALI in a certain sense [32]. In this study, the inflammatory factors gradually increased during the first 24 h after LPS administration compared to the H2. These results suggested that the ALI model was successfully established after intraperitoneal injections of LPS, with the lung injury being obvious at 24 h point. H2 markedly reduced the inflammation of ALI.
In vivo data further indicated that hydrogen gas inhalation protects mice against ALI lethality. It markedly reduces the inflammation and oxidative damages caused by ALI, improves the lung injury caused by LPS stimulation, and inhibits the TLR4 expression in lung tissues. In conclusion, these results show that molecular hydrogen alleviates LPS-induced ALI by reducing lung inflammation and oxidative damage, which may be associated with decreased NF-κB activity. All of the results are consistent with Sun et al. [33] and Du et al. [34], who confirmed that hydrogen could alleviate LPS-induced pulmonary lesions and pulmonary edema and reduce the degree of ALI by inhibiting the release of pro-inflammatory cytokines and oxidative stress. However, Sun’s study results further discussed the apoptosis-related results and the effect of the Nrf2 gene on the NF-KB pathway, while Du et al.explained the effect of the sirt1 gene on the NF-KB pathway.
LPS causes decrements in neutrophilic inflammation and pulmonary function via Toll-like receptor 4 to induce the expression of inflammatory cytokines and chemokines [35, 36]. The mechanism of ALI injury may be associated with bacterial endotoxin (LPS) activation that promotes the interaction of TLR4 and results in activation of NF-κB and release of TNF-α and IL-6 [37]. All of these factors have a pivotal part in inflammatory lung damage. Many studies have proved that inhibiting the activity of NF-κB in different models can alleviate the tissue damage and down-regulated release of cytokines [38, 39]. Therefore, inhibition of NF-κB activation may be an effective choice for protecting ALI.
H2 can directly activate the NF-κB signaling by inhibiting the phosphorylation of IκB-α [40]. In addition, H2 can inhibit the activation of the NF-κB signaling pathway by scavenging the oxygen radicals [41]. Hydrogen gas, a potent antioxidant with rapid gaseous diffusion, effectively reduces cytotoxic free radicals, such as reactive oxygen species (ROS), whilst being mild enough to cause interference with metabolic redox reactions or disrupt cell signaling. Our in vitro data further showed that the elevations of TNF-α, IL-6, and IL-1β mRNA levels were suppressed by H2. Moreover, we found that H2 alleviates the cell damage induced by LPS and reduces the ROS and NO levels of the cell. In addition, the intracellular ROS levels were significantly reduced in the LPS + 60%H2 group, and H2 inhibited TLR4 expression and NF-κB activation in macrophages caused by LPS.
In this study, LPS administration induced massive inflammatory cells and the release of cytokines in the serum, which were attenuated by H2 inhalation treatment. Moreover, we assessed the lung oxidative damage by measuring the level of MDA and NO, and the results showed that hydrogen gas decreased the oxidative lung damage caused by LPS. On the other hand, H2 dramatically inhibited the release of pro-inflammatory cytokines (TNF-α and IL-1β) in LPS-challenged mice and significantly up-regulated anti-inflammatory cytokines (IL-10). Previous studies reported that H2 inhalation could inhibit the release of pro-inflammatory cytokines [42]; however, there are no studies about its effect on anti-inflammatory cytokines.
Conclusions
In vivo and in vitro studies demonstrated that inhalation of H2 could relieve LPS-induced ALI and downregulate the TLR4-mediated NF-κB signaling pathway, thus inhibiting inflammation.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
References
Manicone AM. Role of the pulmonary epithelium and inflammatory signals in acute lung injury. Expert Rev Clin Immunol. 2009;5(1):63–75.
Orfanos SE, et al. Pulmonary endothelium in acute lung injury: from basic science to the critically ill. Intensive Care Med. 2004;30(9):1702–14.
Grommes J, Soehnlein O. Contribution of neutrophils to acute lung injury. Mol Med. 2011;17(3–4):293–307.
Goodman RB, et al. Cytokine-mediated inflammation in acute lung injury. Cytokine Growth Factor Rev. 2003;14(6):523–35.
Dushianthan A, et al. Acute respiratory distress syndrome and acute lung injury. Postgrad Med J. 2011;87(1031):612–22.
Wilson JG, et al. Mesenchymal stem (stromal) cells for treatment of ARDS: a phase 1 clinical trial. Lancet Respir Med. 2015;3(1):24–32.
Fan E, et al. Physical complications in acute lung injury survivors: a two-year longitudinal prospective study. Crit Care Med. 2014;42(4):849–59.
Wan L, et al. Preventive and therapeutic effects of thymol in a lipopolysaccharide-induced acute lung injury mice model. Inflammation. 2018;41(1):183–92.
Lv H, et al. Xanthohumol ameliorates lipopolysaccharide (LPS)-induced acute lung injury via induction of AMPK/GSK3beta-Nrf2 signal axis. Redox Biol. 2017;12:311–24.
Guijarro-Munoz I, et al. Lipopolysaccharide activates Toll-like receptor 4 (TLR4)-mediated NF-kappaB signaling pathway and proinflammatory response in human pericytes. J Biol Chem. 2014;289(4):2457–68.
Yang J, Zhao Y, Shao F. Non-canonical activation of inflammatory caspases by cytosolic LPS in innate immunity. Curr Opin Immunol. 2015;32:78–83.
Yan C, et al. TNF-alpha induction of IL-6 in alveolar type II epithelial cells: Contributions of JNK/c-Jun/AP-1 element, C/EBPdelta/C/EBP binding site and IKK/NF-kappaB p65/kappaB site. Mol Immunol. 2018;101:585–96.
Bhoj VG, Chen ZJ. Ubiquitylation in innate and adaptive immunity. Nature. 2009;458(7237):430–7.
Ben RK, Abdelly C, Savoure A. How reactive oxygen species and proline face stress together. Plant Physiol Biochem. 2014;80:278–84.
Park J, et al. Ganglioside GM3 suppresses lipopolysaccharide-induced inflammatory responses in rAW 264.7 macrophage cells through NF-kappaB, AP-1, and MAPKs signaling. J Cell Biochem. 2018;119(1):1173–82.
Tesoriere L, et al. Indicaxanthin inhibits NADPH oxidase (NOX)-1 activation and NF-kappaB-dependent release of inflammatory mediators and prevents the increase of epithelial permeability in IL-1beta-exposed Caco-2 cells. Br J Nutr. 2014;111(3):415–23.
Youn GS, et al. Overexpression of HDAC6 induces pro-inflammatory responses by regulating ROS-MAPK-NF-kappaB/AP-1 signaling pathways in macrophages. Free Radic Biol Med. 2016;97:14–23.
Xie K, et al. Protective effects of hydrogen gas on murine polymicrobial sepsis via reducing oxidative stress and HMGB1 release. Shock. 2010;34(1):90–7.
Sun Q, et al. Hydrogen-rich saline protects myocardium against ischemia/reperfusion injury in rats. Exp Biol Med (Maywood). 2009;234(10):1212–9.
Mao YF, et al. Hydrogen-rich saline reduces lung injury induced by intestinal ischemia/reperfusion in rats. Biochem Biophys Res Commun. 2009;381(4):602–5.
Chen XL, et al. Superoxide, H2O2, and iron are required for TNF-alpha-induced MCP-1 gene expression in endothelial cells: role of Rac1 and NADPH oxidase. Am J Physiol Heart Circ Physiol. 2004;286(3):H1001–7.
Nakao A, et al. Therapeutic antioxidant medical gas. J Clin Biochem Nutr. 2009;44(1):1–13.
George JF, Agarwal A. Hydrogen: another gas with therapeutic potential. Kidney Int. 2010;77(2):85–7.
Wu G, et al. Hypoxia Exacerbates Inflammatory Acute Lung Injury via the Toll-Like Receptor 4 Signaling Pathway. Front Immunol. 2018;9:1667.
Wang J, et al. Anti-inflammatory effects of apigenin in lipopolysaccharide-induced inflammatory in acute lung injury by suppressing COX-2 and NF-kB pathway. Inflammation. 2014;37(6):2085–90.
Huang CS, et al. Recent advances in hydrogen research as a therapeutic medical gas. Free Radic Res. 2010;44(9):971–82.
Huang CS, et al. Hydrogen inhalation reduced epithelial apoptosis in ventilator-induced lung injury via a mechanism involving nuclear factor-kappa B activation. Biochem Biophys Res Commun. 2011;408(2):253–8.
Huang CS, et al. Hydrogen inhalation ameliorates ventilator-induced lung injury. Crit Care. 2010;14(6):R234.
Kawamura T, et al. Inhaled hydrogen gas therapy for prevention of lung transplant-induced ischemia/reperfusion injury in rats. Transplantation. 2010;90(12):1344–51.
Sun Q, et al. Hydrogen-rich saline provides protection against hyperoxic lung injury. J Surg Res. 2011;165(1):e43–9.
Zhang X, et al. Protective effect of abamectin on acute lung injury induced by lipopolysaccharide in mice. Fundam Clin Pharmacol. 2011;25(6):700–7.
Giebelen IA, et al. Local stimulation of alpha7 cholinergic receptors inhibits LPS-induced TNF-alpha release in the mouse lung. Shock. 2007;28(6):700–3.
Sun R, et al. High concentration of hydrogen gas alleviates Lipopolysaccharide-induced lung injury via activating Nrf2 signaling pathway in mice. Int Immunopharmacol. 2021;101(Pt B): 108198.
Du J, et al. High concentration of hydrogen ameliorates lipopolysaccharide-induced acute lung injury in a sirt1-dependent manner. Respir Physiol Neurobiol. 2022;296: 103808.
Qiu R, et al. Dexmedetomidine restores septic renal function via promoting inflammation resolution in a rat sepsis model. Life Sci. 2018;204:1–8.
Imanifooladi AA, Yazdani S, Nourani MR. The role of nuclear factor-kappaB in inflammatory lung disease. Inflamm Allergy Drug Targets. 2010;9(3):197–205.
Jiao H, et al. Caveolin-1 Tyr14 phosphorylation induces interaction with TLR4 in endothelial cells and mediates MyD88-dependent signaling and sepsis-induced lung inflammation. J Immunol. 2013;191(12):6191–9.
Wang SL, et al. Dexmedetomidine preconditioning plays a neuroprotective role and suppresses TLR4/NF-kappaB pathways model of cerebral ischemia reperfusion. Biomed Pharmacother. 2017;93:1337–42.
Yao H, et al. Dexmedetomidine Inhibits TLR4/NF-kappaB activation and reduces acute kidney injury after orthotopic autologous liver transplantation in rats. Sci Rep. 2015;5:16849.
Song G, et al. H2 inhibits TNF-alpha-induced lectin-like oxidized LDL receptor-1 expression by inhibiting nuclear factor kappaB activation in endothelial cells. Biotechnol Lett. 2011;33(9):1715–22.
Ohsawa I, et al. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat Med. 2007;13(6):688–94.
Xie K, et al. Hydrogen gas improves survival rate and organ damage in zymosan-induced generalized inflammation model. Shock. 2010;34(5):495–501.
Funding
This work was supported by the Shanghai Asclepius Meditec Co., Ltd (project number: MR002-18–006).
Author information
Authors and Affiliations
Contributions
HY and YW contributed to the design and writing of this article. HY, YD, and YF performed the experiments and acquired all data. SM and ZG conducted a literature collection. All the authors participated in the analysis and interpretation of data, critical revision, and final approval of this article. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All experimental procedures were approved by the Experimentation Ethics Committee of the East Hospital, Tongji University School of Medicine in Shanghai, China(permit numbers: SCXK (Shanghai) 2017–0005).
Consent for publication
We claim that this manuscript has been read and approved by all authors.
Competing interests
All the authors declare that they have no conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yin, H., Feng, Y., Duan, Y. et al. Hydrogen gas alleviates lipopolysaccharide-induced acute lung injury and inflammatory response in mice. J Inflamm 19, 16 (2022). https://doi.org/10.1186/s12950-022-00314-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12950-022-00314-x