
Abstract
Background
Apolipoprotein A1 (ApoA1) is the major apoprotein constituent of high-density lipoprotein that can play important roles in tumor invasion and metastasis. The objective of the present study was to evaluate the association of two genetic variants (−75 G/A and +83 C/T) of APOA1 with predisposition to renal cancer.
Methods
A total of 432 subjects, including 216 pathologically-proven renal cancer cases and 216 age- and gender-matched healthy controls, were recruited into this hospital-based case–control study. Genotyping of the APOA1 was performed by polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) combined with gel electrophoresis, and then confirmed by direct sequencing.
Results
Patients with renal cancer had a significantly higher frequency of APOA1 -75 AA genotype [odds ratio (OR) = 2.10, 95 % confidence interval (CI) = 1.18, 3.75; P = 0.01] and APOA1 -75 A allele (OR =1.40, 95 % CI = 1.05, 1.87; P = 0.02) than controls. When stratifying by the distant metastasis status, patients with distant metastasis had a significantly higher frequency of APOA1 -75 AA genotype genotype (OR =2.20, 95 % CI = 1.04, 4.68; P = 0.04).
Conclusion
This study is, to our knowledge, the first to examine prospectively an increased risk role of APOA1 -75 AA genotype and APOA1 -75 A allele in renal cancer susceptibility.
Similar content being viewed by others

Introduction
Renal cancer is the predominant form of malignancy of the kidney and represents 3–4 % of all cancers [1]. In 2015, an estimated 61,560 new cases of kidney and renal pelvis cancers will be diagnosed in the United States [1]. An estimated 14,080 Americans are expected to die from the disease in 2015 [1]. The crude incidence rates of renal cell carcinoma were 7.1 and 3.1 per 100,000 population for men and women respectively in Japan in 1997 [2]. The crude incidence rates of renal cell carcinoma were increased from 4.5 to 5.6 per 100,000 population in China between 1989 and 2008 [3, 4]. Many epidemiological studies have found that environmental factors, such as smoking, diesel exhaust, and various dioxins, may be involved in the development of sporadic renal cancer [5–8]. Although many subjects are exposed to these risk factors during their lifetime, only some of them develop renal cancer, which suggests that genetic susceptibility may play a role in the etiology of renal cancer [9]. The impact of genetic background on renal cancer is still unclear.
Apolipoprotein A1 (ApoA1) is the major apoprotein constituent of high-density lipoprotein (HDL) that can play important roles in tumor invasion and metastasis [10, 11]. Recent findings revealed the crucial roles of ApoA1 in inflammation, tumor growth, angiogenesis, invasion and metastasis [12–14]. APOA1 gene, located on the 11q23-q24, encodes apoA1 [11, 15]. There are several single-nucleotide polymorphisms (SNPs) in the APOA1 gene [16]. Two SNPs (−75 G/A and +83 C/T) of APOA1 play an important role in lipid metabolism [17–19]. It has also been found that APOA1 -75 G/A and +83 C/T genotypes were associated with susceptibility to breast cancer and lymph node metastases occurrence, respectively [20].
To our best knowledge, no reports concerning the role of APOA1 -75 G/A and +83 C/T genotypes on renal cancer risk have been reported yet. We hypothesized that APOA1 -75 G/A and +83 C/T genotypes were associated with renal cancer risk. To test this hypothesis, we performed a prospective hospital-based case–control study to evaluate the association of the APOA1 -75 G/A and +83 C/T genotypes with predisposition to renal cancer.
Materials and methods
Study population
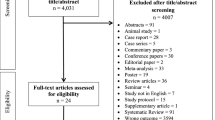
A total of 432 subjects, including 216 pathologically-proven renal cancer cases and 216 age- and gender-matched healthy controls, were recruited into this hospital-based case–control study between February 2012 and December 2014 in the West China Hospital of Sichuan University [21]. All renal cancer cases were histopathologically confirmed. We extracted the following information: tumor grade, tumor classification, lymph node invasion status, distant metastasis status and the pathology of renal cancer. In order to confirm that healthy controls were healthy and free of cancer, volunteers underwent various tests that included physical exams, questionnaires about their health and history, chest X-rays, blood and urine tests for various tumor markers, abdominal ultrasound, gastric endoscopy, and colon enema. The patient or their families/surrogates were interviewed. The Institutional Ethical Committee of the West China Hospital of Sichuan University approved all parts of the study, and informed consent according to the Declaration of Helsinki was obtained from all participants or their families/surrogates.
DNA extraction and genotyping
Genomic DNA was isolated from 20 g/L ethylenediaminetetraacetic acid (EDTA) or sodium citrate anticoagulated 3–5 ml venous blood by the commercially available Qiagen kit (QIAGEN Inc., Valencia, CA, USA) and stored at 4 °C. Genotyping of the APOA1 was performed by polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP). Based on the GenBank reference sequence, the PCR primers designed for the APOA1 -75 G/A and +83 C/T were as follows: 5ʹ-AGG GAC AGA GCT GAT CCT TGA ACT CTT AAG-3ʹ (forward) and 5ʹ-TTA GGG GAC ACC TAG CCC TCA GGA AGA GCA-3ʹ (reverse). The restriction endonuclease enzyme MspI digested the amplified PCR products overnight. Electrophoresis in a 3 % agarose gel followed by ethidium bromide staining and ultraviolet illumination allowed detection of the alleles. The presence of the MspI restriction stie at −75 bp (G allele) and at +83 bp (C allele) in the 433 bp product resulted in four fragments of 45, 66, 113 and 209 bp (Fig. 1). The absence of the restriction site at −75 bp (A allele) resulted in three fragments of 45, 179 and 209 bp (Fig. 1). The absence of the restriction site at +83 bp (T allele) created a larger fragment of 254 bp instead of two fragments of 45 and 209 bp (Fig. 1). For quality control, two independent observers randomly chose 44 samples (22 cases and 22 controls) by computer-generated number scheme. They performed double sampling PCR-RFLP and found no differences, and then confirmed by direct sequencing from Qiagen cleaned up DNA.

Electrophoresis in a 3 % agarose gel after the Mspl digested in renal cancer cases and healthy controls
Statistical analysis
Data are presented as percentages for categorical variables or as means ± standard deviation (SD). Differences between categorical variables were evaluated using Pearson x 2 test, while those between continuous variables were assessed by Student’s t test. The existence of differences in genotypic frequencies between groups was assessed by means of Pearson x 2 test and calculating the odds ratio (OR) with the 95 % confidence intervals (CI). A P-value was considered significant at a level of < 0.05. The Statistical Analysis System software (Version 9.1; SAS Institute Inc., Cary, NC, USA) was used for all statistical tests.
Results
Characteristics of participants
Healthy volunteers and cancer patients were not significantly different in terms of age distribution and gender (Table 1). For renal cancer cases, the tumor grade, tumor classification, lymph node invasion status, distant metastasis status and pathology were presented in a separate paper [21].
APOA1 -75 G/A polymorphisms and renal cancer
Patients with renal cancer had a significantly higher frequency of APOA1 -75 AA genotype [odds ratio (OR) = 2.10, 95 % confidence interval (CI) = 1.18, 3.75; P = 0.01] and APOA1 -75 A allele (OR =1.40, 95 % CI = 1.05, 1.87; P = 0.02) than controls (Table 2). When stratifying by the distant metastasis status, patients with distant metastasis had a significantly higher frequency of APOA1 -75 AA genotype genotype (OR =2.20, 95 % CI = 1.04, 4.68; P = 0.04) (Table 3). When stratifying by the tumor grade, tumor classification, lymph node invasion status and pathology, no significant differences were found (Table 3).
APOA1 + 83 C/T polymorphisms and renal cancer
We did not find any association between APOA1 + 83 C/T polymorphisms and renal cancer risk (Table 2).
Discussion
Many studies have suggested that genetic susceptibility may play a role in the etiology of renal cancer. The DKK3 polymorphisms were associated with renal cancer and that the DKK2 rs17037102 polymorphism might be a predictor for survival in patients with renal cancer after radical nephrectomy [22]. We recently found that IL-6 -174 CC genotype was associated with an increased risk for renal cancer [21]. The functional -31G/C polymorphism in the promoter of survivin might influence the susceptibility and progression of renal cancer in the Chinese population [23]. The polymorphisms of the CYP1B1 gene at codons 119 and 432 might be risk factors for renal cancer, especially in the male population [7]. The CYP1A1 polymorphisms might play an important role in the etiology of renal cancer [24]. The polymorphisms of catechol-O-methyltransferase in men were associated with renal cancer [25]. The R allele of paraoxonase-1 gene Q192R polymorphism might protect against renal cancer [26]. Two SNPs in AGTR1 might be a candidate pathway in renal cancer etiology [27]. Polymorphisms in genes of the renin-angiotensin-aldosterone system (AGTR1 and AGT) influenced renal cell cancer susceptibility [27]. A common variant, rs35252396, at 8q24.21 was associated with renal cell cancer [28].
The APOA1 gene polymorphisms were also associated with many other diseases. It has been found that the APOA1 -75 G/A polymorphism was associated with gallstone disease [29]. The APOA1 -75 G/A and +83 C/T genotypes were also associated with susceptibility to breast cancer and lymph node metastases occurrence, respectively [20]. A pilot study found that APOA1 polymorphisms (−75 G/A and +83 C/T) might be susceptibility to myocardial infarction in a north Indian population [19]. The individuals with the APOA1 -75 A allele were likely to have a lower risk of coronary artery disease as a result of its effect on higher serum concentrations of ApoA1 and HDL-C [30]. The APOA1 -75G/A promoter polymorphism was associated with cognitive performance in multiple sclerosis [31]. It has been found that the APOA1 polymorphisms (−75 G/A and +83 C/T) could be as risk factors for hypertension and obesity in a Brazilian elderly cohort [32]. The APOA1 -75 A allele was associated with an increased risk for Alzheimer’s disease [33]. The APOA1 -75 AA genotype was associated with a higher acute lung injury risk after cardiopulmonary bypass surgery [34].
The exact biological mechanism of the association between the APOA1 -75G/A polymorphisms and the risk of renal cancer is still unclear. ApoA1 can play important roles in tumor growth, angiogenesis, invasion and metastasis [12–14]. Expression of ApoA1 is associated with colonic adenocarcinoma progression, and thus ApoA1 is a potential marker of the aggression [35]. It has also been found that APOA1 -75 G/A and +83 C/T genotypes were associated with susceptibility to breast cancer and lymph node metastases occurrence, respectively [20]. A recent fascinating study reveals an overall protective ability of HDL, specifically APOA1, to induce tumor suppression through both innate and adaptive immune processes in multiple animal tumor models [36]. Tabet et al. demonstrated that HDL’s anti-inflammatory properties were conferred, in part, through HDL-micro-RNA (miR)-223 delivery and translational repression of ICAM-1 in endothelial cells [37]. However, the miR221/222 cluster increases the aggressiveness of tumors in epithelial cancers, through repression of tumor suppressors and through induction of cell motility [38]. APOA1 allelic variety may have an impact on angiogenesis [39]. APOA1 binding protein (AIBP) positively regulates cholesterol efflux from endothelial cells and that effective cholesterol efflux is critical for proper angiogenesis [39]. AIBP is highly expressed in human renal cancer and in 83 % of cancers in general [39]. But there is no study about the interaction of AIBP and the APOA1 alleles. Further research on the possible interaction of AIBP and the APOA1 alleles is necessary.
Some shortcomings of this study should be mentioned. First of all, potential selection bias might have been present, because this is a hospital based case control study and the subjects may not be representative of the general population. Second, this study is limited by its size and lack of replication. Finally, further research on the biological mechanism of the association between the APOA1 -75G/A polymorphisms and the risk of renal cancer is necessary.
In conclusion, to our best knowledge, up to now this study is the first to examine prospectively an increased risk role of APOA1 -75 AA genotype and APOA1 -75 A allele in renal cancer susceptibility.
References
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. 2015;65:5–29.
Marumo K, Satomi Y, Miyao N, Hasegawa M, Tomita Y, Igarashi T, et al. Japanese Society of Renal C: The prevalence of renal cell carcinoma: a nation-wide survey in Japan in 1997. Int J Urol. 2001;8:359–65.
Chen W, Zheng R, Zeng H, Zhang S, He J. Annual report on status of cancer in China, 2011. Chin J Cancer Res. 2015;27:2–12.
Chen WQ, Zheng RS, Zeng HM, Zhang SW, Zhao P, He J. Trend analysis and projection of cancer incidence in China between 1989 and 2008. Zhonghua Zhong Liu Za Zhi. 2012;34:517–24.
Kroeger N, Klatte T, Birkhauser FD, Rampersaud EN, Seligson DB, Zomorodian N, et al. Smoking negatively impacts renal cell carcinoma overall and cancer-specific survival. Cancer. 2012;118:1795–802.
Theis RP, Dolwick Grieb SM, Burr D, Siddiqui T, Asal NR. Smoking, environmental tobacco smoke, and risk of renal cell cancer: a population-based case–control study. BMC Cancer. 2008;8:387.
Sasaki M, Tanaka Y, Okino ST, Nomoto M, Yonezawa S, Nakagawa M, et al. Polymorphisms of the CYP1B1 gene as risk factors for human renal cell cancer. Clin Cancer Res. 2004;10:2015–9.
McLaughlin JK, Gao YT, Gao RN, Zheng W, Ji BT, Blot WJ, et al. Risk factors for renal-cell cancer in Shanghai, China. Int J Cancer. 1992;52:562–5.
Cao Q, Qin C, Ju X, Meng X, Wang M, Zhu J, et al. Chromosome 11q13.3 variant modifies renal cell cancer risk in a Chinese population. Mutagenesis. 2012;27:345–50.
Segrest JP, Li L, Anantharamaiah GM, Harvey SC, Liadaki KN, Zannis V. Structure and function of apolipoprotein A-I and high-density lipoprotein. Curr Opin Lipidol. 2000;11:105–15.
Breslow JL, Ross D, McPherson J, Williams H, Kurnit D, Nussbaum AL, et al. Isolation and characterization of cDNA clones for human apolipoprotein A-I. Proc Natl Acad Sci U S A. 1982;79:6861–5.
Clarke CH, Yip C, Badgwell D, Fung ET, Coombes KR, Zhang Z, et al. Proteomic biomarkers apolipoprotein A1, truncated transthyretin and connective tissue activating protein III enhance the sensitivity of CA125 for detecting early stage epithelial ovarian cancer. Gynecol Oncol. 2011;122:548–53.
Moore LE, Fung ET, McGuire M, Rabkin CC, Molinaro A, Wang Z, et al. Evaluation of apolipoprotein A1 and posttranslationally modified forms of transthyretin as biomarkers for ovarian cancer detection in an independent study population. Cancer Epidemiol Biomarkers Prev. 2006;15:1641–6.
Charet JC, Watine J, Marre A, Charet P. Prognostic value of serum levels of cholesterol and apolipoprotein A1 in pulmonary cancer. Ann Biol Clin (Paris). 1997;55:52.
Arinami T, Hirano T, Kobayashi K, Yamanouchi Y, Hamaguchi H. Assignment of the apolipoprotein A-I gene to 11q23 based on RFLP in a case with a partial deletion of chromosome 11, del(11)(q23.3----qter). Hum Genet. 1990;85:39–40.
Karathanasis SK. Apolipoprotein multigene family: tandem organization of human apolipoprotein AI, CIII, and AIV genes. Proc Natl Acad Sci U S A. 1985;82:6374–8.
Saha N, Tay JS, Low PS, Humphries SE. Guanidine to adenine (G/A) substitution in the promoter region of the apolipoprotein AI gene is associated with elevated serum apolipoprotein AI levels in Chinese non-smokers. Genet Epidemiol. 1994;11:255–64.
Wang XL, Badenhop R, Humphrey KE, Wilcken DE. New MspI polymorphism at +83 bp of the human apolipoprotein AI gene: association with increased circulating high density lipoprotein cholesterol levels. Genet Epidemiol. 1996;13:1–10.
Dawar R, Gurtoo A, Singh R. Apolipoprotein A1 gene polymorphism (G-75A and C + 83 T) in patients with myocardial infarction: a pilot study in a north Indian population. Am J Clin Pathol. 2010;134:249–55.
Hamrita B, Ben Nasr H, Gabbouj S, Bouaouina N, Chouchane L, Chahed K. Apolipoprotein A1–75 G/A and +83 C/T polymorphisms: susceptibility and prognostic implications in breast cancer. Mol Biol Rep. 2011;38:1637–43.
Liu Z, Wang Z, Xiao Y, Lu Y, Lu Y. Association between the interleukin-6 gene polymorphisms and renal cancer risk. Immunol Lett. 2015;164:125–8.
Hirata H, Hinoda Y, Nakajima K, Kikuno N, Yamamura S, Kawakami K, et al. Wnt antagonist gene polymorphisms and renal cancer. Cancer. 2009;115:4488–503.
Qin C, Cao Q, Li P, Ju X, Wang M, Chen J, et al. Functional promoter -31G > C variant in survivin gene is associated with risk and progression of renal cell cancer in a Chinese population. PLoS One. 2012;7:e28829.
Chen J, Cheng M, Yi L, Jiang CB. Relationship between CYP1A1 genetic polymorphisms and renal cancer in China. Asian Pac J Cancer Prev. 2011;12:2163–6.
Tanaka Y, Hirata H, Chen Z, Kikuno N, Kawamoto K, Majid S, et al. Polymorphisms of catechol-O-methyltransferase in men with renal cell cancer. Cancer Epidemiol Biomarkers Prev. 2007;16:92–7.
Uyar OA, Kara M, Erol D, Ardicoglu A, Yuce H. Investigating paraoxonase-1 gene Q192R and L55M polymorphism in patients with renal cell cancer. Genet Mol Res. 2011;10:133–9.
Deckers IA, van den Brandt PA, van Engeland M, van Schooten FJ, Godschalk RW, Keszei AP, et al. Polymorphisms in genes of the renin-angiotensin-aldosterone system and renal cell cancer risk: interplay with hypertension and intakes of sodium, potassium and fluid. Int J Cancer. 2015;136:1104–16.
Gudmundsson J, Sulem P, Gudbjartsson DF, Masson G, Petursdottir V, Hardarson S, et al. A common variant at 8q24.21 is associated with renal cell cancer. Nat Commun. 2013;4:2776.
Dixit M, Choudhuri G, Saxena R, Mittal B. Association of apolipoprotein A1-C3 gene cluster polymorphisms with gallstone disease. Can J Gastroenterol. 2007;21:569–75.
Liao B, Cheng K, Dong S, Liu H, Xu Z. Effect of apolipoprotein A1 genetic polymorphisms on lipid profiles and the risk of coronary artery disease. Diagn Pathol. 2015;10:102.
Koutsis G, Panas M, Giogkaraki E, Karadima G, Sfagos C, Vassilopoulos D. An APOA1 promoter polymorphism is associated with cognitive performance in patients with multiple sclerosis. Mult Scler. 2009;15:174–9.
Chen ES, Mazzotti DR, Furuya TK, Cendoroglo MS, Ramos LR, Araujo LQ, et al. Apolipoprotein A1 gene polymorphisms as risk factors for hypertension and obesity. Clin Exp Med. 2009;9:319–25.
Vollbach H, Heun R, Morris CM, Edwardson JA, McKeith IG, Jessen F, et al. APOA1 polymorphism influences risk for early-onset nonfamiliar AD. Ann Neurol. 2005;58:436–41.
Tu J, Zhang B, Chen Y, Liang B, Liang D, Liu G, et al. Association of apolipoprotein A1–75 G/A polymorphism with susceptibility to the development of acute lung injury after cardiopulmonary bypass surgery. Lipids Health Dis. 2013;12:172.
Tachibana M, Ohkura Y, Kobayashi Y, Sakamoto H, Tanaka Y, Watanabe J, et al. Expression of apolipoprotein A1 in colonic adenocarcinoma. Anticancer Res. 2003;23:4161–7.
Zamanian-Daryoush M, Lindner D, Tallant TC, Wang Z, Buffa J, Klipfell E, et al. The cardioprotective protein apolipoprotein A1 promotes potent anti-tumorigenic effects. J Biol Chem. 2013;288:21237–52.
Tabet F, Vickers KC, Cuesta Torres LF, Wiese CB, Shoucri BM, Lambert G, et al. HDL-transferred microRNA-223 regulates ICAM-1 expression in endothelial cells. Nat Commun. 2014;5:3292.
Chistiakov DA, Sobenin IA, Orekhov AN, Bobryshev YV. Human miR-221/222 in Physiological and Atherosclerotic Vascular Remodeling. Biomed Res Int. 2015;2015:354517.
Fang L, Choi SH, Baek JS, Liu C, Almazan F, Ulrich F, et al. Control of angiogenesis by AIBP-mediated cholesterol efflux. Nature. 2013;498:118–22.
Acknowledgements
Thanks are expressed to all coinvestigators, local project coordinators, research assistants, laboratory technicians, and secretaries/administrative assistants.
Funding
This research received no specific grant from any funding agency in the public, commercial, nor for-profit sectors.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
ZHL, YMX and YPL carried out the molecular genetic studies and drafted the manuscript. LYT and LJ carried out the genotyping. YJW, RCZ and QW participated in the design of the study and performed the statistical analysis. ZHL, YMX, LYT, LJ, YJW, RCZ, QW and YPL conceived of the study, and participated in its design and coordination and helped to draft the manuscript. All authors read and approved the final manuscript.
ZhiHong Liu and YingMing Xiao, are joint first authors.
ZhiHong Liu and YingMing Xiao contributed equally to this work.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Liu, Z., Xiao, Y., Tang, L. et al. Apolipoprotein A1 -75 G/A and +83 C/T polymorphisms and renal cancer risk. Lipids Health Dis 14, 143 (2015). https://doi.org/10.1186/s12944-015-0132-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12944-015-0132-0