
Abstract
Sonic Hedgehog (Shh) signaling cascade is one of the complex signaling pathways that control the accurately organized developmental processes in multicellular organisms. This pathway has fundamental roles in the tumor formation and induction of resistance to conventional therapies. Numerous non-coding RNAs (ncRNAs) have been found to interact with Shh pathway to induce several pathogenic processes, including malignant and non-malignant disorders. Many of the Shh-interacting ncRNAs are oncogenes whose expressions have been increased in diverse malignancies. A number of Shh-targeting miRNAs such as miR-26a, miR-1471, miR-129-5p, miR-361-3p, miR-26b-5p and miR-361-3p have been found to be down-regulated in tumor tissues. In addition to malignant conditions, Shh-interacting ncRNAs can affect tissue regeneration and development of neurodegenerative disorders. XIST, LOC101930370, lncRNA-Hh, circBCBM1, SNHG6, LINC‐PINT, TUG1 and LINC01426 are among long non-coding RNAs/circular RNAs that interact with Shh pathway. Moreover, miR-424, miR-26a, miR-1471, miR-125a, miR-210, miR-130a-5p, miR-199b, miR-155, let-7, miR-30c, miR-326, miR-26b-5p, miR-9, miR-132, miR-146a and miR-425-5p are among Shh-interacting miRNAs. The current review summarizes the interactions between ncRNAs and Shh in these contexts.
Similar content being viewed by others


Introduction
Sonic Hedgehog (Shh) signaling cascade is one of the complicated signaling pathways that administrate the accurately controlled developmental processes in multicellular organisms. It has an important role in the establishment of the outlines of cellular differentiation to regulate multifaceted organ formation. This pathway affects these cellular processes via a cascade that changes the equilibrium between activator and repressor types of glioma-associated oncogene (Gli) transcription factors. A number of Hedgehog (Hh) ligands as well as Patched receptors, Smoothened receptor, Suppressor of fused homolog, Kif7, PKA and cAMP participate in the transfer of signals to the Gli transcription factors. Transfer of the activator form of Gli to the nucleus and its binding with the promoters of target genes lead to the stimulation of the transcription of these genes [1]. Hh gene has been firstly discovered about four decades ago via genetic screen experiments in Drosophila [2].
Shh participates in the tissue regeneration processes and repair mechanism in the post-embryonic period. This pathway has a crucial role in the induction of diverse populations of neurons in the central nervous system, governing several morphogenetic processes in this system [3].
Abnormal regulation of these signals has been shown to be associated with congenital malformations, aberrant tissue regeneration, stem cell renewal and carcinogenesis [4]. Expression, cellular uptake and translocation of the Shh protein as a key Hh ligand precursor have important effects in the regulatory function of Shh signaling [5]. Two other Hedgehog homologues, namely Desert (Dhh) and Indian (Ihh) have been identified in mammals.
Functional studies have shown the importance of Shh signaling in ventral cell type induction. Moreover, disruption of this pathway and recessive mutations have led to cyclopia and severe holoprosencephaly in mice, respectively [6]. In human, heterozygote mutations in Shh have been associated with different clinical features of holoprosencephaly [7].
Shh signaling is also implicated in the regulation of function of normal adult stem cells as well as cancer stem cells [8]. Dysregulation of the Hh signaling pathway has been linked with developmental abnormalities including Gorlin syndrome [9] and cancer [10, 11]. Abnormal activity of this pathway is also involved in the tumor formation and induction of resistance to radio/chemotherapy [12]. Thus, efforts have been made to find novel Shh signaling inhibitors to combat these features [12].
More recently, numerous non-coding RNAs (ncRNAs) have been found to interact with Shh pathway to induce several pathogenic processes, including malignant and non-malignant disorders. The current review summarizes the interactions between ncRNAs and Shh in these contexts. Long non-coding RNAs (lncRNAs), microRNAs (miRNAs) and circular RNAs (circRNAs) are three main regulatory ncRNAs which are discussed in this context. LncRNAs are transcripts with sizes more than 200 nt that regulate expression of genes at different levels. They can regulate chromatin function, influence the assemblage and functions of membraneless nuclear bodies, control the stability and expression of cytoplasmic mRNAs and interfere with signaling pathways [13]. miRNAs have about 22 nt and mainly affect gene expression at post transcriptional level [14]. Finally, circRNAs are made by either typical spliceosome-mediated or lariat-type splicing. They can regulate expression of genes through different mechanisms [15].
For the purpose of preparation of the current review, we searched Google Scholar and PubMed databases with the key words “Shh signaling” OR “Sonic Hedgehog” AND “lncRNA” OR “miRNA” OR “circRNA”. Then, we assessed the abstract of retrieved articles to validate their relevance with the topic. We included studies that assessed function of ncRNAs in cell lines, animal model or clinical samples. A total of 50 studies were included in this review article.
Cell line studies
Non-malignant disorders
The importance of interactions between ncRNAs and Shh pathway has been assessed in different cell lines. This type of interaction has been found to be implicated in the pathophysiology of alopecia. This speculation is based on the results of three-dimensional culture of dermal papilla cells, a group of cells that induce regeneration of hair follicles. Experiments in this type of culture have verified up-regulation of XIST lncRNA and Shh and down-regulation of miR-424. Mechanistically, XIST has been found to sponge miR-424 to increase Shh expression. XIST silencing has led to inhibition of activity of dermal papilla cells, suppression of their proliferation and reduction of ALP activity. In fact, XIST silencing has inhibited Shh mediated hedgehog signals through affecting expression of miR-424 [16].
Human development
Shh-interacting ncRNAs are also involved in the developmental processes. For instance, serum response factor (SRF) controls lineage specification of embryonic stem cell progenitor cells through miR-210-mediated gene silencing. Up-regulation of miR-210 in murine embryonic stem cells-originated embryoid bodies has suppressed cell growth and blocked expression of cardiac progenitor proteins Nkx2.5 and Gata4 and terminal differentiated contractile markers Mlc2v and βMHC. On the other hand, miR-210 silencing has led to activation of cardiac progenitor gene. The effect of miR-210 is exerted through decreasing activity of Shh signaling, which nurtures the cardiac progenitor program. Mechanistically, miR-210 silences Shh activity through targeting 3' UTR of Shh transcript [17]. Activation of Shh/Gli1 signaling pathway through miR-130a-5p/Foxa2 axis has been shown to affect development of fetal lung, thus being involved in the pathogeensis of congenital diaphragmatic hernia [18]. miR-199 is another miRNA that participate in craniofacial development through modulation of Shh pathway [19].
Cancer
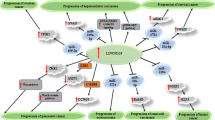
The interaction between ncRNAs and Shh signaling pathway has also been assessed in cancer cell lines. For instance, the tumor suppressor miRNA miR-26a has been found to be down-regulated in breast cancer cell lines. Up-regulation of miR-26a has led to blockade of cell proliferation, clone formation capacity and metastatic aptitude of breast cancer cells, and induction of sensitivity to docetaxel. miR-26a could directly target FAM98A. Up-regulation of this miRNA has resulted in down-regulation of FAM98A, SHH, SMO and GLI1. Taken together, miR-26a suppresses breast carcinogenesis through inhibiting expression of FAM98A, and decreasing activity of Shh pathway [20]. miR-1471 is another down-regulated miRNA in breast cancer cells. This miRNA has been found to be sponged by LOC101930370. LOC101930370 silencing has suppressed progression of breast cancer, while inhibition of miR-1471 has increased aggressive and metastatic abilities of MCF-7 cells. Furthermore, expression levels of SHH and Gli-1 have been significantly decreased following LOC101930370 silencing, and increased by miR-1471 inhibition. Cumulatively, LOC101930370 has been found to increase expression of SHH through sponging miR-1471 [21]. Another study has revealed dysregulation of several lncRNAs in Twist-positive mammosphere cells in breast cancer cell lines. Notably, the Shh-GLI1-related lncRNA-Hh has been among these lncRNAs. Expression of this lncRNA is regulated by Twist. Moreover, lncRNA-Hh can directly target GAS1 to induce Hh activity, which in turn enhances expression of GLI1, and increases SOX2 and OCT4 levels to regulate maintenance of cancer stem cells. The latter is reflected in enhancement of mammosphere-formation efficiency and self-renewal ability in cell lines. Knock down of lncRNA-Hh in Twist-positive breast cancer cells has attenuated activity of Shh-GLI1 signaling and decreased levels of SOX and OCT4 [22]. CircBCBM1 is another example of ncRNAs that can promote metatstatic ability of breast cancer cells through acting as a molecular sponge for miR-125a and modulating expression of BRD4. This circRNA also up-regulate MMP9 levels through enhancing activity of Shh pathway [23]. Figure 1 shows the role of Shh-interacting ncRNAs in breast cancer.

A schematic diagram of the role of several ncRNAs in triggering the Sonic Hedgehog signaling cascade in Breast Cancer. Mounting researches suggest that dysregulation of sonic hedgehog signaling pathway can play a key role in tumorigenesis in breast cancer cells. As an illustration, the recent study has detected that overexpression of lncRNA-Hh can activate Shh-GLI1 signaling and promote the expression levels of SOX2 and OCT4, thereby enhancing cancer stem cells generation in Twist-positive breast cancer cells [22]. Further experiment has validated that lncRNA LOC101930370 can significantly elevate SHH and Gli-1 expression via sponging miR-1471, therefore promoting cell proliferation and metastasis in breast cancer cells by modulating the hedgehog cascade [21]. Moreover, another research has pointed out that miR-26a has a remarkable part in suppressing breast cancer cell proliferation and invasion via downregulating the expression levels of FAM98A, SHH, SMO and GlI1, thereby inactivating the sonic hedgehog pathway in tumor cells [20]
In pancreatic cancer cell lines, miR-132 expression has been found to be up-regulated parallel with down-regulation of Shh levels. Besides, miR-132 mimics could significantly decrease expression of Shh at both transcript and protein levels, facilitating proliferation of pancreatic cancer cells, which has been accompanied by down-regulation of Cyclin-D1, cleaved Caspase-3/9, and suppression of cell apoptosis [24].
Shh-interacting ncRNAs can also affect the pathogenesis of brain tumors. For instance, miR-326 can effectively suppress proliferation, and induce apoptosis in glioma cells via attenuating the activation of the SHH/GLI1 pathway [25]. Moreover, miR-9 via targeting PTCH1 and enhancing expression of GLI1 can trigger the activation of Shh cascade and affect expression of drug efflux transporters, MDR1 and ABCG2 in glioblastoma cells, therefore enhancing Temozolomide resistance in tumor cells [26]. Figure 2 shows the role of Shh-interacting miRNA in glioma/glioblastoma.

A schematic representation of the role of several miRNAs in regulating the sonic hedgehog signaling pathway in glioma. Accumulating evidence has revealed that upregulation of miR-326 in combination with curcumin can effectively contribute in the suppression of proliferation, and elevation of the apoptosis process in glioma cells via attenuating the activation of the SHH/GLI1 pathway [25]. Another finding confirms that miR-9 via targeting PTCH1 and promoting expression of GLI1 can trigger the activation of sonic hedgehog cascade and modulate expression of drug efflux transporters, MDR1 and ABCG2 in glioblastoma cells, therefore enhancing Temozolomide resistance in tumor cells [26]. Furthermore, mounting research has demonstrated that Chidamide can play an important role in inhibiting the expression levels of Shh, Ihh, and Dhh via upregulating miR-338-5p, thereby suppressing the growth rate, migration, and invasion of human malignant glioma cells. In fact, Chidamide exerts its effects by enhancing oxidative stress via the miR-338-5p-mediated regulation of Hedgehog pathway [27]
Additional in vitro studies have revealed participation of Shh-related ncRNAs in several developmental processes as well as carcinogenic processes (Table 1).
Animal studies
Animal studies have shown participation of Shh-related ncRNAs in a variety of non-neoplastic disorders, namely acute myocardial infarction, alopecia, cerebrovascular disorders, diabetes mellitus, inflammatory bowel disease, lung fibrosis, osteoporosis, Parkinson's disease and trigeminal neuralgia as well as different types of cancers (Table 2). For instance, knock-down of miR-802-5p has resulted in reduction of cell apoptosis after myocardial infarction through enhancing activity of Shh signaling, thus decreasing myocardial injury and improving cardiac function [57]. Moreover, experiments in animal models have shown up-regulation of XIST increases dermal papilla cells-mediated hair follicle regeneration via targeting miR-424 to promote Shh expression, thus activating hedgehog signaling [16]. Moreover, miR-153 has been found to decrease expression of PTC expression and enhance activity of Shh signaling pathway to increase angiogenesis in a rat model of cerebral ischemic injury [58].
Experiments in animal models of breast cancer have verified that knock-down of lncRNA-Hh [22] and circBCBM1 [23] can led to reduction of tumor growth. Meanwhile, miR-326 has been shown to increase effects of curcumin in animal models of glioblastoma through modulation of Shh/GLI1 signaling pathway [25].
Studies in clinical samples
Expression assays in clinical samples from a variety of tumor types have indicated down-regulation of Shh-targeting miRNAs such as miR-26a [20] and miR-1471 [21] in breast cancer, miR-129-5p in cervical cancer [28], miR-361-3p in retinoblastoma, miR-26b-5p in gallbladder carcinoma [33], and miR-361-3p in retinoblastoma [53]. In gallbladder cancer, dysregulation of miR-26b-5p has been associated with age and sex of patients, tumor invasion, differentiation degree, tumor location, and TNM stage [33]. Conversely, an expression assay in pancreatic cancer samples has shown up-regulation of miR-132 and down-regulation of Shh [24]. In neuroblastoma samples, down-regulation of CDON and up-regulation of miR-181-a and miR-181-b have been associated with poor overall survival, higher tumor stage and more aggressive phenotype [64].
Shh-related lncRNAs are also involved in the process of keloid formation. Expression assays in keloid tissues and adjacent normal skin epidermis have shown differential expression of 30 lncRNAs and 33 mRNAs between these two sets of samples. Dysregulated lncRNAs included up-regulated lncRNAs AK055628, MIAT, MIR31HG, RP11-264F23.3, and AC073257.2, and downregulated lncRNAs RP11-12M9.3, XLOC_007437, XLOC_009485, RP5-1042I8.7, and HNF1A-AS1 [66].
Table 3 summarizes dysregulation of Shh signaling-related ncRNAs and in clinical samples.
Discussion
Shh signaling is involved in a variety of cellular functions, including tissue development and regeneration, stem cell functions and carcinogenesis. Thus, it is not surprising that ncRNAs that regulate activity of this pathway are implicated in the pathogenesis of a wide range of human disorders. In fact, this pathway represents a prototype of shared pathways between embryogenesis and carcinogenesis.
In the context of malignant disorders, Shh-interacting ncRNAs not only affect cancer progression, but also determine response of cancer cells to a variety of anticancer therapies. Both functions can be explained by the crucial roles of this pathway in the induction of stemness. However, at least in some types of cancers, Shh signaling seems to have protective effects against carcinogenesis. For instance, in pancreatic cancer, expression of the Shh-targeting miRNA miR-132 has been found to be up-regulated parallel with down-regulation of Shh levels [24]. Shh pathway can also induce epithelial-to-mesenchymal transition in gastric, pancreatic, and bladder cancers [67,68,69]. Thus, Shh-interacting ncRNAs might also affect this feature.
It is estimated that one-third of malignancies are correlated with abnormal activity of the Hh signaling pathway [70]. Dysregulation of Hh signaling can contribute to the imitation, growth, metastasis, and apoptosis of several types of cancers. In fact, three patterns of induction of the Hh signaling cascade have been identified in several cancers. These patterns are ligand independent oncogenic Hh pathway, autocrine or juxtacrine and paracrine or reverse paracrine patterns [70].
Moreover, there are several examples of interactions between two classes of ncRNAs in the context of regulation of activity of Shh signals. XIST/miR-424, LOC101930370/miR-1471, circBCBM1/miR-125a, TUG1/miR-132, SNHG6/miR-26b-5p, LINC‐PINT/miR‐425‐5p, and SNHG6/miR-135a-5p are examples of lncRNA/miRNA or circRNA/miRNA pairs that cooperatively regulate activity of Shh pathway. The regulatory impact of these axes on function of Shh pathway should be assessed in different cellular and disease contexts to find whether they act in a context-specific manner or a ubiquitous manner. This has importance in design of novel therapies for each disorder in which abnormal function of Shh pathway has been detected. However, circRNAs have limited roles as miRNA sponges in most cases [71,72,73,74]. In fact, most circRNAs are much less abundant than miRNAs and are not predicted to function as miRNA sponges [75, 76].
Although Shh-related ncRNAs are expected to influence the prognosis and clinical outcome of cancer, this issue has been verified only in the gallbladder cancer, neuroblastoma and laryngeal cancer. Thus, future studies should assess the prognostic roles of these ncRNAs as well as their potential as diagnostic markers.
A number of anti-cancer agents such as propofol have been found to exert their effects through modulation of Shh-related ncRNAs. This agent has been found to simultaneously decrease activity of Shh and PI3K/AKT pathways [48]. Therefore, modulation of expression of Shh-related ncRNAs is a promising anticancer strategy.
Taken together, the data presented above indicates contribution of several ncRNAs in the regulation of Shh pathway and their involvement in the pathogenesis of several disorders. However, this data has some limitations. No comprehensive assessments of different types of ncRNAs using next generation sequencing techniques have been performed. Thus, the interactive networks between different types of ncRNAs and Shh signaling components have not been identified yet.
A number of Hh inhibitors, namely Smo antagonist, Cyclopamine, Sulforaphane, Baicalein, Sangunarine, GANT61, Sonidegib, and PF-04449913 have been used for inhibition of cancer stem cells [77]. Meanwhile, activity of cancer stem cells has been shown to be affected by a number of mentioned ncRNAs. Therefore, combination of mentioned therapeutic modalities with ncRNA-targeted therapies might be regarded as effective options for eradication of cancer stem cells.
Since Shh-related ncRNAs have fundamental roles in the pathogenesis of human disorders, it is possible to down-regulate or up-regulate their expression in order to alter the pathological events in the course of disease evolution. In order to translate the basic science about the role of ncRNAs in the regulation of Shh pathway into clinical application, the following steps should be followed: (1) comprehensive assessment of expression of different classes of ncRNAs in clinical samples; (2) application of system biology methods for analysis of the acquired data; (3) understanding the complex network between different classes of ncRNAs and components of Shh pathway; (4) establishment of in vitro and in vivo models for assessment of the function of each module and (5) finding novel modalities for influencing the expression and activity of these modules.
Availability of data and materials
The analyzed data sets generated during the study are available from the corresponding author on reasonable request.
Abbreviations
- Shh:
-
Sonic Hedgehog
- Gli:
-
Glioma-associated oncogene
- Hh:
-
Hedgehog
- cAMP:
-
Cyclic adenosine 3', 5'-monophosphate
- Dhh:
-
Desert hedgehog
- Ihh:
-
Indian hedgehog
- SRF:
-
Serum response factor
- lncRNAs:
-
Long non-coding RNAs
- miRNAs:
-
MicroRNAs
- circRNAs:
-
Circular RNAs
- GEO:
-
Gene Expression Omnibus
- DM:
-
Diabetes mellitus
- OA:
-
Osteoarthritis
- GBM:
-
Glioblastoma Multiforme
- TNM:
-
Tumor-node-metastasis
References
Skoda AM, Simovic D, Karin V, Kardum V, Vranic S, Serman L. The role of the Hedgehog signaling pathway in cancer: a comprehensive review. Bosn J Basic Med Sci. 2018;18(1):8.
Nüsslein-Volhard C, Wieschaus E. Mutations affecting segment number and polarity in Drosophila. Nature. 1980;287(5785):795–801.
Choudhry Z, Rikani AA, Choudhry AM, Tariq S, Zakaria F, Asghar MW, et al. Sonic hedgehog signalling pathway: a complex network. Ann Neurosci. 2014;21(1):28–31.
Beachy PA, Karhadkar SS, Berman DM. Tissue repair and stem cell renewal in carcinogenesis. Nature. 2004;432(7015):324–31.
Toftgård R. Hedgehog signalling in cancer. Cell Mol Life Sci CMLS. 2000;57(12):1720–31.
Chiang C, Litingtung Y, Lee E, Young KE, Corden JL, Westphal H, et al. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature. 1996;383(6599):407–13.
Roelink H, Augsburger A, Heemskerk J, Korzh V, Norlin S, Altaba AR, et al. Floor plate and motor neuron induction by vhh-1, a vertebrate homolog of hedgehog expressed by the notochord. Cell. 1994;76(4):761–75.
Campbell V, Copland M. Hedgehog signaling in cancer stem cells: a focus on hematological cancers. Stem Cells Cloning Adv Appl. 2015;8:27.
Onodera S, Nakamura Y, Azuma T. Gorlin syndrome: recent advances in genetic testing and molecular and cellular biological research. Int J Mol Sci. 2020;21(20):7559.
Fattahi S, Nikbakhsh N, Ranaei M, Sabour D, Akhavan-Niaki H. Association of sonic hedgehog signaling pathway genes IHH, BOC, RAB23a and MIR195-5p, MIR509-3-5p, MIR6738-3p with gastric cancer stage. Sci Rep. 2021;11(1):1–12.
Zhang J, Fan J, Zeng X, Nie M, Luan J, Wang Y, et al. Hedgehog signaling in gastrointestinal carcinogenesis and the gastrointestinal tumor microenvironment. Acta Pharmaceutica Sinica B. 2021;11(3):609–20.
Carballo GB, Honorato JR, de Lopes GPF. A highlight on Sonic hedgehog pathway. Cell Commun Signal. 2018;16(1):1–15.
Statello L, Guo C-J, Chen L-L, Huarte M. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol. 2021;22(2):96–118.
Macfarlane LA, Murphy PR. MicroRNA: biogenesis, function and role in cancer. Curr Genomics. 2010;11(7):537–61.
Zhao X, Cai Y, Xu J. Circular RNAs: biogenesis, mechanism, and function in human cancers. Int J Mol Sci. 2019;20(16):3926.
Lin B-J, Zhu J-Y, Ye J, Lu S-D, Liao M-D, Meng X-C, et al. LncRNA-XIST promotes dermal papilla induced hair follicle regeneration by targeting miR-424 to activate hedgehog signaling. Cell Signal. 2020;72: 109623.
Zheng G, Tao Y, Yu W, Schwartz RJ. Brief report: SRF-dependent MiR-210 silences the sonic hedgehog signaling during cardiopoesis. Stem Cells. 2013;31(10):2279–85.
Li X, Liu H, Lv Y, Yu W, Liu X, Liu C. MiR-130a-5p/Foxa2 axis modulates fetal lung development in congenital diaphragmatic hernia by activating the Shh/Gli1 signaling pathway. Life Sci. 2020;241: 117166.
Richbourg HA, Hu DP, Xu Y, Barczak AJ, Marcucio RS. miR-199 family contributes to regulation of sonic hedgehog expression during craniofacial development. Dev Dyn. 2020;249(9):1062–76.
Liu T, Wang Z, Dong M, Wei J, Pan Y. MicroRNA-26a inhibits cell proliferation and invasion by targeting FAM98A in breast cancer. Oncol Lett. 2021;21(5):1–9.
Liu X, Zhao T, Bai X, Li M, Ren J, Wang M, et al. LOC101930370/MiR-1471 axis modulates the hedgehog signaling pathway in breast cancer. Cell Physiol Biochem. 2018;48(3):1139–50.
Zhou M, Hou Y, Yang G, Zhang H, Tu G, Du Y-e, et al. LncRNA-Hh strengthen cancer stem cells generation in twist-positive breast cancer via activation of hedgehog signaling pathway. Stem Cells. 2016;34(1):55–66.
Fu B, Liu W, Zhu C, Li P, Wang L, Pan L, et al. Circular RNA circBCBM1 promotes breast cancer brain metastasis by modulating miR-125a/BRD4 axis. Int J Biol Sci. 2021;17(12):3104.
Zhao D, Hou Y, Sun F, Han B, Li S. Effects of miR-132 on proliferation and apoptosis of pancreatic cancer cells via Hedgehog signaling pathway. Eur Rev Med Pharmacol Sci. 2019;23(5):1978–85.
Yin S, Du W, Wang F, Han B, Cui Y, Yang D, et al. MicroRNA-326 sensitizes human glioblastoma cells to curcumin via the SHH/GLI1 signaling pathway. Cancer Biol Ther. 2018;19(4):260–70.
Munoz JL, Rodriguez-Cruz V, Ramkissoon SH, Ligon KL, Greco SJ, Rameshwar P. Temozolomide resistance in glioblastoma occurs by miRNA-9-targeted PTCH1, independent of sonic hedgehog level. Oncotarget. 2015;6(2):1190.
Zhou H, Han L, Wang H, Wei J, Guo Z, Li Z. Chidamide inhibits glioma cells by increasing oxidative stress via the miRNA-338–5p regulation of Hedgehog signaling. Oxidative Med Cell Longevity. 2020;2020:1.
Wang Y-F, Yang H-Y, Shi X-Q, Wang Y. Upregulation of microRNA-129-5p inhibits cell invasion, migration and tumor angiogenesis by inhibiting ZIC2 via downregulation of the Hedgehog signaling pathway in cervical cancer. Cancer Biol Ther. 2018;19(12):1162–73.
Gao J, Zhao G, Li W, Zhang J, Che Y, Song M, et al. MiR-155 targets PTCH1 to mediate endothelial progenitor cell dysfunction caused by high glucose. Exp Cell Res. 2018;366(1):55–62.
Neumann JE, Wefers AK, Lambo S, Bianchi E, Bockstaller M, Dorostkar MM, et al. A mouse model for embryonal tumors with multilayered rosettes uncovers the therapeutic potential of Sonic-hedgehog inhibitors. Nat Med. 2017;23(10):1191–202.
Liu X, Li M, Peng Y, Hu X, Xu J, Zhu S, et al. miR-30c regulates proliferation, apoptosis and differentiation via the Shh signaling pathway in P19 cells. Exp Mol Med. 2016;48(7):e248-e.
Jiang Z, Cushing L, Ai X, Lü J. miR-326 is downstream of Sonic hedgehog signaling and regulates the expression of Gli2 and smoothened. Am J Respir Cell Mol Biol. 2014;51(2):273–83.
Liu X, Wang K, Du H. LncRNA SNHG6 regulating Hedgehog signaling pathway and affecting the biological function of gallbladder carcinoma cells through targeting miR-26b-5p. Eur Rev Med Pharmacol Sci. 2020;24:7598–611.
Bier A, Giladi N, Kronfeld N, Lee HK, Cazacu S, Finniss S, et al. MicroRNA-137 is downregulated in glioblastoma and inhibits the stemness of glioma stem cells by targeting RTVP-1. Oncotarget. 2013;4(5):665.
Li J, Zhang Q, Fan X, Mo W, Dai W, Feng J, et al. The long noncoding RNA TUG1 acts as a competing endogenous RNA to regulate the Hedgehog pathway by targeting miR-132 in hepatocellular carcinoma. Oncotarget. 2017;8(39):65932.
Ghorpade DS, Sinha AY, Holla S, Singh V, Balaji KN. NOD2-nitric oxide-responsive microRNA-146a activates Sonic hedgehog signaling to orchestrate inflammatory responses in murine model of inflammatory bowel disease. J Biol Chem. 2013;288(46):33037–48.
Yuan Z, Xiu C, Liu D, Zhou G, Yang H, Pei R, et al. Long noncoding RNA LINC-PINT regulates laryngeal carcinoma cell stemness and chemoresistance through miR-425-5p/PTCH1/SHH axis. J Cell Physiol. 2019;234(12):23111–22.
Yu F, Zheng Y, Hong W, Chen B, Dong P, Zheng J. MicroRNA-200a suppresses epithelial-to-mesenchymal transition in rat hepatic stellate cells via GLI family zinc finger 2. Mol Med Rep. 2015;12(6):8121–8.
Haque I, Kawsar HI, Motes H, Sharma M, Banerjee S, Banerjee SK, et al. Downregulation of miR-506-3p facilitates EGFR-TKI resistance through induction of sonic hedgehog signaling in non-small-cell lung cancer cell lines. Int J Mol Sci. 2020;21(23):9307.
Sun J, Jia J, Yuan W, Liu S, Wang W, Ge L, et al. LncRNA BLACAT1 accelerates non-small cell lung cancer through up-regulating the activation of Sonic Hedgehog Pathway. Front Oncol. 2021;11:708.
Liu X, Yin Z, Xu L, Liu H, Jiang L, Liu S, et al. Upregulation of LINC01426 promotes the progression and stemness in lung adenocarcinoma by enhancing the level of SHH protein to activate the hedgehog pathway. Cell Death Dis. 2021;12(2):1–13.
Liu M-w, Su M-x, Tang D-y, Hao L, Xun X-H, Huang Y-q. Ligustrazin increases lung cell autophagy and ameliorates paraquat-induced pulmonary fibrosis by inhibiting PI3K/Akt/mTOR and hedgehog signalling via increasing miR-193a expression. BMC Pulmonary Med. 2019;19(1):1–16.
Holla S, Stephen-Victor E, Prakhar P, Sharma M, Saha C, Udupa V, et al. Mycobacteria-responsive sonic hedgehog signaling mediates programmed death-ligand 1-and prostaglandin E2-induced regulatory T cell expansion. Sci Rep. 2016;6(1):1–14.
Miele E, Po A, Begalli F, Antonucci L, Mastronuzzi A, Marras CE, et al. β-arrestin1-mediated acetylation of Gli1 regulates Hedgehog/Gli signaling and modulates self-renewal of SHH medulloblastoma cancer stem cells. BMC Cancer. 2017;17(1):1–12.
Zhang Y, Wang T, Wang S, Xiong Y, Zhang R, Zhang X, et al. Nkx2-2as suppression contributes to the pathogenesis of sonic hedgehog medulloblastoma. Can Res. 2018;78(4):962–73.
Lang Z, Fan X, Lin H, Qiu L, Zhang J, Gao C. Silencing of SNHG6 alleviates hypoxia/reoxygenation-induced cardiomyocyte apoptosis by modulating miR-135a-5p/HIF1AN to activate Shh/Gli1 signalling pathway. J Pharm Pharmacol. 2021;73(1):22–31.
Gibert B, Delloye-Bourgeois C, Gattolliat C-H, Meurette O, Guernevel S, Fombonne J, et al. Regulation by miR181 family of the dependence receptor CDON tumor suppressive activity in neuroblastoma. JNCI: J Nat Cancer Institute. 2014. https://doi.org/10.1093/jnci/dju318.
Akhtar N, Makki MS, Haqqi TM. MicroRNA-602 and microRNA-608 regulate sonic hedgehog expression via target sites in the coding region in human chondrocytes. Arthritis Rheumatol. 2015;67(2):423–34.
Qing Y, Huang M, Cao Y, Du T, Song K. Effects of miRNA-342-3p in modulating Hedgehog signaling pathway of human umbilical cord mesenchymal stem cells by down-regulating Sufu. Oral Dis. 2019;25(4):1147–57.
Huang M, Qing Y, Shi Q, Cao Y, Song K. miR-342-3p elevates osteogenic differentiation of umbilical cord mesenchymal stem cells via inhibiting Sufu in vitro. Biochem Biophys Res Commun. 2017;491(3):571–7.
Nan K, Zhang Y, Zhang X, Li D, Zhao Y, Jing Z, et al. Exosomes from miRNA-378-modified adipose-derived stem cells prevent glucocorticoid-induced osteonecrosis of the femoral head by enhancing angiogenesis and osteogenesis via targeting miR-378 negatively regulated suppressor of fused (Sufu). Stem Cell Res Ther. 2021;12(1):1–13.
Kaur S, Gupta S, Chaudhary M, Khursheed MA, Mitra S, Kurup AJ, et al. let-7 MicroRNA-mediated regulation of Shh signaling and the gene regulatory network is essential for retina regeneration. Cell Rep. 2018;23(5):1409–23.
Zhao D, Cui Z. MicroRNA-361-3p regulates retinoblastoma cell proliferation and stemness by targeting hedgehog signaling. Exp Ther Med. 2019;17(2):1154–62.
Suh HN, Han HJ. Sonic hedgehog increases the skin wound-healing ability of mouse embryonic stem cells through the micro RNA 200 family. Br J Pharmacol. 2015;172(3):815–28.
Liu XS, Chopp M, Wang XL, Zhang L, Hozeska-Solgot A, Tang T, et al. MicroRNA-17-92 cluster mediates the proliferation and survival of neural progenitor cells after stroke. J Biol Chem. 2013;288(18):12478–88.
Zhang H, Tan M, Zhang J, Han X, Ma Y. Propofol inhibits thyroid cancer cell proliferation, migration, and invasion by suppressing SHH and PI3K/AKT signaling pathways via the miR-141–3p/BRD4 Axis. J Healthcare Eng. 2021;2021:1.
Li S, Zhang Y, Sun Y, Zhao H, Wang Y. Inhibition of microRNA-802-5p inhibits myocardial apoptosis after myocardial infarction via Sonic Hedgehog signaling pathway by targeting PTCH1. Eur Rev Med Pharmacol Sci. 2021;25(1):326–34.
Wang B, Xu J, Hu J, Hu M, Huang J, Zhu X. Effects of miR-153 on angiogenesis in MCAO rats through Shh signaling pathway. Eur Rev Med Pharmacol Sci. 2019;23(2):732–9.
Sun Q, Zeng J, Liu Y, Chen J, Zeng Q-C, Chen Y-Q, et al. microRNA-9 and-29a regulate the progression of diabetic peripheral neuropathy via ISL1-mediated sonic hedgehog signaling pathway. Aging (Albany NY). 2020;12(12):11446.
Hyun J, Wang S, Kim J, Kim GJ, Jung Y. MicroRNA125b-mediated Hedgehog signaling influences liver regeneration by chorionic plate-derived mesenchymal stem cells. Sci Rep. 2015;5(1):1–15.
Zhang Z, Li S, Cheng SY. The miR-183∼ 96∼ 182 cluster promotes tumorigenesis in a mouse model of medulloblastoma. J Biomed Res. 2013;27(6):486.
Lin J-C, Liu Z-G, Yu B, Zhang X-R. MicroRNA-874 targeting SUFU involves in osteoblast proliferation and differentiation in osteoporosis rats through the Hedgehog signaling pathway. Biochem Biophys Res Commun. 2018;506(1):194–203.
Wang J, Wang W, Zhai H. MicroRNA-124 enhances dopamine receptor expression and neuronal proliferation in mouse models of Parkinson’s disease via the hedgehog signaling pathway by targeting EDN2. NeuroImmunoModulation. 2019;26(4):174–87.
Hudish LI, Galati DF, Ravanelli AM, Pearson CG, Huang P, Appel B. miR-219 regulates neural progenitors by dampening apical Par protein-dependent Hedgehog signaling. Development. 2016;143(13):2292–304.
Wang X, Wang H, Zhang T, He M, Liang H, Wang H, et al. Inhibition of MicroRNA-195 alleviates neuropathic pain by targeting Patched1 and inhibiting SHH signaling pathway activation. Neurochem Res. 2019;44(7):1690–702.
Huang H, Fu S, Liu D. Detection and analysis of the hedgehog signaling pathway-related long non-coding RNA (lncRNA) expression profiles in keloid. Med Sci Monitor: Int Med J Exp Clin Res. 2018;24:9032–44.
Yoo YA, Kang MH, Lee HJ, Kim B-h, Park JK, Kim HK, et al. Sonic hedgehog pathway promotes metastasis and lymphangiogenesis via activation of Akt, EMT, and MMP-9 pathway in gastric cancer. Cancer Res. 2011;71(22):7061–70.
Xu X, Zhou Y, Xie C, Wei S-m, Gan H, He S, et al. Genome-wide screening reveals an EMT molecular network mediated by Sonic hedgehog-Gli1 signaling in pancreatic cancer cells. PLoS ONE. 2012. https://doi.org/10.1371/journal.pone.0043119.
Islam S, Mokhtari R, Noman A, Uddin M, Rahman M, Azadi M, et al. Sonic hedgehog (Shh) signaling promotes tumorigenicity and stemness via activation of epithelial-to-mesenchymal transition (EMT) in bladder cancer. Mol Carcinog. 2016;55(5):537–51.
Song J, Ge Y, Sun X, Guan Q, Gong S, Wei M, et al. Noncoding RNAs related to the hedgehog pathway in cancer: clinical implications and future perspectives. Mol Cancer. 2022;21(1):115.
Wang YZ. The limited role of most circRNAs as sponges for microRNAs. Cell Mol Life Sci: CMLS. 2022;79(6):299.
Wang Y. Comment on "Circular RNAs and their emerging roles as diagnostic and prognostic biomarkers in ovarian cancer. Cancer Lett. 2020;2020(473):139–47.
Wang YZ. Letter to the Editor: the limited functions and mechanisms of CircASAP1 as a Sponge of Onco-microRNAs. Hepatology (Baltimore, Md). 2021;73(6):2610.
Wang YZ. Letter to the Editor: the limited function of most circRNAs as a microRNA sponge. Hepatology. 2022. https://doi.org/10.1002/hep.32657.
Wang YZ. Comment on "Regulation of nuclear Factor-KappaB (NF-κB) signaling pathway by non-coding RNAs in cancer: Inhibiting or promoting carcinogenesis? [Cancer Lett. 509 (2021 May 2) 63-80]. Cancer Lett. 2021;516:99–100.
Wang Y. Letter to the editor: issues of circular RNAs as MicroRNA Sponges. Hepatology (Baltimore, Md). 2020;72(1):365.
Sari IN, Phi LTH, Jun N, Wijaya YT, Lee S, Kwon HY. Hedgehog signaling in cancer: a prospective therapeutic target for eradicating cancer stem cells. Cells. 2018;7(11):208.
Acknowledgements
This study was financially supported by Grant from Medical School of Shahid Beheshti University of Medical Sciences.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
SGF wrote the manuscript and revised it. MT supervised and designed the study. TK, MS and BMH collected the data and designed the figures and tables. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. Informed consent forms were obtained from all study participants. The study protocol was approved by the ethical committee of Shahid Beheshti University of Medical Sciences. All methods were performed in accordance with the relevant guidelines and regulations.
Consent of publication
Not applicable.
Competing interests
The authors declare they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ghafouri-Fard, S., Khoshbakht, T., Hussen, B.M. et al. Emerging role of non-coding RNAs in the regulation of Sonic Hedgehog signaling pathway. Cancer Cell Int 22, 282 (2022). https://doi.org/10.1186/s12935-022-02702-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-022-02702-y