
Abstract
Background
Escherichia coli heat labile toxin B subunit (LTB) is one of the most popular oral vaccine adjuvants and intestine adsorption enhancers. It is often expressed as a fusion partner with target antigens to enhance their immunogenicity as well as gut absorbability. However, high expression levels of a fusion protein are critical to the outcome of immunization experiments and the success of subsequent vaccine development efforts. In order to improve the expression and functional assembly of LTB-fusion proteins using Saccharomyces cerevisiae, we compared their expression under culture conditions at a sub-physiological temperature 20 °C with their expression under a standard 30 °C.
Results
The assembled expression of LTB-EDIII2 (LTB fused to the envelope domain III (EDIII) of Dengue virus serotype 2), which was expressed at the level of 20 µg/L in our previous study, was higher when the expression temperature was 20 °C as opposed to 30 °C. We also tested whether the expression and functional assembly of a difficult-to-express LTB fusion protein could be increased. The assembled expression of the difficult-to-express LTB-VP1 fusion protein (LTB fused to VP1 antigen of Foot-and-Mouth Disease Virus) dramatically increased, although the total amount of expressed protein was still lower than that of LTB-EDIII2. Slight but significant increase in the expression of well-known reporter protein eGFP, which has previously been shown to be increased by cultivation at 20 °C, was also observed in our expression system. As no significant changes in corresponding transcripts levels and cell growth were observed between 20 °C and 30 °C, we infer that translation and post-translational assembly are responsible for these enhancements.
Conclusions
The effects of lowering the expression temperature from 30 °C to 20 °C on protein expression and folding levels in S. cerevisiae, using several proteins as models, are reported. When heterologous proteins are expressed at 20 °C, a greater amount of (specially, more assembled) functional proteins accumulated than at 30 °C. Although further studies are required to understand the molecular mechanisms, our results suggest that lowering the expression temperature is a convenient strategy for improving the expression of relatively complexly structured and difficult-to-express proteins in S. cerevisiae.
Similar content being viewed by others

Background
Mucosal vaccines, which are typically delivered through nasal or oral routes, offer an attractive solution to some of the challenges of mass vaccination programs due to their cost-effectiveness, safety, and high rate of public acceptance [1, 2]. Despite these benefits, far fewer mucosal than injectable vaccines have been approved. One significant obstacle to mucosal vaccine development is their tendency to induce low immunogenicity, sometimes even promoting the target antigens to develop tolerance. This is the result of difficulties in controlling the quantity of antigens delivered to the immune system through the mucosal interfaces. Thus, specifically in the context of mucosal vaccines, the choice of adjuvants plays a much more crucial role in determining efficacy and protective effect [2]. Among the most potent and widely used mucosal adjuvants are the Vibrio cholera and Escherichia coli toxins and their mutant forms (dmLT and mmCT, respectively) [2,3,4,5]. These adjuvants are unique in that they consistently induce a very strong mucosal immune response specific to the target antigens as well as a systematic response when delivered orally or intranasally [5].
Baker’s yeast, Saccharomyces cerevisiae, is the most popular eukaryotic host for the expression of foreign proteins [6, 7], because as a single cell microorganism, S. cerevisiae combines the advantages of simple prokaryotic systems—including high expression level, ease of genetic manipulation and scale-up, fully developed mass culturing—with those of eukaryotic post-translational modifications and secretion. Moreover, S. cerevisiae is a generally recognized as safe (GRAS) organism, to make its expressed product and processing more applicable without further consideration. In addition, its high-quality protein content and high vitamin levels distinguish S. cerevisiae as a favorable single cell organism useful for live and oral administration of pharmaceutical and feed products [8, 9]. Furthermore, the strong adjuvant properties of yeast derivatives render it an attractive heterologous expression system for vaccine production and development [10,11,12,13,14,15,16,17]. A well-balanced immune response to dengue was observed when a yeast expression system was used to produce dengue virus epitope proteins [18].
One common oral vaccine development strategy is to express target antigens intracellularly fused with the E. coli heat labile toxin B subunit (LTB) in the yeast [13, 16]. This fusion protein is capable of inducing strong systematic as well as mucosal immune responses against the target antigens. However, if the availability of the antigen is limited, such as in a vaccine in which the whole yeast cell is used instead of the protein extract, the strength of the immune responses is compromised [10]. To overcome this problem, it is suggested improving the expression levels of the target antigen. This goal could be achieved through systematic development of various recombinant technologies including codon optimization [19], strong promoters and terminators [20,21,22], and multi-copy expression vectors [23]. In addition, some have suggested altering cultural practices such as acting to change the expression temperature from the physiological temperature [24, 25].
Our previous studies, which were focused on the development of an oral vaccine, used S. cerevisiae to express LTB fused with a Dengue antigen [13] that induced immune responses in mice. We found that oral vaccine efficacy might be further improved by increasing LTB-fusion protein expression. Lowering expression temperatures is an established and popular approach to improve the production of functional heterologous proteins in Escherichia coli [26], as heterologous proteins tend to aggregate at high temperatures [27] but the induction temperature does not seem to improve overall expression level. In animal cells such as CHO, lowering expression temperature also has a positive effect on target protein expression [28,29,30]. Although there is a general consensus, supported by some reports, concerning improved heterologous expression in P. pastoris [31,32,33], Yarrrowia lipolitica [34], Kluyveromyces lactis [35, 36] and S. cerevisiae [24] by temperature reduction, these lacked a systematic approach that would show that this is a widespread phenomenon with practical applicability. As we explored potential means of improving antigen expression using S. cerevisiae, we observed that expression levels and the functional assembly of target proteins increased significantly when expression temperature was lowered from 30 °C to 20 °C. In an attempt to determine the underlying mechanisms, we examined the effect of lowering the expression temperature on three different constructs which collectively covered a wide range of expression levels. We tested our expression system for eGFP as a model reporter protein with a strong expression level [24] in S. cerevisiae whose temperature was lowered to 20 °C. We also tested our previous LTB-EDIII2 fusion protein (LTB fused to envelope domain III (EDIII) of Dengue virus serotype 2), which has a lower expression level (20 g/L) than LTB-scEDIII (LTB fused to the synthetic consensus sequence of EDIII of all four serotypes of Dengue virus) (4 mg/L), as a low expression target [13], and LTB-VP1 (LTB fused to VP1 antigen Foot-and-Mouth Disease Virus) as a difficult-to-express target. Our results revealed that the benefits of lowering expression temperature do not stop at protein assembly but extend to total expression level. This report also highlights a convenient strategy for the modulation of heterologous protein expression in S. cerevisiae, as well as the need to explore the underlying mechanisms so that its potential can be fully exploited.
Results
Expression analyses of eGFP
Since FACS analysis can be easily applied to measure cell fluorescence without further treatment, the expression levels of reporter protein eGFP were compared among transformants (Fig. 1A) by FACS analysis. Twenty transformants were selected from the ura− selective media and then analyzed by FACS. There was little variation in levels of eGFP expression among tested transformants. Among the examined transformants, we selected one for further analysis by FACS and Western blot analysis of the temporal expression of eGFP at 20 °C and 30 °C. As seen in Fig. 1A, the FACS showed that eGFP expression in the sample cultivated at 20 °C was slightly but noticeably stronger on days 1 and 3 than in those cultivated at 30 °C. The expression of eGFP in this culture eventually decreased, and ultimately no significant difference between the 20 °C and 30 °C cultures was observed 5 days after cultivation. Western blot results were consistent with those of the FACS analysis (Fig. 1B).

Expression analysis of yeast codon optimized eGFP under two different expression conditions using FACS (A) and Western blot analysis (B and C). A FACS analysis of expressed eGFP. Inset: yeast cells cultured at 20 °C (left) and 30 °C condition (right) show eGFP expression under UV light. B Western blot of yeast expressing eGFP in 20 °C and 30 °C conditions. Lane 1: purified eGFP; lane 2: S. cerevisiae 2805 strain; lanes 3–5: protein preparation from eGFP-expressing yeast cells harvested after 1, 3, and 5 days, respectively, at 20 °C. Lanes 6–8: protein preparation from eGFP-expressing yeast cells harvested after 1, 3, and 5 days, respectively, at 30 °C. C SDS-PAGE twin gel showing that a similar amount of protein was loaded on each lane
Expression analyses of LTB-EDIII2
We tested LTB-EDIII2 as an exemplar “low expression” LTB-fusion protein. We choose LTB-EDIII2 as its expression was significantly lower than that of a similar LTB-scEDIII fusion protein; approximately, 200 times less LTB-EDIII2 was expressed than LTB-scEDIII (20 g/L vs. 4.0 mg/L). Northern blot and Western blot screenings of all candidate transformants showed that the target proteins were indeed expressed in all of the transformants. From the initial Northern blot (Fig. 2A) screening of LTB-EDIII2 expression, three colonies (#1, #8 and #9) showing intensive hybridizing bands were selected for further temporal expression analysis at the transcription level. At 30 °C, the expression patterns of the three-selected LTB-EDIII2 transformants peaked to 1—3 days after cultivation (Fig. 2B), a result that is consistent with our previous studies [13].

A Northern blot analysis of 12 transformants. 30 µg of total RNA was loaded on each lane. LTB-EDIII2 was used as the probe to detect LTB-EDIII2 expression. Lanes 1–12: LTB-EDIII2 transformants were randomly selected and numbered from 1 to 12. B Temporal expression of LTB-EDIII2 transcript in three selected transformants (transformants #1, #8, and #9 in panel A). Lane numbers indicate the number of days after cultivation at 30 °C and strains are indicated at the top of the lanes. Glyceraldehyde-3-phosphate dehydrogenase gene (GPD) was used as an internal control and bands representing rRNAs in ethidium bromide strained-gel (rRNA) are shown to indicate an equal amount of RNA loaded on each lane
We then selected transformant #8 as the representative transformant used a Northern blot analysis to compare the temporal expression patterns of LTB-EDIII2 at 20 °C and 30 °C. As shown Fig. 3A, no considerable difference in transcript accumulation was observed until 3 days after cultivation at either 20–30 °C. Although a dramatic decrease of in the band intensity of the LTB-EDIII2 transcript was observed at 5 days after cultivation, it was less so once it was normalized to the internal control gene (GPD) expression. Only a slight but significant decrease in band intensity was estimated after the normalization. We then checked the quantitative gene expression levels using real-time RT PCR, and the results indicated that transcript accumulation at 30 °C was stronger than that at 20 °C over the surveyed period. This suggests that transcription of the LTB-EDIII2 gene may not be a major factor in the production of the target protein and, if any, 30 °C is a more favorable condition for transcription than 20 °C (Fig. 3B).

Northern blot analysis (A) and Quantitative real time RT-PCR (qRT-PCR) analysis (B) of LTB-EDIII2 in a selected transformant (#8) under 20 °C and 30 °C conditions. A 20 µg total RNA was loaded on each lane. RNA preparations from cells harvested at day 1, 3, and 5 days after cultivation at 20 °C (Lanes 1–3, respectively) and 30 °C (Lanes 4–6, respectively). GPD was used as an internal control and rRNAs are shown to indicate equal amount of RNA loaded on each lane. B qRT-PCR results of changes in expression of LTB-EDIII2 under 20 °C and 30 °C conditions are shown. Error bars indicate standard deviation based on three independent measurements. ** indicates statistically significant difference between two groups, according to t-test at p = 0.01
To examine the production of corresponding LTB-EDIII2 protein, three selected transformants were subjected to Western blot analysis under both non-denaturing (non-boiling) and denaturing (boiling) conditions. As shown in Fig. 5, LTB-EDIII2 protein expression after 3 days was dramatically different at 20 °C and 30 °C, i.e., the LTB-EDIII2 protein from all three strains was expressed much higher levels at 20 °C than at 30 °C (Fig. 4A). Band intensity was twice or three times as high at 20 °C than at 30 °C (Fig. 4A). Interestingly, the difference in the level of expression between 20 °C and 30 °C under the denatured condition (Fig. 4A), which designed to detect the monomeric LTB-EDIII2, was noticeably high, but still not as high as the difference observed in the non-denatured condition (Fig. 4B), which we tested to detect the assembled oligomeric forms of LTB fusion protein. At least 8 times more assembled LTB-EDIII2 was detected at 20 °C than at 30 °C (Fig. 4B). These results suggest that cultivation at 20 °C not only increased LTB-EDIII2 monomer production but also greatly facilitated the assembly of the expressed monomeric subunit.

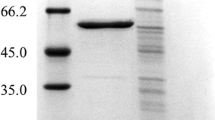
Western blot analysis of three selected LTB-EDIII2 transformants. A LTB-EDIII2 detection under denatured condition. B LTB-EDIII2 detection under non-denatured condition. Densitometry analysis of cross-reacting LTB-EDIII2 bands in the corresponding blot is shown on the right panel. C SDS-PAGE gel shows that an equal amount of protein was loaded on each lane. Lane 1: purified E. coli-expressed EDIII2 as a positive control; Lane 2: a mock transformant as a negative control; Lanes 3, 5 and 7: LTB-EDIII2 transformants #1, #8, and #9 cultured at 20 °C; Lanes 4, 6 and 8: LTB-EDIII2 transformants #1, #8, and #9 cultured at 30 °C. Proteins were prepared from cells cultured for 3 days after the inoculation to the expression medium
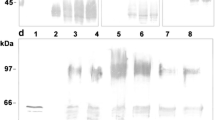
Temporal expression of LTB-EDIII2 in transformant #8 was performed at days 1, 3 and 5 using Western blot analysis (Fig. 5, Additional file 1: Fig. S1) and GM1 ELISA (Fig. 6). The Western analysis indicated that differences in LTB-EDIII2 expression were hardly dependent on culture period. However, significantly more assembled LTB-EDIII2 was produced at 20 °C than at 30 °C (Fig. 5). GM1 ELISA, which quantitatively measures the amount of pentameric LTB-EDIII2, indicated that the yield of the assembled pentameric form of LTB-EDIII2 was 2.5 greater at 20 °C than at 30 °C (Fig. 6). As no significant changes to transcript levels were observed between the 20 °C and 30 °C tests, these results suggest that the improved expression of the target proteins is not be due to transcriptional factors, but rather translational and/or post-translational levels.

Western blot analysis of temporal expression of LTB-EDIII2 from transformant #8. A LTB-EDIII2 expression was resolved under non-denaturing condition at 20 °C and 30 °C. B SDS-PAGE gel showed that an equal amount of protein was loaded on each lane. Lane 1: purified E. coli-expressed LTB as a positive control; Lane 2: a mock transformant cultured for 3 days as a negative control. Proteins were prepared from transformant #8, and cultured for 1, 3, 5 days after inoculation to the expression medium at 20 °C (lanes 3, 5, and 7, respectively) and 30 °C (lanes 4, 6, and 8, respectively)

GM1-ganglioside binding assay (GM1 ELISA) of LTB-EDIII2. The ELISA was conducted by coating 96-wells with GM1 monosialoganglioside as the receptor molecule. Y-axis indicates the percentage of pentameric LTB-EDIII2 in the sample preparations. The coated plates were incubated with the protein preparation of transformant #8 and then cross-reacted with anti-LTB antiserum
Expression analyses of LTB-VP1
As an example of a “difficult-to-express” LTB-fusion protein, we tested the expression of the LTB-VP1. In our attempt to express epitopes of Foot-and-Mouth Disease Virus (FMDV), we found that LTB-VP1 as well as VP1 (comprising aa 725—935 of FMDV (GenBank accession No: AY593823.1)) was difficult-to-express, whereas the remainder of the viral structural proteins (e.g., VP0, VP2, VP3, and VP4) were relatively easier to express (data not shown). We then tested whether the LTB fusion protein (LTB-VP1) could be quantified using GM1 ELISA. The LTB-VP1 transformants were also subjected to a procedure similar to those described above applicable to LTB-EDIII2. Three transformants were selected after a Northern blot analysis, after which one among the three was subjected to temporal expression analysis by additional Northern blot and real-time RT PCR (Additional file 2: Fig. S2). Protein expression was also analyzed by Western blot and GM1 ELISA. As predicted, Western blot analysis using anti-LTB antibody failed to detect discernable bands associated with any monomeric subunit (denatured condition) or multimeric assembly (non-denatured condition) of LTB-VP1 (data not shown). With this indication that levels of LTB-VP1 expression were so low as to be undetectable by Western blot analysis, we performed a more sensitive GM1-ELISA assay. As shown in Fig. 7, a very low GM1-ELISA signal was observed from samples cultured at 30 °C even at the lowest (1/16) dilutions. However, although not as strong as those observed from LTB-EDIII2, the signals indicating assembled LTB-VP1 were so distinctive in the samples cultured at 20 °C that a colorimetric outcome of reaction mixture rendered the change apparent to the naked eye. Based on GM1-ELISA, we estimated the amount of assembled pentameric forms of LTB-VP1 at 20 °C to be 7.8 g/L, which is at least 3 times more than the amount of assembled LTB-VP1.

GM1-ganglioside binding assay (GM1 ELISA) of LTB-VP1. A ELISA was conducted as described earlier. The plates coated with GM1 monosialoganglioside were incubated with the protein preparation of two selected LTB-VP1 transformants and then cross-reacted with anti-LTB antiserum. B Plates are shown to demonstrate colorimetric changes in the well, in contrast with samples from the 30 °C cultures. PC and NC indicate that the E. coli-expressed LTB as a positive control and a mock transformant as a negative control, respectively. Well number 1 indicates the sample of protein preparation without dilution and well numbers (2—8) indicate samples with twofold dilution of the original protein preparation
Discussion
S. cerevisiae has been one of the most popular heterologous expression hosts in biotechnology, and it is increasingly employed in oral vaccine development [10,11,12,13,14,15,16, 37,38,39,40,41]. Our previous study [13] showed that the expression of LTB fused with a synthetic Dengue tetravalent antigen using S. cerevisiae was potentially useful as an oral vaccine against Dengue viruses. However, the intensity of the immune response elicited seem to depend on the bioavailability of the target antigen, i.e. the amount of the target antigen available for absorption by gut mucosal lymphoid organs such as Peyer’s patches. In this paper, to increase the bioavailability of the antigen, we prepared and partially concentrated cell free extract instead of using the entire cell for vaccination. Unfortunately, this step requires extra preparation in terms of effort and equipment, runs the risk of contaminating the oral vaccines, and is therefore undesirable in the context of oral vaccine development. If instead whole yeast cells are used, the expression level of the target antigen must be increased. Although the production of recombinant protein in S. cerevisiae is straightforward, improving the yield of a heterologous protein using this fungus is a more complicated process. While various strategies for improving heterologous expression in S. cerevisiae have been described, such as the selection of strong promoters and enhancers [20, 21], changing terminators [22, 42, 43], engineering protein translocation by overexpressing protein folding enhancing genes [44], or codon optimization [19], each necessitates the extra effort of redesigning the expression systems and adapting the new systems to current laboratory conditions. As an alternative, in one intriguing work expression as well as secretion of GFP in S. cerevisiae was observed to improve by lowering the expression temperature to 20 °C [24]. It remained unclear, however, whether this phenomenon would be universally applicable to all heterologous proteins expressed in the yeast, or only some of them.
In this work, we surveyed three recombinant proteins expressed in the S. cerevisiae 2805 strain. We observed that the yields of all three proteins were increased by lowering the temperature to 20 °C from the yeast physiological temperature at 30 °C during the expression phase [45]. Most importantly, the amount of assembled multimeric LTB-fusion protein increased dramatically, and was particularly more pronounced in the proteins with a multimeric structure or otherwise low expression levels, (e.g., LTB-EDIII2 and LTB-VP1, respectively) and those that are less conspicuous in the protein but possess a relatively simple structure and expressed well (e.g., eGFP). Northern blot and real-time RT PCR of LTB-EDIII2 showed that the increase in the production of the target protein was not due to changes in transcription.
The effects of various types of stress on recombinant protein production in yeast were reviewed [46]. The stress factors that may affect the expression of recombinant proteins can be divided into two groups: metabolic stresses arising from interference with cellular processes and environmental stresses due to culture conditions. The most notable cellular response to metabolic stress is the unfolded protein response, which implicates the limited processing capacity of the endoplasmic reticulum (ER) as well as various quality control mechanisms in degrading unfolded, misfolded, and aggregated heterologous proteins. The use of multicopy expression plasmids may induce extra-metabolic stress on cells and may offset their transcriptional advantage, resulting in low expression levels. In terms of environmental stresses, reducing expression temperature may inhibit protease degradation, leading to an increase in the amount of heterologous proteins [46].
Reducing the expression temperature in E. coli has been widely exploited as a solution to the problem of too many inclusion bodies vs too little soluble expression [47,48,49]. Reduction of the post-induction temperature has been thought to reduce the translational rate, thereby enhancing solubility. High temperatures are also thought to be conducive to hydrophobic aggregation [27]. Some previous studies have also reported benefits associated with expressing heterologous proteins in P. pastoris at low temperatures and proposed that the improvement in the target proteins was due to the enhancement of the protein folding pathway and reduced cell death [50,51,52]. However, Hohenblum et al. [53] showed that cell viability was not affected by temperature. Similar observations have also been reported in animal cells [28,29,30, 54], suggesting that the phenomenon is a universal one. To date, however, attempts to understand the underlying mechanisms involving transcriptomic [55] and proteomic [56] surveys of S. cerevisiae have not yielded a satisfactory explanation for the observed phenomenon. A similar conclusion was also reached by Alexandra Graf [57].
We observed that when E. coli, yeast, or animal cells are forced to express heterologous protein at sub-optimal temperatures, cells grew more slowly than they would have at the optimal temperatures. When the temperature is decreased further, such as to 15 °C or below 10 °C, cell mass decreases dramatically or cells may even stop dividing [55]. However, in the case of yeast, its growth rate at 20 °C is slightly, but not dramatically, lower (lnOD 0.36 at 18 h) from the early growth phase than at 30 °C (lnOD 0.76 at 18 h) (Fig. 8). Moreover, our cultivation procedure for yeast expression was designed to inoculate a large quantity of exponentially growing cells within the nutrient rich media, i.e. successively cultured cells (approximately 105cells per mL) in a total volume of 40 mL YEPD media. This system was designed to support active cell growth and gene expression at different temperatures (20 °C or 30 °C). This inoculation strategy allowed the culture system to reach the stationary phase less than 36 h after incubation regardless of whether the culture was maintained at 20 °C or 30 °C (Fig. 8). Balancing between fermentation and respiration is essential to achieving adequate protein production levels [58, 59]. Accordingly, for the process tested in this paper, 20 °C seems to be the ideal temperature that maximizes protein expression without needlessly compromising cell growth. In addition, the stability of the introduced plasmids in the yeast was measured at the end of the cultivation period by comparing the number of colony forming units (CFUs) per ura− selective and non-selective plate [16, 60]. The plasmid stability of all transformants allowed more than 82% of the plated cells to maintain the plasmids, and we observed no difference in stability between 20 °C and 30 °C samples. These results suggested that plasmid stability should be excluded as potential explanations for the difference in expression levels.

Growth curve of transformants. Cell growth was monitored by measuring optical density at 600 nm using a spectrophotometer (OD600). The growth curves are shown as representative profiles based on duplicated experiments with three replications
Based on the fact that more functional proteins were produced at the sub-optimal temperature, we hypothesize that protein folding plays an important part in the improved expression. Since GPD is a strong constitutive promoter, the target genes were likely overexpressed at the transcriptional level, which was an observation that is consistent with the results of our Northern blot analysis. This may result in the protein folding system becoming overwhelmed. Likewise, the overexpression of proteins involved in protein folding improves the yield of heterologous proteins [44, 61]. In short, although further studies are required for verification, it is likely that the increase in functional protein amounts observed in cultures at low temperatures is a consequence of both the translation machinery as well as the protein folding system.
Conclusions
In this study we reported the effects of a lowered expression temperature (from 30 °C to 20 °C) on protein expression levels and protein folding levels in S. cerevisiae, using several proteins as models. When heterologous proteins were expressed at 20 °C, which is a sub-physiological temperature of S. cerevisiae, more proteins were produced and more assembled functional proteins were accumulated, than when these same proteins were expressed at 30 °C. Although further studies will be required before the molecular mechanisms implicated by this result is fully understood, our results suggest that cell integrity, plasmid stability, and enhanced protein folding play a role. To our knowledge, this is the first report to test this novel strategy for improving heterologous protein production in S. cerevisiae.
Methods
Strains and culture conditions
Escherichia coli Top10 was used throughout the various stages described herein for cloning. BL21 (DE3)-RIPL and BL21 (DE3) were employed to produce recombinant antigens, which were used as standards in Western and ELISA analyses. All E. coli strains were maintained in LB broth or on LB agar supplemented with appropriate antibiotics. Saccharomyces cerevisiae 2805 strain (MATα pep4::HIS3 prb 1-δ Can1 GAL2 his3 ura3-52) was used as a recipient to express all of the LTB fusion proteins mentioned in this report. S. cerevisiae 2805 was maintained in complete medium YEPD (Yeast Extract Peptone Dextrose) before transformation, and subsequently in Ura dropout medium (0.56% Yeast Nitrogen Base with ammonium sulfate, 0.76% KCl, 2% Glucose and 0.14% Yeast synthetic dropout, all in weight/volume unit; and supplemented with 20 mg/L of tryptophan, histidine, leucine and adenine hemisulfate).
Both E. coli and S. cerevisiae were transformed by chemical methods described previously [18]. For yeast expression, a single colony was inoculated into 5 ml of Ura dropout liquid medium and cultured for 48 h at 200 rpm and 30 °C in a shaking incubator. Subsequently, 250 µL of this culture was transferred into 5 ml YEPD medium and cultured for another 16 h under these same conditions. This culture was then inoculated into 40 ml of freshly prepared YEPD medium and grown at either 30 °C or 20 °C at 200 rpm. Biomasses were collected at days 1, 3 and 5 post-inoculation, weighted and stored at − 80 °C for expression analyses.
Plasmid constructions
Yeast codon optimized eGFP (GenBank accession No: ON036474) was synthesized. BamHI and SalI restriction enzyme recognition sites were added before the start codon and after the stop codon, respectively. The resulting fusion gene was cloned in a pGEM T-easy vector for sequence verification. Subsequently the gene was released from the vector by BamHI/SalI and cloned into the pYEGPD-TER vector, which contained a constitutive GPD (glyceraldehyde-3-phosphate dehydrogenase) promoter and a Gal7 terminator (Fig. 9A). Using overlap-extension PCR, genes encoding EDIII2 and VP1 (GenBank accession No: KT452797.1 and ON036475, respectively) were cloned in-frame with the LTB encoding gene. GPGP (GlyProGlyPro) flexible linker [16] and GS3 (Gly4Ser-Gly4Ser-Gly4Ser) flexible linker [62] were inserted between these genes to allow for independent folding of EDIII2 and VP1, respectively. BamHI and SalI restriction enzyme recognition sites were then added before the start codon and after the stop codon, respectively. The resulting fusion genes were cloned using the pGEM T-easy vector for sequence verification. Subsequently, these genes were released from the vector by BamHI/SalI and cloned into the pYEGPD-TER vector (Fig. 9B). The recombinant pYEGPD-TER vectors were transformed into the S. cerevisiae 2805 strain using the lithium acetate method [63]. The primers used for each expression vector are listed in Additional file 3: Table S1.

Schematic representations of expression vectors described in this study. All of the expression cassettes were cloned into the pYEGPD-TER (A) vector [13] using BamHI and SalI restriction enzymes
Northern blot analysis
After screening for positive transformants by colony PCR and E. coli back transformation [18], twelve random transformants were selected for expression analysis. The expression of these transformants at the transcription level was compared using Northern blot to identify those that were most expressed. The three transformants with the highest expression level were then selected for temporal expression analysis. To determine which transformants were most expressed, yeast cells were grown at 30 °C and harvested at day 3 post-inoculation. For temporal expression analysis, yeast cells were grown at either 30 °C or 20 °C and the cells in 40 ml YEPD were harvested at days 1, 3, and 5 post-inoculation, washed twice with dH2O, and then stored at − 80 °C for RNA extraction. Total RNA was extracted as described previously [64]. RNA samples were quantified using a spectrophotometer as 30 µg of each sample was placed on a 1.2% denatured agarose gel. The gels were then blotted onto Amersham Hybond™ membrane, crossed linked with UV, and hybridized in modified Church buffer. Detecting probes were prepared by isothermal (37 °C) amplification of the corresponding genes using random primers and cytosine labelled with 32P isotope.
Real time RT-PCR
Quantitative real time RT-PCR was conducted as previously described to evaluate the expression levels of the target genes [65]. Each expression level was evaluated in triplicate for each transcript, with at least two independent preparations of the same RNA sample. Transcript levels relative to the amount of glyceraldehyde-3-phosphate dehydrogenase (GPD) of S. cerevisiae were used as an internal control. The primer pairs used for each target gene are listed in Additional file 3: Table S1. Statistical analyses were performed by t-test at p = 0.01 using SPSS software (IBM Corp., Armonk, NY). Different letters indicate significant differences between strains.
Western blot analysis
Yeast protein was extracted consistent with methods previously described [18]. 100 or 200 µg of total soluble protein from each sample was mixed with 6 × SDS-PAGE loading buffer and either boiled (denatured) or not boiled (non-denatured) before being loaded onto 10% SDS-PAGE gel.
After gel separation, each gel was then blotted onto nitrocellulose membrane (Hybond™ Cytiva Life Sciences), and the target proteins were detected with specific primary antibodies (either rabbit anti-LTB or mouse anti-dengue envelope domain III) and AP-conjugated secondary antibodies. Western band intensities, which reflect the relative amount of target proteins in the samples, were determined using the Fiji ImageJ software package [66].
GM1 ELISA analyses
GM1 ELISA was used to quantify the amount of pentameric assembled LTB fused antigens. In brief, 96-well microtiter plates were coated with 0.3 µg GM1 (Sigma G-7641) and incubated overnight. The next day, the plates were washed 3 times and blocked with 1% BSA for 1 h at 37 °C. LTB standard and yeast protein samples were added and serially two-fold diluted. The plates were incubated for 2 h at 37°C. After washing, the primary antibody (rabbit anti-LTB, 1:5000 dilution) was added and the plates were incubated for another 2 h at 37 °C. The secondary antibody was then added and the plates were left standing at 37 °C for an additional 2 h. Finally, after three washings, the plates were developed with AP substrate solution (prepared from AP substrate tablets—Sigma S0942-100TAB) and optical density was determined by their absorbance at 405 nm (Multiskan GO microplate reader ThermoFisher Scientific). Statistical analyses were performed by t-test at p = 0.01 using SPSS software (IBM Corp., Armonk, NY).
FACS
Yeast cells expressing eGFP were harvested, washed twice with PBS, and subjected to FACS analysis using a BD FACS CANTOII flow cytometer with a 488 nm excitation laser line. The data was subsequently analyzed using FlowJo V10 software to determine average fluorescent intensities. Statistical analyses were performed by t-test at p = 0.01 using SPSS software (IBM Corp., Armonk, NY).
Availability of data and materials
All data for this study are included in this published article.
Abbreviations
- LTB:
-
Escherichia coli Heat labile toxin B subunit
- LTB-EDIII2 :
-
LTB fused to the envelope domain III (EDIII) of Dengue virus serotype 2
- LTB-VP1:
-
LTB fused to VP1 antigen of Foot-and-Mouth Disease Virus
- LTB-scEDIII:
-
LTB fused to the synthetic consensus sequence of envelope domain III of all four serotypes of Dengue virus
- FMDV:
-
Foot-and-Mouth Disease Virus
- GPD:
-
Glyceraldehyde-3-phosphate dehydrogenase
References
Singh M, O’Hagan D. Advances in vaccine adjuvants. Nat Biotechnol. 1999;17:1075–81.
Lycke N. Recent progress in mucosal vaccine development: potential and limitations. Nat Rev Immunol. 2012;12:592–605.
Firdaus FZ, Skwarczynski M, Toth I. Developments in vaccine adjuvants. Methods Mol Biol. 2022;2412:145–78.
Van der Weken H, Cox E, Devriendt B. Advances in oral subunit vaccine design. Vaccines. 2020;9:1.
Clements JD, Norton EB. The mucosal vaccine adjuvant LT(R192G/L211A) or dmLT. mSphere. 2018;3:e00215-18.
Buckholz RG, Gleeson MA. Yeast systems for the commercial production of heterologous proteins. Biotechnology. 1991;9(11):1067–72.
Strausberg RL, Strausberg SL. Overview of protein expression in Saccharomyces cerevisiae. Curr Protoc Protein Sci. 2001 Chapter 5: Unit5.6.
Bui K, Galzy P. Food yeast. In: Spencer JFT, Spencer DM, editors. Yeast technology. Berlin Heidelberg New York: Springer; 1990. p. 241–65.
Elghandour MMY, Tan ZL, Abu Hafsa SH, Adegbeye MJ, Greiner R, Ugbogu EA, Cedillo Monroy J, Salem AZM. Saccharomyces cerevisiae as a probiotic feed additive to non and pseudo-ruminant feeding: a review. J Appl Microbiol. 2020;128(3):658–74.
Goh S, Kolakowski J, Holder A, Pfuhl M, Ngugi D, Ballingall K, Tombacz K, Werling D. Development of a potential yeast-based vaccine platform for Theileria parva infection in cattle. Front Immunol. 2021;12: 674484.
Gao T, Ren Y, Li S, Lu X, Lei H. Immune response induced by oral administration with a Saccharomyces cerevisiae-based SARS-CoV-2 vaccine in mice. Microb Cell Fact. 2021;20:95.
Cen Q, Gao T, Ren Y, Lu X, Lei H. Immune evaluation of a Saccharomyces cerevisiae-based oral vaccine against Helicobacter pylori in mice. Helicobacter. 2021;26: e12772.
Bal J, Luong NN, Park J, Song KD, Jang YS, Kim DH. Comparative immunogenicity of preparations of yeast-derived dengue oral vaccine candidate. Microb Cell Fact. 2018;17:24.
Bal J, Jung HY, Nguyen LN, Park J, Jang YS, Kim DH. Evaluation of cell-surface displayed synthetic consensus dengue EDIII cells as a potent oral vaccine candidate. Microb Cell Fact. 2018;17:146.
Seif M, Philippi A, Breinig F, Kiemer AK, Hoppstädter J. Yeast (Saccharomyces cerevisiae) polarizes both M-CSF- and GM-CSF-differentiated macrophages toward an M1-Like phenotype. Inflammation. 2016;39:1690–703.
Kim JM, Park SM, Kim JA, Park JA, Yi MH, Kim NS, Bae JL, Park SG, Jang YS, Yang MS, Kim DH. Functional pentameric formation via coexpression of the Escherichia coli heat-labile enterotoxin B subunit and its fusion protein subunit with a neutralzing epitope of ApxIIA exotoxin improves the mucosal immunogenicity and protection against challenge by Actinobacillus pleuropneumoniae. Clin Vaccine Immunol. 2011;18:2168–77.
Shin SJ, Bae JL, Cho YW, Lee DY, Kim DH, Yang MS, Jang YS, Yoo HS. Induction of antigen-specific immune responses by oral vaccination with Saccharomyces cerevisiae expressing Actinobacillus pleuropneumoniae ApxIIA. FEMS Immunol Med Microbiol. 2005;43:155–64.
Nguyen NL, Kim JM, Park JA, Park SM, Jang YS, Yang MS, Kim DH. Expression and purification of an immunogenic dengue virus epitope using a synthetic consensus sequence of envelope domain III and Saccharomyces cerevisiae. Protein Expr Purif. 2013;88:235–42.
Kaishima M, Ishii J, Matsuno T, Fukuda N, Kondo A. Expression of varied GFPs in Saccharomyces cerevisiae: codon optimization yields stronger than expected expression and fluorescence intensity. Sci Rep. 2016;6:35932.
Partow S, Siewers V, Bjørn S, Nielsen J, Maury J. Characterization of different promoters for designing a new expression vector in Saccharomyces cerevisiae. Yeast. 2010;27:955–64.
Blazeck J, Garg R, Reed B, Alper HS. Controlling promoter strength and regulation in Saccharomyces cerevisiae using synthetic hybrid promoters. Biotechnol Bioeng. 2012;109:2884–95.
Curran KA, Karim AS, Gupta A, Alper HS. Use of expression-enhancing terminators in Saccharomyces cerevisiae to increase mRNA half-life and improve gene expression control for metabolic engineering applications. Metab Eng. 2013;19:88–97.
Gnügge R, Liphardt T, Rudolf F. A shuttle vector series for precise genetic engineering of Saccharomyces cerevisiae. Yeast. 2016;33:83–98.
Huang D, Shusta EV. Secretion and surface display of green fluorescent protein using the yeast Saccharomyces cerevisiae. Biotechnol Prog. 2005;21:349–57.
Kimata Y, Iwaki M, Lim CR, Kohno K. A novel mutation which enhances the fluorescence of green fluorescent protein at high temperatures. Biochem Biophys Res Commun. 1997;232:69–73.
Papaneophytou CP, Kontopidis G. Statistical approaches to maximize recombinant protein expression in Escherichia coli: a general review. Protein Expr Purif. 2014;94:22–32.
Kiefhaber T, Rudolph R, Kohler HH, Buchner J. Protein aggregation in vitro and in vivo: a quantitative model of the kinetic competition between folding and aggregation. Biotechnology. 1991;9(9):825–9.
Torres M, Dickson AJ. Combined gene and environmental engineering offers a synergetic strategy to enhance r-protein production in Chinese hamster ovary cells. Biotechnol Bioeng. 2022;119(2):550–65.
Torres M, Akhtar S, McKenzie EA, Dickson AJ. Temperature down-shift modifies expression of UPR-/ERAD-Related genes and enhances production of a chimeric fusion protein in CHO cells. Biotechnol J. 2021;16(2): e2000081.
Kaufmann H, Mazur X, Fussenegger M, Bailey JE. Influence of low temperature on productivity, proteome and protein phosphorylation of CHO cells. Biotechnol Bioeng. 1999;63(5):573–82.
Herrera-Estala AL, Fuentes-Garibay JA, Guerrero-Olazarán M, Viader-Salvadó JM. Low specific growth rate and temperature in fed-batch cultures of a beta-propeller phytase producing Pichia pastoris strain under GAP promoter trigger increased KAR2 and PSA1-1 gene expression yielding enhanced extracellular productivity. J Biotechnol. 2022;20(352):59–67.
Sirén N, Weegar J, Dahlbacka J, Kalkkinen N, Fagervik K, Leisola M, von Weymarn N. Production of recombinant HIV-1 Nef (negative factor) protein using Pichia pastoris and a low-temperature fed-batch strategy. Biotechnol Appl Biochem. 2006;44(Pt 3):151–8.
Yang Z, Zhang Z. Engineering strategies for enhanced production of protein and bio-products in Pichia pastoris: a review. Biotechnol Adv. 2018;36(1):182–95.
Kubiak M, Borkowska M, Białas W, Korpys P, Celińska E. Feeding strategy impacts heterologous protein production in Yarrowia lipolytica fed-batch cultures-Insight into the role of osmolality. Yeast. 2019;36(5):305–18.
Zhou X, Chandarajoti K, Pham TQ, Liu R, Liu J. Expression of heparan sulfate sulfotransferases in Kluyveromyces lactis and preparation of 3’-phosphoadenosine-5’-phosphosulfate. Glycobiology. 2011;21(6):771–80.
Park SW, Kang BH, Lee HM, Lee SJ, Kim HS, Choi HW, Park TJ, Kong KH. Efficient brazzein production in yeast (Kluyveromyces lactis) using a chemically defined medium. Bioprocess Biosyst Eng. 2021;44(4):913–25.
Soutter F, Werling D, Nolan M, Küster T, Attree E, Marugán-Hernández V, Kim S, Tomley FM, Blake DP. A novel whole yeast-based subunit oral vaccine against Eimeria tenella in chickens. Front Immunol. 2022;13: 809711.
So KK, Chun J, Luong NN, Seo HW, Kim DH. Expression of an immunocomplex consisting of Fc fragment fused with a consensus dengue envelope domain III in Saccharomyces cerevisiae. Biotechnol Lett. 2021;43:1895–904.
Zhu L, Lei Z, Xia X, Zhang Y, Chen Y, Wang B, Li J, Li G, Yang G, Cao G, Yin Z. Yeast shells encapsulating adjuvant AS04 as an antigen delivery system for a novel vaccine against Toxoplasma Gondii. ACS Appl Mater Interfaces. 2021;13:40415–28.
Chen C, Hua D, Shi J, Tan Z, Zhu M, Tan K, Zhang L, Huang J. Porcine immunoglobulin Fc fused P30/P54 protein of african swine fever virus displaying on surface of S. cerevisiae elicit strong antibody production in swine. Virol Sin. 2021;36:207–19.
Lei H, Lu X, Li S, Ren Y. High immune efficacy against different avian influenza H5N1 viruses due to oral administration of a Saccharomyces cerevisiae-based vaccine in chickens. Sci Rep. 2021;11:8977.
Curran KA, Morse NJ, Markham KA, Wagman AM, Gupta A, Alper HS. Short synthetic terminators for improved heterologous gene expression in yeast. ACS Synth Biol. 2015;4:824–32.
Ito Y, Yamanishi M, Ikeuchi A, Imamura C, Tokuhiro K, Kitagawa T, Matsuyama T. Characterization of five terminator regions that increase the protein yield of a transgene in Saccharomyces cerevisiae. J Biotechnol. 2013;168:486–92.
Tang H, Bao X, Shen Y, Song M, Wang S, Wang C, Hou J. Engineering protein folding and translocation improves heterologous protein secretion in Saccharomyces cerevisiae. Biotechnol Bioeng. 2015;112:1872–82.
Aguilera J, Randez-Gil F, Prieto JA. Cold response in Saccharomyces cerevisiae: new functions for old mechanisms. FEMS Microbiol Rev. 2007;31:327–41.
Mattanovich D, Gasser B, Hohenblum H, Sauer M. Stress in recombinant protein producing yeasts. J Biotechnol. 2004;113:121–35.
Rosano GL, Ceccarelli EA. Recombinant protein expression in Escherichia coli: advances and challenges. Front Microbiol. 2014;5:172.
Nguyen MT, Krupa M, Koo BK, Song JA, Vu TT, Do BH, Nguyen AN, Seo T, Yoo J, Jeong B, Jin J, Lee KJ, Oh HB, Choe H. Prokaryotic soluble overexpression and purification of human VEGF165 by fusion to a maltose binding protein tag. PLoS ONE. 2016;11: e0156296.
Qing G, Ma LC, Khorchid A, Swapna GV, Mal TK, Takayama MM, Xia B, Phadtare S, Ke H, Acton T, Montelione GT, Ikura M, Inouye M. Cold-shock induced high-yield protein production in Escherichia coli. Nat Biotechnol. 2004;22:877–82.
Li Z, Xiong F, Lin Q, d’Anjou M, Daugulis AJ, Yang DS, Hew CL. Low-temperature increases the yield of biologically active herring antifreeze protein in Pichia pastoris. Protein Expr Purif. 2001;21:438–45.
Jahic M, Wallberg F, Bollok M, Garcia P, Enfors SO. Temperature limited fed-batch technique for control of proteolysis in Pichia pastoris bioreactor cultures. Microb Cell Fact. 2003;2:6.
Shi X, Karkut T, Chamankhah M, Alting-Mees M, Hemmingsen SM, Hegedus D. Optimal conditions for the expression of a single-chain antibody (scFv) gene in Pichia pastoris. Protein Expr Purif. 2003;28:321–30.
Hohenblum H, Borth N, Mattanovich D. Assessing viability and cell-associated product of recombinant protein producing Pichia pastoris with flow cytometry. J Biotechnol. 2003;102:281–90.
Lin CY, Huang Z, Wen W, Wu A, Wang C, Niu L. Enhancing protein expression in HEK-293 cells by lowering culture temperature. PLoS ONE. 2015;10: e0123562.
Tai SL, Daran-Lapujade P, Walsh MC, Pronk JT, Daran JM. Acclimation of Saccharomyces cerevisiae to low temperature: a chemostat-based transcriptome analysis. Mol Biol Cell. 2007;18:5100–12.
Miller MJ, Xuong NH, Geiduschek EP. A response of protein synthesis to temperature shift in the yeast Saccharomyces cerevisiae. Proc Natl Acad Sci USA. 1979;76:5222–5.
Graf A, Dragosits M, Gasser B, Mattanovich D. Yeast systems biotechnology for the production of heterologous proteins. FEMS Yeast Res. 2009;9:335–48.
Huang M, Bai Y, Sjostrom SL, Hallström BM, Liu Z, Petranovic D, Uhlén M, Joensson HN, Andersson-Svahn H, Nielsen J. Microfluidic screening and whole-genome sequencing identifies mutations associated with improved protein secretion by yeast. Proc Natl Acad Sci U S A. 2015;112:E4689–96.
Huang M, Bao J, Hallström BM, Petranovic D, Nielsen J. Efficient protein production by yeast requires global tuning of metabolism. Nat Commun. 2017;8:1131.
Lim YY, Park EH, Kim JH, Park SM, Jang HS, Yoon SW, Kim DH. Enhanced and targeted expression of fungal phytase in Saccharomyces cerevisiae. J Microbiol Biotechnol. 2001;11(6):915–21.
Valkonen M, Penttilä M, Saloheimo M. Effects of inactivation and constitutive expression of the unfolded- protein response pathway on protein production in the yeast Saccharomyces cerevisiae. Appl Environ Microbiol. 2003;69:2065–72.
Chen X, Zaro JL, Shen WC. Fusion protein linkers: property, design and functionality. Adv Drug Deliv Rev. 2013;65:1357–69.
Gietz RD, Schiestl RH. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat Protoc. 2007;2:31–4.
Lim YY, Park SM, Jang YS, Yang MS, Kim DH. Production of a functional mouse interferon g from recombinant Saccharomyces cerevisiae. J Microbiol Biotechnol. 2003;13:537–43.
Park JA, Kim JM, Park SM, Kim DH. Characterization of CpSte11, a MAPKKK gene of Cryphonectria parasitica, and initial evidence of its involvement in the pheromone response pathway. Mol Plant Pathol. 2012;13:240–50.
Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, Preibisch S, Rueden C, Saalfeld S, Schmid B, Tinevez JY, White DJ, Hartenstein V, Eliceiri K, Tomancak P, Cardona A. Fiji: an open-source platform for biological-image analysis. Nat Methods. 2012;9:676–82.
Acknowledgements
We are thankful to the Institute of Molecular Biology and Genetics at Jeonbuk National University for kindly providing the facilities for this research.
Funding
This work was supported in part by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2017R1A6A1A03015876).
Author information
Authors and Affiliations
Contributions
N-LN: designed the experiments and analyzed data, K-KS, NMTL: performed the experiments, D-HK: supervised the experiments and wrote the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that there are no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure. S1
Western blot analysis of temporal expression of LTB-EDIII2 from transformant #8 using anti-Dengue antibody. A LTB-EDIII2 expression was resolved under non-denaturing condition at 20 °C and 30 °C. B SDS-PAGE gel showed that an equal amount of protein was loaded on each lane. Lane 1: purified E. coli-expressed LTB as a positive control; Lane 2: a mock transformant cultured for 3 days as a negative control. Proteins were prepared from transformant #8, and cultured for 1, 3, 5 days after inoculation to the expression medium at 20 °C (lanes 3, 5, and 7, respectively) and 30 °C (lanes 4, 6, and 8, respectively).
Additional file 2: Figure. S2
Northern blot analysis (A) and Quantitative real time RT-PCR (qRT-PCR) analysis (B) of LTB-VP1 in a selected transformant (#4) under 20 °C and 30 °C conditions. A 20 μg total RNA was loaded on each lane. RNA preparations from cells harvested at day 1, 3, and 5 days after cultivation at 20 °C (Lanes 2-4, respectively) and 30 °C (Lanes 5-7, respectively). Lane 1 contains RNA sample from a mock transformant cultured for 3 days at 30 °C as a negative control. GPD was used as an internal control and rRNAs are shown to indicate equal amount of RNA loaded on each lane. B qRT-PCR results of changes in expression of LTB-VP1 under 20 °C and 30 °C conditions are shown. No significant differences between 20 °C and 30 °C were observed.
Additional file 3: Table S1.
List of Primers used in this study.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
So, KK., Le, N.M.T., Nguyen, NL. et al. Improving expression and assembly of difficult-to-express heterologous proteins in Saccharomyces cerevisiae by culturing at a sub-physiological temperature. Microb Cell Fact 22, 55 (2023). https://doi.org/10.1186/s12934-023-02065-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-023-02065-7