
Abstract
Background
Carotenoids are natural pigments with antioxidant properties that have important functions in human physiology and must be supplied through the diet. They also have important industrial applications as food colourants, animal feed additives and nutraceuticals. Some of them, such as β-carotene, are produced on an industrial scale with the use of microorganisms, including fungi. The mucoral Blakeslea trispora is used by the industry to produce β-carotene, although optimisation of production by molecular genetic engineering is unfeasible. However, the phylogenetically closely related Mucor circinelloides, which is also able to accumulate β-carotene, possesses a vast collection of genetic tools with which to manipulate its genome.
Results
This work combines classical forward and modern reverse genetic techniques to deepen the regulation of carotenoid synthesis and generate candidate strains for biotechnological production of β-carotene. Mutagenesis followed by screening for mutants with altered colour in the dark and/or in light led to the isolation of 26 mutants that, together with eight previously isolated mutants, have been analysed in this work. Although most of the mutants harboured mutations in known structural and regulatory carotenogenic genes, eight of them lacked mutations in those genes. Whole-genome sequencing of six of these strains revealed the presence of many mutations throughout their genomes, which makes identification of the mutation that produced the phenotype difficult. However, deletion of the crgA gene, a well-known repressor of carotenoid biosynthesis in M. circinelloides, in two mutants (MU206 and MU218) with high levels of β-carotene resulted in a further increase in β-carotene content to differing extents with respect to the crgA single-null strain; in particular, one strain derived from MU218 was able to accumulate up to 4 mg/g of β-carotene. The additive effect of crgA deletion and the mutations present in MU218 suggests the existence of a previously unknown regulatory mechanism that represses carotenoid biosynthesis independently and in parallel to crgA.
Conclusions
The use of a mucoral model such as M. circinelloides can allow the identification of the regulatory mechanisms that control carotenoid biosynthesis, which can then be manipulated to generate tailored strains of biotechnological interest. Mutants in the repressor crgA and in the newly identified regulatory mechanism generated in this work accumulate high levels of β-carotene and are candidates for further improvements in biotechnological β-carotene production.
Similar content being viewed by others

Background
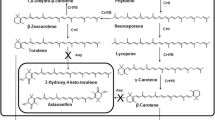
β-carotene is a fat-soluble pigment that has several biological functions and great commercial value due to its diverse uses in food, pharmaceutical products, cosmetics and textiles [1–3]. Thus, the global carotenoid market value was $1.5 billion in 2014 and is expected to reach nearly $1.8 billion in 2019, with a compound annual growth rate of 3.9 % [4]. Microbial β-carotene has attracted much attention because of its rapid generation and lack of influence from geographic location and climate. The microbial β-carotene currently produced by the industry comes from the alga Dunaliella salina [5] or the fungus Blakeslea trispora [6]. B. trispora accumulates large amounts of carotenoids, as do other fungi of the subphylum mucormycotina, but the absence of molecular tools with which to manipulate its genome makes it difficult to improve carotenoid production. In contrast, the genome of Mucor circinelloides, which belongs to the same subphylum, has been sequenced (http://www.genome.jgi.doe.gov/Mucci2/Mucci2.home.html), and procedures for efficient genetic transformation, gene replacement and gene silencing by RNAi [7] allow improvement of carotene production by genetic manipulation.
Fungal carotenoids derive from 3-hydroxy-3-methyl-glutaryl-CoA (HMG-CoA) in the mevalonate pathway [8] and is regulated by complex genetic mechanisms. Light induction of carotenoid biosynthesis is considered to be conserved in filamentous fungi such as Neurospora crassa, Phycomyces blakesleeanus and M. circinelloides [9–11]. Transcription of structural genes for carotenoid biosynthesis increased enormously in response to light, in accordance with the improved carotene content. In the case of N. crassa, a model was established in which induction of carotenogenesis by light is mediated by the white-collar protein complex that is formed with the products of wc-1 and wc-2 genes [12, 13]. Genes homologous to wc-1 and wc-2 have been identified in several fungi, and one of the three homologs to wc-1 of M. circinelloides, mcwc-1c, was also shown to be involved in the induction of carotenogenesis by light [9]. Deletion of mcwc-1c led to only a threefold increase in β-carotene levels, compared to the 21-fold increase in the wild-type strain after light exposure, which indicates that light-induced carotenogenesis in M. circinelloides requires mcwc-1c [9].
Another regulatory gene identified in M. circinelloides is crgA, which codes for a RING finger protein that was regarded as a negative regulator of carotenoid biosynthesis, because its disruption resulted in β-carotene overproduction in the dark as a result of a sharp increase of the carB and carRP mRNA levels [14]. The presence of RING-finger zinc-binding domains, which define a family of ubiquitin ligases that mediate ubiquitylation of target proteins, suggests that CrgA acts as an E3 ubiquitin ligase. In fact, CrgA blocks the function of Mcwc-1b, a protein encoded by a second wc-1 homolog, by mono-ubiquitylation and di-ubiquitylation without degradation [15], which has also been observed in the regulatory process of the budding yeast transcription factor Met4 [16].
The crgA-mediated repression of carotenogenesis and light-induced carotenogenesis are independent regulatory pathways because the crgA mcwc-1c double mutant presented similar levels of β-carotene as the crgA single mutant in the dark but was not able to enhance the production to the same level as the crgA single mutant after illumination [15]. The occurrence of multiple copies of wc-1 genes in mucoromycotina fungi such as M. circinelloides [9], P. blakesleeanus [17], Rhizopus oryzae [17] and Pilobolus crystallinus [18], and the existence of four transduction pathways in charge of different light responses in P. blakesleeanus, suggests that these basal fungi may have developed a more complex light regulatory system than dikaryotic fungi (ascomycetes and basiodiomycetes), which contains a single copy of the wc-1 gene [19, 20]. Moreover, the abundance of genes that regulate carotenogenesis, such as carC, carD, carS, carF and carI in P. blakesleeanus [21–24], suggests that carotene biosynthesis in M. circinelloides may be also regulated by a variety of elements.
In this report, we analysed 34 mutants in which carotenoid biosynthesis was affected to examine the regulatory mechanisms that control carotenoid biosynthesis in M. circinelloides and to generate an overproducing strain of interest for β-carotene production. Eight of these mutants lacked mutations on known structural or regulatory genes, suggesting that they contained mutations in unknown genes involved in carotenoid biosynthesis. Two of these mutants, MU206 and MU218, clearly carried mutations in regulatory genes because they accumulated greater amounts of β-carotene than the wild-type strain. Deletion of crgA in both strains produced an increase in β-carotene levels that was much greater in the crgA-null derivative of MU218, which suggests that the gene affected in MU218 is involved in a regulatory pathway other than that of crgA, whereas the gene mutated in MU206 may participate in the same regulatory pathway as crgA. Moreover, some of the overproducing strains may be of interest for industrial production of β-carotene.
Results
Isolation of mutants affected in the regulation of carotenoid biosynthesis
The mycelium of M. circinelloides has a white-yellowish appearance in the dark because it accumulates a small amount of β-carotene. In the light, the biosynthesis of this pigment is stimulated and the mycelium becomes deep yellow. To identify new genes implicated in the regulation of carotenogenesis, mutagenized spores of the wild-type strain R7B were allowed to follow a complete vegetative cycle to permit expression of recessive mutations, since initial mutants are heterokaryons due to the multinucleate nature of R7B. The recycled spores were used to screen for colour mutants both in the dark and after exposure to light.
After screening about 4 × 105 colonies, 24 colour mutants from N-methyl-N’-nitro-N-nitrosoguanidine (NTG) mutagenesis were isolated and purified by successive vegetative cycles until all of the colonies from each particular mutant showed the very same colour phenotype, indicating that they were true homokaryotic mutants. The strains were named from MU260 to MU269, MU271 to MU278 and MU280 to MU285. After ultraviolet (UV) light treatment, only two colour mutants, MU279 and MU286, were isolated after screening about 5.6 × 105 spores. In this study, we included mutants (MU206, MU212 to MU218) from our collection (University of Murcia, Spain) that were isolated during a previous screening of colonies derived from a different NTG treatment [25]. Most of the mutants showed a white colour in the dark as the wild-type strain but appeared white or pale yellow in the light, whereas the wild-type strain showed a yellow colour (Table 1). In contrast, mutant MU206 and MU218 appeared pale yellow in the dark and yellow and deep yellow, respectively, in the light.
Analysis of carotenogenic genes in mutants
The phenotypes shown by the mutants could be due to mutations in the structural carotenogenic genes (carB and carRP) and in known (mcwc-1c) [9] or unknown regulatory genes, including the genes that code for putative homologs to N. crassa wc-2 [26] and P. blakesleeanus madB [17]. Mutations in the repressor crgA were not considered because none of the mutants showed a bright yellow colour, which is a hallmark of mutations in that gene [14]. Thus, the carB-carRP genomic region, including coding and promoter sequences, and coding and promoter sequences (~200 bp) of mcwc-1c and the four putative mcwc-2 genes (ID78608, ID85699, ID153812 and ID157130) of the 34 colour mutants were amplified, and the polymerase chain reaction (PCR) fragments were directly sequenced without previous cloning to avoid detection of mutations produced by the PCR. The obtained sequences were compared to the corresponding wild-type alleles of the reference genome (http://www.genome.jgi.doe.gov/Mucci2/Mucci2.info.html) to identify mutations. The results of this analysis show that 25 strains presented different mutations in mcwc1c, carB and carRP, including deletions of carB-carRP, characterised by Southern blot hybridisation (data no shown), in two strains (Table 1). Several mutants carried the same mutation suggesting that they derived from an original mutant that was amplified in the spore-recycling step. Surprisingly, no mutations were found in the four mcwc-2 genes. In contrast, nine mutants showed no mutations in the analysed genes and should thus carry mutations in uncharacterised genes that are probably involved in the regulation of carotenoid biosynthesis.
Carotene content in carotenogenic mutants with uncharacterised mutations
To confirm the phenotype of the nine mutants with no mutations in known carotenogenic and putative regulatory genes, their carotene content was determined both in the dark and light and compared with that of the wild-type parental strain R7B. Five of the six strains with a pale yellow colour in light accumulated lower amounts of carotenes than the wild-type strain (Table 2), whereas MU282 showed β-carotene levels similar to those of the wild-type strain even though its mycelium was pale yellow in light. The discrepancy between colour and β-carotene content in this mutant could be caused by a growth defect because its mycelium was less dense than that of the wild-type strain. Scanning spectrophotometry of the carotenoids accumulated in light by the pale yellow mutants showed the characteristic shape and peaks for β-carotene, indicating that this is the main carotene accumulated by all of them, as it happens in the wild-type strain (Additional file 1: Figure S1). The accumulation of mainly β-carotene in the wild-type strain was confirmed by high-performance liquid chromatography (Additional file 2: Figure S2).
In contrast, the white mutant MU264 showed undetectable levels of β-carotene in the dark and light (Table 2; Additional file 1: Figure S1). Special attention was dedicated to this mutant and carotenoids accumulated by this mutant were analysed by HPLC (Additional file 2: Figure S2). No carotenoids were detected in light, including phytoene, indicating that MU264 carries a mutation that blocks completely the carotenoid pathway. In addition to the mutants with low β-carotene levels, two strains, MU206 and MU218, accumulated high levels of β-carotene both in the dark and in light, suggesting that they are affected in genes involved in the regulation of carotenoid biosynthesis. The levels of β-carotene in those strains in the dark were roughly ten times higher than the levels in the wild-type strain R7B, whereas increases in light of 1.5 and 4.5 times were seen in MU206 and MU218, respectively (Table 2).
Characterisation of carotenogenic mutant genomes
The devised strategy for identification of mutations that provoked alterations in β-carotene levels was sequencing of the entire genome followed by comparison to the M. circinelloides reference genome. Although eight strains showed altered β-carotene levels, six strains (MU206, MU216, MU217, MU218, MU264 and MU286) with different pattern of β-carotene accumulation were selected for whole-genome sequencing and two (MU283 and MU285) were preserved for future analyses. In addition to the six strains with altered carotenogenesis, the genome of parental strain R7B was sequenced for use as a reference because this strain was obtained by UV light treatment and could harbour additional mutations to that in the 3-isopropylmalate dehydratase gene (leuA) gene that makes it auxotrophic for leucine [27].
The genome sequences of the seven genomes were obtained using SOLiD technology, and the sequences were mapped against the M. circinelloides reference genome (http://www.genome.jgi.doe.gov/Mucci2/Mucci2.info.html) to detect base substitutions and small deletions and insertion (indels). More than 99.7 % of the reference genome was successfully mapped for each sample, and the mean coverage depth over that mapped region ranged from 39.8 to 53.6× (Additional file 3: Table S1). As a proof of the quality of the genome sequences, the known single-base substitution in the leuA gene that causes a change from glutamic acid to lysine at position 220 [28] was detected in all genomes (Additional files 4, 5: Tables S2, S3). As expected, the R7B genome contained additional mutations, which included three base substitutions that resulted in amino acid changes in a putative E3 ubiquitin-protein ligase (P111S), in a DUF1688 domain–containing protein (D331Y) and in a putative helicase-2 (S308G) (Additional files 4, 5: Tables S2, S3). In addition, 11 base substitutions were found in intergenic regions, one in the intron 1 of basic helix-loop-helix transcription factor-like protein (ID113859) and two others that did not produce amino acid changes (ID76084 and ID164535). Each of these base substitutions was present in the six mutants, except the mutation in the DUF1688 domain–containing protein and two base substitutions in intergenic regions that only appear in MU264 and MU286. The strains without those substitutions were obtained from spores treated with NTG in 1993 and kept frozen until 2009, whereas the strains with the substitutions were isolated in 2009. The genomic DNA from R7B was isolated from the strain used in 2009 and, therefore, it is possible that those changes occurred spontaneously in the R7B strain in our laboratory between 1993 and 2009. In any case, the presence of the mutation in the gene coding for the DUF1688 domain–containing protein in R7B discarded the notion that this mutation is responsible for the phenotype of the mutants.
All mutant strains showed a large number of base substitutions and some indels in comparison to R7B (Table 3; Additional files 4, 5, 6: Tables S2, S3, S4), and most of them corresponded to intergenic regions. Only missense mutations or indels in coding regions were considered to be causative for the phenotypes displayed by the mutants and were analysed in detail. The mutant MU206 presented just one missense mutation (A125 V) in a predicted gene that codes for a protein that only appears in M. circinelloides (Table 3). MU264 carries two missense mutations: one (D380 N) in the gene aspB that codes for a putative septin and the other one (C171Y) in a gene that codes for a putative F-box protein (Table 3). The mutation in the F-box protein affected one of the crucial residues of the F-box domain. In addition to the missense mutations, MU264 has an 11-bp deletion in a gene that codes for a class 3 adenylyl cyclase that provokes truncation of the protein (Table 3). MU286 has one missense mutation (P152L) in a gene that codes for a class 3 lipase and two small deletions that originate truncated proteins, one in a gene that codes for a protein similar to a vacuolar dynamin-like GTPase VpsA and another in a gene that codes for a protein similar to the autophagy-related protein 9 (Table 3). Mutants MU216, MU217 and MU218 have many mutations in several genes (Table 3), and it is difficult to determine which of them is responsible for the mutant phenotype.
To validate the whole-genome sequencing, affected genes from mutants with the fewest affected genes (MU206, MU264 and MU286) were amplified by PCR and sequenced. In all cases, the sequences of the amplified fragments contained the mutations identified in whole-genome sequencing (data not shown), validating the whole-genome sequencing. Moreover, those mutants were transformed with self-replicative plasmids that contained the wild-type alleles corresponding to the mutated genes, and functional complementation was not observed in any case (data not shown), which suggests either the genes in the plasmids were not efficiently expressed or that the phenotype could be provoked for mutations in putative intergenic regions. The lack of complementation is particularly relevant in the case of mutant MU206 because it showed just one base substitution in one gene and was used as a parental strain in the experiments described below.
Generation of crgA null mutants in MU206 and MU218 backgrounds
The enhancement of carotene biosynthesis in MU206 and MU218 (Table 2) suggests that they were affected in genes involved in some regulatory pathway that controls carotenoid biosynthesis. This hypothetical pathway might limit the accumulation of carotenoid in the dark, as with the regulatory pathway in which the crgA gene is the principal element [14, 15]. The lack of the crgA gene led to enrichment of carotene in both dark and light conditions [14], as with MU206 and MU218, although they lacked mutations in the crgA gene (Table 3). Disruption of the crgA gene in these two strains could affect carotenogenesis regulation, in addition to generation of β-carotene–overaccumulating strains of potential interest for industrial production of β-carotene. For this purpose, plasmid pVEN172 [14], which harbours the leuA wild-type allele gene of M. circinelloides flanked by the sequences neighbouring the crgA gene, was adopted in this study. The 6.5-kb fragment released by digestion of pVEN172 with restriction enzyme PstI was used to transform MU206 and MU218, both of which are leuA auxotrophs. Integration of this fragment into the genome of the recipient strains by homologous recombination was expected to cause replacement of the endogenous crgA gene with the wild-type leuA allele gene (Fig. 1).

The genomic structure of crgA wild-type locus and after disruption by homologous recombination. The replacement fragment used to disrupt crgA gene corresponded to a 6.5-kb PstI fragment derived from plasmid pVEN172. The positions of primers CigC F1/crgA R1 used to amplify the disrupted and wild-type crgA locus, and the expected sizes of the corresponding PCR products, are indicated
Transformation of MU206 and MU218 with the replacement fragment produced 27 and 31 transformants, respectively. The initial transformants were grown in a selective medium for several vegetative cycles to identify stable Leu+ transformants, which indicated that exogenous DNA integration had occurred. Finally, four and three homokaryotic transformants that showed a 100 % stable Leu+ phenotype were obtained from MU206 and MU218, respectively. Three transformants derived from MU206 (MU609, MU611 and MU612) presented a deeper yellow colour than the other (MU610). In a similar manner, two transformants derived from MU218 (MU605 and MU606) displayed a deeper yellow colour than the other (MU607). Colour differences could result from different integration events and, therefore, the genomic DNA of all seven homokaryotic transformants and the parental strains MU206 and MU218 were analysed by PCR using primers that amplify both the disrupted and the wild-type crgA locus-producing fragments of 4 and 6.5 kb, respectively. The transformants that showed the deepest yellow colour (MU605, MU606, MU609, MU611 and MU612) showed only the 6.5-kb PCR fragment (Fig. 2) that indicated their lack of a wild-type crgA allele, which had been replaced by the leuA gene (Fig. 1). In contrast, MU607 and MU610 showed the same 4-kb PCR fragment as the parental strains MU206 and MU218 (Fig. 2), which indicated that they contained a wild-type allele of crgA and therefore carried an ectopic integration of the replacement fragment.

Analysis of the crgA locus in knockout mutants. Genomic DNA from the indicated strains was amplified by PCR using primers cigC-F1 and crgA-R1 that amplified wild-type (4.0 kb) and mutant crgA alleles (6.5 kb) (Fig. 1), and the PCR products were analysed by agarose gel electrophoresis. MU605, MU606 and MU607 were derived from MU218, whereas MU609, MU610, MU611 and MU612 were derived from MU206. Marker, λ DNA digested with HindIII. Sizes of the DNA marker relevant fragments are indicated on the left side of the figure
Analysis of carotenogenesis in crgA-null derivatives of MU206 and MU218
The effect of crgA deletion in mutants MU206 and MU218 was analysed by comparison of the β-carotene levels in the obtained transformants, the parental strains and a mutant with deletion of crgA in a wild-type background (MU221) [14]. The obtained transformants and MU221 were prototrophs, whereas the parental strains MU206 and MU218 were leucine auxotrophs. Due to the known effect of leucine on the carotenoid content [9], leucine auxotrophy of the parental strains was complemented by transformation with the autoreplicative plasmid pLEU4-producing strains MU608 and MU613, which were derived from MU218 and MU206, respectively (Table 4).
The β-carotene content both in the dark and in light in all crgA mutants derived from MU206 (MU609, MU611 and MU612) and from MU218 (MU605 and MU606) was higher than that seen in MU221 and the prototrophic MU206 strain (MU613). In contrast, the transformants that carried an ectopic integration of the replacement fragment (MU610 for MU206 and MU607 for MU218) showed β-carotene levels similar to those of the corresponding prototrophic strains (Table 4). Although the simultaneous presence of mutations in MU206 or MU218 and crgA deletion produced an increase in the β-carotene content compared to MU221, the effect was much greater with an MU218 background (Table 4).
High levels of β-carotene can be produced by a high level of expression of structural carotenogenic genes [14]. To determine whether the high β-carotene levels in mutants derived from MU206 and MU218 were linked to a high level of expression of carotenogenic genes, the accumulation of carB and carRP mRNAs was analysed in mycelium grown both in the dark and in light by Droplet Digital PCR [29]. In agreement with the high carotenoid levels, mutants MU206 and MU218, and their corresponding strains with ectopic integrations, accumulated higher levels of carB and carRP mRNAs than the wild-type strain R7B after a light pulse, and also carB mRNA levels in the dark (Fig. 3). However, the carB and carRP mRNA levels in all crgA-knockout mutants were higher than those seen in any strain with a crgA wild-type allele. Thus, MU221 showed a considerable increase in the carB and carRP mRNA levels, in concordance with the high carotene levels accumulated by this strain. Also in agreement with the β-carotene content, the carB and carRP mRNA levels were increased in crgA mutants derived from strain MU206 (M609, and M611) and were increased even more in crgA mutants derived from MU218 (MU605 and MU606) (Fig. 3). Especially interesting was MU606, because it showed the highest carB and carRP mRNA levels, with 27-fold and sixfold increases in the carB mRNA levels and 25-fold and 15-fold increases in the carRP mRNA levels in the dark and light, respectively, with respect to the wild-type strain R7B. Moreover, it accumulated approximately 3 and 4 mg/g dry weight of β-carotene in the dark and in light, respectively. Compared to MU221, the β-carotene production in MU606 was doubled under both dark and light conditions. The disruption crgA gene in MU206 failed to improve the carotene production as significantly as in MU218. The β-carotene content of the transformants of MU206 is similar to that seen in MU221; even MU611, which possesses the highest amount of β-carotene amongst the M206 crgA mutants, rose slightly to 2.8 mg/g in light.

Expression of carB (a) and carRP (b) genes in crgA-null mutants derived from MU206 and MU218. Copy number of the carB (a) and carRP (b) mRNAs in total RNA sample from the indicated strains was determined by Droplet Digital PCR. The carB and carRP mRNA copy numbers were normalised to the actin reference gene. All RNA samples were diluted 10 times and detected in triplicate. MU221, prototrophic crgA mutant derived from R7B; MU218 and MU206, derived from R7B, leuA −; MU605, MU606: crgA-null mutants derived from MU218; MU607, prototrophic strain derived from MU218; MU609 and MU611, crgA-null mutant derived from MU206; MU610, prototrophic strain derived from MU206
Discussion
New regulatory elements in the control of M. circinelloides carotenoid biosynthesis
Several partially characterised regulatory networks control carotenogenesis in fungi that belong to the mucoromycotina subphylum. In P. blakesleeanus, the biosynthesis of carotenoid biosynthesis is regulated by a complex regulatory network triggered with several environmental factors (blue light, sexual interaction, retinol and aromatic chemicals) that act in independent ways [30]. In M. circinelloides, two regulatory pathways have been identified that control different aspects of carotenoid biosynthesis [15]. Thus, the induction of carotenoid biosynthesis by light is mediated by the gene mcwc-1c [9], whereas the basal levels of carotenoids are under the control of a different regulatory pathway in which crgA and mcwc-1b are the main actors [14]. However, there should be an additional unknown regulatory mechanism, as suggested by the ability of light to stimulate carotene biosynthesis in mcwc-1c–null strains, although the effect is very weak [9].
The results of this work support the existence of additional regulatory mechanisms, because eight mutants were isolated that showed altered β-carotene levels even though they lacked mutations in the genes of the two previously known regulatory pathways. Five of the mutants (MU216, MU217, MU283, MU285 and MU286) showed reduced β-carotene levels only in light, which suggests that they are affected by a regulatory pathway involved in the induction by light of carotenoid biosynthesis. However, the absence of common mutations in the sequenced genomes of three of these mutants (MU216, MU217 and MU286) points to the existence of more than one regulatory pathway, although the notion that they carry mutations in different elements of the same pathway, even the mcwc-1c regulatory pathway [9], cannot be dismissed. The other three mutants (MU206, MU218 and MU264) showed alterations of β-carotene both in the dark and in light, although MU206 and MU218 retained their ability to respond to light, which indicates that they carry mutations in genes involved in mechanisms that control the basal carotenoid levels but not its induction by light. Interestingly, MU264 showed a complete blockage of carotenoid synthesis suggesting it defines a pivotal regulatory pathway that controls carotene biosynthesis. Moreover, the absence of shared mutations amongst these mutants suggests that they define either distinct regulatory pathways or different elements of the same pathway. In fact, the genes affected in these mutants, particularly mutants with high β-carotene levels (MU206 and MU218), could code for proteins that participate in the crgA regulatory pathway. The crgA gene represses carotenoid biosynthesis by proteolysis-independent mono-ubiquitilation and di-ubiquitilation of Mcwc-1b, which activates transcription of the structural carotenogenic genes carB and carRP when it is non-ubiquitilated [15]. As a result, deletion of crgA led to significant improvement in carotene accumulation even though carotenogenesis was still induced by light [14]. The β-carotene content in the mutants MU206 and MU218 somehow resembled the phenotype of the crgA-null mutant MU221, which suggests that they could be affected in genes that participate in the same regulatory pathway as the crgA gene. In fact, MU206 and MU218 also showed higher mRNA levels of the structural carotenogenic genes than the wild-type strain (Fig. 3), which suggests that the mutations in these mutants increase the transcription of structural carotenogenic genes in a fashion similar to that of loss-of-function mutations of crgA [14].
To gain insight into the underlying regulatory mechanism altered in mutants MU206 and MU218 and its relationship with the crgA gene, this gene was disrupted in both MU206 and MU218. Disruption of the crgA gene in those mutants resulted in enhancement of both the mRNA levels of carotenogenic genes (carB and carRP) and the β-carotene content in comparison to the recipient strains. Moreover, both MU206-derived and MU218-derived mutants were able to increase both mRNA levels of carotenogenic genes and β-carotene content in response to light, which indicates that the regulatory mechanism that controls induction by light was operating in both types of mutants. However, the expression enhancement of carotenogenic genes in the MU218-derived mutants was much greater than that in the MU206-derived mutants both in the dark and in light, and it was linked to significantly greater accumulation of β-carotene. In fact, the mRNA levels of carotenogenic genes in the crgA-null mutants derived from MU218 greatly exceeded those shown by MU221 both in the dark and in light (Table 4). Even more interesting was that they doubled the amount of β-carotene present in MU221 both in the dark and in light. These results suggest that some of the mutations in MU218 affect one or some of the genes that control the expression of carotenoid genes independently of crgA, thus revealing the existence of an additional mechanism involved in the regulation of carotenoid biosynthesis in M. circinelloides. This mechanism should repress the expression of carotenoid structural genes to a lesser extent than crgA because mRNA levels of carotenogenic genes and carotenoid content are much lower in MU218 than in MU221, particularly in the dark, where there is no interference by light stimulation (Table 4). Therefore, these two regulatory mechanisms may work in parallel, and the mutants for both mechanisms showed consequently higher expression of carotenogenic structural genes and β-carotene content than mutants for just one mechanism. Unfortunately, the presence in MU218 of mutations in several genes renders analysis of the identified regulatory mechanism impossible at this time. Further work is necessary to identify the gene responsible for the phenotype of MU218 and to understand the manner in which it regulates carotenoid biosynthesis.
In contrast, although crgA mutants derived from MU206 also showed higher levels of both mRNAs of carotenogenics genes and β-carotene than MU221 both in the dark and in light, the increases were much more discrete than those seen in the crgA mutants derived from MU218 (Fig. 3; Table 4). Although the existence of a fourth regulatory pathway in the control of carotenogenesis, which is affected in MU206, cannot be discarded, it is more probable that the gene responsible for the MU206 phenotype participates in the crgA regulatory pathway. This idea is supported not only by the discrete increase in expression of carotenogenic genes and carotene content observed in the crgA mutants derived from MU206 over MU221, but also by the similarity in the patterns of β-carotene accumulation shown by MU206 and MU221 (Table 4). Therefore, it is tempting to speculate that MU206 carries a mutation in a gene that acts upstream of crgA or participates with crgA in the repression of carotenogenic structural genes. Further experiments are required to identify the gene responsible for the phenotype of MU206 and understand the relationship with crgA in the regulation of carotenogenesis.
A putative candidate for industrial production of β-carotene
As one of the essential pigments, β-carotene is the precursor of vitamin A, retinal and retinoic acid, which are of critical importance in vision, nutrition and cellular growth [31–33]. β-carotene biosynthesis has been described extensively in the zygomycetes B. trispora, P. blakesleeanus and M. circinelloides, which are promising organisms for industrial production. Compared to the other two fungi, M. circinelloides could be an interesting alternative as a biotechnological β-carotene source because of the efficient tools available for genetic manipulation. This work represents a proof of concept of this idea because the disruption of the crgA gene in mutants (MU206 and MU218) that are affected in the regulation of carotene biosynthesis generated several β-carotene–overaccumulating strains. Amongst these strains, the largest producer of β-carotene, MU606, accumulated 3001.9 ± 103.7 and 4009.23 ± 180.4 μg/g β-carotene of dry mass in the dark and in light, respectively. Therefore, it yielded approximately five and two times more β-carotene than the wild-type strain and MU221. Despite these increases in β-carotene levels, the amount of β-carotene in this strain is far from the main current industrial source of β-carotene [34], B. trispora, which is capable of producing 30 mg/g [35] by mixing two strains of opposite mating types in an optimised enrichment medium. Industrial β-carotene production using M. circinelloides could be simpler than the complicated fermentation process used with B. trispora. In addition, the yeast-like morphology of this dimorphic fungus facilitates biotechnological production [36]. Therefore, the β-carotene–overaccumulating strain obtained in this study is a strong putative candidate for industrial production of β-carotene because additional genetic manipulation or optimisation of growth conditions could generate improved strains comparable to industrial strains of B. trispora.
Conclusions
In conclusion, we adopted a classical forward strategy combined with modern reverse genetic techniques to deepen the regulation of carotenoid biosynthesis and the generation of strong candidate strains of industrial interest for β-carotene production. The results suggest the existence of a new regulatory mechanism that represses carotenoid biosynthesis independently and in parallel to the previously known repressor crgA. Like crgA, it acts at the level of mRNA accumulation keeping the transcript levels of carotenogenic genes low. Although the key gene involved in this new regulatory mechanism is pending on determination, a set of candidates has been identified by whole-genome sequencing. In addition, a double mutant for crgA and the new identified regulatory element was capable of accumulating up to 4 mg/g of β-carotene, being a candidate to be target of further improvements for biotechnological β-carotene production.
Methods
Strains, plasmids and transformation conditions
The strains generated in this work were derived from R7B, which is a leucine auxotroph of M. circinelloides f. lusitanicus CBS277.49 [37] that keeps a wild-type phenotype for carotenoid biosynthesis [25]. Spores of this strain were mutagenized with UV light (50 mJ/cm2) or NTG in conditions that produced a 1–5 % survival rate as previously described [25]. Due to the multinucleate state of R7B spores, spores were allowed to complete a vegetative cycle in complete medium to unveil the expression of recessive mutations. To reduce the probability to isolate the same mutant several times, spores to be mutagenized were split in different tubes and kept separately after recycling.
The strains MU241 and MU221 were also derived from R7B; MU221 is a crgA-null mutant, and MU241 is a prototrophic wild-type strain after replacement of the previous leuA mutant allele with the wild-type allele [14]. Plasmid pLEU4 [37], which is an auto-replicative plasmid that harbours a wild-type leuA allele, was used to complement the leuA auxotrophy of the recipient strains. Plasmid pVEN172 [14] that contains a 6.5-kb PstI fragment of the wild-type allele of leuA gene flanked by DNA regions adjacent to the crgA gene was used to disrupt the crgA gene of MU206 and MU218. Cultures were grown in minimal medium (YNB) [38] or complete medium (YPG) [39]. l-leucine (20 μg/ml) was supplied when required. The transformation procedure has been described previously [40].
Nucleic acid manipulation and analysis
Genomic DNA of M. circinelloides was prepared as described previously [41]. Plasmid was isolated by the rapid boiling method [42] and purified following the manufacturer’s instructions (Thermo Scientific, USA). The presence of mutations in carotenogenic genes in the mutants was analysed by PCR amplification of carB, carRP, mcwc-1c and mcwc-2 genes (ID78608, ID85699, ID153812 and ID157130) using the primers listed in Additional file 7: Table S5 and PfuUltra II Fusion HS DNA Polymerase (Agilent). The PCR fragments were directly sequenced without cloning to avoid interference with mutations produced during PCR, and the obtained sequence was compared to the reference genome of M. circinelloides CBS277.49 (http://www.genome.jgi.doe.gov/Mucci2/Mucci2.info.html).
Linear DNA fragments for transformation were released by treating the plasmid pVEN172 with PstI, and then purified from agarose gel with GeneJET Gel Extraction Kit (Thermo Scientific, USA). To determine whether the fragment was integrated into the genome correctly, PCR amplification in an “Eppendorf Mastercycler personal” was performed with genomic DNA as a template and Herculase II fusion DNA polymerase (Agilent Technologies, USA) following the manufacturer’s instructions. Primers cigC-F1 and crgA-R1 were designed for amplification of the recombinant fragment (Additional file 7: Table S5). The samples were incubated at 95 °C for 3 min for pre-denaturation and subjected to 30 cycles of denaturation (95 °C for 30 s), annealing (55 °C for 30 s) and extension (72 °C for 3 min 30 s) with an additional 5-min extension step.
The transcription level of the structural genes carB and carRP was analysed with the Droplet Digital PCR technique (ddPCR). Total RNA was isolated using Trizol reagent following the instructions of the manufacturer (Invitrogen). Reverse transcription was performed with PrimeScript RT Reagent Kit with gDNA Eraser (Takara), and the ddPCR experiments were carried out with QX200™ ddPCR™ EvaGreen Supermix (BioRad). The PCR reaction mix was partitioned into water-in-oil emulsion droplets with a QX100 Droplet Generator. The droplets were then transferred to a 96-well PCR plate and put into a thermal cycler (Bio Rad C1000). Thermal cycling conditions were [95 °C × 5 min; 40 cycles of (95 °C × 0.5 min, 60 °C × 1 min); 4 °C × 5 min; 98 °C × 5 min (ramp rate set to 2 °C/s)]. The PCR plate was then moved to the QX100 Droplet reader for automatic reading of samples. The copy numbers of carB and carRP genes from various samples were normalised to the actin reference gene (ID157034; scaffold_07: 2052804–2054242).
For whole-genome sequencing of the DNA of carotenogenic mutants, genomic DNA was isolated by cesium chloride gradient [43], sequenced using SOLiD technology and analysed at the Centre for Genomic Research in the University of Liverpool. A sample of mutations was validated by direct sequencing of PCR products from mutated genes in strains MU206, MU264 and MU286 using specific primers (Additional file 7: Table S5). The sequence reads have been deposited in the European Nucleotide Archive and the accession number is PRJEB13169.
Analysis of carotenes
Carotenes were extracted from mycelia grown on YNB solid medium or YNB supplemented with leucine for leucine auxotrophic strains, for 84 h in the dark and 60 h in the dark followed by 24 h in light at 26 °C. The mycelia were washed and dried between paper towels and triturated with a pestle in the presence of liquid nitrogen. After lyophilisation, 2 mg of the freeze-dried mycelium powder was weighed, suspended in 2 ml of methanol and extracted several times with 2 ml of petroleum until the mycelium was colourless [44]. For spectrophotometric quantification of β-carotene, the absorption coefficients given by Davies [45] were applied.
To HPLC analysis of the carotenoids, the organic phase extracted as described above were mixed and evaporated at 30 °C in vacuum. All samples were suspended in 500 µL ethyl acetate and filtered (0.22 μm) prior to HPLC with diode-array detection HPLC–DAD analysis. Carotenoids were identified and quantified using a column ZORBAX Eclipse XDB-C18 end capped 5 μm, 4.6 × 150 mm reverse phase column (Agilent Technologies, USA). The eluents used were (A) acetonitrile: water (9:1, v/v) and (B) ethyl acetate. The column separation was allowed via a series of gradient such as follows: 0–40 % solvent B (0–20 min), 40–60 % solvent B (20–25 min), 60–100 % solvent B (25–25, 1 min), 100 % solvent B (25, 1–35 min) and 100–0 % solvent B (35–50 min) at a flow rate of 1 ml/min. The injection volume was 15 μl each one. Detection of individual carotenoids was made at 284 and 453 nm, which are the wavelength of maximum absorption of phytoene and β-carotene. These compounds were identified by co-chromatography with standards and by elucidation of their spectral characteristics using a photo-diode array detector. The concentrations of carotenoids were estimated on basis of an adequated standard curve using their respective standard compounds [46].
Statistical analysis
All experiments were carried out independently at least three times, and the mean values ± SD were presented. Statistical analysis was performed with one-way analysis of variance with SPSS 20.
References
Sandmann G. Carotenoids of biotechnological importance. In: Schrader J, Bohlmann J, editors. Biotechnology of isoprenoids. Switzerland: Springer; 2015. p. 449–67.
Britton G, Liaaen-Jensen S, Pfander H. Carotenoids volume 5: nutrition and health. Basel: Birkhäuser; 2009.
Hussein G, Sankawa U, Goto H, Matsumoto K, Watanabe H. Astaxanthin, a carotenoid with potential in human health and nutrition. J Nat Prod. 2006;69:443–9.
Crotenoid markert by values. In: The global market for carotenoids. 2015. http://www.bccresearch.com/market-research/food-and-beverage/carotenoids-global-market-report-fod025e.html. Accessed Jul 2015.
Lamers PP, Janssen M, De Vos RC, Bino RJ, Wijffels RH. Exploring and exploiting carotenoid accumulation in Dunaliella salina for cell-factory applications. Trends Biotechnol. 2008;26:631–8.
Feofilova EP. Fungal carotenoids: biological functions and practical use. Prikl Biokhim Mikrobiol. 1993;30:181–95.
Torres-Martínez S, Ruiz-Vázquez RM, Garre V, López-García S, Navarro E, Vila A. Molecular tools for carotenogenesis analysis in the zygomycete Mucor circinelloides. Methods Mol Biol. 2012;898:85–107.
Ye VM, Bhatia SK. Pathway engineering strategies for production of beneficial carotenoids in microbial hosts. Biotechnol Lett. 2012;34:1405–14.
Silva F, Torres-Martínez S, Garre V. Distinct white collar-1 genes control specific light responses in Mucor circinelloides. Mol Microbiol. 2006;61:1023–37.
Lauter F-R, Russo VEA. Blue light induction of conidiation-specific genes in Neurospora crassa. Nucleic Acids Res. 1991;19:6883–6.
Jayaram M, Presti D, Delbrück M. Light-induced carotene synthesis in Phycomyces. Exp Mycol. 1979;3:42–52.
Cheng P, Yang Y, Wang L, He Q, Liu Y. WHITE COLLAR-1, a multifunctional neurospora protein involved in the circadian feedback loops, light sensing, and transcription repression of wc-2. J Biol Chem. 2003;278:3801–8.
He Q, Liu Y. Molecular mechanism of light responses in Neurospora: from light-induced transcription to photoadaptation. Genes Dev. 2005;19:2888–99.
Navarro E, Lorca-Pascual JM, Quiles-Rosillo MD, Nicolás FE, Garre V, Torres-Martínez S, Ruiz-Vázquez RM. A negative regulator of light-inducible carotenogenesis in Mucor circinelloides. Mol Genet Genomics. 2001;266:463–70.
Silva F, Navarro E, Penaranda A, Murcia-Flores L, Torres-Martínez S, Garre V. A RING-finger protein regulates carotenogenesis via proteolysis-independent ubiquitylation of a white collar-1-like activator. Mol Microbiol. 2008;70:1026–36.
Kaiser P, Flick K, Wittenberg C, Reed SI. Regulation of transcription by ubiquitination without proteolysis.: Cdc34/SCF(Met30)-mediated inactivation of the transcription factor Met4. Cell. 2000;102:303–14.
Sanz C, Rodríguez-Romero J, Idnurm A, Christie JM, Heitman J, Corrochano LM, Eslava AP. Phycomyces MADB interacts with MADA to form the primary photoreceptor complex for fungal phototropism. Proc Natl Acad Sci USA. 2009;106:7095–100.
Kubo H. Isolation of madA homologs in Pilobolus crystallinus. Mycoscience. 2009;50:400–6.
Herrera-Estrella A, Horwitz BA. Looking through the eyes of fungi: molecular genetics of photoreception. Mol Microbiol. 2007;64:5–15.
He Q, Cheng P, Yang Y, Wang L, Gardner KH, Liu Y. White collar-1, a DNA binding transcription factor and a light sensor. Science. 2002;297:840–3.
Mehta BJ, Salgado LM, Bejarano ER, Cerda-Olmedo E. New mutants of Phycomyces blakesleeanus for (beta)-carotene production. Appl Environ Microbiol. 1997;63:3657–61.
Revuelta JL, Eslava AP. A new gene (carC) involved in the regulation of carotenogenesis in Phycomyces. Mol Gen Genet. 1983;192:225–9.
Bejarano ER, Parra F, Murillo FJ, Cerdá-Olmedo E. End-product regulation of carotenogenesis in Phycomyces. Arch Microbiol. 1988;150:209–14.
Almeida ER, Cerda-Olmedo E. Gene expression in the regulation of carotene biosynthesis in Phycomyces. Curr Genet. 2008;53:129–37.
Navarro E, Sandmann G, Torres-Martı́nez S. Mutants of the carotenoid biosynthetic pathway of Mucor circinelloides. Exp Mycol. 1995;19:186–90.
Linden H, Macino G. White collar 2, a partner in blue-light signal transduction, controlling expression of light-regulated genes in Neurospora crassa. EMBO J. 1997;16:98–109.
Roncero MIG, Jepsen LP, Stroman P, van Heeswijck R. Characterization of a leuA gene and an ARS element from Mucor circinelloides. Gene. 1989;84:335–43.
Appel KF, Wolff AM, Arnau J. A multicopy vector system for genetic studies in Mucor circinelloides and other zygomycetes. Mol Genet Genomics. 2004;271:595–602.
Hindson BJ, Ness KD, Masquelier DA, Belgrader P, Heredia NJ, Makarewicz AJ, Bright IJ, Lucero MY, Hiddessen AL, Legler TC, et al. High-throughput droplet digital PCR system for absolute quantitation of DNA copy number. Anal Chem. 2011;83:8604–10.
Cerdá-Olmedo E. Phycomyces and the biology of light and color. FEMS Microbiol Rev. 2001;25:503–12.
Manville IA. Nutrition and night blindness. Northwest Med. 1939;38:208–13.
Ross AC. Vitamin A. In: Milner JA, Romaglono DF, editors. Bioactive compounds and cancer. New York: Springer; 2010. p. 335–56.
Ross AC, Gardner EM. The function of vitamin A in cellular growth and differentiation, and its roles during pregnancy and lactation. In: Allen L, King J, Lonnerdal B, editors. Nutrient regulation during pregnancy, lactation, and infant growth. New York: Springer; 1994. p. 187–200.
Kumar A, Vishwakarma HS, Singh J. Microbial pigments: production and their applications in various industries. IJPCBS. 2015;5(1):203–12.
Dufosse L. Microbial production of food grade pigments. Food Technol Biotech. 2006;44:313–21.
McIntyre M, Breum J, Arnau J, Nielsen J. Growth physiology and dimorphism of Mucor circinelloides (syn. racemosus) during submerged batch cultivation. Appl Microbiol Biot. 2002;58:495–502.
Schipper MA. Induced azygospore formation in Mucor (Rhizomucor) pusillus by Absidia corymbifera. Anton Leeuw. 1976;42:141–4.
Dent JG, Graichen ME, Schnell S, Lasker J. Constitutive and induced hepatic microsomal cytochrome P-450 monooxygenase activities in male Fischer-344 and CD rats. Toxicol Appl Pharmacol. 1980;52:45–53.
Bartnicki-Garcia S, Nickerson WJ. Isolation, composition, and structure of cell walls of filamentous and yeast-like forms of Mucor rouxii. Biochim Biophys Acta. 1962;58:102–19.
Gutiérrez A, López-García S, Garre V. High reliability transformation of the basal fungus Mucor circinelloides by electroporation. J Microbiol Methods. 2011;84:442–6.
Ruiz-Pérez VL, Murillo FJ, Torres-Martínez S. PkpA, a novel Phycomyces blakesleeanus serine/threonine protein kinase. Curr Genet. 1995;28:309–16.
Holmes DS, Quigley M. A rapid boiling method for the preparation of bacterial plasmids. Anal Biochem. 1981;114:193–7.
Sambrook J, Russell DW. Molecular cloning: a laboratory manual. Cold Spring Harbor: Cold Spring Harbor Laboratory Press; 2001.
Govind NS, Cerdá-Olmedo E. Sexual activation of carotenogenesis in Phycomyces blakesleeanus. J Gen Microbiol. 1986;132:2775–80.
Davies BH. Carotenoids. In: Holden M, Goodwin TW, editors. Chemistry and biochemistry of plant pigments. London: Academic Press; 1976. p. 38–165.
Fatimah AMZ, Norazian MH, Rashidi O. Identification of carotenoid composition in selected ‘ulam’ or traditional vegetables in Malaysia. Int Food Res J. 2012;19:527–30.
Authors’ contributions
YZ participated in knocking out crgA gene, isolation mutants, carotene content analysis of mutants and the manuscript writing. EN participated in the isolation of carotenogenic mutants, performed their phenotypic characterization, analyzed their genomic sequences, validated the mutations and supervised the complementation experiments. JTCM performed the complementation experiments. LA participated in the HPLC analysis of carotenoids. HC participated in analysis of carotene content. YQC and HZ helped conceive and coordinate the study. STM participated in the isolation of carotenogenic mutants. WC supervised the experiments. VG designed and supervised the experiments and participated in writing of the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We thank J. A. Madrid for technical assistance. This work was funded by the Fundación Séneca (Comunidad Autónoma de la Región de Murcia, Spain), Project no. 08802/PI/08, the Program for New Century Excellent Talents (NCET-13-0831), the Program for Changjiang Scholars and Innovative Research Team in University (IRT1249).
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Yingtong Zhang and Eusebio Navarro contributed equally to this work
Additional files
12934_2016_493_MOESM1_ESM.pptx
Additional file 1: Figure S1. Spectrophotometric scans from 350 nm to 550 nm of total carotenoid extracts from the indicated strains. Absorbance scales are different in each sample.
12934_2016_493_MOESM2_ESM.pptx
Additional file 2: Figure S2. HPLC elution profile of carotenoids accumulated by the indicated strains in light. Carotenoids were detected at 453 nm.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Zhang, Y., Navarro, E., Cánovas-Márquez, J.T. et al. A new regulatory mechanism controlling carotenogenesis in the fungus Mucor circinelloides as a target to generate β-carotene over-producing strains by genetic engineering. Microb Cell Fact 15, 99 (2016). https://doi.org/10.1186/s12934-016-0493-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-016-0493-8