
Abstract
Background
The potential of essential oils (EOs) and of their principal constituents for eradication of biofilm and at the same time the research of new potential acetylcholinesterase inhibitors is gaining increasing interest in last years. The aims of this study were to determine the chemical composition and to evaluate the antibacterial, cytotoxic, and anti-acetylcholinesterase properties of Myrtus communis leaves essential oil and its main constituents.
Methods
Essential oil was obtained by hydrodistillation of M. communis L. leaves and was analyzed by GC and GC–MS. The antimicrobial activity was carried out against both gram-negative and gram-positive bacteria. The microdilution method was used to estimate the minimum inhibitory concentrations (MICs). Then, the capacity of essential oil and its main constituent to inhibit biofilm growth, with the method of O’Toole and Kolterand, and the metabolic activity of biofilm cells through the MTT colorimetric method were evaluated at different times. Moreover, was studied the potential cytotoxic activity against SH-SY5Y cell line with MTT assay and the anti-acetylcholinesterase activity using Ellman’s assay.
Results
Myrtenyl-acetate, 1,8 cineole, α-pinene, and linalool were the main components in the EO. The myrtle EO, at the minimum tested dose (0.4 mg/ml), inhibited S. aureus biofilm by 42.1% and was capable of inhibiting the biofilm cell metabolism in all tested strains, except Staphylococcus aureus. Moreover, the EO showed good cytotoxic and anti-acetylcholinesterase activities IC50 of 209.1 and 32.8 μg/ml, respectively.
Conclusions
The results suggest that myrtle EO and its main constituents could be used as possible products that could act against the resistant pathogenic species E. coli, P. aeruginosa, L. monocytogenes and S. aureus, on the other hand, as possible coadjutants in the treatment of neurological diseases.
Similar content being viewed by others
Background
Myrtle (Myrtus communis L., Myrtaceae) is a plant used worldwide in traditional medicine. It is a spontaneous evergreen shrub or a small tree of Mediterranean area, growing along the coasts, on the islands, and the internal hill [1, 2]. The essential oils (EOs) derived from fresh leaves and/or berries are used as food flavoring (meat, sauces, etc.), for spirits and in the perfume and cosmetic industry [3]. Moreover, in folk medicine, these EOs have an important role in the treatment of gastrointestinal diseases for their astringent proprieties and are used as antimicrobial, antioxidant, and anti-inflammatory agents [4]. Several studies reported that M. communis essential oil and its principal components, such as 1,8-cineole, linalool, eugenol, α-terpineol and γ-terpinene, were active against both Gram-positive and Gram-negative bacteria [5,6,7].
Biofilms are constituted by microbial cells absorbed in a matrix of extracellular polymeric substances. These cells can cause chronic infections, persistent inflammation, and tissue damage [8] and are very different from planktonic cells. In fact, the biofilm is constituted in well-organized hierarchically communities that protect the cells from adverse environmental conditions and antibacterial agents [9, 10].
The potential of EOs and their principal constituents for the eradication of biofilm is gaining increasing interest in recent years as new antibacterial agents, especially for the increasing emergence of bacterial resistance [10, 11]. This phenomenon, as well as the occurrence of the bacterial transformation giving rise to more aggressive bacteria, represent two problems of great impact on human health. Such aspects are encouraging the research toward the identification of new antibacterials, also from natural sources, which can represent alternative and more efficient modes of treatment with respect to the conventional antibacterial drugs, especially when people are faced with particularly aggressive and resistant bacteria. For example, infections by Staphylococcus aureus and Pseudomonas aeruginosa, mainly through biofilm formation, is a problem of great relevance. These infections, carrying on antigen presentation, lead to chronic inflammation and makes common eradication treatments less effective [12]. Similarly acts the uropathogenic Escherichia coli; in fact, its biofilms are problematic to remove from the surface of hospital catheters [13], and Listeria monocytogenes, able to infect food products also by the formation of biofilms [14]. Also, plants are subjected to bacterial infection: for example, the biofilm formed by the Gram-negative Pectobacterium carotovorum causes soft rot in some food plants due to the release of exo-enzymes and increases bacterial resistance during plant disinfection [15].
Different EOs have already been tested in an attempt to find potential inhibitors of the formation of biofilm [16,17,18]. Conversely, few studies are reported on the identification of those extracts and/or EOs capable of affecting the metabolism of the cells present in the biofilm organization, which profoundly differ from planktonic cells of the same species due to complex phenotypic and metabolic changes that regulate some cell events, such as adhesion, sporulation, starvation survival, rough-smooth phase variations, etc. This multifaceted scenario also led to differences in the susceptibility of planktonic and biofilm bacterial cells to antimicrobial agents.
Since the 1990s, a very promising field of application of EOs is the study of their cytotoxic properties against several cancer cell lines [19]. Nevertheless, few studies reported the cytotoxicity of the EO of M. communis against several cell lines: human cervix adenocarcinoma (HeLa), human breast cancer (MCF-7), Mus musculus mastocytoma cells (P815), and human colon colorectal adenocarcinoma (HT29) [20,21,22]. Until now, no research has been carried out on the possible cytotoxicity of M. communis or of any plants belonging from Myrtaceae family against neuroblastoma, the most common tumor among children less than one year of age [23].
Moreover, in the last years, the research of natural substances for therapeutic aims highlights a potential source of acetylcholinesterase (AChE) inhibitors in plants [24]. This interest derives from the traditional use of several plants to treat neurodegenerative diseases [25]. Several EOs and their constituents have been investigated for their inhibition activity of AChE [26], but only one study reported the possible anti-acetylcholinesterase action of myrtle EO [27].
Therefore, the present study was carried out to determine the chemical composition of the EO from leaves of M. communis leaves and to investigate on antimicrobial, antibiofilm, cytotoxic, and anti-acetylcholinesterase activities of myrtle EO and its main constituents.
Materials and methods
Plant material
Leaves of M. communis were collected in Bellosguardo (Salerno, Italy) in February 2020. The plant was identifiedby Prof. Vincenzo De Feo, full professor at University of Salerno . A voucher specimen (DF/2020/314) is stored in the herbarium of the Medical Botany Laboratory, University of Salerno. The use of M. communis L. leaves in the present study compiles with international guidelines; permissions or licenses were not necessary to collect this plant species.
Isolation of essential oil
Fresh leaves were subjected to steam distillation for 3 h according to the standard procedure described by the European Pharmacopoeia [28]. The distillation furnished yellow oil in 0.9 % yield on a dry mass basis. The oil was solubilized in n-hexane, filtered over anhydrous sodium sulphate, and stored under N2 at +4 °C in dark-sealed vial until analysis.
GC-FID analysis
Analytical gas chromatography was performed on a Perkin-Elmer Sigma-115 gas chromatograph equipped with a FID and a data handling processor. The separation was achieved using a HP-5 MS fused-silica capillary column (30 m × 0.25 mm i.d., 0.25 µm film thickness). Column temperature: 40 °C, with 5 min initial hold, and then to 270 °C at 2 °C/min, and finally at 270 °C (20 min); injection mode splitless (1 µL of a 1:1000 n-hexane solution). Injector and detector temperatures were 250 °C and 290 °C, respectively. Analysis was also run by using a fused silica HP Innowax polyethylene glycol capillary column (50 m × 0.20 mm i.d., 0.25 µm film thickness). In both cases, helium was used as carrier gas (1.0 ml/min) [29].
GC/MS analysis and identification of constituents
GC/MS analyses were performed on an Agilent 6850 Ser. II apparatus, fitted with a fused silica DB-5 capillary column (30 m × 0.25 mm i.d., 0.33 µm film thickness), coupled to an Agilent Mass Selective Detector MSD 5973; ionization energy voltage 70 eV; electron multiplier voltage energy 2000 V. Mass spectra were acquired in the range 40–500 amu, scan time 5 scans/s. Gas chromatographic conditions were as reported above; transfer line temperature, 295 °C. Most constituents were identified by comparison of their Kovats retention indices (Ri) [calculated in relation to a series of n-alkanes (C10–C35)], with either those of the literature [30,31,32,33], by accurate analysis of mass spectra on both columns and by their comparison with those of authentic compounds available in our laboratories by means of NIST 08 and Wiley 275 libraries. The components’ relative concentrations were obtained by peak area normalization.
Microorganisms and culture conditions
Three Gram negative (E. coli DSM 8579, P. aeruginosa ATCC 50071, P. carotovorum DSM 102074) and two Gram positive (S. aureus DMS 25923, L. monocytogenes ATCC 7644) bacterial strains, provided by DSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany) were used. Bacteria were grown in Luria Bertani (LB) broth (Sigma, Milano, Italy) for 18 h at 37 °C and 80 rpm (Corning LSE, Pisa, Italy). P. carotovorum was grown at 28 °C and 80 rpm. Bacteria were serially diluted in sterile physiological solution and spread onto Muller-Hinton agar plates. Using an aseptic method, a single colony of each strain was transferred into a 10 ml sterile tube containing the iso-sensitized broth and placed for the growth in incubator for 18 h at 80 rpm (Corning) and different temperatures depending on the strain. Pellet was washed and resuspended in 10 ml sterile saline solution. The optical density was recorded at 600 nm, and serial dilutions were performed to ensure an optical density at 0.5.
Minimal Inhibitory Concentration (MIC)
The Minimal Inhibitory Concentration (MIC) values were calculated following the method of Saker and coworkers [34] and Fratianni and coworkers [35] on microtiter-plates. A resazurin solution, used as an indicator solution, was prepared by dissolving 270 mg in 40 ml of sterile distilled water. Through the use of a vortex mixer, the indicator was perfectly dissolved so to have a homogeneous solution. The EO, and its main components, myrtenyl acetate, 1,8 cineole, α-pinene, and linalool, were dissolved in sterile DMSO. Two-fold serial dilutions were prepared to obtain 50 μL of EO, myrtenyl acetate, 1,8 cineole, α-pinene, and linalool, in serially descending concentrations in each well. Thirty-five μL of 3.3 × strength iso-sensitized broth and 5 μL of resazurin solution were supplemented, to reach a final volume/well of 240 μL with different volumes of sterile Muller-Hinton broth (Sigma-Aldrich, Milano, Italy) previously set. Lastly, 10 μL of bacterial suspension were added to each well to get a concentration of about 5 × 105cfu/ml, to obtain a final volume of 250 μL. Sterile DMSO and tetracycline (dissolved in DMSO, 1 mg/ml) were used as negative and positive control, respectively. Multiwell plates were prepared in triplicate and incubated at 37 °C for 24 h. The color changes were assessed visually. The lowest concentration at which the color change (from dark purple to colorless) was visualized, indicated the MIC value that thus represented also the lowest concentration of compound giving rise to a marked reduction in the appearance of growth compared to the growth control.
Biofilm inhibitory activity
The effects of the EO and its main components, myrtenyl acetate, 1.8 cineole, α-pinene, and linalool, at concentrations ranging from 0.4 to 2.0 mg/ml, were evaluated using the method of Fratianni and coworkers [35] using 96-well microtiter plates. In each well, the overnight bacterial cultures were adjusted to 0.5 McFarland with fresh culture broth. Ten µL of the diluted cultures were distributed in each well; then different volumes of the samples and Muller-Hinton broth were added, to reach a final volume of 250 µL/well. Microplates were completely covered with parafilm to avoid the evaporation of samples with relative loss of volume and incubated for 48 h at 37 °C (except P. carotovorum that was incubated at 28 °C). Planktonic cells were removed, and the attached cells were gently washed twice with sterile physiological saline. After that, 200 µL of methanol were added to each well, retaining it for 15 min to fix the sessile cells. After discharge of methanol, each plate was left until complete dryness of samples. Staining of the adhered cells was performed by adding 200 µL of 2% w/v crystal violet solution to each well. After 20 min, wells were gently washed with the sterile physiological solution and left to dry. Two hundred microliters of glacial acetic acid 20% w/v were added to allow the release of the bound dye. The absorbance was measured at OD = 540 nm (Varian Cary Spectrophotometer model 50 MPR, Cernusco sul Naviglio, Italy). The percent value of biofilm inhibition was calculated respect to control (cells grown without the presence of the samples). Triplicate tests were done, and the average results were taken for reproducibility.
Metabolic activity of biofilm cells
The effects of different concentrations of the myrtle EO, and its main components, myrtenyl acetate, 1.8 cineole, α-pinene and linalool, ranging from 0.4 to 2.0 mg/ml on the metabolic activity of biofilm cells was evaluated through the MTT colorimetric method, using 96-well microtiter plates [35] The overnight bacterial cultures, grown at the due temperatures, were adjusted to 0.5 McFarland and treated as above described. After 48 h incubation, bacterial suspension was removed, and 150 µL of PBS and 30 µL of 0.3% MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, Sigma, Milan, Italy] were added, keeping microplates at 37 °C. After 2 h, MTT solution was removed and, after two washing steps with 200 μL of sterile physiological solution, 200 µL of DMSO were added to allow the dissolution of the formazan crystals, which were measured at OD = 570 nm (Varian Cary Spectrophotometer model 50 MPR, Cernusco sul Naviglio, Italy). Triplicate tests were done and the average results were taken for reproducibility.
MTT assay
Human neuroblastoma (SH-SY5Y) cancer cells purchased from ATCC Bioproducts (ATCC, Manassas,VA, USA) were cultured in Roswell Park Memorial Institute Medium (RPMI) supplemented with 1% L-glutamine, 10% heat-inactivated fetal bovine serum (FBS),1% penicillin/streptomycin (all from Sigma Aldrich) at 37°C in an atmosphere of 95% O2 and 5% CO2.
Cells were plated (5 × 103) in 96-well culture plates in 150 µL of culture medium and incubated at 37 °C in a humidified atmosphere of 95% O2 and 5% CO2. The day after, a 150 µL aliquot of serial dilutions of the EO and its main constituents (500–25 µg/ml) was added to the cells and incubated for 24 h. DMSO was used as a control. Cell viability was assessed through MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide) assay. Briefly, 30 µL of MTT (5 mg/ml) were added and the cells were incubated for an additional 3 h. After that, cells were lysed, and the dark blue crystals were solubilized with 30 µL of a solution containing 50%, v/v, N,N-dimethylformamide, 20%, w/v, SDS with an adjusted pH of 4.5. The optical density (OD) of each well was measured with a microplate spectrophotometer (Thermo Scientific Multiskan GO, Monza, Italy) equipped with a 520 nm filter. Cell viability in response to treatment was calculated as a percentage of control cells treated with DMSO at the final concentration 0.1% [36].
Anti-acetylcholinesterase activity
AChE inhibitory activity assay was performed according to a previously described spectrophotometric method [37] with minor modifications [38]. Briefly, in a total volume of 1 ml, 415 µL of Tris-HCl buffer 0.1 M (pH 8), 10 µL of different concentrations of extract dissolved in methanol, and 25 µL of AChE solution (0.28 U/ml) were incubated for 15 min at room temperature. Seventy-five microliters of a solution of AChI (1.83 mM) and 475 µL of DTNB were added, and the final mixture was incubated for 30 min at room temperature.
Absorbance was measured at 405 nm by a spectrophotometer (Thermo Scientific Multiskan GO, Monza, Italy). Galanthamine was used as a positive control. Bidistilled water, instead of the EO or galanthamine, was used as a negative control. The inhibition rate (%) of AChE activity was calculated by comparison with the negative control by using the following equation:
Statistical analysis
All experiments were carried out in triplicate. Data of antimicrobial, cytotoxic, and anti-acetylcolinesterase activities were statistically analyzed using GraphPad Prism 6.0 software (GraphPad Software Inc., San Diego, CA, United States) followed by the comparison of means (two-way ANOVA) using Dunnett’s multiple comparison test, at the significance level of p < 0.05.
Results
Phytochemical analyses
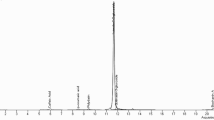
Hydrodistillation of three samples of the leaves of M. communis provided pale-yellow oils in 0.33 ± 0.05% yield on a dry mass basis. The GC profile of essential oil is present in Fig. 1. Table 1 shows the chemical composition of the EO; compounds are listed according to their elution order on a HP-5MS.

GC–MS chromatogram of the leaves essential oil of M. communis L
Altogether, 59 compounds were identified, accounting for 98.4% of the total oil. Oxygenated monoterpenes were the main components (71.7%), followed by monoterpenes hydrocarbons (18.6%) and sesquiterpenes hydrocarbons (2.6%). Myrtenyl-acetate (29.8%), 1,8-cineole (21.9%), α-pinene (14.7%) and linalool (9.1%) were the main constituents. Other compounds, in a lesser amount, were heptyl isobutanoate (3.2%), geranyl-acetate (2.6%), α-terpineol (2.3%), (Z)-caryophyllene (1.3%) and α-humulene (1.1%).
Antibacterial activity
Through the resazurin test, the minimal inhibitory concentration (MIC) necessary to block the growth of the bacteria used as tester strains was evaluated. Results are shown in Table 2.
The whole EO exhibited an effective inhibitory activity that did never exceed 6 mg/ml (in E. coli). The four main components of the EO showed a MIC higher than of the EO, and in some cases, values reach up 10 mg/ml. The whole EO perhaps could act as an antibacterial agent due to the synergistic action of more components that, considered individually, did not exhibit the same potency.
The capacity of the EOs to block or limit the formation of biofilm (through the use of the crystal violet assay) and the effect of the EOs on the bacterial cell metabolism (observed by the MTT test) were also evaluated. Results are shown in Tables 3 and 4, respectively.
The addition of the different doses (final concentration 0.4, 1, and 2 mg/ml, calculated after the MIC analysis), was carried out at three different times: at zero time before the formation of the biofilm (to evaluate the ability of the EO and its main components to block the formation of biofilm ab origine); after 24 h growth (to evaluate the capacity to act on mature biofilm) and after 48 h growth (when the biofilm is extremely mature); therefore, in the presence of different metabolic and microbial cell growth conditions (Table 3).
At zero time, the addition of the myrtle EO determined, at the highest concentration, a biofilm-inhibitory action, ranging between 26.25% (against E. coli) up to 66.67% (against P. aeruginosa). Interestingly, at the lowest concentration (0.4 mg/ml) the oil was practically ineffective against P. carotovorum, P. aeruginosa and showed very low effectiveness against L. monocytogenes and E. coli.
The influence exerted by the single components on the whole EO activity seemed different according to the bacterial species. Thus, α-pinene and linalool showed an inhibitory biofilm activity at 2 mg/ml, equal to 20.76% and 33.68%, respectively. On the other hand, the inhibitory biofilm action of the EO against L. monocytogenes could be due to α-pinene and myrtenyl acetate.
All EO components were active against P. carotovorum, except myrtenyl acetate, which showed an inhibitory biofilm activity only of 2.08%. On the contrary, the four main components resulted both in blocking or limiting the formation of the biofilm by P. aeruginosa and S. aureus. In the latter case, α-pinene determined an inhibitory efficacy of 91.39%.
The addition of the EO after 24 hours did not determine an effective inhibition of the biofilms. E. coli was completely insensitive to myrtle EO. Similar behavior was observed in the case of P. aeruginosa (inhibitory efficacy = 4.97%), P. carotovorum (inhibitory efficacy = 2.8%), and S. aureus (inhibitory efficacy of only 1.1%). Only L. monocytogenes still seemed, albeit weakly, sensitive to the presence of the myrtle EO (15.99%).
E. coli was completely insensitive to the presence of linalool and α-pinene (which instead were effective when added at zero time). L. monocytogenes seemed influenced by α-pinene (efficacy = 28.98%) and linalool, but not by myrtenyl acetate as at time zero. The addition of EO main components the 24 h mature P. aeruginosa biofilm had a low inhibitory efficacy ranging from 6.33% for myrtenyl acetate to 16.61% for 1,8 cineole. The best inhibitory efficacy on the 24 h mature S. aureus biofilm was showed by myrtenyl acetate (inhibitory efficacy = 50.62 % at 2 mg/ml).
The activity of the EO against the 48 h bacterial biofilms revealed a still different situation. An efficacy boost of the EO, which acted against E. coli with an inhibitory efficacy of 42.49% was observed. Only S. aureus returned to be sensitive to the action of the whole EO, albeit with a low efficacy (22.06%), but the four main components showed much more efficacy ranging between 36.48% (α-pinene) and 64.66% (linalool). Also, against 48 h biofilm of L. monocytogenes the EO was less active (4.09%), than linalool that exhibited, if tested alone, a strong inhibition (75.77%).
A similar setting was observed with regard to P. carotovorum on which the EO exhibited an efficacy of 8.29%, although three of the four main components showed an efficacy ranging between 23.70% (myrtenyl acetate) and 36.81% (1.8 cineole) against this ultra-mature biofilm.
P. aeruginosa was completely insensitive to the action of the EO, although some of the main components were capable of acting even partially on the 48 h biofilms.
All these data seem to indicate that the inhibitory efficacy was very strong if the EO was added at zero time, with a strong inhibition of biofilm formation. When the EO was added to 24 h biofilm, its effectiveness decreased drastically and increased again in 48 h biofilm.
A different scenario appeared when the effects of EO myrtle and its main components were evaluated on the metabolism of microbial cells, in the same at zero 24h and 48 h times, namely in the absence and in the presence of a mature or ultra-mature biofilm, respectively. The results are shown in Table 4.
The addition of the EO at time zero determined a different behavior on bacterial cells. It was effective against four of the five microorganisms tested and, at the highest concentration, the inhibitory effect on cellular metabolism ranged between 37.59% (E. coli) and 68.06% (L. monocytogenes). S. aureus was the only resistant strain.
In E. coli, myrtenyl acetate and linalool had a good activity (metabolic inhibition = 61.34% instead 1,8 cineole resulted completely ineffective.
As regards L. monocytogenes, it could be hypothesized that the bacterial metabolism was influenced by all four components, which showed an inhibitory efficacy ranging between 41.73% (myrtenyl acetate) and 67.71% (α-pinene). α-Pinene (36.81%) and 1,8 cineole (35.65 %) had lower efficacy than EO vs P. carotovorum; in fact, the EO showed an inhibitory efficacy on microbial metabolism of 60.82%. The influence of the myrtenyl acetate against P. aeruginosa resulted in a good inhibitory metabolic activity (96.99%). Instead, 1,8 cineole was completely ineffective. Instead, against the metabolism of L. monocytogenes 24 h biofilm the more active compounds were myrtenyl acetate (50.89%) and α-pinene (43.73%). For P. carotovorum, all components inhibited its metabolism, except 1,8 cineole, which was completely ineffective.
The 24 h biofilm of S. aureus was very sensitive to the EO (65.30%) and to the presence of myrtenyl acetate (69.77%) more than to the presence of the other 3 components, whose inhibitory efficacy did not go beyond 44.92%.
The addition of the samples on ultra-mature 48 h biofilms showed, once again, a different behavior. In this case, S. aureus continued to be influenced by the presence of the whole EO that, although to a lesser extent, with inhibition of 44.96%. Once again, the EO proved effective against L. monocytogenes, inhibiting the cellular metabolism by 60.83% (with a loss of about 8% compared to the previous condition).
This means that, in the case of S. aureus, the action of the whole EO seemed to be mainly influenced by the status of the biofilm, and that the EO acted more effectively on mature (24 h) ultra-mature (48 h) biofilms. In the case of L. monocytogenes, the EO seemed to have no "preferences", acting in all three times in a similar way, albeit slightly decreasing on ultra-mature biofilms.
The EO resulted completely ineffective against P. carotovorum and its inhibitory efficacy against metabolism of P. aeruginosa progressively decreased from a mature biofilm to an ultra-mature biofilm.
Cytotoxic activity
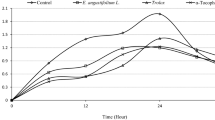
The cytotoxicity of M. communis EO, myrtenyl acetate, α-pinene, 1,8 cineole, and linalool was evaluated using an MTT assay performed on the human neuroblastoma cell line (SH-SY5Y). After 24 h of treatment, the essential oil and its main constituents revealed very different cytotoxic activities, as reported in Fig. 2.

Cell viability calculated as percentage after MTT assay. Cells were treated with different concentrations (500–25 μg/ml) of M. communis essential oil, myrtenyl-acetate, α-pinene, 1,8 cineole and linalool for 24 h and solvent (DMSO, 0.1%) alone. Data are the mean ± SD of three experiments *p < 0.05, ** p < 0.01, **** p < 0.0001 vs. DMSO
M. communis essential oil and α- pinene showed significant cytotoxicity at all concentrations tested even if their IC50 values are very different from each other; in fact, IC50 of essential oil was 209.1 μg/ml and for α- pinene was > 3 mg/ml. Linalool had significant activity against SH-SY5Y cells at concentrations ranging from 100 to 500 μg/ml, and showed an IC50= 356.3 μg/ml. Instead, myrtenyl-acetate had not cytotoxic activities with an IC50 > 2 g/ml. Treatment of SH-SY5Y neuroblastoma cells with the EO for 24 h resulted in a stronger cytotoxic activity respect to its principal constituents; these results suggested a synergic action between the EO components.
Anti-acetylcholinesterase activity
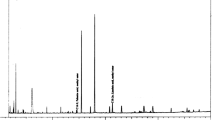
The acetylcholinesterase inhibitory activity of M. communis EO and its main constituents was evaluated by Ellman’s spectrophotometric method. Except for linalool, all tested substances showed AChE inhibitory activity and were able to inhibit in vitro the enzyme in a concentration-dependent manner (Fig. 3).

Dose-dependent inhibitory activity of M. communis EO, myrtenyl-acetate, α-pinene and 1,8 cineole against AChE. Data are given as mean ± SD (n = 3). *p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001 vs. Galantamine
The IC50 values are reported in Table 5. 1,8-Cineole and α-pinene exhibited the most promising activities with IC50 values of 13.5 and 15.0 µg/ml, respectively. Interesting activity against AChE was also observed for the whole EO (IC50 = 32.8 µg/ml), whereas a weak AChE inhibitory activity was found for myrtenyl acetate (IC50 = 111.4 µg/ml).
Anyway, the activities of myrtle EO and its main constituents are good if compared with galantamine, which showed an IC50 of 0.7 μg/ml.
Discussion
Phytochemical analyses
Bradesi and coworkers hypothesized two chemotypes of M. communis, according to the presence/absence of myrtenyl acetate [39]. Our sample seems to belong to the first chemotype because it was characterized by a high content of this compound (29.80%).
Our results agree with Boelens and Jimenez, who reported the chemical composition of a Spanish myrtle essential oil with a high percent of myrtenyl-acetate (>30.0%) and a lower content of α-pinene (< 8.50%) [39]. Moreover, in the EO from an Algerian wild myrtle, myrtenyl-acetate (38.7%), 1,8-cineole (12.7%), and α-pinene (13.7%) were the main components [40]
Chalchat and coworkers studied the EOs of M. communis from different regions of the Mediterranean area and showed that in the essential oils from Tunisia and Corsica, α-pinene (51.2-52.9%; 53.5-56.7%), 1,8-cineole (24.1-24.7%; 18.8-21.3%) and limonene (6.1-7.3%; 5.0-5.2%) were the main constituents while myrtenyl-acetate was present in little amounts (0.1-0.3%; 0.8%); instead 1,8-cineole (32.5-37.5%) and myrtenyl-acetate (14.8-21.1%) were the principal constituents in the Moroccan and coast of Montenegro EOs [3, 41].Another myrtle EO from Tunisia showed two predominant components 1,8 cineole-55.09% and α-pinene 33.14%, and myrtenyl acetate was absent [42,43,44].Badra and coworkers reported limonene (33.4%) as the principal constituents of the EO from a myrtle sample collected in northeastern Algeria; this component was also present in different amounts in all previously cited studies but not in our sample [45].
Italian samples of EO showed many differences between regions. Two myrtle EOs from two locations of Liguria (Italy) presented α-pinene as the principal constituent (41.6% and 28.9%, respectively); myrtenyl-acetate and myrtenol were absent [45]. Moreover, the EOs from of 52 genotypes of M. communis growing in the same collection field at Oristano (Sardinia, Italy) showed α-pinene, limonene, 1,8-cineole, α-terpineol, and linalool as main components with few differences among samples [46,47,48,49]; in our sample limonene was absent and α-terpineol present in low percentage (2.1%). These results suggested that the Sardinian myrtle EOs belong to the α-pinene, 1,8-cineole, limonene chemotype and are characterized by the lack of myrtenyl acetate. Instead, in nine samples of EOs from M. communis leaves myrtenyl acetate was present as the main or second main compound depending on chemotype [50].
The chemical composition of myrtle essential oil is highly variable due to several factors such as growing conditions (climate, altitude, humidity, temperature, etc.), geographical position, and season or vegetative period of the plant [51]. Moreover, a close link exists among light-shade conditions, essential oil yield, and morphological parameters [52].
Antibacterial activity
The results of different antibacterial activities highlighted that the chemical composition of the EO was ineffective or weakly effective against the tested Gram-negative bacteria. At the same time, it acted more strongly on the metabolism of the tested Gram-positive, albeit with different efficacy, according to the microorganism [18].
The data obtained suggested that myrtle EO seemed to be very effective ab origine on the biofilm formation if added at zero time, while its effectiveness has practically been canceled by adding it to mature biofilms (after 24 hours of growth). The addition of the EO to ultra-mature biofilms (after 48 hours of growth) seemed to increase again the effectiveness of EO myrtle, which was particularly effective against E. coli. When it was unable to act against the formation of the biofilm, the EO proved, however, effective by inhibiting bacterial metabolism in mature and ultra-mature biofilms, demonstrating that this EO was able to act both on young biofilms and mature and ultra-mature biofilms (through the action on bacterial cellular metabolism).
M. communis EO is known for its antimicrobial activity, generally ascribed to its chemical composition rich in monoterpene hydrocarbons and oxygenated monoterpenes such as linalool, carvacrol, α-pinene, and 1,8-cineole [53]. Like Berka-Zougali and coworkers [54], we observed a wide spectrum of action of myrtle EO. Moreover, other researchers reported a weak antibacterial activity of the EO against S. aureus and E. coli, due to the presence of its major components, such as 1,8-cineole and α-pinene [55]. Few studies are available in the literature on the ability of the myrtle EO to inhibit the formation of biofilms by pathogens [56,57,58]. Cannas and coworkers reported its efficacy in inhibiting biofilm formation by different Candida species, such as C. albicans, C. parapsilosis, and C. tropicalis [59]. Moreover, an isopropyl acetate extract obtained from myrtle leaves has proven effective in limiting the formation of biofilms by Propionibacterium acnes, also by acting on already mature biofilms.
The biofilm formation is responsible for several consequences, such as the production of exopolysaccharides, swimming, and swarming motility [60, 61]. Extracts and essential oils from several medicinal plants have been exploited as antibiofilm agents for pathogenic biofilm forming bacteria and fungi. They offer a virtually large and sustainable resource of very interesting classes of biologically active compounds. The prevention and/or control of biofilms by plant derivatives can occur through one or several mechanisms affecting the structure or the metabolism of the bacterial cells. There is a huge trend in relation to the identification of natural products that could possess anti-biofilm activity. myrtenol is a component of myrtle essential oil, demonstrated antibiofilm activity against S. aureus [62]. Myrtle EO was proved to block the metalloproteinase matrix activity [63], a mechanism involved in biofilm formation, as demonstrated by Xsia Tay and coworkers [64]. It has been used in combination with the EOs of Alchemilla vulgaris and Eucalyptus sp., to limit the formation of biofilms by Peptostreptococcus stomatis [65]. It was also shown inhibitory activity against S. mutans, S. sanguinis, and S. salivarius, therefore with a potential field of applicability in infections of teeth and oral cavity [66]. It is possible to hypothesize that myrtle EO and its main constituents analyzed in the present research could operate, depending on the strain, damaging the bacterial cell wall functions, or negatively affecting bacterial metabolism and enzymatic processes [60]. The myrtle EO may represent a product with a broad power against the pathogenic species E. coli, P. aeruginosa, L. monocytogenes, and S. aureus, resulting in particular scientific and practical interest due to the increased number of microbial species showing resistance to antibiotics [18, 53]. It is also important to highlight the activity of the EO against the Gram-negative P. carotovorum, known mainly as an agro-food pathogen infecting some of the most common crops such as potato, pineapple, and maize [58]. As is known, once the "niches" of biofilm, containing cells and other material (nucleic acids, proteins, polysaccharides) are formed, the cells present inside the biofilm tend to modify its metabolic pathways, not only to become more resistant than the homologous planktonic cells but also to express a greater "virulence". This means that starting from this step, it is more difficult to eradicate a possible infection using synthetic antibiotics, conventionally effective against the corresponding planktonic cells [60, 67]. In our study, the myrtle EO was able both to inhibit the viability of the cells present to the inner of biofilm, and/or to modulate the metabolic pathway that leads to greater bacterial virulence. The myrtle EO and its main components were studied in the antibacterial activity of these pathogens during the life of a biofilm, from immature to ultra-mature, associating the biofilm formation test (carried out with violet crystal) with an assessment of the biofilm metabolic activity (evaluated using the MTT reduction assay). This test has been used to evaluate the cytotoxicity of the myrtle EO in antimalarial assays [68] and to evaluate some biological properties, including the cytotoxicity of EOs from different areas of Algeria [69], Morocco [21], or Saudi Arabia [70]. The study performed by Alves and coworkers on thyme EOs demonstrated their ability to inhibit the biofilm formation and their influence on the viability of the cells entrapped within the biofilm, so to avoid or limit the subsequent cell changes occurring in [71]. The myrtle EO has been tested to inhibit the bacterial quorum-sensing mechanism [72]. The broad range of activity lead to incorporate it into chitosan edible films as an effective and safe strategy to deliver the oil to the foods [73]. In our experiments, the presence of myrtle EO, distinctly reduced the metabolic activity of cells in biofilms after 48 h of incubation. Our data also provided a preliminary indication that myrtle EO could affect the metabolic activity, as previously reported for the EO of Rosmarinus officinalis, rich in 1,8 cineole and α-pinene [74].
Cytotoxic activity
Few studies reported the potential cytotoxic activity of M. communis EO on cancer cell lines [20,21,22] and no one on SH-SY5Y cells. However, our sample was more active (IC50 = 209.1 μg/ml) than myrtle EOs reported in the literature. Scazzocchio et al. showed no toxicity of a commercial myrtle EO on HeLa cells after a 24 h treatment [20]; Harassi and coworkers reported moderate cytotoxicity of two Moroccan myrtle EOs against MCF7 and P815 cells, with IC50 ranging from 4.0 to 6.25 μg/ml for MCF7 and from 53.9 to 260 μg/ml for P815 cells after 48 h [21]. On HT29 cell line, M. communis EO from Yemen reached after 72 h an IC50 of 110 μg/ml [22]. Anyway, our IC50 value was > 20 μg/ml indicating that the essential oil was not cytotoxic as judged by the criterion set by the National Cancer Institute that stated that only natural substances with IC50 < 20 μg/ml were considered to be cytotoxic against the treated cells [75].
Anti-acetylcholinesterase activity
Only one study has been reported on the possible neuroprotective effects of M. communis EO. Sicilian EOs from the leaves of Myrtus communis L. stored in a collection orchard located at the experimental station ‘Orleans’ of the University of Palermo (Italy) showed lower acetylcholinesterase inhibitory activity than our sample, with IC50 values ranging from 96.0 to 520.2 μg/ml [27]. Another study regarding different myrtle leaf extracts displayed a moderate AChE inhibitory activity of M. communis [76].
No studies are available in the literature concerning the possible inhibitory AChE activity for myrtenyl acetate and linalool. Instead, our results confirm the good inhibitory activity for α pinene and 1,8 cineole. In fact, Dohi and coworkers reported IC50 values of 0.022 and 0.015 mg/ml, respectively [26].
Our findings revealed that the inhibitory activity of the whole EO results from a synergistic activity between the constituents. Further studies will be carried out to determine the possibility to use myrtle EO and/or its main constituents as coadjutant in the treatment of neurological disease.
Conclusions
This study provides a phytochemical profile for the EO from the leaves of M. communis from Cilento area, never investigated before. Moreover, the effects of this EO on the biofilm formation and biofilm cells metabolic activity, the cytotoxicity on SH-SY5Y cells and its possible activity as an anti-acetylcholinesterase inhibitor were evaluated. Results could open new perspectives for the application of M. communis EO as the potentialproduct against the resistant pathogenic species E. coli, P. aeruginosa, L. monocytogenes, and S. aureus. Moreover, the results obtained with cytotoxicity on SH-SY5Y cells used as a model of neuronal cells and the good activities as acetylcholinesterase inhibitors made myrtle EO and its main constituents candidates for further studies on their possible use as coadjutants in the treatment of neurological diseases.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- AChE:
-
Acetylcholinesterase
- AChI:
-
Acetylthiocholine iodide
- DMSO:
-
Dimethyl sulfoxide
- DNTB:
-
5,5-Dithio-bis-(2-nitrobenzoic acid)
- EO:
-
Essential oil
- FBS:
-
Foetal bovine serum
- FID:
-
Flame ionization detector
- GC:
-
Gas chromatography
- IC:
-
Inhibitory concentration
- MIC:
-
Minimum inhibitory concentrations
- MS:
-
Mass spectrometry
- MTT:
-
3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide
- OD:
-
Optical density
- PBS:
-
Phosphate buffered saline
- RPMI:
-
Roswell Park Memorial Institute
References
Cannas S, Molicotti P, Ruggeri M, Cubeddu M, Sanguinetti M, Marongiu B, Zanetti S. Antimycotic activity of Myrtus communis L. towards Candida spp. from clinical isolates. J Infect Dev Ctries. 2013;7:295–8.
Alipour G, Hosseinzadeh H, Dashti S. Review of pharmacological effects of Myrtus communis L. and its active constituents. Phytother Res. 2014;28:1125–36. https://doi.org/10.1002/ptr.5122.
Chalchat JC, Garry RF, Michet A. Essential oils of Myrtle (Myrtus communis L) of the Mediterranean litoral. J Essen Oil Res. 1998;10(613):617. https://doi.org/10.1080/10412905.1998.9700988.
Gortzi O, Lalas S, Chinou I, Tsaknis J. Reevaluation of bioactivity and antioxidant activity of Myrtus communis extract before and after encapsulation in liposomes. Euro Food Res Technol. 2008;226:583–90. https://doi.org/10.1007/s00217-007-0592-1.
Oyedemi SO, Okoh AI, Mabinya LV, Pirochenva G, Afolayan AJ. The proposed mechanism of bactericidal action of eugenol, α-terpineol and γ-terpinene against Listeria monocytogenes, Streptococcus pyogenes, Proteus vulgaris and Escherichia coli. Afr J Biotechnol. 2009;8:1280–90.
Cherrat L, Espina L, Bakkali M, García-Gonzalo D, Pagán R, Laglaoui A. Chemical composition and antioxidant properties of Laurus nobilis L. and Myrtus communis L. essential oils from Morocco and evaluation of their antimicrobial activity acting alone or in combined processes for food preservation. J Sci Food Agric. 2013;94:1197–204. https://doi.org/10.1002/jsfa.6397.
Ghnaya AB, Chograni H, Messoud C, Boussaid M. Comparative chemical composition and antibacterial activities of Myrtus communis L. essential oils isolated from tunisian and algerian population. J Plant Pathol Microbiol. 2013;4:186.
Høiby N, Bjarnsholt T, Givskov M, Molin S, Ciofu O. Antibiotic resistance of bacterial biofilms. Int J Antimicrob Agents. 2010;35:322–32. https://doi.org/10.1016/j.ijantimicag.2009.12.011.
Lewandowski Z, Caldwell DE, Korber DR, Lappin-Scott HM. Microbial biofilms. Annu Rev Microbiol. 1995;49:711–45.
Grumezescu AM. Essential oils and nanotechnology for combating microbial biofilms. Curr Org Chem. 2013;17:90–6.
Nazzaro F, Fratianni F, d’Acierno A, Coppola R, Ayala-Zavala FJ, da Cruz AG, De FeoV. Essential oils and microbial communication. In Essential Oils-Oils of Nature. Intech Open, London, UK, 2019
Camporese A. In vitro activity of Eucalyptus smithii and Juniperus communis essential oils against bacterial biofilms and efficacy perspectives of complementary inhalation therapy in chronic and recurrent upper respiratory tract infections. Infez Med. 2013;21:117–24.
Bernal-Mercado AT, Gutierrez Pacheco MM, Encinas Basurto D, MataHaro V, Lopez Zavala AA, Islas Osuna MA, Gonzalez-Aguilar GA, Ayala Zavala JF. Synergistic mode of action of catechin, vanillic and protocatechuic acids to inhibit the adhesion of uropathogenic Escherichia coli on silicone surfaces. J Appl Microbiol. 2019;128:387–400. https://doi.org/10.1111/jam.14472.
Djordjevic D, Wiedmann M, McLandsborough LA. Microtiter plate assay for assessment of Listeria monocytogenes biofilm formation. App Environ Microbiol. 2002;68:2950–598. https://doi.org/10.1128/AEM.68.6.2950-2958.2002.
Pär R. Davidsson, Tarja Kariola, Outi Niemi, E. T. Palva. Pathogenicity of and plant immunity to soft rot pectobacteria Front Plant Sci. 2013;4:121. https://doi.org/10.3389/fpls.2013.00191.
Gutierrez-Pacheco MM, Gonzalez-Aguilar GA, Martinez-Tellez MA, Lizardi-Mendoza J, Madera-Santana TJ, Bernal-Mercado AT, Vazquez-Armenta FJ, Ayala-Zavala JF. Carvacrol inhibits biofilm formation and production of extracellular polymeric substances of Pectobacterium carotovorum subsp. carotovorum. Food Control. 2018;89:210. https://doi.org/10.1016/j.foodcont.2018.02.007.
Hosseini Nezhad M, Alamshahi L, Panjehkeh N. Biocontrol efficiency of medicinal plants against Pectobacterium carotovorum, Ralstonia solanacearum and Escherichia coli. Open Conf Proc J. 2012;3:46–51. https://doi.org/10.2174/1876326X01203020046.
Nazzaro F, Fratianni F, Coppola R. Quorum sensing and phytochemicals. Int J Mol Sci. 2013;14:12607–19. https://doi.org/10.3390/ijms140612607.
Nazzaro F, Fratianni F, De Martino L, Coppola R, De Feo V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals. 2013;6:1451–74. https://doi.org/10.3390/ph6121451.
Baser KHC, Buchbauer G. Handbook of essential oils: science, technology, and applications. Florida: CRC Press; 2015.
Scazzocchio F, Garzoli S, Conti C, Leone C, Renaioli C, Pepi F, Angiolella L. Properties and limits of some essential oils: chemical characterisation, antimicrobial activity, interaction with antibiotics and cytotoxicity. Nat Prod Res. 2016;30:1909–18. https://doi.org/10.1080/14786419.2015.1086346.
Harassi Y, Tilaoui M, Idir A, Frédéric J, Baudino S, Ajouaoi S, Ait Mouse H, Zyad A. Phytochemical analysis, cytotoxic and antioxidant activities of Myrtus communis essential oil from Morocco. J Complement Integr Med. 2019;2019:16. https://doi.org/10.1515/jcim-2018-0100.
Anwar S, Crouch RA, Awadh Ali NA, Al-Fatimi MA, Setzer WN, Wessjohann L. Hierarchical cluster analysis and chemical characterisation of Myrtus communis L essential oil from Yemen region and its antimicrobial, antioxidant, and anti-colorectal adenocarcinoma properties. Nat Prod Res. 2017;31:158–2163. https://doi.org/10.1080/14786419.2016.1277346.
Heck JE, Ritz B, Hung RJ, Hashibe M, Boffetta P. The epidemiology of neuroblastoma: a review. Paediatr Perinat Epiidemiol. 2009;23:125–43. https://doi.org/10.1111/j.1365-3016.2008.00983.x.
Mukherjee PK, Kumar V, Mal M, Houghton PJ. Acetylcholinesterase inhibitors from plants. Phytomedicine. 2007;14:289–300. https://doi.org/10.1016/j.phymed.2007.02.002.
Ingkaninan K, Temkitthawon P, Chuenchom K, Yuyaem T, Thongnoi W. Screening for acetylcholinesterase inhibitory activity in plants used in Thai traditional rejuvenating and neurotonic remedies. J Ethnopharmacol. 2003;89:261–4. https://doi.org/10.1016/j.jep.2003.08.008.
Dohi S, Terasaki M, Makino M. Acetylcholinesterase inhibitory activity and chemical composition of commercial essential oils. J Agric Food Chem. 2009;57:4313–8. https://doi.org/10.1021/jf804013j.
Maggio A, Loizzo MR, Riccobono L, Bruno M, Tenuta MC, Leporini M, Tundis R. Comparative chemical composition and bioactivity of leaves essential oils from nine Sicilian accessions of Myrtus communis L. J Essent Oil Res. 2019;31:546–55. https://doi.org/10.1080/10412905.2019.1610089.
Council of Europe. European Pharmacopeia, 5th ed. Strasbourg Cedex: Council of Europe; 2004.
Della Pepa T, Elshafie HS, Capasso R, De Feo V, Camele I, Nazzaro F, Scognamiglio MR, Caputo L. Antimicrobial and phytotoxic activity of Origanum heracleoticum and O. majorana essential oils growing in Cilento (Southern Italy). Molecules. 2019;24(14):2576. https://doi.org/10.3390/molecules24142576.
Jenning W, Shibamoto T. Qualitative Analysis of Flavor and Fragrance Volatiles by Glass Capillary Gas Chromatography. San Francisco: Academic Press; 1980.
Davies N. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicon and Carbowax 20M phases. J Chromatogr. 1990;503:1–24.
Adams R. Identification of Essential Oil Components by Gas Chromatography/mass Spectroscopy. Carol Stream, IL: Allured Publishing; 2007.
Goodner KL. Practical retention index models of OV-101, DB-1, DB-5, and DB-Wax for flavor and fragrance compounds. LWT Food Sci Technol. 2008;41:951–8. https://doi.org/10.1016/j.lwt.2007.07.007.
Sarker SD, Nahar L, Kumarasamy Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods. 2007;42:321–4. https://doi.org/10.1016/j.ymeth.2007.01.006.
Fratianni F, Cozzolino A, De Feo V, Coppola R, Ombra MN, Nazzaro F. Polyphenols, antioxidant, antibacterial, and biofilm inhibitory activities of Peel and Pulp of Citrus medica L., Citrus bergamia, and Citrus medica cv. Salò Cultivated in Southern Italy Molecules. 2019;24:4577. https://doi.org/10.3390/molecules24244577.
Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods. 1983;65:55–63. https://doi.org/10.1016/0022-1759(83)90303-4.
Ellman GL, Courtney KD, Andres VJr, Featherstone RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol. 1961;7:88–95. https://doi.org/10.1016/0006-2952(61)90145-9.
Albano S, Lima AS, Miguel MG, Pedro LG, Barroso JG, Figueiredo AC. Antioxidant, anti-5-lipoxygenase and antiacetylcholinesterase activities of essential oils and decoction waters of some aromatic plants. Rec Nat Prod. 2012;6:35–48.
Boelens BH, Jimenez R. The chemical composition of Spanish Myrtle oils. Part II J Essent Oil Res. 1992;4:349–53. https://doi.org/10.1080/10412905.1997.10554245.
Touaibia M. Composition and anti-inflammatory effect of the common myrtle (Myrtus communis L.) essential oil growing wild in Algeria. Phytothérapie, 2017 https://doi.org/10.1007/s10298-017-1100-9
Mulas M, Melis RAM. Essential oil composition of myrtle (Myrtus communis) leaves. J Herbs Spices Med Plants. 2011;17(1):21–34. https://doi.org/10.1080/10496475.2011.556986.
Bekhechi C, Watheq Malti CE, Boussaïd M, Achouri I, Belilet K, Gibernau M, Casanova J, Tomi F. Composition and chemical variability of Myrtus communis leaf oil from northwestern Al-geria. Nat Prod Commun. 2019;14(5):1934578X19850030. https://doi.org/10.1177/1934578X19850030.
Mimica-Dukić N, Bugarin D, Grbo-vić S, Mitić-Ćulafić D, Vuković-Gačić B, Orčić D, Jovin E, Couladis M. Essential oil of Myrtus communis L. as a potential antioxidant and antimutagenic agents. Molecules. 2010;15(4):2759–70. https://doi.org/10.3390/molecules15042759.
Kaya DA, Ghica MV, Dănilă E, Öztürk Ş, Türkmen M, Albu Kaya MG, Dinu-Pîrvu CE. Selection of optimal operating conditions for extraction of Myrtus Communis L. essential oil by the steam distillation method. Molecules. 2020;25:2399. https://doi.org/10.3390/molecules25102399.
Flamini G, Cioni PL, Morelli I, Maccioni S, Baldini R. Phytochemical typologies in some populations of Myrtus communis L. on Caprione Promontory (East Liguria, Italy). Food Chem. 2004;85:599–604. https://doi.org/10.1016/j.foodchem.2003.08.005.
Usai M, Marchetti M, Culeddu N, Mulas M. Chemotaxonomic evaluation by volatolomics analysis of fifty-two genotypes of myrtus communis L. Plants. 2020;9:1288. https://doi.org/10.3390/plants9101288.
Tuberoso CI, Barra A, Angioni A, Sarritzu E, Pirisi FM. Chemical composition of volatiles in Sardinian myrtle (Myrtus communis L.) alcoholic extracts and essential oils. J Agric Food Chem. 2006;22:1420–6. https://doi.org/10.1021/jf052425g.
Tateo F, Picci V. Prime indagini sulla caratterizzazione GLCMS dell’olio essenziale di mirto di Sardegna (Mirtus communis L). RiV Soc Ital Sci Aliment. 1982;11:53–8.
Pirisino G, Mule A, Moretti MDL, Satta M. Yield and chemical composition of essential oil from self-sown Myrtus communis L from Cuglieri (Sardinia). RiV Itali EPPOS. 1996;7:159–69.
Siracusa L, Napoli E, Tuttolomondo T, Licata M, La Bella S, Gennaro MC, Ruberto GA. Two-year bio-agronomic and chemotaxonomic evaluation of wild sicilian myrtle (Myrtus communis L.) berries and leaves. Chem biodiversity. 2019;16:e1800575. https://doi.org/10.1002/cbdv.201800575.
Andrade EHA, Alves CN, Guimarães EF, Carreira LMM, Maia JGS. Variability in essential oil composition of Piper dilatatum L. C Rich Biochem Syst Ecol. 2011;39:669–75. https://doi.org/10.1016/j.bse.2011.05.021.
Fadil M, Farah A, Ihssane B. Chemometric investigation of light-shade effects on essential oil yield and morphology of Moroccan Myrtus communis L. Springerplus. 2016;5:1062. https://doi.org/10.1186/s40064-016-2749-5.
Sadiki M, Balouiri M, Barkai H, Maataoui H, Koraichi S, Elabed S. Synergistic antibacterial effect of Myrtus communis and Thymus vulgaris essential oils fractional inhibitory concentration. Int J Pharm Pharmac Sci. 2014;6:121–4.
Berka-Zougali B, Ferhat MA, Hassani A, Chemat F, Allaf KS. Comparative study of essential oils extracted from Algerian Myrtus communis L. leaves using microwaves and hydrodistillation. Int J Mol Sci. 2012;13:4673–95. https://doi.org/10.3390/ijms13044673.
Inouye S, Takizawa T, Yamaguchi H. Antibacterial activity of essential oils and their major constituents against respiratory tract pathogens by gaseous contact. J Antimicr Chemoth. 2001;47:565–73. https://doi.org/10.1093/jac/47.5.565.
Heras B, Scanlon MJ, Martin JL. Targeting virulence not viability in the search for future antibacterials. Br J Pharmacol. 2015;79:208. https://doi.org/10.1111/bcp.12356.
Hartmann R, Singh KP, Pearce P, Mok R, Boya Song B, Francisco Díaz-Pascual F, Dunkel J, Drescher K. Emergence of three-dimensional order and structure in growing biofilms. Nat Phys. 2019;15:251. https://doi.org/10.1038/s41567-018-0356-9.
Mehrsorosh H, Gavanji S, Larki B, Mohammadi MD, Karbasiun A, Bakhtari A, Hashemzadeh F, Mojiri A. Essential oil composition and antimicrobial screening of some Iranian herbal plants on Pectobacterium carotovorum. Global NEST J. 2014;16:240–51.
Cannas S, Molicotti P, Usai D, Maxia A, Zanetti S. Antifungal, anti-biofilm and adhesion activity of the essential oil of Myrtus communis L against Candida species. Nat Prod Res. 2014;28:2173–7. https://doi.org/10.1080/14786419.2014.925892.
Nazzaro F, Fratianni F, d’Acierno A, De Feo V, Ayala Zavala FJ, Gomez- Cruz A, Granato G, Coppola R. Effect of polyphenols on microbial cell-cell communications. In Quorum Sensing. Molecular Mechanism and Biotechnological Application; ed Tommonaro, G.; Amsterdam:Elsevier; 2019 https://doi.org/10.1016/B978-0-12-814905-8.00008-3
Schonewille E, Nesse LL, Hauck R, Windhorst D, Hafez HM, Vestby LK. Biofilm building capacity of Salmonella enterica strains from the poultry farm environment. FEMS Immunol Med Microbiol. 2012;65:360–5. https://doi.org/10.1111/j.1574-695X.2012.00966.x.
Cordeiro L, Figueiredo P, Souza H, Sousa A, Andrade FJr, Barbosa-Filho P, Lima E. Antibacterial and Antibiofilm Activity of Myrtenol against Staphylococcus aureus. Pharmaceuticals. 2020;13:133. https://doi.org/10.3390/ph13060133.
Zeidán-Chuliá F, Rybarczyk-Filho JL, Gursoy M, Könönen E, Uitto VJ, Gursoy OV, Cakmakci L, Moreira JCF, Ulvi K. Bioinformatical and in vitro approaches to essential oil-induced matrix metalloproteinase inhibition. Pharm Biol. 2012;50:675–86. https://doi.org/10.3109/13880209.2012.677847.
Xsia Tay C, Yiling Quah S, Nee Lui J, Soo Hoon Yu V, SooTan K. Matrix Metalloproteinase inhibitor as an antimicrobial agent to eradicate Enterococcus faecalis Biofilm. J Endod. 2015;41:858–63. https://doi.org/10.1016/j.joen.2015.01.032.
Marshall-Jones, Z.; Baillon, M.-L. ; Buckley, C. Orally hygiene composition comprising myrtle. 2010 US Patent 20100061944A1.
Rasaie N, Esfandiari E, Rasouli S, Abdolahian F. Antimicrobial effect of Myrtus communis. L. essential oils against oral microorganism. Jentashapir J Health Res. 2017;9:e12032.
Kerekes EB, Deák E, Takó M, Tserennadmid R, Petkovits T, Vágvölgyi C, Krisch J. Anti‐biofilm forming and anti‐quorum sensing activity of selected essential oils and their main components on food‐related micro‐organisms. J Appl Microbiol. 2013;115:933–42. https://doi.org/10.1111/jam.12289.
Naghibi F, Esmaeili S, Rain Abdullah N, Nateghpour M, Taghvai M, Kamkar S, Mosaddegh M. In Vitro and In Vivo Antimalarial evaluations of myrtle extract, a plant traditionally used for treatment of parasitic disorders. BioMed Res Int. 2013;2013: 316185. https://doi.org/10.1155/2013/316185.
Bouzabata A, Cabral C, Gonçalves MJ, Cruz MT, Bighelli A, Cavaleiro C, Casanova J, Tomi F, Salgueiro L. Myrtus communis L. as source of a bioactive and safe essential oil. Food Chem Toxicol. 2015;75:166–72. https://doi.org/10.1016/j.fct.2014.11.009.
Mohamed ME, Mohafez OM, Khalil HE, Alhaider IA. Essential oil from myrtle leaves growing in the Eastern part of Saudi Arabia: components, anti-inflammatory and cytotoxic activities. J Essent Oil Bear Plants. 2019;22:4877–92. https://doi.org/10.1080/0972060X.2019.1645046.
Alves M, Gonçalves MJ, Zuzarte M, Alves-Silva JM, Cavaleiro C, Cruz MT, Salgueiro L. Unveiling the antifungal potential of two Iberian thyme essential oils: effect on C albicans germ tube and preformed biofilms. Front Pharmacol. 2019;10:446. https://doi.org/10.3389/fphar.2019.00446.
Aleksic V, Knezevic P. Antimicrobial and antioxidative activity of extracts and essential oils of Myrtus communis L. Microbiol Res. 2014;169:240–54. https://doi.org/10.1016/j.micres.2013.10.003.
Myszka K, Sobieszczańska N, Olejnik A, Majcher M, Szwengiel A, Wolko Ł, Juzwa W. Studies on the anti-proliferative and anti-quorum sensing potentials of Myrtus communis L. essential oil for the improved microbial stability of salmon-based products. LWT. 2020;127:109380. https://doi.org/10.1016/j.lwt.2020.109380.
Miladi H, Mili D, Slama RB, Zouari S, Ammar E, Bakhrouf A. Antibiofilm formation and anti-adhesive property of three mediterranean essential oils against a foodborne pathogen Salmonella strain. Microb pathog. 2016;93:22–31. https://doi.org/10.1016/j.micpath.2016.01.017.
Geran RI, Greenberg NH, Macdonald MM, Schumacher AM, Abbott BJ. Protocols for screening chemical agents and natural products against animal tumours and other biological systems. Cancer Chemother Rep. 1972;3:59–61.
Acknowledgements
Not applicable
Funding
This research received no external funding.
Author information
Authors and Affiliations
Contributions
Conceptualization, V.D.F and F.N; chemical characterization F.C., L.C. and G.A; cytotoxic and anti-acetylcholinesterase activities L.C, F.C, G.A and G.V; antimicrobial activity F.N and F.F; validation, L.C, F.N. and F.F, writing—original draft preparation, L.C, F.N. and V.D.F; writing—review and editing, V.D.F. All authors have read and approved final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Caputo, L., Capozzolo, F., Amato, G. et al. Chemical composition, antibiofilm, cytotoxic, and anti-acetylcholinesterase activities of Myrtus communis L. leaves essential oil. BMC Complement Med Ther 22, 142 (2022). https://doi.org/10.1186/s12906-022-03583-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-022-03583-4