
Abstract
Background
Little is known about DNMT3A expression and its prognostic significance in childhood B cell acute lymphoblastic leukemia (B-ALL).
Methods
We determined DNMT3A mRNA expression in 102 children with B-ALL. Correlations with relapse-free survival (RFS) and common clinical characteristics were analyzed. DNMT3A was stably knocked out by CRISPR/Cas9 gene editing technology in Reh and 697 B-ALL cell lines. Cell proliferation activity after treated with daunorubicin (DNR) was determined by CCK8 assay in DNMT3A KO Reh and 697 cell lines.
Results
DNMT3A expression in B-ALL patients who were in continuous complete remission (CCR) was higher than in those who got relapse (P = 0.0111). Receiver operating characteristic curve showed prognostic significance of DNMT3A expression (P = 0.003). Low expression of DNMT3A (≤ 0.197) was significantly correlated with poor RFS (P < 0.001) in children with B-ALL. Knock-out of DNMT3A in Reh and 697 cell lines significantly increased IC50 of DNR (P = 0.0201 and 0.0022 respectively), indicating elevated resistance to DNR.
Conclusion
Low expression of DNMT3A associates with poor prognosis in children with B-ALL. Knock-out of DNMT3A confers resistance to DNR on leukemic cells.
Similar content being viewed by others

Background
B cell acute lymphoblastic leukemia (B-ALL) is the most common childhood malignancy. Although cure rate of childhood B-ALL has been greatly improved with risk-adjusted therapy [1, 2], relapsed leukemia is still a leading cause of death for children mainly due to therapy resistance [2,3,4]. Thus, it is of great significance to clarify the mechanisms of therapy resistance and relapse of B-ALL.
DNA methyltransferase 3A (DNMT3A) catalyzes de novo DNA methylation and plays important roles in the pathogenesis of malignancies including leukemia. Furthermore, DNMT3A mutations in acute myeloid leukemia (AML) and T cell ALL are associated with poor prognosis of the patients [5,6,7,8,9,10,11]. Our previous studies have shown that DNMT3A mutations can be found in a few of children with B-ALL, and are correlated with poor prognosis [12]. However, the expression level of DNMT3A and its prognostic significance in B-ALL remains unclear.
In this study, we assessed the relationship between expression level of DNMT3A and prognosis in Chinese childhood B-ALL. Moreover, CRISPR/Cas9 has been used to knock out DNMT3A gene in leukemic Reh and 697 cell lines in order to explore the role of DNMT3A expression playing in resistance to chemotherapeutic drugs. We showed that low expression of DNMT3A was correlated with poor treatment outcome, knock-out of this gene resulted in obvious resistance to DNR, a common chemotherapeutic drug in treatment of ALL.
Methods
Patients
From July 2010 to May 2014, a total of 226 consecutive childhood patients with newly diagnosed B-ALL were admitted to Beijing Children’s Hospital. The criterion for the patient’s inclusion was ≥70% leukemic cells in BM samples [13, 14].
One hundred two B-ALL patients with available diagnostic bone marrow (BM) samples were enrolled in this study. One hundred twenty-four patients not fulfilling the inclusion criterion were excluded from this study. No difference was found between patients included and excluded in terms of patients’ characteristics and survival to suggest selection bias (Supplemental Table S1). BM samples from 11 patients in continuous CR were collected and used as control. ALL patients were diagnosed and treated in accordance with the Chinese Children’s Leukemia Group ALL 2008 Protocol (CCLG-ALL 2008) at Beijing Children’s Hospital [4].
Among the 102 patients, there were 64 boys and 38 girls, aged from 1 to 13 years with a median age of 4. Chromosome karyotype analysis was performed in 66 patients and the karyotype results were interpreted according to the International System for Human Gytogenomic Nomenclature guide-lines [15]. Of the 66 patients, normal and abnormal karyotype was seen in 22 and 32 patients respectively and no metaphase schizophrenia was found in the 12 patients. Fusion gene was detected in all 102 patients by a nested multiplex reverse transcription polymerase chain reaction (RT-PCR) system, as described by Gao C. et al. [16]. Thirty-four patients carried 4 types of fusion genes including ETV6-RUNX1, TCF3-PBX1, BCR-ABL1, and FUS-ERG. The details of stratification and treatment according to CCLG-ALL 2008 were described previously [4, 17]. Ninety-four patients were in continuous complete remission (CR), 8 patients relapsed 2 to 62 months after diagnosis. The follow-up time ranged from 1.67 to 92 months (median, 59 months). MRD at d33 (the end of induction of remission) and d78 (before consolidation therapy) were detected using RQ-PCR targeted at Ig/TCR (immunoglobulin and T cell receptor gene rearrangements) according to European MRD (Minimal residual disease) laboratory guidelines [18,19,20,21]. Informed consents were obtained from all the children’s parents or legal guardians.
Cell lines
Human B-ALL cell lines Reh and HEK293T cell were purchased from National Infrastructure of Cell Line Resource (No. 3101HUMTCHu131 and 1101HUM-PUMC000010, respectively); 697 cell line was a kind gift from Dr. Suning Chen at the first affiliated Hospital of Soochow University (Suzhou, China). Reh and 697 were cultured in RPMI 1640 (GIBCO, USA) supplemented with 10% fetal bovine serum (FBS, AusGeneX, Brisbane) and 1% penicillin/streptomycin. HEK293T cell was cultured in DMEM supplemented with 10% FBS and 1% penicillin/streptomycin. All cells were maintained at 37 °C in a humidified atmosphere containing 5% CO2.
Nucleic acid extraction
Mononucleated cells were separated from 1 ml of patients’ BM aspirate by centrifugation with Ficoll 400 (MD Pacific Technology CO., Ltd.) and stored at -70˚C until use. Total RNA of samples was extracted and reverse transcribed using Trizol Reagent (Invitrogen, USA) and MMLV reverse transcriptase (Promega, USA) according to the manufacturers’ instructions respectively. Genomic DNA of Reh and 697 cell lines were extracted using a Blood & Cell Culture DNA Midi Kit (TIANGEN, China) according to the manufacturer’s protocol.
Quantitative analysis of DNMT3A expression
Real-time quantitative polymerase chain reaction (RQ-PCR) was performed using Power SYBR™ Green PCR Master Mix (Applied Biosystems 4,367,659) by an ABI Prism 7500 Sequence Detection System (Applied Biosystems, Foster City, CA, USA). GUS (β-Glucuronidase) expression was used as an internal control. The cycling condition included pre-denaturation at 95 °C for 30 s, followed by 40 cycles of 5 s at 95 °C, 30 s at 55 °C and 30 s at 72 °C. Primers were shown in Table 1. We used the cDNA samples obtained from 697 cell line as a calibrator. The relative expression of DNMT3A was calculated by the method of 2−ΔΔCt. The levels of DNMT3A and GUS were tested in triplicates.
Lenti DNMT3A-sgRNA-Cas9 constructs
The cDNA sequence encoding sgRNA which targets a conserved sequence in exon 7 of human DNMT3A gene was synthesized and subcloned into LentiCRISPR-v2 plasmid (Addgene 52961, kindly provided by Dr. Jian Huang at Temple University, Philadelphia, PA) to make the lentiDNMT3A-sgRNA-Cas9 construct. Briefly, the forward and reverse primers including 20 bp target DNMT3A sequence and BsmbI sticky ends were annealed and inserted into the lentiCRISPR-v2 plasmid digested with FastDigest Esp3I (Thermo Fisher Scientific, #FD0454) (Fig. 1b). sgRNA primer sequences have been reported by Gundry MC et al. previously and were shown in Table 1 [22].

Schematic diagram of sgRNA targeting DNMT3A. a The structure of DNMT3A gene and the three common transcripts. Black vertical lines: exons. Horizontal lines: introns. Arrow: the location of sgRNA targeting exon 7. b The structure of lentiCRISPR v2 plasmid. The arrows indicate the sgRNA sequence
Lentivirus production and infection
To produce lentivirus, 6 μg of transfer plasmid lentiDNMT3A-sgRNA-Cas9 or control plasmid lenti-CRISPR-v2 were co-transfected into HEK 293 T cells with 4.5 μg of packaging plasmids psPAX2 (AddGene 12260) and 3 μg of VSV-G (AddGene 8454) using FuGENE® 6 Transfection Reagent (Promega E2692) according to the manufacturer’s instructions. After incubation for 48 h, the culture supernatants containing lentivirus were harvested and filtered with 0.45 μm filter and stored at − 80 °C. The Reh and 697 cell lines (5 × 105) was infected with the lentivirus at an M.O.I. of 40 separately, using spin-transduction (centrifuging the plate coated with 8 μg/ml polybrene (SANTA CRUZ) at 1200 g for 2 h at 25 °C), then were cultured for 24 h in the incubator. On the next day, the medium was changed with fresh RPMI 1640 complete medium and the cells were cultured for another 24 h.
T7EN1 assays for quantifying frequencies of indel mutations
Lentivirus-infected cells were selected by 1 μg/ml puromycin for 2 days. Genomic DNA was extracted and used to amplify the genomic region flanking the DNMT3A sgRNA target site with KAPA2G Robust HotStart ReadyMix (KAPA BIOSYSTEMS KK5702) and PCR primers listed in Table 1. Then T7EN1 assay was performed using T7 Endonuclease I (NEB #M0302L) according to the Instruction Manual. The digested DNA was analyzed on electrophoresis system using a 2% agarose gel.
Western blotting
A fraction of lentivirus-infected cells was lysed in NE-PER® Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific, USA). The lysates were denatured in 5 × SDS loading buffer by boiling at 95 °C for 10 min and were subjected on a NuPAGE™ 4–12% Bis–Tris Protein Gels (Invitrogen). After transferred to Biotrace NT nitrocellulose Transfer Membrane (PALL, 66,485), the expression of proteins was detected using following antibodies: 1:300 DNMT3A (D23G1) Rabbit mAb (No. 3598; CST) alone was incubated firstly, then 1:300 DNMT3A (D2H4B) Rabbit mAb (No. 32578; CST) and 1:2000 Lamin B1 Mouse mAb (No. 66095–1-Ig; Proteintech) were mixed and incubated together on the next day in the same blot after finishing the secondary antibody incubation and band scanning for DNMT3A(D23G1) mAb. Secondary antibody included Goat anti-Mouse IgG (H + L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor 680 (No. A21058; Invitrogen) and Goat anti-Rabbit IgG (H + L) Highly Cross-Adsorbed Secondary Antibody, Alexa Fluor Plus 800 (No. A32735; Invitrogen). The bands were scanned by LICOR Odyssey CLX.
Cell viability
The lentivirus-infected Reh and 697 cell lines were plated into 96-well plate separately, 104/well. After treatment with 100 μl DNR, the cells were cultured for 24 h at 37 °C in a humidified atmosphere containing 5% CO2. The concentrations of DNR for Reh were 0.0009, 0.0055, 0.0111, 0.0222, 0.0443, 0.1773 μM; and for 697 were 0.0089, 0.0177, 0.0887, 0.1773, 0.8866, 1.7731 μM. Twenty microliters of Cell Counting Kit-8 (CCK-8, Yeasen 40203ES60*, Shanghai, China) solution were added to each well and mixed gently. After incubation for 1 h, optical density (OD) at 450 nm was determined using a Spectra MAX 190 microplate reader. After calibrated with non-cellular background, cell viability was calculated using a non-treatment control regarded as 100% of cell viability.
Statistical analysis
Receiver Operating Characteristic (ROC) curve was used to decide the cut-off value of low- and high-expression of DNMT3A (DNMT3Alow and DNMT3Ahigh) in leukemic cells of children with B-ALL. Fisher’s exact test was used to test the differences in clinical characteristics and relapse rates between DNMT3Alow and DNMT3Ahigh patients. Relapse free survival (RFS) was defined as the date of leukemia diagnosis to the date of recurrence. Survival estimates were calculated using the Kaplan–Meier method, and the groups were compared using the log-rank test. The independent prognostic significance of DNMT3A expression and the common clinical features was analyzed by Cox proportional hazards model (Method: Enter). All data were analyzed with the SPSS 16.0 software package and a P value < 0.05 was considered statistically significant. The fitting curves of inhibitory effects of DNR on cell proliferation were plotted by GraphPad Prism 8, and half maximal inhibitory concentration (IC50) was also calculated by the software.
Results
DNMT3A expression in childhood B-ALL
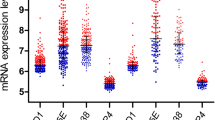
Firstly, we determined DNMT3A expression in 102 newly diagnosed (ND) B-ALL patients and 11 patients in continuous CR (control) by relative quantitative PCR. As a result, DNMT3A expression in ND patients with B-ALL, ranged from 0.0006594 to 1.713 with a median of 0.4363, was significantly higher than that in control patients (range: 0.08055 to 0.1865, median: 0.1147; P = 0.0004, Fig. 2a). Interestingly, DNMT3A expression in ND B-ALL patients who got relapse was significantly decreased compared with that in patients who were in CCR at the last follow-up (P = 0.0111, Fig. 2b).

DNMT3A expression in ND B-ALL patients and controls. a DNMT3A expression was significantly increased in ND B-ALL patients compared with that of controls. b DNMT3A expression was significantly decreased in ND B-ALL patients who relapsed compared with that in ND patients in CCR
Low expression of DNMT3A indicated poor prognosis in ND B-ALL patients
ROC curve analysis was performed to evaluate the prognostic value of DNMT3A expression. When regarded as a continuous value, DNMT3A expression had a good predictive significance for relapse of B-ALL in children, with an area under curve (AUC) of 0.819 (95% CI: 0.686–0.952; P = 0.003), Fig. 3A), which indicated that DNMT3A expression could be a potential prognostic biomarker for ND B-ALL patients.

Prognostic significance of DNMT3A expression in 102 children with B-ALL. a ROC curve analysis of DNMT3A expression with relapse as an event. b The patients in DNMT3Alow group had poorer RFS than those in DNMT3A.high group (P < 0.001)
According to the ROC curve, 0.197 was determined as the optimal cutoff value for DNMT3A expression level, with a sensitivity and specificity of 75% and 87.2% respectively. Using this cut-off, we divided 102 patients into two groups, 22 cases with low DNMT3A expression (≤ 0.197, DNMT3Alow) and 80 cases with high DNMT3A expression (> 0.197, DNMT3Ahigh). There was a significantly higher relapse rate in DNMT3Alow group (6 out of 22 vs 2 out of 88, Fisher’s exact test, P = 0.001). Moreover, poor RFS was also observed in the DNMT3Alow group (57.3% ± 17.9% vs. 97.2% ± 1.9%, P < 0.001) (Fig. 3b). Among other common clinical features, only clinical risk classification had a trend of correlation with RFS (P = 0.053, Table 2), indicating its important prognostic value.
In multivariate analysis for prognosis (Table 2), DNMT3A expression, Age, WBC at diagnosis, MRD at day 33 and clinical risk classification were used as covariates in a cohort of 96 cases due to no MRD result about 6 patients. Adjusted for all these factors, DNMT3A expression remained an independent prognostic factor for RFS of patients with B-ALL (HR = 36.824, 95% CI: 3.746 ~ 362.033, P = 0.002, Table 2). In addition, high risk was also an independent prognostic factor for RFS (P = 0.022, Table 2). These findings indicated that low expression of DNMT3A in leukemic cells at diagnosis could be a useful indicator for disease relapse in childhood B-ALL.
Comparison of clinical features between DNMT3A low and DNMT3A high patients
In the next step, we analyzed the correlation of DNMT3A expression with common clinical characteristics such as age, gender, white blood cell (WBC) count at diagnosis and fusion genes. However, no correlation was found between DNMT3A expression and above clinical characteristics (Table 3).
We further analyzed the association of DNMT3A expression with MRD at d33, MRD at d78 and risk classification respectively, but no significant correlation between them was found (Fisher’s exact test, P > 0.05, Table 3).
Knock-out of DNMT3A enhanced resistance of Reh and 697 cell lines to DNR
To confirm the correlation of low expression of DNMT3A with poor prognosis of children with B-ALL, firstly, we disrupted DNMT3A in Reh and 697 cell lines separately. T7 endonuclease I (T7EN1) assay showed high efficiency of the sgRNA to direct Cas9-mediated ablation of DNMT3A (Fig. 4a and b). Furthermore, as expected, Western blotting indicated that DNMT3A expression was remarkably reduced after infection with DNMT3A-sgRNA lentivirus (Fig. 4c and d).

LentiCRISPR/Cas9 mediated editing of DNMT3A gene in Reh and 697 cell lines. a and b T7e1 assay analysis of specific sgRNA-mediated indels at DNMT3A locus in Reh and 697 cell lines separately. The lower migrating bands marking by a white arrow represent the disrupted gene alleles. c and d Expression of three DNMT3A protein variants was significantly reduced in Reh and 697 cell lines infected by DNMT3A-sgRNA lentivirus separately
DNR is one of the main chemotherapeutic drugs in induction therapy of B-ALL. We next tested whether knock-out of DNMT3A gene could cause Reh and 697 cell lines to be tolerant to DNR by CCK8 assay. These cells were treated by different concentrations of DNR for 24 h. It was shown that IC50 was significantly increased in the DNMT3A-knockout cells, indicating decreased cell viability (Fig. 5a, Control vs. DNMT3A KO, 0.02159 vs. 0.02892 μM, P = 0.0201; Fig. 5b, Control vs. DNMT3A KO, 0.1190 vs. 0.1865 μM, P = 0.0022). These results demonstrated that sgRNA mediated Cas9 knock-out of DNMT3A can causes Reh and 697 cell lines to be resistant to DNR, implying that DNMT3A expression plays an important role in the sensitivity of B-ALL leukemic cells to chemotherapeutic drugs such as DNR.

Knock-out of DNMT3A gene increased resistance of Reh and 697 cell lines to DNR. IC50 of DNR significantly increased in DNMT3A-knockout cells (Independent-samples T test, P = 0.0201 in Reh cell line (a) and P = 0.0022 in 697 cell line (b)). The standard errors of the means are shown (n = 3 experiments for each drug concentration)
Discussion
In recent two decades, DNMT3A mutations have been found in approximately 20% of adult AML patients, 9% of adult T-ALL and 0 ~ 1.4% of childhood AML, and the hotspots of mutations are mainly located in exon 23 which encodes the catalytic methyltransferase domain [5,6,7,8,9,10,11]. DNMT3A mutations are associated with poor prognosis and used for risk stratification in AML [5,6,7,8,9,10], and is associated with increased age and adverse outcome in adult T-ALL [11]. However, few studies focused on the role of DNMT3A in B-ALL. Our previous study has shown that DNMT3A mutations can be found in exon 23 and its adjacent intron regions in a few of children with B-ALL (5/182, 2.7%), and may have adverse impact on prognosis [12].
As there are only a few B-ALL patients with DNMT3A mutations, we sought to determine the prognostic significance of DNMT3A expression in B-ALL. As expected, low expression was associated with relapse in 102 patients with B-ALL. Furthermore, knock-out of DNMT3A increased IC50 of DNR in Reh and 697 cell lines, indicating the relationship of low expression of DNMT3A and chemoresistance.
It was reported that Dnmt3a loss in HSCs leads to hypomethylation of genes with a causal role in cancer, such as Runx1 and Gata3. Runx1 promotes murine erythroid progenitor proliferation and inhibits differentiation by preventing Pu.1 downregulation [23]. Gata3 targets Runx1 in the embryonic hematopoietic stem cell niche [24]. Thus, previous studies and ours’ suggest that deletion or low-expression of DNMT3A result in differentiation inhibition of HSCs and allow HSCs to be propagated indefinitely in vivo [25,26,27,28], which may play an important role in leukemogenesis and resistance to chemotherapy. This may provide us with an alternative target of therapy for childhood B-ALL.
It has been reported that DNMT3A expression is directly transactivated by transcription factor WT1 in Wilms’ tumor cells [29] and is negatively regulated by p53 at the transcriptional level in lung cancer [30]. In fact, overexpression of the WT1 transcript was demonstrated in children with B-ALL at diagnosis [31], which may contribute to increased expression of DNMT3A in leukemic cells. However, increased expression of TP53 by 2 to 20-fold higher in pediatric primary B-ALL than in healthy controls [32] would inhibit DNMT3A expression. Thus, the regulation mechanism of DNMT3A expression is quite complicated in different types of cancer cells and needs to be clarified especially in childhood B-ALL.
There are some limitations or weakness in this study. Firstly, as this is a retrospective study and only patients with > 70% of bone marrow infiltration were selected, the findings were based on a small number of cases. A prospective study with large sample size and long-term follow-up are necessary to confirm the prognostic significance of DNMT3A. Secondly, the influence of treatment compliance could not be assessed. More attention should be paid to this point in the prospective study. Thirdly, we only focused on DNMT3A expression, ignoring its DNA methyltransferase activity, the role of which in chemoresistance of B-ALL cells should be clarified in future study.
Conclusions
In summary, we associated low expression of DNMT3A with poor prognosis in Chinese pediatric patients with B-ALL. Furthermore, the knockout of DNMT3A conferred less sensitivity to daunorubicin in leukemic cell lines. Successful disruption of DNMT3A in Reh and 697 cell lines may facilitate the studies on mechanism of relapse and chemotherapeutic resistance for childhood B-ALL. Future prospective studies with large sample size, long-term follow-up, more leukemia cell lines and more mechanism research were recommended to confirm the DNMT3A role in childhood with B-ALL.
Availability of data and materials
All data generated and analyzed during this study are included in this manuscript and original data as supplemental part.
Change history
21 April 2023
A Correction to this paper has been published: https://doi.org/10.1186/s12885-023-10862-x
Abbreviations
- B-ALL:
-
B cell acute lymphoblastic leukemia
- RFS:
-
Relapse-free survival
- DNR:
-
Daunorubicin
- CCR:
-
Continuous complete remission
- DNMT3A:
-
DNA methyltransferase 3A
- AML:
-
Acute myeloid leukemia
- BM:
-
Bone marrow
- CCLG-ALL 2008:
-
Chinese Children’s Leukemia Group ALL 2008 Protocol
- CR:
-
Continuous complete remission
- Ig/TCR:
-
Immunoglobulin And T cell receptor gene rearrangements
- MRD:
-
Minimal residual disease
- RQ-PCR:
-
Real-time quantitative polymerase chain reaction
- ROC:
-
Receiver Operating Characteristic
- RFS:
-
Relapse free survival
- WBC:
-
White blood cell
- T7EN1:
-
T7 endonuclease I
References
Maloney KW, Devidas M, Wang C, Mattano LA, Friedmann AM, Buckley P, et al. Outcome in Children With Standard-Risk B-Cell Acute Lymphoblastic Leukemia: Results of Children’s Oncology Group Trial AALL0331. J Clin Oncol. 2020;38:602–12.
Hunger SP, Mullighan CG. Acute Lymphoblastic Leukemia in Children. N Engl J Med. 2015;373:1541–52.
Kimura S, Mullighan CG. Molecular markers in ALL: Clinical implications. Best Pract Res Clin Haematol. 2020;33:101193.
Cui L, Li ZG, Chai YH, Yu J, Gao J, Zhu XF, et al. Outcome of children with newly diagnosed acute lymphoblastic leukemia treated with CCLG-ALL 2008: The first nation-wide prospective multicenter study in China. Am J Hematol. 2018;93:913–20.
Park DJ, Kwon A, Cho BS, Kim HJ, Hwang KA, Kim M, et al. Characteristics of DNMT3A mutations in acute myeloid leukemia. Blood Res. 2020;55:17–26.
Li W, Cui L, Gao C, Liu S, Zhao X, Zhang R, et al. DNMT3A mutations in Chinese childhood acute myeloid leukemia. Medicine (Baltimore). 2017;96:e7620.
Brunetti L, Gundry MC, Goodell MA. DNMT3A in Leukemia. Cold Spring Harb Perspect Med. 2017;7:a030320.
Ho PA, Kutny MA, Alonzo TA, Gerbing RB, Joaquin J, Raimondi SC, et al. Leukemic mutations in the methylation-associated genes DNMT3A and IDH2 are rare events in pediatric AML: a report from the Children’s Oncology Group. Pediatr Blood Cancer. 2011;57:204–9.
Yang L, Rau R, Goodell MA. DNMT3A in haematological malignancies. Nat Rev Cancer. 2015;15:152–65.
Shah MY, Licht JD. DNMT3A mutations in acute myeloid leukemia. Nat Genet. 2011;43:289–90.
Bond J, Touzart A, Lepretre S, Graux C, Bargetzi M, Lhermitte L, et al. DNMT3A mutation is associated with increased age and adverse outcome in adult T-cell acute lymphoblastic leukemia. Haematologica. 2019;104:1617–25.
Li W, Gao C, Cui L, Liu S, Zhao X, Zhang R, et al. DNMT3A mutations and prognostic significance in childhood acute lymphoblastic leukemia. Leuk Lymphoma. 2015;56:1066–71.
Carroll WL, Bhojwani D, Min DJ, Raetz E, Relling M, Davies S et al. Pediatric acute lymphoblastic leukemia. Hematol Am Soc Hematol Educ Program. 2003;2003(1):102–31.
Jiao Y, Cui L, Gao C, Li W, Zhao X, Liu S, et al. CASP8AP2 is a promising prognostic indicator in pediatric acute lymphoblastic leukemia. Leuk Res. 2012;36:67–71.
Stevens-Kroef M, Simons A, Rack K, Hastings RJ. Cytogenetic Nomenclature and Reporting. Methods Mol Biol. 2017;1541:303–9.
Gao C, Zhao XX, Li WJ, Cui L, Zhao W, Liu SG, et al. Clinical features, early treatment responses, and outcomes of pediatric acute lymphoblastic leukemia in China with or without specific fusion transcripts: a single institutional study of 1,004 patients. Am J Hematol. 2012;87:1022–7.
Zhao X, Gao C, Cui L, Li W, Liu S, Zhang R, et al. Quantitative monitoring of minimal residual disease in childhood acute lymphoblastic leukemia using TEL-AML1 fusion transcript as a marker. Pediatr Investig. 2018;2:223–9.
Szczepanski T, van der Velden VH, Hoogeveen PG, de Bie M, Jacobs DC, van Wering ER, et al. Vdelta2-Jalpha rearrangements are frequent in precursor-B-acute lymphoblastic leukemia but rare in normal lymphoid cells. Blood. 2004;103:3798–804.
van Dongen JJ, Langerak AW, Bruggemann M, Evans PA, Hummel M, Lavender FL, et al. Design and standardization of PCR primers and protocols for detection of clonal immunoglobulin and T-cell receptor gene recombinations in suspect lymphoproliferations: report of the BIOMED-2 Concerted Action BMH4-CT98-3936. Leukemia. 2003;17:2257–317.
van der Velden VH, van Dongen JJ. MRD detection in acute lymphoblastic leukemia patients using Ig/TCR gene rearrangements as targets for real-time quantitative PCR. Methods Mol Biol. 2009;538:115–50.
Cui L, Li Z, Wu M, Li W, Gao C, Deng G. Combined analysis of minimal residual disease at two time points and its value for risk stratification in childhood B-lineage acute lymphoblastic leukemia. Leuk Res. 2010;34:1314–9.
Gundry MC, Brunetti L, Lin A, Mayle AE, Kitano A, Wagner D, et al. Highly Efficient Genome Editing of Murine and Human Hematopoietic Progenitor Cells by CRISPR/Cas9. Cell Rep. 2016;17:1453–61.
Willcockson MA, Taylor SJ, Ghosh S, Healton SE, Wheat JC, Wilson TJ, et al. Runx1 promotes murine erythroid progenitor proliferation and inhibits differentiation by preventing Pu.1 downregulation. Proc Natl Acad Sci U S A. 2019;116:17841–7.
Fitch SR, Kapeni C, Tsitsopoulou A, Wilson NK, Gottgens B, de Bruijn MF, et al. Gata3 targets Runx1 in the embryonic haematopoietic stem cell niche. IUBMB Life. 2020;72:45–52.
Jeong M, Park HJ, Celik H, Ostrander EL, Reyes JM, Guzman A, et al. Loss of Dnmt3a Immortalizes Hematopoietic Stem Cells In Vivo. Cell Rep. 2018;23:1–10.
Gu T, Lin X, Cullen SM, Luo M, Jeong M, Estecio M, et al. DNMT3A and TET1 cooperate to regulate promoter epigenetic landscapes in mouse embryonic stem cells. Genome Biol. 2018;19:88.
Zhang X, Su J, Jeong M, Ko M, Huang Y, Park HJ, et al. DNMT3A and TET2 compete and cooperate to repress lineage-specific transcription factors in hematopoietic stem cells. Nat Genet. 2016;48:1014–23.
Challen GA, Sun D, Jeong M, Luo M, Jelinek J, Berg JS, et al. Dnmt3a is essential for hematopoietic stem cell differentiation. Nat Genet. 2011;44:23–31.
Szemes M, Dallosso AR, Melegh Z, Curry T, Li Y, Rivers C, et al. Control of epigenetic states by WT1 via regulation of de novo DNA methyltransferase 3A. Hum Mol Genet. 2013;22:74–83.
Lin RK, Wang YC. Dysregulated transcriptional and post-translational control of DNA methyltransferases in cancer. Cell Biosci. 2014;4:46.
Chen JS, Hsiao CC, Sheen JM, Cheng CN. Comparison of minimal residual disease (MRD) estimated by flow cytometry and by real-time quantitative PCR of Wilms tumor gene 1 (WT1) transcript expression in children with acute lymphoblastic leukemia. Leuk Res. 2007;31:1351–7.
Oh L, Hainaut P, Blanchet S, Ariffin H. Expression of p53 N-terminal isoforms in B-cell precursor acute lymphoblastic leukemia and its correlation with clinicopathological profiles. BMC Cancer. 2020;20:110.
Acknowledgements
The authors would like to thank all staff members in the hematology oncology center and in the laboratory of hematologic diseases who have taken care of the patients and given the help for the studies.
Funding
This study was supported by the grants from National Natural Science Foundation of China [No. 81870114 and 81970135], Beijing Natural Science Foundation of China [No. 7194263 and 7202044], Scientific Research Common Program of Beijing Municipal Commission of Education [No. KM201810025025], and Beijing Municipal Science & Technology Commission [No. Z181100001718100]. The roles these funders had were showed in part of “Authors’ contributions” in detail.
Author information
Authors and Affiliations
Contributions
ZGL designed this research and revised the manuscript. WJL performed experiments, interpreted/analyzed data, prepared figures and wrote the manuscript. CJW also performed part of the experiments and LC helped in analyzing data and preparing figures. As co-corresponding authors, WL and RDZ contributed equally to study design and organized informed consent. SGL and XXZ contributed patient material and provided clinical data. All authors have read and approved the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This study was submitted to and approved by our institutional ethics committee of Beijing Children’s Hospital Affiliated to Capital Medical University.
Informed consents were obtained from all the children’s parents or legal guardians.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original version of this article was revised: The given name of author Shuguang Liu has been corrected.
Supplementary Information
Additional file 1: Table S1.
Comparison of clinical characteristics in children with B-cell acute lymphoblastic leukemia include or excluded in this study.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, W., Liu, S., Wang, C. et al. DNMT3A low-expression is correlated to poor prognosis in childhood B-ALL and confers resistance to daunorubicin on leukemic cells. BMC Cancer 23, 255 (2023). https://doi.org/10.1186/s12885-023-10724-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12885-023-10724-6