
Abstract
Background
Schizophrenia is a severe neurodevelopmental disorder with a complex genetic and environmental etiology. Abnormal glutamate ionotropic N-methyl-D-aspartate receptor (NMDA) type subunit 1 (NR1) may be a potential cause of schizophrenia.
Methods
We conducted a case-control study to investigate the association between the GRIN1 gene, which encodes the NR1 subunit, and the risk of schizophrenia in a northern Chinese Han population using Sanger DNA sequencing. The dual luciferase reporter assay was used to detect the influence of two different haplotypes on GRIN1 gene expression.
Results
Seven SNPs (single nucleotide polymorphisms), including rs112421622 (− 2019 T/C), rs138961287 (− 1962--1961insT), rs117783907 (−1945G/T), rs181682830 (−1934G/A), rs7032504 (-1742C/T), rs144123109 (−1140G/A), and rs11146020 (−855G/C) were detected in the study population. Rs117783907 (−1945G/T) was associated with the occurrence of schizophrenia as a protective factor. The genotype frequencies of rs138961287 (− 1962--1961insT) and rs11146020 (−855G/C) were statistically different between cases and controls (p < 0.0083). The other four variations were not shown to be associated with the disease. Two haplotypes were composed of the seven SNPs, and distribution of T-del-G-G-C-G-G was significantly different between the case and control groups. However, the dual luciferase reporter assay showed that neither of the haplotypes affected luciferase expression in HEK-293 and SK-N-SH cell lines.
Conclusions
The GRIN1 gene may be related to the occurrence of schizophrenia. Additional research will be needed to fully ascertain the role of GRIN1 in the etiology of schizophrenia.
Similar content being viewed by others


Background
Schizophrenia is a serious genetic disease characterized by emotional impairment, cognitive deficits, and social dysfunction [1]. A large schizophrenia genome-wide association study (GWAS) recently indicated that multiple genes involved in glutamatergic neurotransmission were relevant to schizophrenia [2]. Glutamate is the primary excitatory neurotransmitter in the central nervous system (CNS) and exerts a physiological role by binding to several glutamate receptors. Of the various glutamate receptors, NMDA receptors have received more attention due to their pivotal role in axonal formation, long-term potentiation, and excitotoxicity [3, 4]. Studies have shown that NMDA receptors are closely related to learning and memory [5]. Moreover, a review [6] pointed out that abnormal NMDA receptors are associated with Alzheimer’s disease, Huntington’s disease, and epilepsy. Phenylcyclohexyl piperidine (PCP), which blocks a NMDA glutamate receptor subtype [7], induces a psychotomimetic state that closely resembles schizophrenia [8]. These findings imply that NMDA receptor dysfunction might be involved in the etiology of the CNS disorders [9].
NMDA receptors are heterotetramers composed of two NR1 subunits and NR2 or/and NR3 subunits [10]. The NR1 subunit, which is encoded by the GRIN1 gene, is a functional subunit of the NMDA receptor and is widely distributed throughout the brain [11]. Mice that expressed only 5% normal levels of the NR1 subunit showed increased activity, dullness, and social and sexual deficiencies. Moreover, these behavioral changes were similar to those observed in animal models of schizophrenia [12]. mRNA and protein levels of NR1 subunits were shown to be decreased in the postmortem brain of schizophrenic patients [13, 14]. These studies indicate a potential association between the GRIN1 gene and schizophrenia. Although one study [15] identified 143 differentially expressed proteins in the anterior cingulate cortex between schizophrenia patients and healthy controls, it did not include the NR1 subunit. In addition, genetic association studies showed no significant difference in genotypic and allelic frequency distribution of the GRIN1 gene between schizophrenic and healthy controls in Japanese and Chinese populations [16, 17].
The role of GRIN1 in the etiology of schizophrenia remains uncertain, and genetic association studies of the GRIN1 gene and schizophrenia in the northern Chinese Han population are relatively deficient. We conducted a case-control study to investigate the association between GRIN1 and the risk of schizophrenia in a northern Chinese Han population using Sanger DNA sequencing. Furthermore, the effects of two different haplotypes located in the 5′ promoter region of the GRIN1 gene on protein expression were detected by dual luciferase reporter assay.
Methods
Samples
Blood samples from 316 northern Han Chinese healthy unrelated volunteers (157 females, 159 males, mean age 44 ± 14.3) were provided by China Medical University. Questionnaires showed that there was no history of mental illness within three generations. Blood samples from 309 northern Han Chinese patients with schizophrenia (156 females, 153 males, average age 41 ± 14.6) were provided by the Third People’s Hospital of Liaoning Province. The diagnosis of schizophrenia was in accordance with The Diagnostic and Statistical Manual of Mental Disorders (fourth edition). To confirm the diagnoses, two independent senior psychiatrists reviewed psychiatric medical records. Genomic DNA was extracted from peripheral blood by the standard phenol-chloroform method. The study was approved by the Ethics Committee of China Medical University, and written informed consent was obtained from each participant and/or patient guarantor.
PCR amplification
Polymerase chain reaction (PCR) was used to amplify the GRIN1 fragment, including the 5′ flanking and untranslated regions. The nucleotide position of the target fragment amplified was from − 2334 to + 86 (with ATG + 1). Genomic DNA (1 μL, about 30 ng) was amplified under the following reaction contents: 1 μL (5 pmol) each of sense and antisense primers, 2 μL (3 nmol) of dNTP mix, 0.2 μL (about 0.5 U) of PrimeSTAR® HS DNA polymerase (Takara, Dalian, China) and 10 μL 2 × Prime STAR HS GC buffer. Sterilized deionized water was added to a volume of 20 μL. PCR cycling conditions were 94 °C for 1 min; 30 cycles at 98 °C for 10 s, at 60 °C for 5 s, and 72 °C for 2 min 30 s; and 72 °C for 10 min. PCR products were separated by 1% agarose gel electrophoresis.
DNA sequencing
DNA was sequenced using Sanger DNA sequencing (Taihe Biotechnology Co. Ltd. Beijing China). Primer information was shown in Table 1.
Construction of pGL-3 recombinant vector
The target fragment located at − 2143 - + 222 (with ATG + 1) was amplified using a PrimerSTAR® kit (Takara, Dalian, China). The sense primer was 5’ GGCTAGCCTGAACATTTAGCGATCA 3′ and the antisense primer was 5’ CAGATCTGGCATTGAGCTGAATCTTC 3′. The primers contained NheI or BglII restriction endonuclease sites at the 5′ end. By using the pGM-T Ligation® Kit (TIANGEN, Beijing, China), PCR products purified from agarose gel were cloned into pGM-T vectors, and the recombinant vectors were subsequently transformed into JM109 competent cells. The pGM-T recombinant plasmids extracted by SanPrep® Column Enodotoxin-Free Plasmid Mini-Preps Kit (Sangon Biotech, Shanghai, China) were subjected to Sanger DNA sequencing to ensure correct insertion of the target fragment.
The two different haplotype pGM-T recombinant vectors were subcloned into the pGL-3 Basic Vector (Promega, Madison, Wisconsin, USA) using NheI or BglII.
Cell culture
Human embryonic kidney cell line HEK-293 and neuroblastoma cell line SK-N-SH were used to test luciferase activity of the pGL-3 recombinant vectors. HEK-293 cells were cultured in HyClone® DMEM high glucose medium with 10% fetal bovine serum (Thermo Fisher Scientific, Massachusetts, USA). SK-N-SH cells were cultured in KeyGRN BioTECH® DMEM high glucose medium (with 0.011 g/L sodium pyruvate) with 15% fetal bovine serum. Cells were seeded in 24-well plates (2 × 105 cells per well). According to the manufacturer’s protocol (Invitrogen, California, USA), Lipofectamine®3000 reagent was used to co-transfect the pGL-3 recombinant plasmids containing the two haplotypes with the Renilla luciferase-expressing vector pRL-TK (Promega) into the two cell lines. Cells were harvested after 24 h in culture. Firefly luciferase activity (LUC value) was measured and normalized to renilla luciferase activity (TK value). Each assay was performed in triplicate in the two cell lines.
Statistical analysis
SPSS 20.0 software (IBM, Armonk, NY, USA) was used to analyze genotype frequency, allele frequency, haplotype frequency, and LUC/TK values (relative fluorescence intensity). Haploview 4.2 software (Broad Institute, Cambridge, MA, USA) was used for the Hardy–Weinberg equilibrium test and to confirm haplotypes. The χ2 test was used to examine the distribution of genotypes, alleles, and haplotypes between groups. The threshold for polymorphism multiple correction was p = 0.05/6, and the haplotype multiple test threshold after Bonferroni correction was 0.05/2. Independent sample T test was used to compare the relative fluorescence intensity of the two haplotypes. Relative fluorescence intensity was expressed as the mean ± standard deviation, and p < 0.05 (two sided) represented a significant difference. Power analysis was conducted using PS program [18] statistical software. Detection of GRIN1 gene expression in different tissues was performed with the GTEx database (https://gtexportal.org/home/).
Results
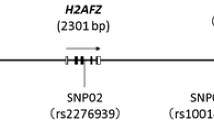
Seven common SNPs (Fig. 1), including rs112421622 (− 2019 T/C), rs138961287 (− 1962--1961insT), rs117783907 (−1945G/T), rs181682830 (−1934G/A), rs7032504 (-1742C/T), rs144123109 (−1140G/A) and rs11146020 (−855G/C), were detected in the 5′ promoter region of the GRIN1 gene in healthy Chinese Han individuals. The distribution of all seven SNPs was in accordance with Hardy-Weinberg equilibrium in the control group (p > 0.05). Results of linkage disequilibrium analysis using Haploview 4.2 software are presented in Fig. 2, showing that rs138961287 (− 1962 - -1961insT) and rs117783907 (−1945G/T) are in linkage disequilibrium (D’ = 0.99, r2 = 0.94). Given that there were six independent SNPs, the threshold for polymorphism multiple correction was p = 0.05/6 = 0.0083.

Schematic diagram of the distribution of seven SNP loci in the GRIN1 gene. The detailed position of the fragment used for Sanger DNA sequencing in the GRIN1 gene 5′ promoter region was located between − 2334 bp and + 86 bp. * denotes the SNP not included in the genetic association study because the minimum allele frequency was less than 0.01

Linkage disequilibrium diagram of seven SNP loci in the GRIN1 gene. Left is D’ in the control group, and the red grid represents D’ = 1. Right is r2 in the control group, and the black grid represents r2 = 1
Both genotype and allele distribution (Additional file 1) of rs117783907 (−1945G/T) were significantly different between the case and control groups (p < 0.0083). The frequency of the T allele in rs117783907 (−1945G/T) in the case group (15.2%) was much lower than in the control group (21.7%). The genotype frequencies of rs138961287 (− 1962--1961insT) and rs11146020 (−855G/C) were statistically different between cases and controls (p < 0.0083). Rs112421622 (− 2019 T/C), rs181682830 (−1934G/A), rs7032504 (-1742C/T), and rs144123109 (−1140G/A) were detected only in two genotypes and were determined to be unrelated to schizophrenia. The power of (− 1962 - -1961insT), rs117783907 (−1945G/T), and rs11146020 (−855G/C) were all > 0.70, and the power of rs117783907 (−1945G/T) in particular reached 0.982 (Additional file 1). According to the GTEx database, the GRIN1 gene is highly expressed in brain tissue compared with other tissues (Fig. 3).

GRIN1 gene expression in different tissues. TPM on the vertical axis represents the transcript quantification value, and the horizontal axis represents different tissues. The GRIN1 gene was highly expressed in brain tissue compared with other tissues
A total of seven SNPs formed six haplotypes, but after excluding four with very low frequencies (< 5%), only haplotypes T-del-G-G-C-G-G and T-insT-T-G-C-G-C remained. Therefore, the haplotype multiple test threshold after Bonferroni correction was 0.05/2 = 0.025. We found a significant difference in the haplotype frequency distribution of T-del-G-G-C-G-G between case and control groups (Additional file 2), with p values of 0.005. Specifically, the frequency of haplotype T-del-G-G-C-G-G in the case group (77.8%) was much higher than in the control group (70.9%), increasing the disease risk (OR = 1.442, 95% CI: 1.116–1.862). Haplotype T-insT-T-G-C-G-C was not related to schizophrenia.
Comparing the LUC/TK values of the two haplotypes (Fig. 4), we found that the relative fluorescence intensity of the recombinant vector T-del-G-G-C-G-G was higher than T-T-T-G-C-G-C in HEK-293 cells. However, the difference was not statistically significant (p = 0.178). In SK-N-SH cells, no statistical difference in relative fluorescence intensity was found between the recombinant vectors T-del-G-G-C-G-G and T-T-T-G-C-G-C (p = 0.956).

Relative fluorescence intensity of two different recombinant haplotypes in HEK-293 and SK-N-SH cells. a and b represent the relative fluorescence intensity (LUC/TK values) of two haplotypes in HEK-293 cells and SK-N-SH cells, respectively. Del-G-G represents the haplotype T-del-G-G-C-G-G; T-T-C represents the haplotype T-insT-T-G-C-C. There was no significant difference in relative fluorescence intensities between the T-del-G-G-C-G-G and T-insT-T-G-C-G-C haplotypes in HEK-293 (p = 0.178) or SK-N-SH cells (p = 0.959)
Discussion
In the present study, we examined the association between the GRIN1 gene and the risk of schizophrenia in a northern Chinese Han population. Using Sanger DNA sequencing, we detected seven SNPs, including rs112421622 (− 2019 T/C), rs138961287 (− 1962--1961insT), rs117783907 (−1945G/T), rs181682830 (−1934G/A), rs7032504 (-1742C/T), rs144123109 (−1140G/A) and rs11146020 (−855G/C). Novel mutations were not detected.
Both genotype and allele distribution of rs117783907 (−1945G/T) were significantly different between the case and control groups (p < 0.0083). The genotype frequencies of rs138961287 (− 1962--1961insT) and rs11146020 (−855G/C) were statistically different between the case and control groups (p < 0.05), indicating that rs138961287 (− 1962--1961insT), rs117783907 (−1945G/T) and rs11146020 (−855G/C) are related to schizophrenia. Another association study in a northern Chinese Han population reported that the C allele frequency of rs11146020 reduced the risk of schizophrenia [19], although this allele was reported to be associated with schizophrenia as a risk factor in an Italian population [20]. Furthermore, a meta-analysis [21] suggested that the C variation allele of rs11146020 might be associated with an increased risk for developing schizophrenia.
The risk of developing psychosis increases with the accumulation of many genetic risk variants and exposure to multiple adverse environmental factors [22]. In different ethnic groups, the effects of environmental and genetic factors and their interactions on the risk of disorders may vary [21]. It has been shown that some genetic polymorphisms might be associated with altered risk in schizophrenia [23] and immune system diseases among different ethnicities [24,25,26]. Interestingly, recent evidence suggests that patients with schizophrenia might display signs of typical autoimmune processes associated with impaired functioning of microRNAs [27]. Thus, the opposite effect of this same C allele of rs11146020 (−855G/C) on schizophrenia in different ethnic groups is likely due to genetic heterogeneity.
There have been no studies involving the two SNPs in linkage disequilibrium rs138961287 (− 1962--1961insT) and rs117783907 (−1945G/T). The frequency of the T allele of rs117783907 (−1945G/T) in the case group was significantly lower than in the control group, indicating that it could be a protective factor in individuals of northern Han Chinese descent with schizophrenia.
According to the GTEx database, the GRIN1 gene was highly expressed in brain tissues compared to other tissues, suggesting that GRIN1 may play an important role in the CNS. Pranita et al. [28] found no significant risk for schizophrenia at the rs11146020 C allele. However, schizophrenic individuals in that study were being treated with antipsychotic drug therapy. Rice et al. [29] reported no association between the GRIN1 gene and schizophrenia. However, the sample size was relatively small, with only five cases in an Asian population. Georgi et al. [30] detected distribution of four single nucleotide polymorphisms and one microsatellite marker at GRIN1. They found significant associations between schizophrenia and these polymorphisms in single-marker and haplotype-based analyses. Chanasong et al. [31] found that the A allele of rs1126442 was associated with increased risk of METH-dependent psychosis in a Thai population. Leuba et al. [32] indicated that patients with Alzheimer’s disease had significantly lower NR1 subunit levels in the entorhinal cortex and the frontal cortex. Wu et al. [33] genotyped six polymorphisms of GRIN1 and GRIN2B and found that the two genes in conjunction with each other were associated with Parkinson’s disease in a Chinese population.
We defined two haplotypes (frequencies > 0.5) in the 5′ promoter region of the GRIN1 gene. Haplotype T-del-G-G-C-G-G was statistically associated with increased risk of schizophrenia. Transcription factors regulate gene expression by interacting with cis-regulatory elements. Rice et al. [29] and Begni et al. [20] found that rs11146020 (−855G/C) may change the + 1 G of the transcription factor NF-κB consensus sequence (GGGG). However, we failed to detect significantly different luciferase expression between the two haplotypes in both the HEK-293 and SK-N-SH cell lines. In fact, the non-significant effects of the two haplotypes on luciferase expression is not inconsistent with the conclusion that the GRIN1 gene is associated with schizophrenia. Because more than one transcription factor binds to cis-regulatory elements in the 5′ promoter region of the GRIN1 gene, they likely reinforce or offset each other. A genome-wide association study [2] recently showed that multiple genes involved in the glutamatergic system and the DRD2 gene in the dopamine system acted on schizophrenia, implying that the two systems do not work independently in schizophrenia. Similarly, it is reasonable to speculate that GRIN1 and other interactional genes likely affect the function of GRIN1 polymorphisms. Further studies are needed to verify this postulation.
There were several limitations in this study. First, in addition to the seven SNPs, another polymorphism (Fig. 1), rs1003711879 (− 289--287delGCC), was also detected. Unfortunately, due to the relatively small sample size, rs1003711879 (− 289 - -287delGCC) could not be included in the genetic association study because the frequency did not reach 1%. Second, only HEK-293 and SK-N-SH cell lines were used for functional experiments, and use of additional related cell lines may provide more information.
Conclusions
Using Sanger DNA sequencing, we illustrated that rs11146020, rs138961287, and rs117783907 in the promoter region of the GRIN1 gene are associated with schizophrenia in a northern Chinese Han population. Haplotype T-del-G-G-C-G-G might significantly increase the risk of schizophrenia, while T-insT-T-G-C-G-C is likely not related to disease risk. However, the dual luciferase reporter assay showed that neither of the haplotypes affected luciferase expression. Future efforts will be needed to ascertain the role of GRIN1 in the etiology of schizophrenia.
Abbreviations
- 95% CI:
-
95% confidence interval
- ATG:
-
transcription start site
- CDS:
-
coding sequence
- OR:
-
odds ratio
- SNP:
-
single nucleotide polymorphism
References
Xu Y, Ren J, Ye H. Association between variations in the disrupted in schizophrenia 1 gene and schizophrenia: A meta-analysis. Gene. 2018;651:94–9.
Schizophrenia Working Group of the Psychiatric Genomics C. Biological insights from 108 schizophrenia-associated genetic loci. Nature. 2014;511:421–7.
Rabacchi S, Bailly Y, Delhaye-Bouchaud N, Mariani J. Involvement of the N-methyl D-aspartate (NMDA) receptor in synapse elimination during cerebellar development. Science. 1992;256:1823–5.
Wood MW, VanDongen HM, VanDongen AM. The 5′-untranslated region of the N-methyl-D-aspartate receptor NR2A subunit controls efficiency of translation. J Biol Chem. 1996;271:8115–20.
Morris RG. NMDA receptors and memory encoding. Neuropharmacology. 2013;74:32–40.
Hansen KB, Yi F, Perszyk RE, Menniti FS, Traynelis SFNMDA. Receptors in the Central Nervous System. Methods Mol Biol. 2017;1677:1–80.
Moghaddam B, Krystal JH. Capturing the angel in “angel dust”: twenty years of translational neuroscience studies of NMDA receptor antagonists in animals and humans. Schizophr Bull. 2012;38:942–9.
Javitt DC, Zukin SR. Recent advances in the phencyclidine model of schizophrenia. Am J Psychiatry. 1991;148:1301–8.
Zhou Q, Sheng M. NMDA receptors in nervous system diseases. Neuropharmacology. 2013;74:69–75.
Vyklicky V, Korinek M, Smejkalova T, Balik A, Krausova B, Kaniakova M, et al. Structure, function, and pharmacology of NMDA receptor channels. Physiol Res. 2014;63(Suppl 1):S191–203.
Wang H, Wang X, Li Y, Yu H, Wang C, Feng C, et al. Chronic ethanol exposure induces SK-N-SH cell apoptosis by increasing N-methyl-D-aspartic acid receptor expression and intracellular calcium. Exp Ther Med. 2018;15:3791–800.
Mohn AR, Gainetdinov RR, Caron MG, Koller BH. Mice with reduced NMDA receptor expression display behaviors related to schizophrenia. Cell. 1999;98:427–36.
Weickert CS, Fung SJ, Catts VS, Schofield PR, Allen KM, Moore LT, et al. Molecular evidence of N-methyl-D-aspartate receptor hypofunction in schizophrenia. Mol Psychiatry. 2013;18:1185–92.
Catts VS, Derminio DS, Hahn CG, Weickert CS. Postsynaptic density levels of the NMDA receptor NR1 subunit and PSD-95 protein in prefrontal cortex from people with schizophrenia. NPJ Schizophr. 2015;1:15037.
Focking M, Lopez LM, English JA, Dicker P, Wolff A, Brindley E, et al. Proteomic and genomic evidence implicates the postsynaptic density in schizophrenia. Mol Psychiatry. 2015;20:424–32.
Tani A, Kikuta R, Itoh K, Joo A, Shibata H, Ninomiya H, et al. Polymorphism analysis of the upstream region of the human N-methyl-D-aspartate receptor subunit NR1 gene (GRIN1): implications for schizophrenia. Schizophr Res. 2002;58:83–6.
Hung CC, Chen HY, Chen CH. Systematic mutation analysis of the human glutamate receptor, ionotropic, N-methyl-D-aspartate 1 gene(GRIN1) in schizophrenic patients. Psychiatr Genet. 2002;12:225–30.
Xu FL, Ding M, Yao J, Shi ZS, Wu X, Zhang JJ, et al. Association between mitochondrial DNA variations and schizophrenia in the northern Chinese Han population. PloS one. 2017;12:e0182769.
Zhao X, Li H, Shi Y, Tang R, Chen W, Liu J, et al. Significant association between the genetic variations in the 5′ end of the N-methyl-D-aspartate receptor subunit gene GRIN1 and schizophrenia. Biol Psychiatry. 2006;59:747–53.
Begni S, Moraschi S, Bignotti S, Fumagalli F, Rillosi L, Perez J, et al. Association between the G1001C polymorphism in the GRIN1 gene promoter region and schizophrenia. Biol Psychiatry. 2003;53:617–9.
Saadat M. N-methyl-D-aspartate receptor NR1 subunit gene (GRIN1) G1001C polymorphism and susceptibility to schizophrenia: A meta-analysis. EXCLI J. 2010;9:11–6.
Zwicker A, Denovan-Wright EM, Uher R. Gene-environment interplay in the etiology of psychosis. Psychol Med. 2018;48:1925–36.
Lam M. Comparative genetic architectures of schizophrenia in East Asian and European populations; 2018.
Liu X, Qin H, Wu J. Xu J. Association of TNFAIP3 and TNIP1 polymorphisms with systemic lupus erythematosus risk: A meta-analysis. Gene. 2018;668:155–65.
Hua J, Huang W. Peptidylarginine deiminase 4 -104C/T polymorphism and risk of rheumatoid arthritis: A pooled analysis based on different populations. PloS one. 2018;13:e0193674.
Lu C, Xu K, Guo H, Peng K, Yang Z, Hao YQ, et al. The relationship of PADI4_94 polymorphisms with the morbidity of rheumatoid arthritis in Caucasian and Asian populations: a meta-analysis and system review. Clin Rheumatol. 2018;37:289–96.
Ermakov EA, Ivanova SA, Buneva VN, Nevinsky GA. Blood-Derived RNA- and microRNA-Hydrolyzing IgG Antibodies in Schizophrenia Patients. Biochemistry (Mosc). 2018;83:507–26.
Pai P, Arathil P, Kotambail A, Nair R, Gupta M, Moily NS, et al. Association of GRIN1, ABCB1, and DRD4 genes and response to antipsychotic drug treatment in schizophrenia. Psychiatr Genet. 2015;25:135–6.
Rice SR, Niu N, Berman DB, Heston LL, Sobell JL. Identification of single nucleotide polymorphisms (SNPs) and other sequence changes and estimation of nucleotide diversity in coding and flanking regions of the NMDAR1 receptor gene in schizophrenic patients. Mol Psychiatry. 2001;6:274–84.
Georgi A, Jamra RA, Klein K, Villela AW, Schumacher J, Becker T, et al. Possible association between genetic variants at the GRIN1 gene and schizophrenia with lifetime history of depressive symptoms in a German sample. Psychiatr Genet. 2007;17:308–10.
Chanasong R, Thanoi S, Watiktinkorn P, Reynolds GP, Nudmamud-Thanoi S. Genetic variation of GRIN1 confers vulnerability to methamphetamine-dependent psychosis in a Thai population. Neurosci Lett. 2013;551:58–61.
Leuba G, Vernay A, Kraftsik R, Tardif E, Riederer BM, Savioz A. Pathological reorganization of NMDA receptors subunits and postsynaptic protein PSD-95 distribution in Alzheimer’s disease. Curr Alzheimer Res. 2014;11:86–96.
Wu SL, Wang WF, Shyu HY, Ho YJ, Shieh JC, Fu YP, et al. Association analysis of GRIN1 and GRIN2B polymorphisms and Parkinson’s disease in a hospital-based case-control study. Neurosci Lett. 2010;478:61–5.
Acknowledgements
We would like to express our gratitude to Professor Hao Pang for his theoretical guidance.
Funding
This work was supported by the National Natural Science Foundation of China (No. 81601653). The funders had no role in the design of the study nor in the collection, analysis, and interpretation of data or in writing the manuscript.
Availability of data and materials
Raw data of the study will be available upon request after publication.
Author information
Authors and Affiliations
Contributions
YPL, XCZ, and YL designed the study. YPL performed the assay and drafted the manuscript. JXX, JFX, and XX were involved in the sample collection. MD performed statistical analysis. JY and BJW revised the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Written informed consent was obtained from all subjects and/or patient guarantors. The study was approved by the Ethics Committee of China Medical University.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
Genotype and allele distribution of SNPs in the GRIN1 gene (significance threshold = 0.0083). a Frequency is shown in brackets (%); b P value shown in bold reaches a significant level (P < 0.0083); the genotype and allele frequency distribution of -1945G/T and the genotype frequency of -855G/C and − 1962 - -1961insT were significantly different between the case and control groups. (DOC 77 kb)
Additional file 2:
Haplotypes formed by the seven SNPs in GRIN1 and disease risk (significance level = 0.025). Del-G-G represents the haplotype T-del-G-G-C-G-G, T-T-C represents the haplotype T-insT-T-G-C-C.The frequency distribution of haplotype T-del-G-G-C-G-G was significantly different between the case group and the control group, with p values of 0.005. Specifically, the frequency of haplotype T-del-G-G-C-G-G in the case group (77.8%) was much higher than in the control group (70.9%), increasing the disease risk (OR = 1.442, 95% CI: 1.116–1.862). Haplotype T-insT-T-G-C-G-C was not related to schizophrenia (XLSX 12 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Liu, Yp., Ding, M., Zhang, Xc. et al. Association between polymorphisms in the GRIN1 gene 5′ regulatory region and schizophrenia in a northern Han Chinese population and haplotype effects on protein expression in vitro. BMC Med Genet 20, 26 (2019). https://doi.org/10.1186/s12881-019-0757-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12881-019-0757-3