
Abstract
Background
Hypertension is a multifactorial disease involving both environmental and genetic Factros. G894 T eNOS polymorphism has been suggested to be responsible for reduced NO synthesis, and EH development. The objective of our case-control study is to evaluate the potential association of G894 T eNOS polymorphism with Essential Hypertension (EH) susceptibility, among a sample of Moroccan patients.
Methods
One hundred forty five hypertensive patients were recruited from the department of Cardiology, University Hospital Center Ibn Rochd, Casablanca, Morocco, and compared to 184 apparently healthy subjects. DNA samples were genotype by PCR-RFLP method using MboI restriction enzyme.
Results
Our results showed a positive correlation between G894 T eNOS distribution and Alcohol and Obesity rik factors (P = 0.009 and 0.02 respectively). Patients with elevated Cardio Vascular Risk (CVR) carried out the higher frequency of homozygous mutant genotype TT (62.2%) and T mutant allele (77.8%), compared to median and low CVR groups. G894 T eNOS distribution was significantly associated to a high risk of EH occurrence under the GT and TT genotypes (OR [95% CI] = 20.2 [7.7–52.4], P < 0.0001; OR [95% CI] = 332.5 [98.2–1125.4], P < 0.0001 respectively), and the 3 genotypic transmission models (Dominant: OR [95% CI] = 43.2 [17.9–104.09], P < 0.0001; Recessive: OR [95% CI] = 47.7 [18.6–122.3]; P < 0.0001; Additive: OR [95% CI] = 14.02 [9.6–20.45], P < 0.0001).
Conclusion
Our study suggests a strong association of G894 T eNOS polymorphism with susceptibility to EH in Morocco. Studies trying to identify contributing genes may be very useful and allow recognizing the vulnerable individuals and classifying patients in subgroups with definite genetic and pathogenic mechanisms to achieve better prevention and therapeutics.
Similar content being viewed by others

Background
Hypertension is a multifactorial disease involving both environmental and genetic components. Alterations in endothelial-derived Nitric Oxide (NO) production have been associated with numerous diseases [1, 2], and, in humans, can be genetically determined by the presence of different polymorphisms in the eNOS gene.
Synthesized by endothelial cells, NO contributes to the vasodilatation process and regulation of Blood Pressure (BP) [1]. Its production in vascular endothelium cells is controlled by endothelial Nitric Oxide Synthase (eNOS) gene, from L-arginine oxidation [3].
The gene encoding eNOS is located on chromosome 7q35–36, and composed of 25 introns and 26 exons that encode a 135 KDa protein, containing 1203 amino acids (Fig. 1) [4, 5]. eNOS gene is highly polymorphic. The G894 T (Glu298Asp) variant, located in exon 7, is the most described and has been suggested to be responsible for reduced NO synthesis, and EH development [6]. This variant alters the primary structure of the protein and has the potential to alter one or more functional properties of the enzyme directly. Two different studies have shown the eNOS protein containing Asp at position 298 to be subject to selective proteolytic cleavage in endothelial cells and vascular tissues [7, 8]. If this observation is correct, the cleaved fragments would be expected to lack NO synthase activity. However, two other reports suggest that this observation might be an artifact [9, 10]. The Glu298Asp SNP affects also eNOS localization to caveolar membrane [11].

Scheme of the gene encoding endothelial nitric oxide synthase gene (eNOS) (a and b) [18]. Scheme of the gene encoding endothelial nitric oxide synthase (eNOS or NOS3). The human NOS3 gene (located at 7q35-36) contains 26 exons that span 21 kb. Exons are described by number. AUG: the transcription start site. Three specific polymorphisms in the NOS3 gene are marked by arrows. c Scheme of the eNOS protein
As there is lack of data concerning the potential association of G894 T eNOS polymorphism with EH susceptibility in North African populations and especially in the Moroccan one, we have preceded to the evaluation of its effect on EH predisposition in a sample of Moroccan patients compared to healthy subjects.
Methods
Study population
Blood samples were collected from 145 hypertensive patients, recruited from the department of Cardiology, University Hospital Center Ibn Rochd, Casablanca, Morocco, and compared to 184 apparently healthy subjects. Patients with a mean systolic blood pressure (SBP) ≥140 mmHg, mean diastolic blood pressure (DBP) ≥90 mmHg or taking antihypertensive therapy were declared hypertensive and were recruited. However, those presenting symptoms or signs suggesting secondary origins of hypertension were excluded from the study. For healthy subjects, they did not show any abnormalities concerning the physical status, blood pressure and family history of hypertension. Patients’ Clinical data were collected and an informed consent, approved by the Ethical Committee of the University of Hassan II, School of Medicine, Casablanca, was signed by each patient and control before entering the study.
DNA extraction
Venous blood from all participants in this study was collected in EDTA tubes. Samples were stored at − 20 °C until extraction of DNA. Genomic DNA was extracted from blood leukocytes using the standard method of salting out [12].
Genotype determination
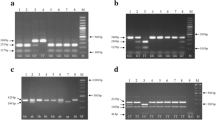
We used PCR-RFLP to genotype samples for +894G/T eNOS polymorphism as previously described by [13]. We have proceeded to an amplification of 50–100 ng of extracted DNA, followed by digestion using MboI restriction enzyme, which gave rise to three profiles: homozygous wild type GG (one fragment of 206 bp), heterozygous GT (three fragments of 206, 119 and 87 bp), and homozygous mutated TT (two fragments of 119 and 87 bp) (Fig. 2). The digested products were separated on 3% agarose gel electrophoresis stained with Ethidium Bromide (BET), and visualized with UV rayons.

PCR-RFLP analysis on agarose gel of G894T eNOS polymorphism. M: size marker; Lane 1: GG; Lane 2: GT; Lane 3: TT
Statistical analysis
Statistical analysis was performed using SPSS 21.0 software. Chi square test (χ2) and a P value < 0.05 were used to determine statistical significance of association/non-association between genotypes and classical risk factors. Hardy–Weinberg Equilibrium test (HWE) was performed in both cases and controls groups for the analyzed polymorphism. Odd ratio (OR) was calculated to estimate the association between genotypes and EH risk, using a Confidence Interval (CI) of 95%. Significance was approved at P-value less than 0.05.
Results
Genotypic distribution of the G894 T eNOS polymorphism was in Hardy-Weinberg Equilibrium (HWE) among both cases and controls (Table 1). The average age of our patients was 61.15 ± 10.23 Vs 54.15 ± 2 for healthy subjects. There was a predominance of female in the cases group compared to male (80% Vs 20% respectively).
Table 1 shows the correlation between clinical parameters of our patients and their G894 T eNOS genotypes distribution. There was a statistically significant association only with Alcohol (P = 0.009) and Obesity (P = 0.02); a tendency to a significant association was found with age (P = 0.07), Physical activity (P = 0.05) and personal cardiovascular history (P = 0.07).
Correlation of cardiovascular risk (CVR) status to G894 T eNOS allelic and genotypic distributions, shows no statistically significant association (P = 0.5), even patients with elevated CVR carried out the higher frequency of homozygous mutant genotype TT (62.2%) and T mutant allele (77.8%), compared to median and low CVR groups (Table 2).
Table 3 reports the allelic and genotypic distribution of G894 T eNOS polymorphism, among cases and controls. Our results showed that 77.93% of patients were carrying the mutant allele T, 59.27% of them having the homozygous mutant profile, and 37.24% that were heterozygous. The majority of healthy subjects (79.89%) carried out the wild type allele G, with 63.04% of them having the homozygous wild type genotype GG. G894 T eNOS polymorphism was significantly associated to a high risk of HTA occurrence under the GT and TT genotypes (OR [95% CI] = 20.2 [7.7–52.4], P < 0.0001; OR [95% CI] = 332.5 [98.2–1125.4], P < 0.0001 respectively), and also the 3 genotypic transmission models (Dominant: OR [95% CI] = 43.2 [17.9–104.09], P < 0.0001; Recessive: OR [95% CI] = 47.7 [18.6–122.3]; P < 0.0001; Additive: OR [95% CI] = 14.02 [9.6–20.45], P < 0.0001).
Discussion
The human eNOS gene is highly polymorphic. Results about the association of eNOS Single Nucleotide Polymorphisms with Essential Hypertension development are often controversial and inconclusive [14,15,16,17,18]. Many investigators studied the relationship between G894 T eNOS polymorphism and hypertension. There is an apparent discrepancy among results in these association studies [19,20,21,22,23,24,25]. It was found to be associated with increased risk of hypertension in Caucasians [23, 24], but in another two studies it was not (in neither Caucasians nor Japaneses) [26, 27].
NO is known to be an endothelium-derived relaxing factor and a local blood flow regulator in healthy humans [23, 24, 28]; these facts led to the hypothesis that a reduction in the activity of the endothelial L-arginine–NO pathway could initiate or contribute to the development of Essential Hypertension by promoting and sustaining the observed increase in peripheral resistance [14, 29].
Some researchers demonstrated that inhibition of eNOS elevates Blood Pressure in healthy humans, and disruption of the eNOS gene leads to Hypertension in mice [15, 16]. Analyses of genetically engineered animals deficient in eNOS expression (eNOS−/−) provide support for the importance of eNOS-derived NO, especially in cardiovascular homeostasis. For instance, eNOS−/− mice develop numerous vascular disease states, including systemic hypertension [15, 30]. Moreover, patients with Essential Hypertension have either diminished whole body NO production or increased inactivation leading to lower plasma levels [31,32,33,34]. These results strongly implicate genetic alterations in the eNOS gene in the pathogenesis of human EH [35].
To investigate the potential implication of G894 T eNOS polymorphism in EH, we performed a case-control study among a sample of Moroccan EH patients. Our results showed –first- that Alcohol and obesity were the clinical parameters significantly associated with G894 T eNOS distribution (P = 0.009 and 0.02 respectively); a tendency to a statistically significant association was also found with ‘Age’, Physical activity’ and ‘Personal history of cardiovascular disease’ (P = 0.07, 0.05 and 0.07 respevtively) (Table 1). As known, the pathogenesis of hypertension may be explained by the interaction of many factors such as sodium intake, overweight, alcohol, smoke and the genetic background of subjects [36, 37].
Correlation between cardiovascular risk (CVR) status and G894 T eNOS distribution showed no statistically significant association (P = 0.5); even patients having elevated CVR carried out the higher frequency of mutant homozygous genotype TT (62.2%) (Table 2).
We found a statistically very significant association of this variant of eNOS gene with increased risk of EH development in our study sample, under the GT and TT genotypes (OR [95% CI] = 20.2 [7.7–52.4], P < 0.0001; OR [95% CI] = 332.5 [98.2–1125.4], P < 0.0001 respectively), and also the 3 genotypic transmission models (Dominant: OR [95% CI] = 43.2 [17.9–104.09], P < 0.0001; Recessive: OR [95% CI] = 47.7 [18.6–122.3]; P < 0.0001; Additive: OR [95% CI] = 14.02 [9.6–20.45], P < 0.0001). However, studies results about the association of this variant with susceptibility to EH are controversial and divergent [14,15,16,17]. Many of these studies suggested that inhibition of eNOS pathway lead to BP elevation by low NO production [14, 29]. Study of Wenru Tang et al... [35], analyzing the association of G894 T eNOS polymorphism with EH predisposition, among two Chinese populations (Hani and Yi), reported that –in Hani population- the 894 T allele was associated with increased risk of pathology development, while, in Yi population, the association was found with the 894G allele. Shoji M et al [21] also reported an association of the 894 T allele with EH susceptibility and elevated BP among Japanese subjects. Several studies reported similar results [23, 28], when others found controversial results [19, 20, 22, 26, 27, 38].
This inconsistency in results can be explained by the fact that many of these studies didn’t take in consideration the rest of risk factors that may influence the susceptibility to EH, such as ethnicity (Table 4), age, gender, obesity… [39]. eNOS enzyme acts as a homodimer, functionally composed of two major domains: a C-terminal reductase domain and an N-terminal oxygenase domain. Besides the cofactors tetrahydrobiopterin, flavin adenine dinucleotide, flavin mononucleotide, and calmodulin, eNOS catalytic activity requires the presence of a heme domain [40]. Studies analyzing the structure of eNOS protein have demonstrated that the Glu298 variant of eNOS humain gene is located on the catalytic site of the heme part of the protein, in contact with the protein-protein interaction zone, sensitively affected by Glu298Asp substitution [41]. In his study, Tesauro et al. suggests that the Glu298 and Asp298 variants are differentially treated in cells: Asp298 is prone to natural cleavage by proteases that cleave the eNOS protein in the Glu/Asp 298 substitution site [42]. These findings suggest that Glu/Asp 298 substitution may contribute to the pathogenesis of EH and modulate the therapeutic response of some patients against anti-hypertensive agents [43], leading to different BP responses and cardiovascular outcomes [44, 45].
In the present study, some limitations have to be noted; first the small sample size of patients and controls that was analyzed (145 and 184 respectively); the lack of serum NO concentration measurement; It should also be noted that hypertension is a complex disease; other candidate genes may contribute to the susceptibility of hypertension, and thus must be analyzed. Further studies including larger sample sizes, and overcoming the current limitations might be very useful to better understand the genetic predisposition to hypertension in our population.
Conclusion
Our study is the first in Morocco to evaluate the association of G894 T eNOS polymorphism with EH risk of occurrence. Our results suggest a strong association of this variant of eNOS gene with EH susceptibility in our study sample. Genetic factors contribute to 30–50% of BP variability in human EH. Studies trying to identify contributing genes may provide useful information and allow recognizing the vulnerable individuals and classifying patients in subgroups with definite genetic and pathogenic mechanisms to achieve better prevention and therapeutics. In this context, further analyses still be needed, in order to investigate eNOS adjacent markers in a wider context; future studies should also focus on gene–gene and gene–environment interactions, as well as haplotype patterns.
Abbreviations
- BP:
-
Blood Pressure
- CI:
-
Confidence Interval
- EH:
-
Essential Hypertension
- eNOS:
-
endothelial oxide nitric synthase
- G894 T eNOS:
-
Substitution G/T on 894 position of eNOS gene sequence
- OR:
-
Odds Ratio
- PCR:
-
Polymerase Chaine Reaction
- RFLP:
-
Restriction Fragment Length Polymorphism
References
Idrissi HH, Hmimech W, Diakite B, Korchi F, Baghdadi D, Habbal R, Nadifi S. Association of G894T eNOS, 4G/5G PAI and T1131C APOA5 polymorphisms with susceptibility to myocardial infarction in Morocco. Meta Gene. 2016;9:56–61.
Diakite B, Hamzi K, Slassi I, Yahyaoui EL, Mohammed ELA, Moulay MF, Habbal R, Sellama N, GMRAVC. G894T Endothelial Nitric Oxide Synthase Polymorphism and Ischemic Stroke in Morocco. Meta Gene. 2014;2:349–57.
Williams S. The association of the Glu298Asp polymorphism of endothelial nitric oxide synthase (eNOS) with outcome after subarachnoid hemorrhage. Doris Duke Medical Students J. 2003–2004;3:52–6.
Colombo MG, Andreassi MG, Paradossi U, Botto N, Manfredi S, Masetti S, et al. Evidence for association of a common variant of the endothelial nitric oxide synthase gene (Glu298!Asp polymorphism) to the presence, extent, and severity of coronary artery disease. Heart. 2002;87:525–8.
Hingorani AD. Polymorphisms in endothelial nitric oxide synthase and atherogenesis: John French lecture 2000. Atherosclerosis. 2001;154:521–7.
Tanus-Santos JE, Desai M, Flockhart DA. Effects of ethnicity on the distribution of clinically relevant endothelial nitric oxide variants. Pharmacogenetics. 2001;11:719–25
Persu A, Stoenoiu MS, Messiaen T, Davila S, Robino C, El-Khattabi O, et al. Modifier effect of eNOS in autosomal dominant polycystic kidney disease. Hum Mol Genet. 2002;11:229–41.
Tesauro M, Thompson WC, Rogliani P, Qi L, Chaudhary PP, Moss J. Intracellular processing of endothelial nitric oxide synthase isoforms associated with differences in severity of cardiopulmonary diseases: cleavage of proteins with aspartate vs. glutamate at position 298. Proc Natl Acad Sci U S A. 2000;6:2832–5.
Fairchild TA, Fulton D, Fontana JT, Gratton JP, McCabe TJ, Sessa WC. Acidic hydrolysis as a mechanism for the cleavage of the Glu (298)/asp variant of human endothelial nitric-oxide synthase. J Biol Chem. 2001;276:26674–9.
McDonald DM, Alp NJ, Channon KM. Functional comparison of the endothelial nitric oxide synthase Glu298Asp polymorphic variants in human endothelial cells. Pharmacogenetics. 2004;14:831–9.
Joshi MS, Mineo C, Shaul PW, Bauer JA. Biochemical consequences of the NOS3 Glu298Asp variation in human endothelium: altered caveolar localization and impaired response to shear. FASEB J. 2007;21:2655–63.
Miller SA, Dykes DD, Polesky HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988;16:1215.
Hingorani AD, Liang CF, Fatibene J, Lyon A, Monteith S, Parsons A, et al. A common variant of the endothelial nitric oxide synthase (Glu298→asp) is major risk factor for coronary artery disease in the UK. Circulation. 1999;100:1515–20.
Hingorani AD. Endothelial nitric oxide synthase polymorphisms and hypertension. Curr Hypertens Rep. 2003;5:19–25.
Huang PL, Huang Z, Mashimo H, Bloch KD, Moskowitz MA, Bevan JA, et al. Hypertension in mice lacking the gene for endothelial nitric oxide synthase. Nature. 1995;377:239–42.
Kojda G, Laursen JB, Ramasamy S, Kent JD, Kurz S, Burchfield J, et al. Protein expression, vascular reactivity and soluble guanylate cyclase activity in mice lacking the endothelial cell nitric oxide synthase: contributions of NOS isoforms to blood pressure and heart rate control. Cardiovasc Res. 1999;42:206–13.
Bode-Boger SM, Boger RH, Kielstein JT, Loffler M, Schaffer J, Frolich JC. Role of endogenous nitric oxide in circadian blood pressure regulation in healthy humans and in patients with hypertension or atherosclerosis. J Investig Med. 2000;48:125–32.
Cecilia Vecoli. Endothelial Nitric Oxide Synthase Gene Polymorphisms in Cardiovascular Disease. Vitamins and Hormones, Vitam Horm. 2014;96:387-406. https://doi.org/10.1016/B978-0-12-800254-4.00015-5.
Zhao Q, Su SY, Chen SF, Li B, Gu DF. Association study of the endothelial nitric oxide synthase gene polymorphisms with essential hypertension in northern Han Chinese. Chinese Med J. 2006;119:1065e1071.
Kishimoto T, Misawa Y, Kaetu A, Nagai M, Osaki Y, Okamoto M, et al. eNOS Glu298Asp polymorphism and hypertension in a cohort study in Japanese. Prev Med. 2004;39:927e931.
Shoji M, Tsutaya S, Saito R, Takamatu H, Yasujima M. Positive association of endothelial nitric oxide synthase gene polymorphism with hypertension in northern Japan. Life Sci. 2000;66:2557e2562.
Kato N, Sugiyama T, Morita H, Nabika T, Kurihara H, Yamori Y, et al. Lack of evidence for association between the endothelial nitric oxide synthase gene and hypertension. Hypertension. 1999;33:933e936.
Lacolley P, Gautier S, Poirier O, Pannier B, Cambien F, Benetos A. Nitric oxide synthase gene polymorphisms, blood pressure and aortic stiffness in normotensive and hypertensive subjects. J Hypertens. 1998;16:31e35.
Miyamoto Y, Saito Y, Kajiyama N, Yoshimura M, Shimasaki Y, Nakayama M, et al. Endothelial nitric oxide synthase gene is positively associated with essential hypertension. Hypertension. 1998;32:3e8.
Chen W, Srinivasan SR, Li S, Boerwinkle E, Berenson GS. Genderspecific influence of NO synthase gene on blood pressure since childhood: the Bogalusa heart study. Hypertension. 2004;44:668e673.
Benjafield AV, Morris BJ. Association analyses of endothelial nitric oxide synthase gene polymorphisms in essential hypertension. Am J Hypertens. 2000;13:994–8.
Moe KT, Lim ST, Wong P, Chua T, DeSilva DA, Koh TH, Wong MC, Chin-Dusting J. Association analysis of endothelial nitric oxide synthase gene polymorphism with primary hypertension in a Singapore population. J Hum Hypertens. 2006;20:956–63.
Cosentino F, Patton S, d’Uscio LV, Werner ER, Werner-Felmayer G, Moreau P, et al. Tetrahydrobiopterin alters superoxide and nitric oxide release in prehypertensive rats. J Clin Invest. 1998;101:1530e1537.
Vallance P, Collier J, Moncada S. Effects of endothelium-derived nitric oxide on peripheral arteriolar tone in man. Lancet. 1989;2:997–1000.
Duplain H, Burcelin R, Sartori C, Cook S, Egli M, Lepori M, et al. Insulin resistance, hyperlipidemia, and hypertension in mice lacking endothelial nitric oxide synthase. Circulation. 2001;104:342–5.
Forte P, Copland M, Smith LM, Milne E, Sutherland J, Benjamin N. Basal nitric oxide synthesis in essential hypertension. Lancet. 1997;349:837–42.
Node K, Kitakaze M, Yoshikawa H, Kosaka H, Hori M. Reduced plasma concentrations of nitrogen oxide in individuals with essential hypertension. Hypertension. 1997;30:405–8.
Moncada S, Higgs A. The L-arginine-nitric oxide pathway. N Engl J Med. 1993;329:2002–12.
Shimasaki Y, Yasue H, Yoshimura M, Nakayama M, Kugiyama K, Ogawa H, et al. Association of the missense Glu298Asp variant of the endothelial nitric oxide synthase gene with myocardial infarction. J Am Coll Cardiol. 1998;31:1506–10.
Tang W, Yang Y, Wang B, Xiao C. Association between a G894T polymorphism of eNOS gene and Essential hypertension in Hani and Yi minority groups of China. Arch Med Res. 2008;39:222e225.
Snieder H, Harshfield GA, Treiber FA. Heritability of blood pressure and hemodynamics in African- and European-American youth. Hypertension. 2003;41:1196–201.
Mein CA, Caulfield MJ, Dobson RJ, Munroe PB. Genetics of essential hypertension. Hum Mol Genet. 2004; 13 (Spec No 1):R169–R175 (Epub 2004 Feb 5).
Zintzaras E, Kitsios G, Stefanidis I. Endothelial NO synthase gene polymorphisms and hypertension: a meta-analysis. Hypertension. 2006;48:700e710.
Chen W, Srinivasan SR, Berenson GS. Plasma renin activity and insulin resistance in African American and white children: the Bogalusa heart study. Am J Hypertens. 2001;14:212e217.
Forstermann U, Sessa WC. Nitric oxide synthases: Regulation and function. European Heart Journal. 2012;33:829–37.
Fischmann TO, Hruza A, Niu XD, Fossetta JD, Lunn CA, Dolphin E, et al. Structural characterization of nitric oxide synthase isoforms reveals striking active-site conservation. Nature Struct Biol. 1999;6:233e242.
Tesauro M, Thompson WC, Rogliani P, Qi L, Chaudhary PP, Moss J. Intracellular processing of endothelial nitric oxide synthase isoforms associated with differences in severity of cardiopulmonary diseases: cleavage of proteins with aspartate vs. glutamate at position 298. Proc Natl Acad Sci U S A. 2000;97:2832e2835.
Jáchymová M, Horký K, Bultas J, Kozich V, Jindra A, Peleska J, Martásek P. Association of the Glu298Asp polymorphism in the endothelial nitric oxide synthase gene with essential hypertension resistant to conventional therapy. Biochem Biophys Res Commun. 2001;284:426–30.
Arnett DK, Claas SA, Glasser SP. Pharmacogenetics of antihypertensive treatment. Vasc Pharmacol. 2006;44:107–18.
Cooke GE, Doshi A, Binkley PF. Endothelial nitric oxide synthase gene: prospects for treatment of heart disease. Pharmacogenomics. 2007;8:1723–34.
Nagib El-Kilany GE, Nayel E, Hazzaa S. Nitric oxide synthase gene G298 allele. Is it a marker for microvascular angina in hypertensive patients? Cardiovasc Radiat Med. 2004;5:113–8.
Krex D, Fortun S, Kuhlisch E, et al. The role of endothelial nitric oxide synthase (eNOS) genetic variants in European patients with intracranial aneurysms. J Cereb Blood Flow Metab. 2006;26:1250–5.
Afrasyap L, Ozturk G. NO level and endothelial NO synthase gene polymorphism (Glu298Asp) in the patients with coronary artery disease from the Turkish population. Acta Biochim Biophys Sin (Shanghai). 2004;36:661–6.
Moon J, Yoon S, Kim E, et al. Lack of evidence for contribution of Glu298Asp (G894T) polymorphism of endothelial nitric oxide synthase gene to plasma nitric oxide levels. Thromb Res. 2002;107:129–34.
Walch K. Kolbus A. Hefler-Frischmuth K. Polymorphisms of the endothelial nitric oxide synthase gene in premenopausal women with polycystic ovary syndrome. Maturitas. 2008;61:256–9.
Li R, Lyn D, Lapu-Bula R, et al. Relation of endothelial nitric oxide synthase gene to plasma nitric oxide level, endothelial function, and blood pressure in African Americans. Am J Hypertens. 2004;17:560–7.
Hillermann R, Carelse K, Gebhardt GS. The Glu298Asp variant of the endothelial nitric oxide synthase gene is associated with an increased risk for abruptio placentae in pre-eclampsia. J Hum Genet. 2005;50:415–9.
Nassereddine S, Idrissi HH, Habbal R, Abouelfath R, Korch F, Haraka M, Karkar A, Nadifi S. The polymorphism G894 T of endothelial nitric oxide synthase (eNOS) gene is associated with susceptibility to essential hypertension (EH) in Morocco. BMC Med Genet. https://doi.org/10.1186/s12881-018-0638-1.
Acknowledgements
We thank gratefully all the staff of our Human Genetics and Molecular Pathologies Laboratory (LGPM), University Hassan II, School of Medicine, Casablanca, for their support and encouragements, and the Hassan II Academy of Science and Technology for its financial support.
Availability of data and materials
The datasets used and/or analysed during the current study is available from the corresponding author on reasonable request.
Author’s contributions
SN, HHI and RhA carried out the molecular genetics study and performed the statistical analysis and interpretation of results. FK and MH were involved in collection of data and blood samples from the Department of Cardiology, University Hospital Center Ibn Rochd, Casablanca, Morocco. RH and SN are the directors of the study; they critically revised the manuscript, provided important intellectual contribution and gave final approval of the version to be published. All authors read and approved the final version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Patients’ Clinical data were collected and an informed consent, approved by the Ethical Committee of the University of Hassan II, School of Medicine, Casablanca, was signed by each patient and control before entering the study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Nassereddine, S., Hassani Idrissi, H., Habbal, R. et al. The polymorphism G894 T of endothelial nitric oxide synthase (eNOS) gene is associated with susceptibility to essential hypertension (EH) in Morocco. BMC Med Genet 19, 127 (2018). https://doi.org/10.1186/s12881-018-0638-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12881-018-0638-1