
Abstract
Background
Peroxiredoxins (Prxs) are a large family of antioxidant enzymes that respond to biotic and abiotic stress by decomposing reactive oxygen species (ROS). In this study, the stress tolerance function of the Th2CysPrx gene was further analysed. It lays a foundation for further studies on the salt tolerance molecular mechanism of T. hispida and improved salt tolerance via transgenic plants.
Results
In this study, the stress tolerance function of the Th2CysPrx gene was further analysed. The results of transgenic tobacco showed higher seed germination rates, root lengths, and fresh weight under salt stress than wild-type tobacco. Simultaneously, physiological indicators of transgenic tobacco and T. hispida showed that Th2CysPrx improved the activities of antioxidant enzymes and enhanced ROS removal ability to decrease cellular damage under salt stress. Moreover, Th2CysPrx improved the expression levels of four antioxidant genes (ThGSTZ1, ThGPX, ThSOD and ThPOD).
Conclusions
Overall, these results suggested that Th2CysPrx enhanced the salt tolerance of the transgenic plants. These findings lay a foundation for further studies on the salt tolerance molecular mechanism of T. hispida and improved salt tolerance via transgenic plants.
Similar content being viewed by others
Background
Abiotic stresses, such as drought, salinity, and extreme temperature, among others, negatively affect the growth and yield of plants, resulting in very large economic losses. Among the harmful environmental stresses, salt stress in particular leads to slower plant growth and declines in cultivated plant production [1]. In the long-term evolutionary process, some plants, such as Atriplex canescens [2], Halostachys caspica [3], Suaeda salsa [4], and Salicornia brachiata [5], among others, have gradually adapted to a salt stress environment and can grow in dry and saline land.
Tamarix hispida is a typical woody halophyte that can form natural forests in saline alkali soil with 1% salt content. In addition, it can also endure drought stress, which makes it an ideal material to clone genes related to drought and salt tolerance and to study the salt tolerance mechanism of woody halophytes [6]. Many previous studies have been conducted to examine the mechanism of salt and drought tolerance and the function of stress resistance genes of T. hispida. For example, TheIF1A, ThDREB, ThZFP1, ThGSTZ1 and ThPOD3 in T. hispida increase salt and drought tolerance by regulating superoxide dismutase (SOD) and peroxidase (POD) activities to reduce reactive oxygen species (ROS) accumulation [7,8,9,10,11]. These results all suggest that scavenging of ROS plays an important role in the response of T. hispida to salt stress.
POD, SOD, CAT (Catalase), GPX (Glutathione peroxidase) and GST (Glutathione S-transferases) are important ROS-scavenging genes. Gao et al. [12] found that ThGSTZ1 improved tolerance to abscisic acid ABA and methyl viologen (MV) stress by augmenting the activities and expression levels of ThGPX, ThSOD, and ThPOD and the ROS-scavenging capacity. The NtSOD, NtAPX, NtCAT, NtPOX, and NtGST genes play roles in eliminating ROS and increasing stress tolerance in plant under stress. Fortunately, overexpressed wheat TaWRKY44 was found to activate the above five genes [13]. In Betula platyphylla, BplMYB46 could increase the ROS-scavenging capacity and proline content to improve salt and osmotic tolerance by affecting the expression of genes, including POD, SOD and P5CS (Δ1-pyrroline-5-carboxylate synthetase) [14]. Zhang et al. [15] found that under NaCl stress, ALA in tomatoes increased the activity of ROS-scavenging antioxidant enzymes and the expression of the SOD, APX and POD genes encoding these enzymes.
Peroxiredoxins (Prxs) are a family of non-haem peroxidases that are widely present in animals, plants and microorganisms. The biological function of these peroxidases is to regulate the balance of ROS and intracellular signal transduction through hydrogen peroxide (H2O2), alkyl hydroperoxides, and peroxynitrite [16,17,18,19]. In plants, based on the number and position of conserved Cys residues, Prxs are grouped into four classes: 1CysPrx, 2CysPrx, type-II Prx and PrxQ. The 1CysPrxs have only one conserved Cys residue, and PrxQ contains two cysteine residues that are catalytically active and linked by intramolecular disulphide bonds [20]. Both 2-Cys Prx and type-II Prx have two conserved Cys residues, but the difference between them is that 2-Cys Prx is a stromal protein [21], and Prx-II has various isoforms [22].
Prxs operate in a particular way during plant growth, development and stress tolerance. For example, Kim et al. [23] found that 2CysPrxs could eliminate H2O2 by participating in an alternative water-water cycle and protect photosynthetic structures against oxidative damage under environmental restriction. Pea chloroplast 2CysPrx and mitochondrial Prx IIF affect the structure and peroxidase activity of photosynthetic structures [24]. Kim et al. [25] found that 2CysPrx from Oryza sativa could increase tolerance to ROS-induced oxidative stress by improving cellular redox homeostasis. Overexpression of 2CysPrx in tall fescue plants increases resistance to oxidative stress and antioxidant activity [26]. Prx also detoxifies ROS and modulates signalling responses [27]. Interestingly, there is no direct evidence that the Prxs gene of T. hispida is involved in ROS scavenging and the abiotic stress response. In a previous study, Gao et al. [28] cloned four Prxs genes from T. hispida. Real-time quantitative PCR (qRT-PCR) analysis indicated that these genes could respond to several abiotic stresses and ABA application. However, Th2CysPrx displayed a unique expression pattern under the studied stress conditions. In the present study, the role of Th2CysPrx in salt stress was further demonstrated, and elucidated the molecular mechanism of this gene under salt stress. It also provided potential application prospects for molecular breeding to improve salt tolerance.
Results
Overexpression of Th2CysPrx improves salt stress tolerance in transgenic tobacco
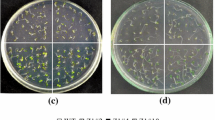
To study the biological role of Th2CysPrx in the salt stress response, the 14 resistant lines were obtained. The qRT-PCR results showed that Th2CysPrx gene were overexpressed in all 14 lines (Additional file 1: Figure S1). Two representative T3 homozygous transgenic lines (Line 7 and Line 11) were randomly selected for further salt tolerance analysis. The germination and growth of transgenic and wild-type (WT) tobacco were compared during exposure to normal and 125 mM NaCl stress conditions. The results showed that, under normal conditions, there was no obvious difference in the germination rates and seedling growth between transgenic and WT plants (Fig. 1a-c). Under salt stress, however, the germination rate and seedling growth of transgenic plants was significantly better than those of WT plants. Under NaCl stress, the germination rates of transgenic lines were 89.7% (Line 7) and 84.5% (Line 11), while that of WT plants was only 53.5% (Fig. 1d). The chlorophyll content of the transgenic lines under NaCl treatment was 1.54- and 1.68-fold greater, respectively, than that of the WT plants (Fig. 1e). The average fresh weight of the transgenic lines under NaCl treatment was 2.15- and 2.10-fold greater, respectively, than that of the WT plants (Fig. 1g). The average root length of transgenic lines under NaCl treatment was 1.99- and 1.87-fold greater, respectively, than that of WT plants (Fig. 1h). It was apparent that salt stress significantly inhibited the growth of transgenic and WT plants. However, the growth of transgenic plants was significantly superior to that of WT plants. These results indicated that the expression of Th2CysPrx in tobacco could significantly increase the salt tolerance of the plants.

Growth of Th2CysPrx transgenic lines and WT tobacco under NaCl stress. a The distribution chart of the WT, Line7 and 11 in the plates. b, c Seed germination of Th2CysPrx-transformed lines and WT plants under normal conditions (1/2 MS) and salt (125 mM NaCl) stress for 10 d. d Seed germination rates assay. e Chlorophyll contents assay. f The growth phenotype of Th2CysPrx transformed and WT plants. Analysis of fresh weight (g) and root length (h). Control: under normal conditions. All experiments were repeated three times. Data are the mean ± SD of three independent experiments. Asterisks (*, P < 0.05) indicate significant difference compared with WT plants
The Th2CysPrx gene significantly improves ROS scavenging and reduces cell damage in transgenic tobacco
The H2O2 and O2− concentration in the transgenic lines and WT plants were examined by 3,3′-diaminobenzidine (DAB) and nitro blue tetrazolium (NBT) staining, respectively. The results showed that under normal condition, there was no significant variation in the ROS generated by the three lines. In contrast, under salt stress, the WT plants exhibited deeper staining, suggesting much greater ROS accumulation in the WT than the transgenic lines under stress conditions (Fig. 2a). 2,7-dichlorofluorescin diacetate (H2DCF-DA) staining of the leaves of the transgenic and WT lines was also performed after 1 and 2 h of salt stress, respectively. There were no obvious differences in ROS levels in intact guard cells under normal conditions. However, after salt stress, the WT plants showed increased ROS accumulation in guard cells compared with the transgenic lines (Fig. 2b). The transgenic lines exhibited lower ROS content than the WT plants. These results indicated that Th2CysPrx led to a significant reduction of ROS accumulation in plant cells under salt stress.

Under 200 mM NaCl stress, histochemical staining analysis of ROS accumulation and cell damage of transgenic lines and WT plants in 2-month-old. a Diaminobenzidine (DAB) and nitroblue tetrazolium (NBT) revealed the accumulation of O2− and H2O2 in the leaves of transgenic (Line7 and 11) and (WT) plants subjected to salt stress, Evans Blue staining analysis of cell death. b H2DCF-DA staining of guard cells. All experiments were repeated three times
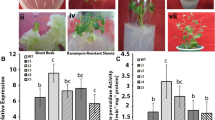
Furthermore, SOD and POD activities in the transgenic and WT plant lines were measured under salt stress. The results revealed no significant differences between two independent transgenic lines and WT plants under normal conditions. In contrast, SOD and POD activities were higher in the two transgenic lines than in the WT plants under NaCl stress. Specifically, the SOD activities of the transgenic lines were 1.17- and 1.10-fold higher than that of WT (Fig. 3a). And the POD activity values in the transgenic lines were 1.40- and 1.46-fold higher, respectively, than those in the WT plants (Fig. 3b).

Physiological index analysis of 2-month-old seedlings of 35S::Th2cys transgenic and WT plants under salt conditions. a Chlorophyll contents assay. b SOD activity. c POD activity. d MDA content. e Electrolyte leakages. Normally watered plants were used as controls. CK: Tobacco plants transformed with empty pROKII; 35::Th2Cys: Tobacco overexpressing Th2CysPrx; All experiments were repeated three times. Data are the mean ± SD of three independent experiments. Asterisks (*, P < 0.05) indicate significant difference compared with WT plants
Similarly, there were no obvious differences between the transgenic and WT plant lines based on Evans blue staining, malondialdehyde (MDA) contents and electrolyte leakage (EL) rates under normal conditions. Under salt stress conditions, deeper Evans blue staining was observed, and the MDA content and electrolyte leakage were significantly increased. However, Evans blue staining was weaker and the MDA content and EL rate were lower in the transgenic than in the WT plant lines. Under salt stress, the EL of the transgenic lines was 71.85 and 60% that of the WT plants, respectively (Fig. 3c). The MDA contents of the transgenic lines were 81.56 and 73.76% that of the WT plants, respectively (Fig. 3d).
Taken together, these results indicated that Th2CysPrx in tobacco improved salt stress tolerance by increasing ROS scavenging and preventing cell damage to maintain better plant growth. The transgenic lines exhibited lower ROS content and less damage than the WT plants, suggesting that Th2CysPrx directly affected ROS scavenging and cell protection during NaCl treatment.
Use of transient overexpression of Th2CysPrx in T. hispida to further evaluate the results in transgenic tobacco
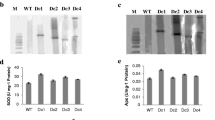
To confirm the results of heterologous expression in tobacco, Th2CysPrx was transiently transferred into T. hispida. qRT-PCR analysis showed that the expression of the Th2CysPrx gene in 35S::Th2cys overexpression T. hispida was 12.38-fold that of the control, indicating that the transient overexpression line of T. hispida was successfully obtained (Fig. 4a). Then, biochemical staining and related physiological indexes were analysed and compared between 35S::Th2cys and CK plants. The DAB and NBT staining results showed no significant difference between 35S::Th2cys and CK plants prior to salt stress. However, CK plants showed darker staining than 35S::Th2cys plants after salt stress, with darker staining after 2 h than 1 h (Fig. 4b).

Physiological index analysis of 1-month-old seedlings of CK, 35S::Th2cys under 150 mM NaCl stress. a The relative expression levels of Th2CysPrx as measured by qRT-PCR. The data was processed using the 2-ΔΔCT method. b DAB and NBT staining. c SOD activity levels. d POD activity levels. e MDA content. f Electrolyte leakages. CK: T. hispida plants transformed with empty pROKII; 35::Th2cys: T. hispida overexpressing Th2CysPrx; All experiments were repeated three times. Data are the mean ± SD of three independent experiments
Similarly, the SOD and POD activity levels also showed no difference between 35S::Th2cys and CK plants under normal conditions. Under salt stress, SOD activity in CK plants was 80.69% of that in 35S::Th2cys plants (Fig. 4c), and POD activity in CK plants was 70.51% of that in 35S::Th2cys plants (Fig. 4d). Under normal conditions, there was no significant difference in terms of MDA or electrolyte leakage content between 35S::Th2cys and CK plants. However, 35S::Th2cys plants showed significantly lower MDA level and electrolyte leakage than CK plants, and the MDA content and electrolyte leakage in 35S::Th2cys plants was 74.85 and 67.70% of that in CK plants (Fig. 4e, f). Altogether, these results showed that Th2CysPrx conferred salt tolerance to the transgenic tobacco and T. hispida plants.
Additionally, the expression levels of four antioxidant genes (ThGSTZ1, ThGPX, ThSOD and ThPOD) in transiently transfected Th2CysPrx T. hispida were analysed by qRT-PCR. The results showed that these genes shared similar expression profiles with Th2CysPrx in transiently transfected T. hispida seedlings, all of which were upregulated after NaCl stress. The expression levels of these genes were 1.22, 1.55, 1.50, and 1.86-fold that of CK plants, respectively (Fig. 5). These results indicated that Th2CysPrx overexpression affected ThGSTZ1, ThGPX, ThSOD and ThPOD gene expression. Thus, these results indicated that the expression of Th2CysPrx altered the expression of other stress-related genes, suggesting that salt stress tolerance regulation involves a complex network.

The relative expression levels of ThGSTZ1, ThGPX, ThSOD, ThPOD in WT and 35::Th2CysPrx exposed to NaCl stress. a The relative expression levels of ThGSTZ1. b The relative expression levels of ThGPX. c The relative expression levels of ThSOD. d The relative expression levels of ThPOD. CK: T. hispida plants transformed with empty pROKII; 35::Th2cys: T. hispida overexpressing Th2CysPrx; The error bars were calculated from three independent replicates of the qRT-PCR
Discussion
Under adversity, plants usually accumulate ROS, which leads to damage to protein synthesis and stability to produce cellular macromolecules and membrane lipids and the generation of oxidative stress [29]. The 2CysPrx plays an important role in scavenging ROS and regulating signal transduction, acting as a molecular chaperone and DNA damage response. In previous studies, 2CysPrx served as a barrier for H2O2. Rapeseed 2CysPrx activates chloroplast fructose-1,6-bisphosphatase (FBPase) and participates in the Calvin cycle [30]. Kim et al. [31] Showed that Arabidopsis 2CysPrx can protect citrate synthase (CS) from heat-induced aggregation and function as a molecular chaperone. Banerjee et al. [32] found that overexpression of cyanobacterium Anabaena 2CysPrx could decrease intracellular ROS levels under abiotic stress to reduce cell damage. Overexpression of 2CysPrx from mungbean can efficiently eliminate cellular ROS to improve tolerance to MV stress in Arabidopsis [33]. However, in the process of preventing DNA damage, some organisms use the ROS pathway, and some plants protect the cells from oxidative damage through the non-ROS pathway. For example, 2CysPrx-induced tobacco gene silencing largely affects the regeneration of ascorbic acid and thus the biosynthesis of abscisic acid (ABA) [17]. Above researches showed the 2CysPrx has different functions, most of which are research on glycogen plants, but little exploration in woody halophytes. Therefore, we need to explore whether Th2CysPrx prevents cell damage through the ROS pathway in woody halophyte T.hispida.
Vidigal et al. [17] demonstrated that 2CysPrx was the key to H2O2 clearance and regulated ABA signalling downstream of ROS genes. Many studies have shown that ABA has important biological functions and is an important signalling molecule under abiotic stresses [34,35,36]. The expression of genes that participate in the ABA signalling pathway can increase plant abiotic tolerance [37, 38]. In this study, compared with WT plants, transgenic Th2CysPrx tobacco exhibited increased germination rates, root lengths and chlorophyll contents under salt stress (Fig. 1). Overexpression of Th2CysPrx resulted in better SOD and POD activities, and reduced ROS accumulation. Additionally, the expression levels of the ThGSTZ1, ThGPX, ThSOD and ThPOD genes were markedly upregulated in transgenic T. hispida under salt stress, which indicated that the Th2CysPrx gene could improve salt tolerance by increasing antioxidant enzyme and strengthening ROS scavenging activities, leading to reduced ROS accumulation.
We also found that the activities and expression levels of SOD and POD in CK and transgenic T. hispida were not consistent under normal condition. However, the overall trend between the activities and expression levels is consistent under NaCl stress. The results showed the studied ThSOD and ThPOD gene may be key effective gene among SOD and POD family genes responding to NaCl stress (Figs. 2, 3, 4, 5).
It remains unclear whether the Th2CysPrx gene improves plant ROS scavenging by activating the expression of ABA signalling pathway genes. So, in future studies, we will further analyze the salt tolerance mechanism of the Th2CysPrx gene in T.hispida, while comparing the differences in abiotic stress in plants between 2CysPrx in tobacco and T.hispida.
Conclusion
Prxs have an important place in plant growth and development, as well as in responses to stress. However, there is no direct evidence to demonstrate that the Prxs gene of T. hispida is involved in ROS scavenging and abiotic stress responses. In the present study, a Th2CysPrx was isolated from T. hispida, and transgenic tobacco and T. hispida showed advantages with respect to morphological, physiological, and biochemical traits under salt stress. Additionally, the expression levels of four antioxidant genes (ThGSTZ1, ThGPX, ThSOD and ThPOD) were significantly higher in overexpressed Th2CysPrx of T. hispida than in the control. Altogether, the results indicated that Th2CysPrx increased salt tolerance via increasing the expression and activity of antioxidant enzymes and improved ROS scavenging ability. Future studies should assess whether Th2CysPrx participates in the ABA signalling pathway.
Methods
Plant materials and growth conditions
Tobacco seeds (Longjiang 911) were obtained from the Tobacco Science Research Institute of Heilongjiang Province [39]. And kept in our laboratory. Tobacco seeds stored at 4 °C for 3–5 days were sterilized with 70% (w:v) ethanol for 1 min followed by 3% sodium hypochlorite for 5 min, rinsed eight times with sterile water, and sown onto plates containing half-strength Murashige and Skoog (1/2 MS) medium. They were then placed in a cabinet with a controlled environment (16 h light: 8 h dark) at 22 °C. One week later, the seedlings were transplanted to pots containing a mixture of perlite and soil (1:3 v/v) and grown in a greenhouse (14 h light: 10 h dark) at 24 °C with 70% relative humidity.
T. hispida seeds (the Turpan Desert Botanical Garden, Xinjiang, 293 China) was planted in a greenhouse in Harbin (China). Seeds for propagation of plant material were harvested from these T. hispida plants, and planted on 1/2 MS medium and grown in a greenhouse (14 h light: 10 h dark) at 24 °C with 70% relative humidity and a photon flux density of 250 μE m− 2 s− 1.
Generation of transgenic plants
The ORF of Th2CysPrx was amplified and cloned into pROKII vector (referred to as 35S::Th2cys). The primers are shown in Table 1. The 35S::Th2cys was introduced into Agrobacterium tumefaciens EHA105 by electroporation, and the transgenic tobacco was further obtained by the agrobacterium-mediated leaf disc transformation method [40]. Fifteen lines were generated in the T0 generation. The two T3 homozygous transgenic lines (Line 7 and Line 11) were randomly selected for further analysis.
Concurrently, Th2CysPrx was also transiently overexpressed in T. hispida. Specifically, it was transiently transformed into one-month-old T. hispida seedlings with 35S::Th2cys to overexpress Th2CysPrx and with empty pROKII plasmid (used as a control, CK) according to Zheng et al. [41] with some modifications. In particular, single colonies of A. tumefaciens strain EHA105 harbouring 35S::Th2cys or empty pROKII were cultured to an OD600 = 0.8, and the cells were harvested by centrifugation. The transformants used [1/2 MS + 3% (w/v) sucrose + 150 μM acetosyringone + 0.01% (w/v) Tween 20] were adjusted to an OD600 = 0.8. T. hispida was immersed in transformation solution and incubated at 25 °C for 4 h. Subsequently, 2% sucrose was used to quickly wash the seedlings once for 1 min, after which they were planted vertically on 1/2 MS solid medium for 12, 24, 36 or 48 h. Total RNA was isolated from every samples using the CTAB method [42], and 1 mg of RNA was reverse transcribed with the PrimeScript™ RT reagent Kit (TaKaRa, China). The resulting cDNA product was diluted to 10x and used as a template for the qRT-PCR analyses. Real-time RT-PCR was conducted using a Bio-Rad (MJ) Opticon™2 System (Bio-Rad, Hercules, USA) Actin (FJ618517), α-tubulin (FJ618518), and β-tubulin (FJ618519) were used as internal controls to normalize the amount of total RNA present in each reaction. The primers used are listed in Table 1. The PCR conditions were 94 °C for 30 s, 45 cycles of 94 °C for 12 s, 58 °C for 30 s, 72 °C for 40 s, and 80 °C for 1 s for plate reading. A melting curve was generated for each sample at the end of each run to assess the purity of the amplified products. To determine the expression of Th2CysPrx and several antioxidant genes. The primers are listed in Table 1. The reaction system and procedure for qRT-PCR were performed according to Gao et al. [28]. The 2-ΔΔCt method was used to calculate the relative gene expression [43].
Analysis of stress tolerance
The T3 generation seeds of Th2CysPrx transgenic tobacco were surface sterilized and sown on 1/2 MS agar medium or 1/2 MS with 125 mM NaCl. Germination was recorded after 10 d. In addition, 3-day-old seedlings sown on 1/2 MS were transferred to 1/2 MS medium or 1/2 MS with 125 mM NaCl for 2 weeks to compare the fresh weight and root length between transgenic and wild type (WT) lines.
Physiological analysis
Seven-day-old tobacco seedlings sown on 1/2 MS were transferred to a mixture of perlite and soil (1:3 v/v). After 6 weeks, the tobacco seedling roots were watered with the solution of 200 mM NaCl. Simultaneously, the seedlings were watered with water as a control. After 7 d, the leaves of stressed and control tobacco were harvested.
One-month-old seedlings of the transient transgenic T. hispida seedlings (35S::Th2cys and CK) exposed to 150 mM NaCl for 12 h were harvested, and the physiological index was measured, respectively. The SOD, POD activities and MDA contents were measured according to Wang et al. [44]. The EL was measured according to Ben-Amor et al. [45]. The chlorophyll contents were measured following the method of Lichtenthaler [46].
Detection of ROS and cell death
The transgenic and WT/control young leaves were collected after 0, 1 or 2 h of NaCl treatment, and histochemical staining analysis was carried out immediately. To detect superoxide accumulation, hydrogen peroxide accumulation, and cell death, leaves were infiltrated with DAB, NBT and Evans blue staining according to the method described in detail by Zhang et al. [47] and Kim et al. [48]. Evaluation of ROS production in intact guard cells was performed by staining with H2DCF-DA (Sigma-Aldrich) [49].
Statistical analysis
Each experiment was repeated at least three times independently. Error bars represent standard deviations. Differences were compared using Student’s t-test. P < 0.05 was considered significant, which is indicated by *.
Availability of data and materials
All data and materials generated or analyzed during this study are included in this article or are available from the corresponding author on reasonable request.
Change history
19 August 2020
An amendment to this paper has been published and can be accessed via the original article.
Abbreviations
- SOD:
-
Superoxide dismutase
- POD:
-
Peroxidase
- ROS:
-
Reactive oxygen species
- CAT :
-
Catalase
- GPX :
-
Glutathione peroxidase
- GST :
-
Glutathione S-transferases
- MV:
-
Methyl viologen
- P5CS :
-
Δ1-pyrroline-5-carboxylate synthetase
- Prxs:
-
Peroxiredoxins
- H2O2 :
-
Hydrogen peroxide
- qRT-PCR:
-
Real-time quantitative PCR
- WT:
-
Wild-type
- DAB:
-
3,3′-Diaminobenzidine
- NBT:
-
Nitro blue tetrazolium
- H2DCF-DA:
-
2,7-Dichlorofluorescin diacetate
- MDA:
-
Malondialdehyde
- EL:
-
Electrolyte leakage
- FBPase:
-
Fructose-1,6-bisphosphatase
- CS:
-
Citrate synthase
- ABA:
-
Abscisic acid
- 1/2 MS:
-
Half-strength murashige and skoog
References
Elsawy HIA, Mekawy AMM, Elhity MA, Abdel-Dayem SM, Abdelaziz MN, Assaha DVM, et al. Differential responses of two Egyptian barley (Hordeum vulgare L.) cultivars to salt stress. Plant Physiol Biochem. 2018;127:425–35.
Li JT, Yu G, Sun XH, Zhang XH, Liu JL, Pan HY. AcEBP1, an ErbB3-binding protein (EBP1) from halophyte Atriplex canescens, negatively regulates cell growth and stress responses in Arabidopsis. Plant Sci. 2016;248:64–74.
Zhang S, Zeng YL, Yi XY, Zhang YF. Selection of suitable reference genes for quantitative RT-PCR normalization in the halophyte Halostachys caspica under salt and drought stress. Sci Rep. 2016;6:30363.
Zhang X, Liu XX, Wu L, Yu GH, Wang XE, Ma HX. The SsDREB transcription factor from the succulent halophyte Suaeda salsa enhances abiotic stress tolerance in transgenic tobacco. In J Genomics. 2015;5:1–13.
Tiwari V, Chaturvedi AK, Mishra A, Jha B. Introgression of the SbASR-1 gene cloned from a halophyte Salicornia brachiata enhances salinity and drought endurance in transgenic groundnut (Arachis hypogaea) and acts as a transcription factor. Plos One. 2015;10(7):e0131567.
Zhang TQ, Zhao YL, Wang YC, Liu ZY, Gao CQ. Comprehensive analysis of MYB gene family and their expressions under abiotic stresses and hormone treatments in. Front Plant Sci. 2018;9:1303.
Yang GY, Yu LL, Wang YC, Wang C, Gao CQ. The translation initiation factor 1A (TheIF1A) from Tamarix hispida is regulated by a Dof transcription factor and increased abiotic stress tolerance. Front Plant Sci. 2017;8:513.
Yang GY, Yu LL, Zhang KM, Zhao YL, Guo YC, Gao CQ. A ThDREB gene from Tamarix hispida improved the salt and drought tolerance of transgenic tobacco and T. hispida. Plant Physiol Biochem. 2017;113:187–97.
Zang DD, Wang C, Ji XY, Wang YC. Tamarix hispida zinc finger protein ThZFP1 participates in salt and osmotic stress tolerance by increasing proline content and SOD and POD activities. Plant Sci. 2015;235:111–21.
Yang GY, Wang YC, Xia DA, Gao CQ, Wang C, Yang CP. Overexpression of a GST gene (ThGSTZ1) from Tamarix hispida, improves drought and salinity tolerance by enhancing the ability to scavenge reactive oxygen species. Plant Cell Tiss Org. 2014;117(1):99–112.
Guo XH, Jiang J, Wang BC, Li HY, Wang YC, Yang CP, et al. ThPOD3, a truncated polypeptide from Tamarix hispida, conferred drought tolerance in Escherichia coli. Mol Biol Rep. 2010;37(3):1183–90.
Gao CQ, Yang GY, Guo YC, Zhao YL, Yang CP. Overexpression of ThGSTZ1 from Tamarix hispida improves tolerance to exogenous ABA and methyl viologen. Trees. 2016;30:1935–44.
Wang XT, Zeng J, Li Y, Rong XL, Sun JT, Sun T, et al. Expression of TaWRKY44, a wheat WRKY gene, in transgenic tobacco confers multiple abiotic stress tolerances. Front Plant Sci. 2015;6:615.
Guo HY, Wang YC, Wang LQ, Hu P, Wang YM, Jia YY, et al. Expression of the MYB transcription factor gene, BplMYB46, affects abiotic stress tolerance and secondary cell wall deposition in Betula platyphylla. Plant Biotechnol J. 2017;15(1):107–21.
Zhang ZP, Miao MM, Wang CL. Effects of ALA on photosynthesis, antioxidant enzyme activity, and gene expression, and regulation of proline accumulation in tomato seedlings under NaCl stress. J Plant Growth Regul. 2015;34(3):637–50.
Wang L, Guo HY, Zhang N, Ma ZH, Jiang SG, Zhang DC. Molecular characterization and functional analysis of a peroxiredoxin 1 cDNA from golden pompano (Trachinotus ovatus). Dev Comp Immunol. 2015;51(2):261–70.
Vidigal P, Martin-Hernandez AM, Guiu-Aragonés C, Amâncio S, Carvalho L. Selective silencing of 2Cys and type-IIB Peroxiredoxins discloses their roles in cell redox state and stress signaling. Integr Plant Biol. 2015;57(6):591–601.
Liu XL, Xi QY, Yang L, Li HY. Molecular cloning and sequence analysis of Peroxiredoxin cDNA from American White Shrimp (Litopenaeus vannamei). Adv Mater Res. 2011;343–344:1255–64.
Tripathi BN, Bhatt I, Dietz KJ. Peroxiredoxins: a less studied component of hydrogen peroxide detoxification in photosynthetic organisms. Protoplasma. 2009;235(1–4):3–15.
Kong W, Shiota S, Shi YX, Nakayama H, Nakayama K. A novel peroxiredoxin of the plant Sedum lineare is a homologue of Escherichia coli bacterioferritin co-migratory protein (Bcp). Biochem J. 2000;351(Pt 1):107–14.
Baier M, Dietz KJ. The plant 2-Cys peroxiredoxin BAS1 is a nuclear-encoded chloroplast protein: its expressional regulation, phylogenetic origin, and implications for its specific physiological function in plants. Plant J. 1997;12(1):179–90.
Horling F, König J, Dietz KJ. Type II peroxiredoxin C a member of the peroxiredoxin family of Arabidopsis thaliana: its expression and activity in comparison with other peroxiredoxins. Plant Physiol Biochem. 2002;40(6–8):491–9.
Kim SY, Jang HH, Lee JR, Sung NR, Lee HB, Lee DH, et al. Oligomerization and chaperone activity of a plant 2-Cys peroxiredoxin in response to oxidative stress. Plant Sci. 2009;177(3):227–32.
Calderón A, Lázaro-Payo A, Iglesias-Baena I, Camejo D, Lázaro JJ, Sevilla F, et al. Glutathionylation of pea chloroplast 2-Cys Prx and mitochondrial Prx IIF affects their structure and peroxidase activity and sulfiredoxin deglutathionylates only the 2-Cys Prx. Front Plant Sci. 2017;8:118.
Kim IS, Kim YS, Yoon HS. Expression of salt-induced 2-Cys peroxiredoxin from Oryza sativa increases stress tolerance and fermentation capacity in genetically engineered yeast Saccharomyces cerevisiae. Appl Microbiol Biotechnol. 2013;97(8):3519–33.
Kim KH, Alam I, Lee KW, Sharmin SA, Kwak SS, Lee SY, et al. Enhanced tolerance of transgenic tall fescue plants overexpressing 2-Cys peroxiredoxin against methyl viologen and heat stresses. Biotechnol Lett. 2010;32(4):571–6.
Horling F, Lamkemeyer P, König J, Finkemeier I, Kandlbinder A, Baier M, et al. Divergent light-, ascorbate-, and oxidative stress-dependent regulation of expression of the peroxiredoxin gene family in Arabidopsis. Plant Physiol. 2003;131(1):317–25.
Gao CQ, Zhang KM, Yang GY, Wang YC. Expression analysis of four peroxiredoxin genes from Tamarix hispida in response to different abiotic stresses and exogenous abscisic acid (ABA). Int J Mol Sci. 2012;13(3):3751–64.
Wang FZ, Wang QB, Kwon SY, Kwak SS, Su WA. Enhanced drought tolerance of transgenic rice plants expressing a pea manganese superoxide dismutase. J Plant Physiol. 2005;162(4):465–72.
Caporaletti D, D'Alessio AC, Rodriguez-Suarez RJ, et al. Non-reductive modulation of chloroplast fructose-1,6-bisphosphatase by 2-Cys peroxiredoxin. Biochem Biophys Res Commun. 2007;355:722–7.
Kim SY, Jang HH, Lee JR, et al. Oligomerization and chaperone activity of a plant 2-Cys peroxiredoxin in response to oxidative stress. Plant Sci. 2009;177:227–32.
Banerjee M, Chakravarty D, Ballal A. Redox-dependent chaperone/ peroxidase function of 2-Cys-Prx from the cyanobacterium Anabaena PCC7120: role in oxidative stress tolerance. BMC Plant Biol. 2015;15:60.
Cho CW, Chung E, Heo JE. Molecular characterization of a 2-Cys peroxiredoxin induced by abiotic stress in mungbean. Plant Cell Tiss Org. 2012;108(3):473–84.
Saxena I, Srikanth S, Chen Z. Cross talk between H2O2 and interacting signal molecules under plant stress response. Front Plant Sci. 2016;7:570.
León J, Castillo MC, Coego A, Lozano-Juste J, Mir R. Diverse functional interactions between nitric oxide and abscisic acid in plant development and responses to stress. J Exp Bot. 2014;65(4):907–21.
Wang YL, Ma FW, Li MJ, Liang D, Zou J. Physiological responses of kiwifruit plants to exogenous ABA under drought conditions. Plant Growth Regul. 2011;64(1):63–74.
Li XY, Kong XG, Huang Q, Zhang Q, Ge H, Zhang L, et al. CARK1 phosphorylates subfamily III members of ABA receptors. J Exp Bot. 2019;70(2):519–28.
Li ZX, Liu C, Zhang Y, Wang BM, Ran QJ, Zhang JR. The bHLH family member ZmPTF1 regulates drought tolerance in maize by promoting root development and ABA synthesis. J Exp Bot. 2019;70(19):5471–86.
Chen RP, Qiu EJ, Song BG, Jiao QM, Sun YK, Huang ZR. Breeding and characteristics of a new flue-cured tobacco variety Longjiang 911. China Tobacco Science. 2002;04:23–7.
Bastaki NK, Cullis CA. Floral-dip transformation of flax (Linum usitatissimum) to generate transgenic progenies with a high transformation rate. J Vis Exp. 2014;94:52189.
Zheng L, Liu GF, Meng XN, Li YB, Wang YC. A versatile agrobacterium-mediated transient gene expression system for herbaceous plants and trees. Biochem Genet. 2012;50(9–10):761–9.
Gao CQ, Wang YC, Jiang B, Liu GF, Yu LL, Wei ZG. A novel vacuolar membrane H+-ATPase c subunit gene (ThVHAc1) from Tamarix hispida confers tolerance to several abiotic stresses in Saccharomyces cerevisiae. Mol Biol Rep. 2011;38(2):957–63.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−delta delta C(T)) method. Methods. 2001;25(4):402–8.
Wang YC, Gao CQ, Liang YN, Wang C, Yang CP, Liu GF. A novel bZIP gene from Tamarix hispida mediates physiological responses to salt stress in tobacco plants. Plant Physiol. 2010;167(3):222–30.
Ben-Amor M, Flores B, Latché A, Bouzayen M, Pech JC, Romojaro F. Inhibition of ethylene biosynthesis by antisense ACC oxidase RNA prevents chilling injury in Charentais cantaloupe melons. Plant Cell Environ. 1999;22(12):1579–86.
Lichtenthaler HK. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods Enzymol. 1987;148C(11–12):350–82.
Zhang X, Wang L, Meng H, Wen HT, Fan YL, Zhao J. Maize ABP9 enhances tolerance to multiple stresses in transgenic Arabidopsis by modulating ABA signaling and cellular levels of reactive oxygen species. Plant Mol Biol. 2011;75(4–5):365–78.
Kim M, Ahn JW, Jin UH, Choi D, Paek KH, Pai HS. Activation of the programmed cell death pathway by inhibition of proteasome function in plants. J Biol Chem. 2003;278(21):19406–15.
Hua DP, Wang C, He JN, Liao H, Duan Y, Zhu ZQ, et al. A plasma membrane receptor kinase, GHR1, mediates abscisic acid- and hydrogen peroxide-regulated stomatal movement in Arabidopsis. Plant Cell. 2012;24(6):2546–61.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China (No.31670679), The Province in Heilongjiang Outstanding Youth Science Fund (JC2017004) and Heilongjiang Touyan Innovation Team Program (Tree Genetics and Breeding Innovation Team). The funding bodies played no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Author information
Authors and Affiliations
Contributions
CG designed the research and revised the manuscript; YW, ZL, PW and BJ performed research; YW, XL, JW and WD mainly conducted data analysis and wrote the paper and all authors read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1 Figure S1
. The relative expression levels of Th2CysPrx gene in the WT and transgenic tobacco as measured by qRT-PCR. The data was processed using the 2-ΔΔCT method. WT: the wild type tobacco. Line1–14: different transgenic tobacco lines; All experiments were repeated three times. The error bars represent the standard deviation.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, Y., Liu, Z., Wang, P. et al. A 2-Cys peroxiredoxin gene from Tamarix hispida improved salt stress tolerance in plants. BMC Plant Biol 20, 360 (2020). https://doi.org/10.1186/s12870-020-02562-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-020-02562-6