
Abstract
Background
In plants, ERF genes participate in a variety of regulatory pathways, such as plant growth and biotic and/or abiotic stress responses. Although the genome of Chinese white pear (‘Dangshansuli’) has been released, knowledge regarding the ERF family in pear, such as gene functions, evolutionary history and expression patterns, remains limited.
Results
In our study, a total of 155 members of ERF families were identified in pear (Pyrus bretschneideri). The Ka and Ks values suggested that whole-genome duplication (WGD) and dispersed duplication have effectively contributed to the expansion of the pear ERF family. Gene structure and phylogeny analysis divided the PbrERF family into 12 groups, and their gene functions were predicted by comparative analysis. qRT-PCR was carried out to verify the relative expression levels of 7 genes in group III using wild and cultivated pear fruits at three key developmental stages. Wild samples had higher expression of these genes than cultivated samples, especially at the enlarged fruit stage. The transcriptome data of pear seedlings subjected to dehydration treatment further revealed that 4 of the 7 genes responded to drought conditions.
Conclusion
The AP2/ERF gene family is greatly expanded in pear. Comparative analysis revealed the probability of ERF genes performing functional roles in multiple pathways. Expression analysis at different stages of pear fruit development in wild and cultivated samples indicated that genes in group III might be involved in abiotic and/or biotic stresses. Further transcriptome data on seedlings subjected to drought treatment verified the potential role of ERF genes in stress response. These results will provide a valuable reference for understanding the function and evolution of the ERF family in higher plants.
Similar content being viewed by others


Background
The AP2/ERF superfamily, plant-specific transcription factors, is characterized by a large number of members with great functional divergence. Members contain one or several conserved AP2/ERF domains consisting of approximately 60 to 70 amino acids in the DNA-binding region. Based on the number of conserved domains, the superfamily can be classified into three types, the AP2, ERF, and RAV gene families [1]. In the AP2 family, each protein sequence contains two AP2/ERF domains. In the ERF family, only a single AP2/ERF domain occurs in each protein sequence, while members of the RAV family contain not only an AP2/ERF domain but also a B3 domain—a conserved DNA-binding domain present in other plant-specific transcription factors. Furthermore, the ERF family can be divided into two major subfamilies, CBF/DREB and ERF [2]. In the ERF subfamily, the conserved nucleotide sequence AGCCGCC of the GCC-box [3] is found in the promoter regions of pathogenesis-related (PR) genes, which regulate gene expression in disease resistance response pathways [4]. In the DREB family, the conserved binding sequence CCGAC typically binds to cis-acting elements and participates in responses to abiotic stress (cold and drought) and in the regulation of plant hormones such as ABA and ethylene [5, 6] via the regulation of gene expression. Previous studies have shown that AP2 genes might regulate plant organ growth and development pathways, such as flower development and the determination of seed size [7,8,9]. Genes in the RAV family were predicted to participate in the response to ethylene [10] and to biotic and/or abiotic stresses [11, 12].
The release of high-throughout sequencing data has enabled identification and analysis of gene families at the genome-wide level. To date, whole-genome identification and analysis of the ERF gene family have been performed in many species, such as castor bean [13], Arabidopsis [14], poplar [15], grape [16], rice [17], wheat [18], cucumber [19], and soybean [20]. Pear, one of the most important fruits, in the Rosaceae family, is widely distributed worldwide. However, knowledge of the ERF family in pear remains severely limited. The recent completion of pear genome sequencing and assembly [21] provides us an opportunity to identify and dissect the AP2/ERF family. This investigation will provide insights into the function and evolution of the AP2/ERF family in pear.
In our study, we used the genome sequence data of pear to explore the AP2/ERF family on the whole-genome level. A total of 191 unigenes were identified as candidate members of the AP2/ERF family, including 155 ERF unigenes, 26 AP2 unigenes, and nine RAV unigenes. Phylogenetics, gene structure, and predicted function were characterized for the AP2/ERF family, as well as gene expression patterns. These results build a solid foundation for the future exploration of ERF gene functions in pear.
Methods
Identification of AP2/ERF gene family
Based on the pear genome project (http://peargenome.njau.edu.cn/) [21], we downloaded all protein sequences to identify members of the AP2/ERF gene family. The Arabidopsis AP2/ERF genes were identified in a previous study [22], and their amino acid sequences were downloaded from the Plant Transcription Factor Database (PlantTFDB) (http://planttfdb.cbi.pku.edu.cn). We used two approaches to obtain members of the ERF family in Pyrus bretschneideri. First, a Hidden Markov Model search (HMMsearch) was performed using the HMM profile with the AP2 domain (PF00847). Second, a BLASTP alignment against all pear protein sequences was used to perform an extensive search for candidate AP2/ERF genes using ERF protein sequences from tobacco and Arabidopsis thaliana as queries. For all sequences searched using these two methods, we first removed the redundant sequences and incomplete sequences. Then, we used the SMART tool (http://smart.embl-heidelberg.de/) and the InterProScan tool (http://www.ebi.ac.uk/Tools/pfa/iprscan/) to detect the presence or absence of the AP2/ERF domain in the candidate protein sequences. Protein sequences with one or more AP2/ERF domains were identified and retained as putative members of the AP2/ERF family for subsequent analyses. The location information on each pear AP2/ERF gene was obtained from the pear genome database. Then, the data were displayed by plotting a graph using Circos software [23].
Phylogenetics, gene structure and motif analyses
An un-rooted phylogenetic tree was constructed using MEGA6.0 [24] with neighbor-joining (NJ) criteria and verified using the maximum likelihood (ML) method, and 1000 bootstrap replicates were performed based on multiple alignments of the full-length amino acid (AA) sequences of all AP2/ERF genes in pear and Arabidopsis using ClustalW [25]. Based on the alignments of CDS sequences with the corresponding full-length genomic sequences, the gene structures of the AP2/ERF family were displayed using an online website: Gene Structure Display Server (GSDS) (http://gsds.cbi.pku.edu.cn/). Moreover, conserved motifs were detected in pear AP2/ERF family members using the motif analysis tool MEME (http://meme-suite.org/tools/meme) with the default parameters except for two: motif site distribution, any number of repetitions; maximum number of motifs, 30.
Synteny analysis and calculation of Ka and Ks values
We used a method distinct from that used in the Genome Duplication Database (PGDD) (http://chibba.agtec.uga.edu/duplication/) [26] to perform the synteny analysis. First, to identify the candidate homologous gene pairs (E < 1e-5, top 5 matches), a BLASTP alignment was carried out across the whole genome. The potential homologous gene pairs identified were then loaded into the software MCScanX with the default parameters [27, 28] to identify syntenic chains. We also used MCScanX to further distinguish the WGD/segmental, dispersed proximal, and tandem duplication event types in the ERF gene family.
Furthermore, candidate homologous gene pairs identified from the same synteny block were used as the input for the software KaKs_Calculator 2.0 [29] to calculate the Ka and Ks values. The software parameters were set as follows: YN as the Method (−m) and Standard Code as the Genetic code table (−c). Then, we used a python script written in-house to obtain the Ka and Ks values of the identified syntenic genes.
Gene expression analyses
The RNA-Seq data on ‘Dangshansuli’ obtained from our previous study [21] and downloaded from the pear genome website (http://peargenome.njau.edu.cn) were used to analyze the expression of PbrAP2/ERF genes at six different developmental stages of pear fruit: S1 (15 DAF), S2 (36 DAF), S3 (80 DAF), S4 (110 DAF), S5 (145 DAF), and S6 (167 DAF). The RNA-Seq data from five different dehydration treatments of pear seedlings obtained in the previous study were also downloaded [30]. The heatmaps were plotted in R using the heatmap.2 function based on the logarithmically (log2) transformed reads per kilobase per million (RPKM) values of each AP2/ERF gene.
RNA extraction and cDNA synthesis
We collected six pear accessions including three wild accessions of P. pyrifolia, ‘Matanggengzi’ (‘MTGZ’), ‘Baitanggengzi’ (‘BTGZ’), and ‘Tiantanggengzi’ (‘TTGZ’), and three cultivated accessions, ‘Huanghuali’ (‘HH’), ‘Lipuxueli’ (‘LPXL’), and ‘Liuchengfengshan’ (‘LCFS’), at three fruit developmental stages (small fruit stage, 52 DAF; enlarged fruit stage, 94 DAF; mature fruit stage, 128 DAF) for qRT-PCR analysis. First, we mixed the pear fruit samples of the same developmental stages from wild or cultivated genotypes. Then, the Plant Total RNA Isolation Kit Plus (FOREGENE Co. Ltd.) was used to extract the total RNA from the mixed samples of pear fruit. In the process, we carried out an improved step that was proposed in our previous study [31]. To obtain higher-quality RNA from pear fruit at the late stage of development, with high water content, less water (40 μl) was used to elute the RNA from the filtration column. Then, the total RNA was adjusted to the same concentration, and based on the adjusted RNA, first-strand cDNA was synthesized using TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix (TransGen Biotech Co. Ltd.).
Quantitative real-time PCR analysis (qRT-PCR)
Seven pairs of the most reliable primers (Additional file 1) were designed to amplify the seven candidate gene sequences using online software from NCBI (National Center for Biotechnology Information) (https://www.ncbi.nlm.nih.gov/tools/primer-blast/). According to the method described in a previous study [31], the LightCycler 480 SYBR GREEN I Master (Roche) was used to perform the qRT-PCR analysis. A 20 μl mixed reaction system was constructed, each containing 100 ng of template cDNA, 0.5 μM of each primer and 10 μl of LightCycler 480 SYBR GREEN I Master. All reactions were carried out in 96-well plates with four replicates for each cDNA sample. We set the qRT-PCR conditions as follows: first 5 min at 95 °C for pre-incubation, 55 cycles at 95 °C for 3 s, 60 °C for 10 s, and 72 °C for 30 s, and then 3 min at 72 °C for extension. Finally, the step of fluorescence signal data collection was carried out at 60 °C. Pyrus Actin (accession No. AF386514) and Pyrus GAPDH were used as the internal control genes. The average threshold cycle (Ct) of each cDNA sample was calculated using the running results displayed on the computer. Meanwhile, the relative expression levels of seven genes were calculated using the 2-∆∆Ct method described in a previous study [32].
Results
Identification of AP2/ERF genes in pear genome
Members of the AP2/ERF family were searched for in the Chinese white pear (P. bretschneideri) genome using two strategies: Hidden Markov Model search (HMMsearch) using the HMM profile (PF00847) of the AP2 domain and BLASTP search using ERF proteins from tobacco and Arabidopsis thaliana as queries. A total of 240 sequences were matched as candidate AP2/ERF genes across the whole genome. Among these candidates, we removed 41 AP2/ERF genes located in unanchored scaffolds, as well as two (Pbr039133.1 and Pbr002083.1) containing one Amb_all domain and one X8 domain, respectively. Meanwhile, six genes were removed because of incomplete domains. Consequently, 191 non-redundant and complete AP2/ERF genes were surveyed in our study. Based on their domain structures, the AP2/ERF superfamily genes were classified into three families: ERF (one AP2 domain), AP2 (two AP2 domains), and RAV (one AP2 and one B3 domain). Of these 191 genes, 22 genes were assigned to the AP2 gene family and encode proteins containing two repeated AP2/ERF domains. Interestingly, although the gene Pbr038562.1 contains two AP2/ERF domains, it is more closely related to the ERF type (Fig. 1). Eight genes were assigned to the RAV gene family and were predicted to encode one AP2/ERF domain and one B3 domain. Meanwhile, 161 genes encode proteins containing only a single AP2/ERF domain. Of these 161 genes, 155 were assigned to the ERF family. Of the remaining six genes, Pbr033071.1 (Pbr2AP2–3), Pbr025458.1 (Pbr10AP2–14), Pbr023949.1 (Pbr1AP2–1), and Pbr022083.1 (Pbr1AP2–2) also encode a single AP2/ERF domain, but are distinct from the ERF family and instead clustered into the AP2 family. Therefore, these six genes were further analyzed as AP2 genes (Additional file 2). Similarly, the gene Pbr030666.1 (Pbr9RAV7) encodes an AP2/ERF domain but is more similar to the RAV family. Finally, the gene Pbr002042.1 (Pbr14solo-1) was assigned as a soloist, with low homology to other AP2/ERF genes, although it includes an AP2/ERF-like domain sequence. A previous study showed that more AP2/ERF genes were identified in apple, including 51 AP2 genes, six RAV genes, 195 ERF genes, and seven soloists [33], supporting great expansion of the AP2/ERF family in both pear and apple. To distinguish each family member, we named these genes according to classification of the family and the order of the chromosome locations (Additional file 2). For example, genes in the ERF family were named from Pbr1ERF1 to Pbr17ERF155. Furthermore, the location information of each gene was used to display the distribution of the PbrERF genes in the pear genome. The results showed that all 155 PbrERF genes were distributed on chromosomes 1 through 17 as shown in Fig. 2.

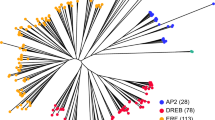
Phylogenetic tree of 191 AP2/ERF family proteins in pear. Red font represents the AP2 group; blue represents the RAV group; green represents the ERF group; black represents Soloist

Distribution and collinearity of the AP2/ERF members in the pear genome. Red lines mark the positions of genes on chromosomes. Blue lines indicate collinearity relationships among genes
Phylogenetics, gene structure, and conserved motif analyses
To explore the evolutionary relationships among the 191 AP2/ERF genes identified in pear, we constructed a phylogenetic tree using MEGA 6.0 based on multiple sequence alignments of 122 Arabidopsis ERF genes obtained from a previous study [7] and 191 pear AP2/ERF genes. The resulting phylogenetic tree allowed us to split the AP2/ERF family into 12 distinct clades from I to Xb-L, which were strongly supported by bootstrap values as shown in Fig. 3. Based on the classification of the AP2/ERF family in Arabidopsis thaliana [7], the DREB subfamily and the ERF subfamily were clearly separated, consisting of clades I-IV and clades V-Xb-L, respectively. In addition, the AP2 subfamily in pear contained 26 members; the clade RAV subfamily included nine members; and Soloist was separated (Table 1; Fig. 4a). Further, we found that most of the clades and subclades consisted of genes from both the pear and Arabidopsis AP2/ERF families, which indicated that the AP2/ERF genes are homologous and evolved from a common ancestor between pear and Arabidopsis.

Phylogenetic trees of pear and Arabidopsis AP2/ERF proteins. The full-length sequences of all AP2/ERF proteins were aligned using ClustalX, and the phylogenetic tree was generated using MEGA 6.0 with the NJ method

Analyses of gene structure and conserved motifs within each AP2/ERF clade in pear. a Clades identified in AP2/ERF family. b Exon/intron structures of pear AP2/ERF genes. Yellow boxes represent the exons. Blue boxes represent UTRs. Black lines connecting two exons indicate introns. c The distribution of conserved motifs within each AP2/ERF gene. Boxes in different colors represent different conserved motifs, and their relative positions are displayed
Gene structure analyses provided further evidence to support the phylogenetic topology groupings of gene families and showed that members of the AP2 family had intron numbers from six to ten, eight members in the RAV family had lost all introns, and Pbr9RAV7 had one. Most members of the ERF subfamily had only one exon and two UTR regions, but most members of groups V, VII, and X had two exons and one intron. Meanwhile, the Pbr14solo-1 gene contained seven introns and eight exons. Further, intron positions were found to be conserved in the AP2 family, although the number of introns varied. Likewise, most of the ERF family members shared the same or similar intron patterns, with most introns located in the AP2/ERF domain regions, as shown in Fig. 4b. In general, many conserved motifs could be detected in the protein sequences of transcription factors and may participate in activating gene expression as potential DNA-binding sites. In previous studies, diverse conserved motifs have been identified in rice and Arabidopsis, and their functions have been investigated [15, 22]. To characterize potential conserved motifs from the amino acid sequences of the AP2/ERF members, we used the Multiple Em for Motif Elicitation (MEME) tool [34] to analyze the 191 AP2/ERF amino acid sequences in pear. A total of 30 conserved motifs were detected and named 1–30 (see Fig. 4c). The results showed that most members of each group contained the same motifs. The gene structures provided reliable evidence to support and validate previous phylogenetic groupings.
Function prediction of each group in the PbrERF gene family
A previous study predicted the function of each group of the ERF family in Arabidopsis [22]. In our study, phylogenetic analysis allowed us to identify putative orthologous and paralogous ERF genes in pear and Arabidopsis. In general, homologous genes share similar gene structures and are clustered in the same clades, in which the genes possess similar functions. To predict the gene functions of each group in the pear ERF family, we constructed a phylogenetic tree via comparative genomics to define the ERF groups of orthologs from pear and Arabidopsis (Fig. 3). In group I, although the functions of all Arabidopsis ERF genes are unknown, the At1g78080 protein and the WXP1 protein share conserved motifs, and the overexpression of gene WXP1 has been reported as an activator in the regulation of wax synthesis in Medicago truncatula [35]. Therefore, we predicted that ERF genes of group I might play an important role in wax accumulation in pear as well. The functions of Arabidopsis group III ERF genes have been extensively studied and include potentially crucial functional roles in the response to cold, drought, and salt stresses [36,37,38]. Although the functions of group III protein in pear are unknown, these proteins may also participate in abiotic stress responses as transcriptional activators, based on the similar and conserved motif regions. In group IV, DREB2A (At5g05410) and DREB2B (At3g11020) have been reported to regulate DRE-mediated transcription as transcription factors [37], and the gene AtERF#052 (ABI4) participated in ABA signaling [39] and sugar response pathways [40, 41]. In group V, overexpression of the gene WIN1/SHN1 (At1g15360) contributed to the accumulation of leaf epidermal wax [42, 43], and genes SHN2 (At5g11190) and SHN3 (At5g25390) had similar functions. All genes of group V shared a conserved motif structure. Therefore, other pear ERF genes of group V may reasonably be hypothesized to play important roles in the pathway of wax accumulation as well. In addition, the fewer PbrERF genes in group V in pear than apple [33] (4 genes vs. 19 genes) might be due to its functional redundancy in controlling wax accumulation. The proteins in group VI share conserved motifs in the N-terminal region. In previous studies, the tobacco Tsi1 protein [44] and tomato Pti6 proteins [45] exhibited the similar characteristic gene structure features to members of group VI, and the genes Tsi1 and Pti6 have been reported to respond to abiotic and/or biotic stresses by regulating the expression of key genes in the pathway. In the group VI-L, all proteins also have the two conserved motifs that characterize group VI. In group VII, the gene AtEBP (At3g16770) was identified as a key gene interacting with the bZIP transcription factor OBF4 in vitro, although the function of this interaction remained unknown [46]. In addition, gene structure analyses showed that all genes in group VII have only a single intron and share completely consistent motifs in the 5′-flanking region of the AP2/ERF domain (Fig. 4). In group VIII, AtERF4 (At3g15210) and AtERF7 (At3g20310) were shown to negatively regulate gene expression in response to ABA, jasmonic acid, and ethylene [47,48,49]. Moreover, the genes LEP (At5g13910) [50] and ESR1/DRN (At1g12980) [51, 52] participate in the regulation of organ differentiation and plant development. Thus, we hypothesized that other pear genes in group VIII might have similar functions. In group X, Arabidopsis ABR1 (At5g64750) was reported in a previous study to participate in ABA response as a repressor, and knockout of the gene ABR1 resulted in an excitatory response to ABA in root growth and seed germination processes [53].
Gene duplication and synteny analyses in the PbrERF gene family
Five gene duplication types can be detected in a genome, including whole-genome duplication (WGD), singleton duplication, tandem duplication, proximal duplication, and dispersed duplication. These duplication events are the major driving force in the expansion of gene families [54]. Therefore, to explore the origins of duplicate genes in the PbrERF gene family, we dissected the duplication type of each member of the PbrERF family by using the software package MCScanX. Each gene in the PbrERF family was assigned to one of the five gene duplication types. Among them, 66.45% (103) of the pear PbrERF genes were retained from WGD or segmental duplication events, compared to only 5.16% (8) from tandem, 19.35% (30) from dispersed, and 9.03% (14) from proximal duplication events (Additional file 3).
To further provide additional evidence for a WGD event as the major force contributing to the expansion of the PbrERF gene family, a method developed from the one used in the Plant Genome Duplication Database (PGDD) was used to identify synteny blocks across the whole pear genome. In our analysis, a total of 75 duplicated gene pairs (Additional file 4) were found in the PbrERF gene family. The similarity for each pair ranged from 30.22%~ 100.00%. Meanwhile, considering that orthologs often retain equivalent functions over the course of evolution, we examined the orthologous relationships of ERF genes between pear and Arabidopsis using the same method. A total of 84 genes from pear, with the exception of Soloist, have one or several putative orthologs in Arabidopsis (Additional file 5). Meanwhile, all of them were subdivided into the same group as their orthologs in Arabidopsis, which further supported the results of the phylogenetic analysis. Among these 84 genes, eight belong to the AP2 family, 75 to the ERF family and only one to the RAV family. A previous study reported no Arabidopsis orthologs for the RAV family genes in Chinese plum [55]. Synteny analysis can be effectively used to provide strong support for putative paralogous or orthologous genes found through phylogenetic analyses.
Estimation of dates and driving forces of evolution
The synonymous substitution rate per site, that is, the Ks value, is usually used to estimate the evolutionary dates of WGD events. Previous studies have reported that the pear genome has undergone two WGD events: an ancient WGD (Ks ~ 1.5–1.8), which was inferred to have occurred ~ 140 MYA [56], and a recent WGD (Ks ~ 0.15–0.3), which was inferred to have occurred 30–45 MYA [21]. Therefore, we estimated the dates of the expansion of the PbrERF family by calculating the Ks value in our study. The mean Ks values of the PbrERF gene pairs duplicated in the WGD event in the syntenic region, ranging from 0.01 to 3.37, are shown in Additional file 4. Furthermore, 65 (86.67%) pairs of duplicated genes were distributed at the two Ks value peaks (Fig. 5). Thus, these duplicated gene pairs may have arisen from the same recent (30–45 MYA) and ancient (~ 140 MYA) WGDs, which led to the expansion of the PbrERF gene family.

Distribution of mean Ks values of 75 pairs of PbrMYB genes. The x-axis represents the mean Ks value; y-axis represents the density of the distribution. The data were plotted using R
Positive selection (Darwinian selection) drives the evolution of animals and plants via accumulation of new advantageous mutations, which are then spread throughout the population. In contrast, negative selection, that is, purifying selection, is an evolutionary process to remove deleterious mutations [57]. To further determine whether one or both selection processes have driven the evolution of the PbrERF family in pear, the Ka values (nonsynonymous substitutions per site) and Ka/Ks ratio of homologs were also calculated using coding sequences (CDS) of genes in the PbrERF family. The direction and magnitude of selection could be measured using the Ka/Ks ratio: values less than one indicate negative selection, equal to one indicates neutral selection, and greater than one indicates positive selection [58]. In these analyses, all 75 gene pairs possessed Ka/Ks ratios less than one except for the gene pair Pbr017391.1-Pbr030208.1, whose Ka/Ks ratio was greater than one, implying that purifying selection has driven PbrERF family evolution as the primary force. Meanwhile, we also proposed that the genes Pbr017391.1 and Pbr030208.1 might play important roles related to the evolution of plant phenotypic traits such as fruit size and sugar or acid content.
Expression analyses of AP2/ERF genes in pear
Transcriptome sequencing (RNA-Seq) data from six different developmental stages of ‘Dangshansuli’ pear fruit were downloaded from our pear genome database (http://peargenome.njau.edu.cn) [21]. The expression patterns of 191 pear AP2/ERF genes are shown in Fig. 6 (ERF family) and Fig. 7 (AP2 and RAV families, Soloist). Among these 191 genes, expression was not detected at any stage for 51 genes, and 140 genes were expressed in at least one pear fruit stage. Seventy-eight genes were detected at all six stages, although not all expression levels were high (Additional file 6).

Expression heatmap of pear ERF family genes at six developmental stages of fruit. The expression levels of AP2/ERF genes were measured by RNA-Seq analysis at six different stages: 15 DAF (S1), 36 DAF (S2), 80 DAF (S3), 110 DAF (S4), 145 DAF (S5), and 167 DAF (S6). The AP2/ERF gene family was subdivided into 12 groups: I, II, III, IV, V, VI, VI-L, VII, VIII, IX, X, and Xb-L. The color scale at the top right represents RPKM normalized by log2. Light green indicates a low expression level, gray indicates a medium level, and red indicates a high level. The graph was plotted in R using the heatmap.2 function

Expression heatmap of level of pear AP2, RAV, and Soloist genes. The expression levels of the AP2, RAV, and Soloist genes were measured by RNA-Seq analysis at six different stages: 15 DAF (S1), 36 DAF (S2), 80 DAF (S3), 110 DAF (S4), 145 DAF (S5), and 167 DAF (S6). The color scale at the top right represents RPKM normalized by log2. Light green indicates a low expression level, gray indicates a medium level, and red indicates a high level. The graph was plotted in R using the heatmap.2 function
The expression pattern of PbrAP2/ERF genes is varied, even within a single group. For example, group IX can be divided into three subgroups based on expression patterns. Five genes (Pbr4ERF25, Pbr7ERF61, Pbr1ERF2, Pbr12ERF101, and Pbr10ERF78) constitute a subgroup in which all genes were highly expressed in all six stages and are clustered together. Six genes (Pbr1ERF3, Pbr12ERF102, Pbr12ERF99, Pbr5ERF37, Pbr10ERF84, and Pbr1ERF1) constitute a second subgroup, in which all genes were expressed at low levels in each stage. However, the remaining genes can be clustered together and were expressed at low levels in one to four stages. Interestingly, in all groups, we could always detect one to six highly expressed genes at all fruit developmental stages.
Generally, wild plants have higher resistance than cultivated plants. To verify whether our functional cluster analysis was reliable, we selected seven genes of group III, predicted to be involved in abiotic and/or biotic stress response, for qRT-PCR analysis in wild and cultivated pears at three different developmental stages (small fruit stage, 52 Days After Flowering (DAF); enlarged fruit stage, 94 DAF; mature fruit stage, 128 DAF). The results indicated that these seven genes are significantly differently expressed in wild and cultivated pears, especially during the small and enlarged fruit stages (Fig. 8), and that the expression levels in wild pears are far higher than in cultivated pears. Accordingly, we could conclude that genes in group III regulated the abiotic stress response pathway via positive regulation and that resistance-related genes gradually lose their function during fruit ripening.

Relative expression levels of seven genes involved in the regulation of biotic and abiotic stresses. qRT-PCR analysis was performed to measure the expression levels of seven genes in subgroup III. Blue bars represent wild pear, and red bars represent cultivated pear. The x-axis represents the three stages, and the y-axis represent the relative expression levels of the genes, shown as the mean ± SD from three replications
We also used transcriptome data from dehydration treatment [30], in which pear seedlings were dehydrated for 0 (D0), 1 (D1), 3 (D3) and 6 (D6) hours at 26 °C, followed by recovery in water at 26 °C for 24 h (DH24), to validate these seven candidate stress-related genes. The results showed that four of the seven genes were differentially expressed in two or more comparisons from five libraries. As shown in Fig. 9, the gene Pbr4ERF24 was differentially expressed in nine comparisons, and its expression level peaked when the seedlings were dehydrated for six hours. The gene Pbr1ERF9 was differentially expressed in six comparisons, D0-VS-D3, D0-VS-D6, D1-VS-D6, D3-VS-D6, D3-VS-DH24, and D6-VS-DH24. Its gene expression peak also appeared at six hours of dehydration treatment. The gene Pbr2ERF12 was differentially expressed in six comparisons, D0-VS-D6, D0-VS-DH24, D1-VS-D6, D1-VS-DH24, D3-VS-D6, and D3-VS-DH24, while the gene Pbr7ERF63 was differentially expressed only in the D0-VS-D1 and D1-VS-DH24 comparison. These results strongly supported the involvement of ERF genes in the response to drought stress in pear.

The RPKM values of stress-related ERF genes in subgroup III. Transcriptome data from dehydration treatments were used to measure the expression levels of four genes. The x-axis represents the different comparisons, and the y-axis represents the RPKM values. Note: A-VS-B, blue bar represents A, red bar represents B
Discussion
As an important plant-specific transcription factor, the AP2/ERF gene family has been widely studied in many plants, such as Arabidopsis [14], poplar [15], grape [16], and rice [17]. However, relatively few pear ERF genes were investigated in previous studies. Herein, 191 members of the AP2/ERF superfamily were identified from pear genome, including 155 members from the ERF family. Previous studies have showed that similar numbers of members have been identified in plant species, a total of 202 members of AP2/ERF superfamily including 169 members of ERF family in poplar, and 180 members of AP2/ERF superfamily including 145 members of ERF family in rice. Meanwhile, these three species have the similar genome size, 527 Mb in pear, 480 Mb in poplar and 466 Mb in rice, indicating that the number of members in AP2/ERF superfamily were closely related to genome size in the three species. In the present study, some unanchored ERF genes and genes with incomplete domain were removed. Although this decreased the potential number of members in ERF family of pear, it made the candidate ERF genes more reliable.
Most of PbrERF genes were duplicated and retained from WGD event, and the extensive dispersed duplication was found in pear ERF family. A recent lineage-specific WGD event (30~ 45 MYA) [21] has likely contributed to the higher proportions of WGD-type PbrERF gene duplications observed in pear. Meanwhile, it also indicated that WGD events played major roles in the evolution and expansion of the PbrERF gene family in pear. Interestingly, previous analyses showed that recent gene duplication appears to be involved in the expansion of ERF family in apple [22], further supporting similar expansion patterns of the ERF family in both pear and apple. The calculation of Ks of ERF gene pairs further supported that most of ERF genes duplicated from the same recent and ancient WGD events. In addition, higher Ks values (1.92–3.37) were found for six (8.00%) duplicated gene pairs, and lower Ks values (0.004–0.01) were found for four (5.33%) gene pairs (Additional file 4), which suggested that other duplication events have occurred to drive the evolution of the ERF family in pear.
Phylogenetic tree of full-length amino acid sequences of ERF in pear and Arabidopsis revealed that most of subgroups in ERF family included both of genes from pear and Arabidopsis, indicating these ERF genes pre-date the species divergence. Genes in the same subgroups share the similar gene structures, which decide the similar gene functions. Therefore, comparison with known function of ERF genes in Arabidopsis could help to identify candidate orthologous genes of pear and predict their gene functions. As previous study has reported that most of subgroups in Arabidopsis were predicted involving in plant growth and stress responses [22], a plant without fruit. It is relatively limited to use Arabidopsis ERF genes as the queries in phylogenetic tree to identify the functional genes in pear fruit growth and development. However, RNA-Seq data provided us a complete expression profile of ERF genes in different fruit developmental stages, which showed at least one gene, in all subgroups, highly expressed at all stages. Gene expression can also provide us important clues to perform the gene function prediction [59]. The function prediction analysis indicated most of the AP2/ERF gene family members to be involved in abiotic and/or biotic stress response. Therefore, we predicted that the genes of each group that were highly expressed throughout growth and development might play more important roles in stress response. qRT-PCR analysis showed that seven genes in group III indeed differently expressed between wild and cultivated pears. Furthermore, transcriptome data from dehydration treatment [23] validated that four of seven PbrERF genes were differentially expressed in different treatments, while the remaining three genes might play more important roles in other stress responses.
Conclusion
This is the first comprehensive study on AP2/ERF gene family in pear aiming to help clarifying the gene function, evolution and expression pattern. The AP2/ERF gene family is greatly expanded in pear, and WGD event plays the important role. Function predication and expression divergence between duplicated genes revealed that the ERF genes are involved in multiple regulation pathways, multiple evidence supported that genes in group III might be involved in responses to abiotic stress. These results will lay a valuable foundation to understand the function and evolution of the ERF gene family in pear and other related species.
References
Riechmann JL, Heard J, Martin G, Reuber L, Jiang C, Keddie J, Adam L, Pineda O, Ratcliffe OJ, Samaha RR, et al. Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Science. 2000;290(5499):2105–10.
Yoh S, Qiang L, Joseph GD, Hiroshi A, Kazuo S, Kazuko Y-S. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Biochem Biophys Res Commun. 2002;290(3):998–1009.
Ohmetakagi M, Shinshi H. Ethylene-inducible DNA binding proteins that interact with an ethylene-responsive element. Plant Cell. 1995;7(2):173–82.
Hao D, Ohme-Takagi M, Sarai A. Unique mode of GCC box recognition by the DNA-binding domain of ethylene-responsive element-binding factor (ERF domain) in plant. J Biol Chem. 1998;273(41):26857–61.
Yamaguchi-Shinozaki K, Shinozaki K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell. 1994;6(2):251–64.
Chao J, Iu B, Singh J. Requirement of a CCGAC cis-acting element for cold induction of the BN115 gene from winter Brassica napus. Plant Mol Biol. 1996;30(3):679–84.
Elliott RC, Betzner AS, Huttner E, Oakes MP, Tucker WQ, Gerentes D, Perez P, Smyth DR. AINTEGUMENTA, an APETALA2-like gene of Arabidopsis with pleiotropic roles in ovule development and floral organ growth. Plant Cell. 1996;8(2):155–68.
Jofuku KD, Omidyar PK, Zorana G, Okamuro JK. Control of seed mass and seed yield by the floral homeotic gene APETALA2. Proc Natl Acad Sci U S A. 2005;102(8):3117–22.
Nole-Wilson S, Krizek BA. DNA binding properties of the Arabidopsis floral development protein AINTEGUMENTA. Nucleic Acids Res. 2000;28(21):4076–4082(4077).
Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Huaming C, Paul S, Stevenson DK, Justin Z, Pascual B, Rosa C. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science. 2003;301(5633):653–7.
Sohn KH, Lee SC, Jung HW, Hong JK, Hwang BK. Expression and functional roles of the pepper pathogen-induced transcription factor RAV1 in bacterial disease resistance, and drought and salt stress tolerance. Plant Mol Biol. 2006;61(6):897–915.
Chia-Wen L, Ruey-Chih S, Chiu-Ping C, Su-Juan Y, Tsai-Hung H, To-Chun C, Ming-Tsair C. Tomato RAV transcription factor is a pivotal modulator involved in the AP2/EREBP-mediated defense pathway. Plant Physiol. 2011;156(1):213–27.
Xu W, Li F, Ling L, Liu A. Genome-wide survey and expression profiles of the AP2/ERF family in castor bean (Ricinus communis L.). BMC Genomics. 2013;14(5):600–1.
Yoh S, Qiang L, Dubouzet JG, Hiroshi A, Kazuo S, Kazuko YS. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Biochem Biophys Res Commun. 2002;290(3):998–1009.
Jing Z, Cai B, Peng RH, Bo Z, Jin XF, Yong X, Feng G, Fu XY, Tian YS, Wei Z. Genome-wide analysis of the AP2/ERF gene family in Populus trichocarpa. Biochem Biophys Res Commun. 2008;371(3):468–74.
Licausi F, Giorgi FM, Zenoni S, Osti F, Pezzotti M, Perata P. Genomic and transcriptomic analysis of the AP2/ERF superfamily in Vitis vinifera. BMC Genomics. 2010;11(51):719.
Rashid M, Guangyuan H, Guangxiao Y, Hussain J, Xu Y. AP2/ERF transcription factor in Rice: genome-wide canvas and Syntenic relationships between monocots and eudicots. Evol Bioinformatics Online. 2012;8(4):321–55.
Zhuang J, Chen JM, Yao QH, Xiong F, Sun CC, Zhou XR, Zhang J, Xiong AS. Discovery and expression profile analysis of AP2/ERF family genes from Triticum aestivum. Mol Biol Rep. 2011;38(2):745–53.
Hu L, Liu S. Genome-wide identification and phylogenetic analysis of the ERF gene family in cucumbers. Gene Molecul Biol. 2011;34(4):624–33.
Zhang G, Ming C, Chen X, Xu Z, Shan G, Li LC, Li A, Guo J, Long M, Ma Y. Phylogeny, gene structures, and expression patterns of the ERF gene family in soybean (Glycine max L.). J Exp Bot. 2008;59(15):4095–107.
Wu J, Wang Z, Shi Z, Zhang S, Ming R, Zhu S, Khan MA, Tao S, Korban SS, Wang H. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 2013;23(2):396–408.
Toshitsugu N, Kaoru S, Tatsuhito F, Hideaki S. Genome-wide analysis of the ERF gene family in Arabidopsis and Rice. Plant Physiol. 2006;140(2):411–32.
Krzywinski M, Schein J, Birol I, Connors J, Gascoyne R, Horsman D, Jones SJ, Marra MA. Circos: an information aesthetic for comparative genomics. Genome Res. 2009;19(9):1639–45.
Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. [J] Molecul Biol Evol. 2016;33(7):1870.
Larkin MA, Blackshields G, Brown NP, et al. Clustal W. Clustal X version 2.0[J]. Bioinformatics. 2007;23(21):2947–8.
Lee T-H, Tang H, Wang X, Paterson AH. PGDD: a database of gene and genome duplication in plants. Nucleic Acids Res. 2013;41(D1):D1152–8.
Tang H, Wang X, Bowers JE, Ming R, Alam M, Paterson AH. Unraveling ancient hexaploidy through multiply-aligned angiosperm gene maps. Genome Res. 2008;18(12):1944–54.
Wang Y, Tang H, DeBarry JD, Tan X, Li J, Wang X, T-h L, Jin H, Marler B, Guo H. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012;40(7):e49.
Wang D, Zhang Y, Zhang Z, Zhu J, Yu J. KaKs_Calculator 2.0: a toolkit incorporating gamma-series methods and sliding window strategies. Genom, proteom bioinformat. 2010;8(1):77–80.
Li KQ, Xu XY, Huang XS. Identification of differentially expressed genes related to dehydration resistance in a highly drought-tolerant pear, Pyrus betulaefolia, as through RNA-Seq. PLoS One. 2016;11:e0149352.
Li X, Xue C, Li J, et al. Genome-wide identification, evolution and functional divergence of MYB transcription factors in Chinese white pear (Pyrus bretschneideri)[J]. Plant Cell Physiol. 2016;57(4):824–47.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method [J]. Methods. 2001;25(4):402–8.
Girardi CL, Rombaldi CV, Cero JD, et al. Genome-wide analysis of the AP2 / ERF, superfamily in apple and transcriptional evidence of ERF involvement in scab pathogenesis[J]. Sci Hortic. 2013;151(2):112–21.
Bailey TL, Williams N, Misleh C, Li WW. MEME: discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006;34(suppl 2):W369–73.
Zhang JY, Broeckling CD, Blancaflor EB, Sledge MK, Sumner LW, Wang ZY. Overexpression of WXP1, a putative Medicago truncatula AP2 domain-containing transcription factor gene, increases cuticular wax accumulation and enhances drought tolerance in transgenic alfalfa (Medicago sativa). Plant J. 2005;42(5):689–707.
Gilmour SJ, Zarka DG, Stockinger EJ, Salazar MP, Houghton JM, Thomashow MF. Low temperature regulation of the Arabidopsis CBF family of AP2 transcriptional activators as an early step in cold-induced COR gene expression. Plant J. 1998;16(4):433–42.
Liu Q, Kasuga M, Sakuma Y, Abe H, Miura S, Yamaguchi-Shinozaki K, Shinozaki K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell. 1998;10(8):1391–406.
Haake V, Cook D, Riechmann J, Pineda O, Thomashow MF, Zhang JZ. Transcription factor CBF4 is a regulator of drought adaptation in Arabidopsis. Plant Physiol. 2002;130(2):639–48.
Finkelstein RR, Wang ML, Lynch TJ, Rao S, Goodman HM. The Arabidopsis abscisic acid response locus ABI4 encodes an APETALA2 domain protein. Plant Cell. 1998;10(6):1043–54.
Huijser C, Kortstee A, Pego J, Weisbeek P, Wisman E, Smeekens S. The Arabidopsis SUCROSE UNCOUPLED-6 gene is identical to ABSCISIC ACID INSENSITIVE-4: involvement of abscisic acid in sugar responses. Plant J. 2000;23(5):577–85.
Arenas-Huertero F, Arroyo A, Zhou L, Sheen J, Leon P. Analysis of Arabidopsis glucose insensitive mutants, gin5 and gin6, reveals a central role of the plant hormone ABA in the regulation of plant vegetative development by sugar. Genes Dev. 2000;14(16):2085–96.
Aharoni A, Dixit S, Jetter R, Thoenes E, van Arkel G, Pereira A. The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell. 2004;16(9):2463–80.
Broun P, Poindexter P, Osborne E, Jiang C-Z, Riechmann JL. WIN1, a transcriptional activator of epidermal wax accumulation in Arabidopsis. Proc Natl Acad Sci U S A. 2004;101(13):4706–11.
Park JM, Park C-J, Lee S-B, Ham B-K, Shin R, Paek K-H. Overexpression of the tobacco Tsi1 gene encoding an EREBP/AP2–type transcription factor enhances resistance against pathogen attack and osmotic stress in tobacco. Plant Cell. 2001;13(5):1035–46.
Zhou J, Tang X, Martin GB. The Pto kinase conferring resistance to tomato bacterial speck disease interacts with proteins that bind a cis-element of pathogenesis-related genes. EMBO J. 1997;16(11):3207–18.
Büttner M, Singh KB. Arabidopsis thaliana ethylene-responsive element binding protein (AtEBP), an ethylene-inducible, GCC box DNA-binding protein interacts with an ocs element binding protein. Proc Natl Acad Sci. 1997;94(11):5961–6.
McGrath KC, Dombrecht B, Manners JM, Schenk PM, Edgar CI, Maclean DJ, Scheible W-R, Udvardi MK, Kazan K. Repressor-and activator-type ethylene response factors functioning in jasmonate signaling and disease resistance identified via a genome-wide screen of Arabidopsis transcription factor gene expression. Plant Physiol. 2005;139(2):949–59.
Yang Z, Tian L, Latoszek-Green M, Brown D, Wu K. Arabidopsis ERF4 is a transcriptional repressor capable of modulating ethylene and abscisic acid responses. Plant Mol Biol. 2005;58(4):585–96.
Song C-P, Agarwal M, Ohta M, Guo Y, Halfter U, Wang P, Zhu J-K. Role of an Arabidopsis AP2/EREBP-type transcriptional repressor in abscisic acid and drought stress responses. Plant Cell. 2005;17(8):2384–96.
van der Graaff E, Dulk-Ras A, Hooykaas P, Keller B. Activation tagging of the LEAFY PETIOLE gene affects leaf petiole development in Arabidopsis thaliana. Development. 2000;127(22):4971–80.
Banno H, Ikeda Y, Niu Q-W, Chua N-H. Overexpression of Arabidopsis ESR1 induces initiation of shoot regeneration. Plant Cell. 2001;13(12):2609–18.
Kirch T, Simon R, Grünewald M, Werr W. The DORNRÖSCHEN/ENHANCER OF SHOOT REGENERATION1 gene of Arabidopsis acts in the control of meristem cell fate and lateral organ development. Plant Cell. 2003;15(3):694–705.
Pandey GK, Grant JJ, Cheong YH, Kim BG, Li L, Luan S. ABR1, an APETALA2-domain transcription factor that functions as a repressor of ABA response in Arabidopsis. Plant Physiol. 2005;139(3):1185–93.
Maher C, Stein L, Ware D. Evolution of Arabidopsis microRNA families through duplication events. Genome Res. 2006;16(4):510–9.
Du D, Hao R, Cheng T, Pan H, Yang W, Wang J, Zhang Q. Genome-wide analysis of the AP2/ERF gene family in Prunus mume. Plant Mol Biol Report. 2013;31(3):741–50.
Fawcett JA, Maere S, Van de Peer Y. Plants with double genomes might have had a better chance to survive the cretaceous-tertiary extinction event. Proc Natl Acad Sci. 2009;106(14):5737–42.
Starr TK, Jameson SC, Hogquist KA. Positive and negative selection of T cells. Annu Rev Immunol. 2003;21(1):139–76.
Yang Z. PAML 4: phylogenetic analysis by maximum likelihood. Mol Biol Evol. 2007;24(8):1586–91.
Peng X, Zhao Y, Li X, Wu M, Chai W, Sheng L, Wang Y, Dong Q, Jiang H, Cheng B. Genomewide identification, classification and analysis of NAC type gene family in maize. J Genet. 2015;94(3):377–90.
Acknowledgments
We thank Dr. Jing Fan and Prof. Hongju Hu for providing the wild and cultivated sand pear materials to support our works. We also thank the group members in the Centre of Pear Engineering Technology Research, State Key Laboratory of Crop Genetics and Germplasm Enhancement at the Nanjing Agricultural University.
Funding
The work was funded by the National Science Foundation of China (31672111), the earmarked fund for the China Agriculture Research System (CARS-28), the Science Foundation of Jiangsu Province for Distinguished Young Scholars (BK20150025) and The Six Talent Peaks Project in Jiangsu Province (2014-NY-025).
Availability of data and materials
All sequence information on pear is available at the pear genome database (http://peargenome.njau.edu.cn/). All Arabidopsis protein sequences were downloaded from The Arabidopsis Information Resource (TAIR) (https://www.arabidopsis.org). Transcriptome sequencing (RNA-Seq) data on six different developmental stages of pear fruit have been published and can be downloaded from our website (http://peargenome.njau.edu.cn). All data generated or analyzed during this study are included in this article and its supplementary information files. Six fruit samples collected from wild and cultivated accessions at three different developmental stages were provided by the Research Institute of Fruit and Tea, Hubei Academy of Agricultural Science, Wuhan, China.
Author information
Authors and Affiliations
Contributions
XL carried out the experiments, data analysis, and figure preparation and drafted the manuscript. ST and MM performed the RNA extraction and qRT-PCR analysis. SW and SZ provided valuable suggestions and discussed this manuscript. And XH provided the transcriptome data from the dehydration treatment of pear seedlings. JW managed and designed the research.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
The seven pairs of primer sequences for amplifying functional genes in group III for qRT-PCR analysis (XLS 36 kb)
Additional file 2:
The gene name, gene ID, domain description, start position, end position, chromosome, and gene length of 191 pear AP2/ERF genes. (XLS 60 kb)
Additional file 3:
The duplication event types of 155 pear ERF genes. (XLS 54 kb)
Additional file 4:
The Ka, Ks, and Ka/Ks values of 75 pear ERF gene pairs. (XLS 42 kb)
Additional file 5:
The orthologous relationships between pear and Arabidopsis ERF genes. (XLS 49 kb)
Additional file 6:
The RPKM (reads per kilobase per million) values for PbrAP2/ERF gene expression. (XLS 98 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Li, X., Tao, S., Wei, S. et al. The mining and evolutionary investigation of AP2/ERF genes in pear (Pyrus). BMC Plant Biol 18, 46 (2018). https://doi.org/10.1186/s12870-018-1265-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-018-1265-x