
Abstract
Background
Lesion mimics (LMs) are disease-like symptoms that occur randomly on plant green leaves in the absence of pathogens. A previous study showed that LMs are related to enhanced resistance to a broad spectrum of diverse pathogen races and programmed cell death (PCD). Stripe rust is a globally epidemic fungal disease that can substantially reduce the quality and yield of crops. The development of resistant cultivars is an economical and environmentally friendly way to enhance the adaptability and yield stability of crops instead of the use of fungicide applications.
Results
In this study, a novel LM gene affording Pst resistance was identified and mapped with molecular markers developed for marker-assisted selection (MAS)-based wheat breeding. In this study, a novel LM gene named lm4, which is closely linked (8.06 cM) to SSR markers Xgwm210 and Xgwm455, was identified by using a Yanzhan 1/Neixiang 188 RIL population. The genetic distance of lm4 was then narrowed such that it was flanked by SSR markers with 0.51 cM and 0.77 cM intervals. Two SSR markers, lm4_01_cib and lm4_02_cib, were developed based on the content in the Chinese Spring genome database and wheat 660 K SNP results; these markers can be used to conduct MAS of LMs in wheat. The results also showed that lm4 significantly improved the resistance of stripe rust in wheat.
Conclusions
Therefore, lm4 is associated with stripe rust resistance, which may provide theoretical support for future crop disease-resistance breeding and for understanding the plant apoptosis mechanism.
Similar content being viewed by others

Background
Lesion mimics (LMs), which are also referred to as hypersensitive reaction-like (HRL) traits, occur spontaneously in leaf tissue without attack by any plant pathogens. LMs may provide enhanced plant resistance to a broad spectrum of diverse pathogen races [1, 2]. LMs exhibit different phenotypes, such as their color and size, with respect to the timing and conditions [3]. Previous studies have reported that LM traits exist in several plant species, such as maize [4, 5], Arabidopsis [6, 7], barley [8], and rice [9]. Studies of lesion mimics have provided insight into the activation of programmed cell death (PCD) or defense response pathways in plants [10]. Some LM mutants spontaneously express defense response genes involved in plant disease resistance signaling pathways [3, 11].
LM mutants have also been reported to be resistant to virulent pathogen races, which supports the direct use of LM mutants in crop disease-resistance breeding [12]. However, only a few studies concerning lesion mimics in wheat have been reported [2, 10]. Previous studies have reported that the C591 mutant (M8) is a stable flecking mutant [13]. Another leaf flecking mutant (M66) showed enhanced resistance to powdery mildew, stripe rust and brown rust [14,15,16]. Kamlofski et al. reported a hypersensitive-like (HPL) trait that was similar to the lesion-mimic phenotype and enhanced resistance to leaf rust [12]. A dominant gene (lm) from wheat cultivar Ning 7840 was located on chromosome 1BL and provided resistance to leaf rust in adult plants [2]. Two light-dependent lesion-mimic genes (lm1 and lm2) have been subsequently mapped onto 3BS and 4BL [17]. Wang et al. reported that a novel light-dependent lesion-mimic mutation (lm3) was closely linked to the SSR marker Xbarc203 on chromosome 3BL, and the resulting wheat mutants exhibited enhanced resistance to powdery mildew [18]. To date, just a few lesion-mimic-related genes have been characterized in bread wheat. Our knowledge of the effects of LMs on wheat disease resistance is limited, and the chromosome locations of the genes underlying the LM trait have not been determined [2]. Therefore, characterizing LM genes and elucidating their functions is of great significance to understand both the whole signal transduction pathway of programmed cell death and disease resistance mechanisms in crop plants.
Stripe rust is an and airborne fungal disease caused by Puccinia striiformis f. sp. tritici (Pst) and occurs worldwide [19]. Stripe rust can significantly reduce the quality and yield of crops [20]. The development of cultivars exhibiting durable tolerance to various pathogens is an economical and environmental way to enhance the adaptability and yield stability of crops instead of the use of fungicide applications [21,22,23,24]. The objectives of this study were to (1) identify LM genes in wheat and map them, (2) investigate the probable effects of the lm gene on Pst resistance and important agronomic traits, and (3) develop molecular markers that are useful in MAS and gene cloning in the future.
Results
Phenotypic and genetic analysis of lesion mimics in the RIL population
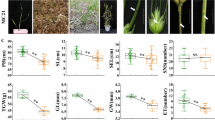
In this study, lesion-mimic (LM) traits likely appear as small yellow spots (disease-like symptoms) randomly spread throughout the green leaves of wheat (Fig. 1a, b, c). LMs appear without any plant pathogens, and LM spots started at approximately the fifth-leaf stage of wheat plants. In the current study, we found lesion-mimicking phenomena among Yanzhan 1/Neixiang 188 RILs. In the present study, the lesion-mimic trait was classified into scores of 0–4 based on the spread number and severity of yellow spots on the wheat leaves (Fig. 1d). According to their LM scores, all the RILs were then divided into two groups from 2015 to 2018: the normal-phenotype (LM0–1) group and the LM-phenotype (LM2–4) group. The segregation ratio of the two groups of LM traits in 2015–2018 was tested by the chi-square fitness test (Table 1). The segregation of the normal and LM phenotypes in the population conformed to a 1:1 ratio (p > 0.05) in all the environments. We crossed several LM4- and LM0-phenotype RILs, and the F1s showed lesion-mimic traits in their leaves (Fig. 1c). These results suggest that the lesion-mimic phenotype in Yanzhan 1/Neixiang 188 RILs is seemingly controlled by a single dominant gene. The dominant lesion-mimic gene identified in this study was named lm4.

a, b Lesion mimic phenotype and stripe rust on wheat leaves; c The phenotype of F1 crossed by LM4 and LM0 (LM4 and LM0 wheat lines were from Yanzhan1/Neixiang188 RILs); d Phenotype and classification of LM
Relation between lesion mimics and stripe rust resistance
In this study, a significant negative correlation was investigated between the lesion-mimic score and stripe rust IT value in 2016–2017 (r = − 0.53 ~ − 0.66, p < 0.01) (Table 2). As the degree of LM increased, the wheat stripe rust IT value significantly decreased. In breeding programs, plants with LMs are essentially highly resistant to stripe rust. Therefore, these results indirectly indicate that lm4 plays an important role in wheat stripe rust resistance, which can provide new insights or theoretical support for future disease-resistance breeding.
Lesion-mimic effects on agronomic traits
The effects of lesion mimics on the agronomic traits of wheat plants were investigated at Shifang and Maerkang in 2016–2018 in Sichuan Province (Table 3). Except for plant height (PH) in 2018 there were no significant effects of lesion mimics on spikelet number (SPI), number of sterile spikelets per spike (SSNS), grain number per spike (GNS), 1000-grain weight (TGW), or spike length (SL) of wheat. In general, these results reflect that LMs have no impact on agronomic traits of wheat, including yield traits.
Chromosomal location of the lesion-mimic gene
A total of 252 SSR markers were used to construct a genetic linkage map of 198 RILs (linkage map obtained from CAAS) in this study. lm4 was preliminarily localized to 2DS. However, the genetic distance was not close (8.06 cM), and the gene was flanked by SSR markers Xgwm210 and Xgwm455 (Fig. 2a, b); the LOD value was 30.1, and the phenotypic variation explained (PVE) was 50.8% (Fig. 2a). Therefore, a wheat 660 K SNP array was used to develop new molecular markers to narrow the physical genetic distance. Several SSR primers (markers) linked to lm4 on the 2DS chromosome were developed from the results of the wheat 660 K SNP array (Table S1, S2, Additional file 1). The genetic distance of lm4 was then narrowed such that the gene was flanked by SSR markers lm4_01_cib and lm4_02_cib (Fig. 2c); the genetic distances were 0.51 cM and 0.77 cM, respectively. The LOD value was 19.4, and the PVE was 37.1%. lm4 was ultimately delimited to an approximately 50 Mb regions on the basis of the Chinese Spring chromosome 2D genome sequence. The lm4 gene identified in this study is a novel lesion-mimic gene, which is different from the previously reported LM gene.

The genetic map of the region around lm4 on chromosome 2DS. LOD curves with data from different years (2015–2018 Shifang and Maerkang) (a), initial (b) and advanced (c) genetic linkage maps
Discussion
A novel lesion-mimic gene and mapping
Lesion mimics constitute a disease-like phenomenon that occurs in plant leaves without any pathogen infection, injury or obvious stress [25]. The phenotype of LMs in the current study is similar to that of yellow spot lesions on wheat leaves and lesion spots at booting in the fifth or sixth leaf stage of wheat. The previously reported LM gene in wheat, lm, was located on 1BL [2], lm1 and lm2 were located on 3BS and 4AL [17], and lm3 was mapped onto 3BL [18]. Unlike the previously reported lesion-mimic genes, the LM gene found in this study (lm4) is a novel type of lesion-mimic gene in wheat; this gene was mapped to 2DS and is a dominant LM gene derived from Yanzhan 1/Neixiang 188 RILs. The phenotype of the lesion mimic (lm4) in this study was also different from that of the HLP mutant induced by EMS [12]. Although LM traits are expressed at about the fifth or sixth leaf stage of wheat, LMs are the result of a natural mutation, and the types of lesion manifested also differed in this study: the LM phenotype induced by EMS involves small white spots (1 ~ 2 mm) on the leaves [12]. Therefore, LMs constitute a novel type of lesion-mimic trait that is different from that of the EMS-induced mutant, and lm4 is also different from previously reported LM genes.
Relationships between LMs and yield traits in wheat
Previous studies have shown that most lesion mimics have a negative effect on agronomic crop traits, especially those affecting yield production, although HLP mutants have been excluded [12, 16, 26]. In the current study, the agronomic traits, including yield traits (SPI, SSNS, GNS and TGW), of the wheat RILs were not significantly reduced by the appearance of a lesion-mimic phenotype compared with the phenotype of normal wheat lines (Table 3). Breeders aim to develop disease-resistant and high-yielding crop varieties. In this study, we found that lm4 significantly improved stripe rust resistance in wheat and did not affect major yield-related traits. Thus, this gene could be used as a potential tool for future disease-resistance breeding.
Effects of lesion mimics on stripe rust resistance
Stripe rust is a major fungal disease that threatens the quality and yield of wheat [27]. Controlling the spread of stripe rust and breeding new resistant varieties to improve the quality and yield of wheat is the main goal of breeders. In addition to disease resistance genes for specific races, studies on certain disease resistance-related genes have gradually attracted increased amounts of attention in recent years. Li et al. reported that lm (derived from Ning 7840 and located on 1BL) can enhance leaf rust resistance in wheat [2]. lm1 and lm2 were mapped to 3BS and 4AL, respectively, and are correlated with improved powdery mildew resistance [17]. The recently located LM gene lm3 (mapped onto 3BL) provides resistance to powdery mildew in adult plants [18]. These results provide new insight into the molecular mechanism of LM to improve broad-spectrum resistance in wheat, which may be helpful for screening candidate genes underlying the LM trait in this species. In this study, lm4 was found to be a novel lesion-mimic gene that is related to enhancing stripe rust resistance in wheat. Certain QTLs for stripe rust, Fusarium head blight resistance and leaf rust have been reported to be located on 2DS in wheat, close to lm4 [28,29,30,31,32,33]. In this study, a significant correlation was found between LMs and stripe rust resistance in the field (r = − 0.61, p < 0.01). The potential functions of lm4 in response to the above diseases in wheat deserve to be further studied.
Signaling pathways related to lesion mimics
In recent years, studies have reported that lesion mimics resembling a hypersensitivity reaction may enhance the resistance of plants by certain defense signaling pathways involved in plant disease resistance or stress resistance. Some LMs are associated with the production of ROS, which respond to cell death signals [3, 34, 35]. The inducible defense response of plants exists mainly to provide plants with an optimal defense system by relying on signaling pathways, such as those involving salicylic acid, jasmonic acid and ethylene, and the cross-talk between them. The salicylic acid-dependent pathway leads to cell death. In plants, cell death may play an important role in resistance to pathogens [36].
In addition, studies have reported that LMs may be associated with the programmed cell death signaling pathway. Pathogens have difficulty invading necrotic spots; therefore, LMs could improve plant disease resistance [37]. Plants have complex systems for regulating cell death, and these systems have a purpose in plant development against pathogens and environmental stress [37]. These results indicate that the mechanism of LM-enhanced plant resistance may be caused by associations with resistance genes or may involve signaling pathways to regulate plant defense responses and the programmed cell death pathway in plants. In this study, we mapped lm4 to a 50 Mb interval on 2DS and identified 18 predicted candidate genes (Table S3). Among these candidate genes, TraesCS2D02G090600 is related to the physiological defense response and immunity-related protein activity; TraesCS2D02G091200 and TraesCS2D02G092200 are involved in regulating cell death and defense upon pathogen recognition; and TraesCS2D02G090900 and TraesCS2D02G091100 are related to signal transduction. TraesCS2D02G091000, TraesCS2D02G091600, TraesCS2D02G091900 and TraesCS2D02G092100 function in response to stimuli, and TraesCS2D02G091300 and TraesCS2D02G091400 are related to leaf senescence and chloroplasts, respectively.
Lesion-mimic mutants can be a powerful tool to study their involvement in cell death. In addition to this genetic approach, physiological and biochemical characterization of the corresponding proteins was performed to identify the function of LM genes. This work should provide insight into cell death, defense or development through the determination of the biochemical functions of these proteins, their subcellular localization and their interacting proteins [3]. In the present study, fine mapping or gene cloning are needed for an improved understanding of the resistance mechanism and function of lm4. Studying the LM gene and its function is crucial for understanding the signaling pathways involved in plant apoptosis and disease resistance mechanisms.
Conclusions
A novel lesion-mimic gene (lm4) was identified by using a Yanzhan 1/Neixiang 188 RIL population. This gene is closely linked to SSR markers lm4_01_cib and lm4_02_cib, separated by 0.51 cM and 0.77 cM, respectively, intervals on 2DS. SSR markers were developed based on the content of Chinese Spring genome database and wheat 660 K SNP results, and these markers can be used for MAS of LM in wheat. In this study, we found that LMs were related to enhanced resistance to stripe rust in wheat. Therefore, resistance-related gene mapping (cloning) or resistant-cultivar breeding is an economical and environmentally friendly way to enhance the adaptability and yield stability of crops instead of the use of fungicide applications. In the present study, lm4 was associated with stripe rust resistance, and 18 candidate genes were chosen to analyze potential functions. LM gene cloning is required to understand the functions and disease resistance mechanism in wheat. This study may provide new ideas or theoretical support for future crop plant disease-resistance breeding and for understanding the plant apoptosis mechanism.
Methods
Plant materials
A total of 198 wheat recombinant inbred lines (RILs) of the Yanzhan 1 × Neixiang 188 mapping population (obtained from the Chinese Academy of Agricultural Sciences [CAAS]) were used for linkage analysis. The mapping population for this study was planted at the experimental station of the Chengdu Institute of Biology, Chinese Academy of Sciences, in Shifang (SF) and Maerkang (Ma) during the growing seasons of 2015 to 2018, according to local legislation in Sichuan Province (2015–2016, 2016–2017, 2017–2018 at SF; 2016–2017 and 2017–2018 at Ma). Twenty seeds of each accession were planted in a row. The stripe rust-susceptible wheat line Minxian 169 (obtained from the Chengdu Institute of Biology, Chinese Academy of Sciences), a control, was inserted after every 9 rows. Each experiment was arranged in a randomized complete block design, with two replicates, at Shifang and Maerkang from 2015 to 2018.
Evaluations of lesion-mimic phenotypes and agronomic traits
In total, 198 RILs and 2 parents (Yanzhan 1 and Neixiang 188) were evaluated for their lesion-mimic (LM) phenotype at SF (104°17′ E, 31°13′ N) and Ma (102°11′ E, 31°92′ N) in Sichuan Province from 2015 to 2018. The lesion-mimic phenotypes were arbitrarily subdivided into 5 scores based on flag leaf symptoms according to the methods of Yao et al. [17], with modifications. No visible lesions (specks) were recorded as 0 (the parental phenotype); few specks and low severity (< 25%) were recorded as 1; some specks and moderate severity (25–50%) were recorded as 2; large specks and high severity (50–75%) were recorded as 3; and a large number of specks and very high severity (> 75%) were recorded as 4. Plants with scores of 0 or 1 were considered normal, and those with scores of 2 or higher were classified as having lesion-mimic phenotypes.
Agronomic traits of the RILs were investigated by our team at the Chengdu Institute of Biology, Chinese Academy of Sciences, during the Shifang cropping seasons. These parameters included plant height (PH), spikelet number (SPI), number of sterile spikelets per spike (SSNS), grain number per spike (GNS), 1000-grain weight (TGW), and spike length (SL). Three to five plants of each wheat line were evaluated, and their means were used for analysis.
Evaluation of stripe rust resistance
All 198 RIL lines and the 2 parents were evaluated for stripe rust at Shifang and Maerkang from 2016 to 2018. Mixtures of Pst spores from races Pst-CYR32, Pst-CYR33, Pst-SU11, Pst-Hybrid46 and Pst-G22 (provided by SAAS) were suspended in 0.05% Tween 20 and sprayed onto four-leaf-stage wheat seedlings.
In the adult stage, stripe rust response types (ITs) were identified, and each environment was evaluated at least twice, mainly from 20 weeks to 23 weeks after sowing. Stripe rust infection types (ITs) were evaluated based on typical 0–4 classification systems [38].
Lesion-mimic gene mapping
Seedling leaves of the 198 RIL lines and two parents (Yanzhan 1, Neixiang 188) were collected, and genomic DNA was extracted from each sample using the CTAB method [39]. The quality and quantity of the DNA were determined using 1.0% agarose gel electrophoresis and a spectrophotometer (NanoDrop ND-1000, Thermo Scientific, Wilmington, DE). Two hundred and fifty-two polymorphic SSR markers covering 21 wheat chromosomes were used to genotype the mapping population to identify the chromosomal location of the LM gene (Additional file 1). Information about the SSR markers is available on the Grain Genes website (http://wheat.pw.usda.gov).
Based on the phenotypic evaluations, 10 wheat lines with an LM score of 0 and 10 RILs with an LM score of 4 were used to prepare two bulks representing extreme phenotypes. The DNA of these lines along with the parental lines was genotyped by 660 K SNP arrays at China Golden Marker Corporation (Beijing; http://www.cgmb.com.cn). Various SNP markers located on 2DS associated with lesion mimics were identified from the SNP typing results (Additional file 1). Whole wheat genome sequences were searched by SNP-tagged probe sequences (https://www.ncbi.nlm.nih.gov/) or according to the possible physical intervals on 2DS obtained from the SNP analysis search of the Chinese Spring genomic intervals (https://urgi.versailles.inra.fr/jbrowseiwgsc/gmod_jbrowse/?data=myData%2FIWGSC_RefSeq_v1.0&loc=chr2D%3A1..651852609&tracks=DNA&highlight=). A matched scaffold sequence was obtained, and repeated a DNA analysis was performed using the SSR Hunter 1.3 program (Li Qiang and Wan Jianmin 2005). The DNA sequences of both ends of the repeats were obtained, and primers were designed using Primer Premier 6.0 software (Canada). These primers (Table S1) were used for PCR- and electrophoresis-based analyses, and primers suitable for polymorphism were selected as molecular markers to obtain genotypes in the genetic population (Table S2).
PCR was conducted in a total volume of 20 μl comprising 200 ng of DNA template, 10 μl of 2× Es Taq MasterMix (Kangwei Century, China), 0.6 μl of 10 μM forward primer and 0.6 μl of 10 μM reverse primer. The amplification procedure was as follows: 94 °C for 5 min; 35 cycles of denaturation at 94 °C for 30 s, 45–60 °C (adjusted according to the primers) for 30 s, and 72 °C for 45 s; and then 72 °C for a total extension of 10 min. The separation of the PCR products was carried out by 1% agarose gel electrophoresis or 8% nondenaturing polyacrylamide gel electrophoresis. The agarose gel electrophoresis was performed with ethidium bromide (EB), and the polyacrylamide gel electrophoresis was performed with silver nitrate [40, 41].
Data analysis
All phenotypic data were recorded in Microsoft Office Excel 2013 for statistical analysis. One-way analysis of variance (ANOVA) was conducted to evaluate the variance and significance between groups by using SPSS 20.0 and GraphPad Prism 5.0. The genetic segregation ratio of normal (LM0–1) and lesion-mimic phenotypes (LM2–4) was tested by the chi-square test. Mean phenotypes of LMs and stripe rust scores for each RIL collected from each individual experiment were used for QTL analysis. The inclusive composite interval mapping of additive (ICIM-ADD) QTL method was used, and a walking speed of 1.0 cM with a stepwise regression probability of 0.001 was chosen for QTL detection. The threshold for declaring a significant QTL was determined by 1000-permutation tests. The LOD score to determine significant QTLs was 3.5 in all environments, and a LOD threshold of 3.5 was the criterion selected for a significant QTL. Linkage map construction and QTL mapping were performed using QTL IciMapping V4.1 software, and the genetic distance between markers was measured using centimorgans (cM) [42]. The threshold of the logarithm of odds value was set to 3.0 to determine linkage between markers, with a maximum recombination fraction at 0.4.
Availability of data and materials
The dataset and materials presented in the investigation are available from the supplementary tables and additional file 1.
Abbreviations
- LM:
-
Lesion mimic
- PCD:
-
Programmed cell death
- MAS:
-
Marker-assisted selection
- HRL:
-
Hypersensitive reaction-like
- RIL:
-
Recombinant inbred line
- PVE:
-
Phenotypic variation explained
- PH:
-
Plant height
- SPI:
-
Spikelet number
- SSNS:
-
Number of sterile spikelets per spike
- GNS:
-
Grain number per spike
- TGW:
-
1000-grain weight
- SL:
-
Spike length
- ITs:
-
Infection type
- Yr:
-
Stripe rust or yellow rust
- Pst:
-
Puccinia striiformis f. sp. tritici
References
Bruggeman Q, Raynaud C, Benhamed M, Delarue M. To die or not to die? Lessons from lesion mimic mutants. Front Plant Sci. 2015;6:24-45.
Li T, Bai G. Lesion mimic associates with adult plant resistance to leaf rust infection in wheat. Theor Appl Genet. 2009;119(1):13–21.
Lorrain S, Vailleau F, Balaque C, Roby D. Lesion mimic mutants: keys for deciphering cell death and defense pathways in plants? Trends Plant Sci. 2003;8(6):263–71.
Hoisington DA, Neuffer MG. Walbot V. disease lesion mimics in maize. 1. Effect of genetic background, temperature, developmental age, and wounding on necrotic spot formation with les1. Dev Biol. 1982;93(2):381–8.
Penning BW, Johal GS, McMullen MD. A major suppressor of cell death, slm1, modifies the expression of the maize (Zea mays L.) lesion mimic mutation les23. Genome. 2004;47(5):961–9.
Greenberg JT, Guo AL, Klessig DF, Ausubel FM. Programmed cell-death in plants-a pathogen-triggered response activated coordinately with multiple defense functions. Cell. 1994;77(4):551–63.
Li Y, Chen L, Mu J, Zuo J. LESION SIMULATING DISEASE1 interacts with catalases to regulate hypersensitive cell death in Arabidopsis. Plant Physiol. 2013;163(2):1059–70.
McGrann GRD, Steed A, Burt C, Nicholson P, Brown JKM. Differential effects of lesion mimic mutants in barley on disease development by facultative pathogens. J Exp Bot. 2015;66(11):3417–28.
Takahashi A, Kawasaki T, Henmi K, Shii K, Kodama O, Satoh H, Shimamoto K. Lesion mimic mutants of rice with alterations in early signaling events of defense. Plant J. 1999;17(5):535–45.
Li L, Shi X, Zheng F, Wu D, Li A-A, Sun F-Y, Li C-C, Wu J-C, Li T. Transcriptome analysis of Dlm mutants reveals the potential formation mechanism of lesion mimic in wheat. Eur J Plant Pathol. 2016;146(4):987–97.
Li L, Shi X, Zheng F, Li C, Wu D, Bai G, Gao D, Wu J, Li T. A novel nitrogen-dependent gene associates with the lesion mimic trait in wheat. Theor Appl Genet. 2016;129(11):2075–84.
Kamlofski CA, Antonelli E, Bender C, Jaskelioff M, Danna CH, Ugalde R, Acevedo A. A lesion-mimic mutant of wheat with enhanced resistance to leaf rust. Plant Pathol. 2007;56(1):46–54.
Nair SK, Tomar SMS. Genetical and anatomical analyses of a leaf flecking mutant in Triticum aestivum L. Euphytica. 2001;121(1):53–8.
Boyd LA, Minchin PN. Wheat mutants showing altered adult plant disease resistance. Euphytica. 2001;122(2):361–8.
Boyd LA, Smith PH, Wilson AH, Minchin PN. Mutations in wheat showing altered field resistance to yellow and brown rust. Genome. 2002;45(6):1035–40.
Kinane JT, Jones PW. Isolation of wheat mutants with increased resistance to powdery mildew from small induced variant populations. Euphytica. 2001;117(3):251–60.
Yao Q, Zhou R, Fu T, Wu W, Zhu Z, Li A, Jia JZ. Characterization and mapping of complementary lesion-mimic genes lm1 and lm2 in common wheat. Theor Appl Genet. 2009;119(6):1005–12.
Wang F, Wu W, Wang D, Yang W, Sun J, Liu D, Zhang A. Characterization and genetic analysis of a novel light-dependent lesion mimic mutant, lm3, showing adult-plant resistance to powdery mildew in common wheat. Plos One. 2016;11(5):e0155358.
Rosewarne GM, Herrera-Foessel SA, Singh RP, Huerta-Espino J, Lan CX, He ZH. Quantitative trait loci of stripe rust resistance in wheat. Theor Appl Genet. 2013;126(10):2427–49.
Wellings CR. Global status of stripe rust: a review of historical and current threats. Euphytica. 2011;179(1):129–41.
Johnson R. Durable resistance-definition of, genetic-control, and attainment in plant-breeding. Phytopathology. 1981;71(6):567–8.
Kou Y, Wang S. Wang, broad-spectrum and durability: understanding of quantitative disease resistance. Curr Opin Plant Biol. 2010;13(2):181–5.
Anand A, Schmelz EA, Muthukrishnan S. Development of a lesion-mimic phenotype in a transgenic wheat line overexpressing genes for pathogenesis-related (PR) proteins is dependent on salicylic acid concentration. Mol Plant-Microbe Interact. 2003;16(10):916–25.
Mu J, Huang S, Liu S, Zeng Q, Dai M, Wang Q, Wu J, Yu S, Kang Z, Han D. Genetic architecture of wheat stripe rust resistance revealed by combining QTL mapping using SNP-based genetic maps and bulked segregant analysis. Theor Appl Genet. 2019;132(2):443–55.
Johal GS, Lee EA, Close PS, Coe EH, Neuffer MG, Briggs SP. A tale of 2 mimics-transposon mutagenesis and characterization of 2 disease lesion mimic mutations of maize. Maydica. 1994;39(1):69–76.
Kjaer B, Jensen HP, Jensen J, Jorgensen JH. Associations between 3 ML-O powdery mildew resistance genes and agronomic traits in barley. Euphytica. 1990;46(3):185–93.
Chen XM. Epidemiology and control of stripe rust puccinia striiformis f. sp tritici on wheat. Can J Plant Pathol Rev Can Phytopathol. 2005;27(3):314–37.
Agenbag GM, Pretorius ZA, Boyd LA, Bender CM, Prins R. Identification of adult plant resistance to stripe rust in the wheat cultivar Cappelle-Desprez. Theor Appl Genet. 2012;125(1):109–20.
Powell NM, Lewis CM, Berry ST, MacCormack R, Boyd LA. Stripe rust resistance genes in the UK winter wheat cultivar Claire. Theor Appl Genet. 2013;126(6):1599–612.
Pawar SK, Sharma D, Duhan JS, Saharan MS, Tiwari R, Sharma I. Mapping of stripe rust resistance QTL in Cappelle-Desprez x PBW343 RIL population effective in northern wheat belt of India. 3 Biotech. 2016;6:76-83.
Bokore FE, Cuthbert RD, Knox RE, Randhawa HS, Hiebert CW, DePauw RM, Singh AK, Singh A, Sharpe AG, N'Diaye A, Pozniak CJ, McCartney C, Ruan Y, Berraies S, Meyer B, Munro C, Hay A, Ammar K, Huerta-Espino J, Bhavani S. Quantitative trait loci for resistance to stripe rust of wheat revealed using global field nurseries and opportunities for stacking resistance genes. Theor Appl Genet. 2017;130(12):2617–35.
Zou J, Semagn K, Chen H, Iqbal M, Asif M, N'Diaye A, Navabi A, Perez-Lara E, Pozniak C, Yang RC, Graf RJ, Randhawa H, Spaner D. Mapping of QTLs associated with resistance to common bunt, tan spot, leaf rust, and stripe rust in a spring wheat population. Molecular Breeding. 2017;37(12):144-57.
Eldakak M, Das A, Zhuang Y, Rohila JS, Glover K, Yen Y. A Quantitative proteomics view on the function of Qfhb1, a major QTL for Fusarium head blight resistance in wheat. Pathogens. 2018;7(3):58-84.
Jabs T, Dietrich RA, Dangl JL. Initiation of runaway cell death in an Arabidopsis mutant by extracellular superoxide. Science. 1996;273(5283):1853–6.
Torres MA, Dangl JL, Jones JDG. Arabidopsis gp91(phox) homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc Natl Acad Sci U S A. 2002;99(1):517–22.
Tomiyama K. Research on the hypersensitive response. Annu Rev Phytopathol. 1983;21:1–12.
Johal GS, Hulbert SH, Briggs SP. Disease lesion mimics of maize-a model for cell-death in plants. Bioessays. 1995;17(8):685–92.
McIntosh RA, Wellings CR, Park RF. Wheat rusts: An atlas of resistance genes. East Melbourne, Victoria, Australia: Commonwealth Scientific and Industrial Re-search Organisation (CSIRO); 1995.
Abu Almakarem AS, Heilman KL, Conger HL, Shtarkman YM, Rogers SO. Extraction of DNA from plant and fungus tissues in situ. BMC research notes. 2012;5:266.
An ZW, Xie LL, Cheng H, Zhou Y, Zhang Q, He XG, Huang HS. A silver staining procedure for nucleic acids in polyacrylamide gels without fixation and pretreatment. Anal Biochem. 2009;391(1):77–9.
Liu R, Lu J, Zhou M, Zheng SG, Liu ZH, Zhang CH, Du M, Wang MX, Li YF, Wu Y, Zhang L. Developing stripe rust resistant wheat (Triticum aestivum L.) lines with gene pyramiding strategy and marker-assisted selection. Genet Resour Crop Evol. 2020;67:381–91.
Meng L, Li H, Zhang L, Wang J. QTL IciMapping: integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015;3(3):269–83.
Acknowledgments
We thank Xia Xianquan (Sichuan Academy of Agricultural Sciences) for helping conduct field inoculations.
Funding
This work was supported by the ‘Strategic Priority Research Program’ of the Chinese Academy of Sciences (grant number XDA24030401-2), the “13th Five-year Plan” for National Key Research and Development (grant number 2017YFD0100902), and the “13th Five-year Plan” for Wheat Crops Breeding in Sichuan Province. The funders had no role in the study design, data collection, and analysis; the decision to publish; or the preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
LZ and YW established the experimental design and provided the plant material. RL performed the experiments, the 660 K SNP array data and the statistical analyses. RL and JL measured the agronomic traits of the wheat population. SGZ and RL analyzed the 660 K SNP array data. RL wrote the manuscript. MD, CHZ, MXW, YFL and JYX read the article and modified it. All the authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liu, R., Lu, J., Zheng, S. et al. Molecular mapping of a novel lesion mimic gene (lm4) associated with enhanced resistance to stripe rust in bread wheat. BMC Genom Data 22, 1 (2021). https://doi.org/10.1186/s12863-021-00963-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12863-021-00963-6