
Abstract
Background
The study was designed to explore the correlation of the asymmetric regulation between periaqueductal gray (PAG) and bilateral trigeminal nucleus caudalis (TNC) in migraine rats through studying the changes of metabolites in pain regulatory pathway of acute migraine attack.
Methods
Thirty male Sprague–Dawley (SD) rats were randomly divided into three groups: blank, control, model groups. Then, blank group was intraperitoneally injected with ultrapure water, while control group injected with saline and model group injected with Glyceryl Trinitrate (GTN). Two hours later, PAG and bilateral TNC were removed respectively, and metabolite concentrations of PAG, Left-TNC, Right-TNC were obtained. Lastly, the differences of metabolite among three brain tissues were compared.
Results
The relative concentrations of rNAA, rGlu, rGln, rTau, rMI in PAG or bilateral TNC had interaction effects between groups and sites. The concentration of rLac of three brain tissues increased in migraine rats, however, the rLac of LTNC and RTNC increased more than that of PAG. Besides, the concentrations of rNAA and rGln increased in RTNC, while rGABA decreased in RTNC.
Conclusions
There is correlation between PAG, LTNC and RTNC in regulation of pain during acute migraine attack, and the regulation of LTNC and RTNC on pain is asymmetric.
Similar content being viewed by others

Background
Migraine is a debilitating neurological disorder that leads to reduce the quality of life, increase anxiety and depression, and increase brain infarction and cognitive dementia, affecting an estimated 100 million people worldwide [1, 2]. With the increase of the global population, the prevalence of migraine is also on the rise which increases the burden of the health, living and economic worldwide [3]. However, the pathogenesis of migraine is complex and needs to be further studied. At present, trigeminal vascular reflex theory is believed to be the core mechanism of migraine, the central projections of injurious primary afferent fibers such as dura enter the caudal canal of medulla oblongata in the brainstem through the trigeminal tract, terminating mainly in the trigeminal nucleus caudalis (TNC), so the inhibition of TNC has been shown to be a predictor of efficacy in the treatment of migraine [4,5,6,7]. The research on TNC is mainly conducted on both sides. However, with the in-depth research on the left and right brain functions, it is found that the two hemispheres have different functions [8], at the same time, previous studies [9] have found that after electroacupuncture stimulation of the left and right thalamus in the treatment of migraine, the values of NAA /Cr and Cho/Cr in the left thalamus are significantly increased compared with those before treatment. In addition, there is almost no statistical difference in Cho/ Cr while the values of NAA/ Cr in the right thalamus increased compared with those before treatment. This result suggests that the pathogenesis of migraine may also have differences between left and right regulatory pathways.
The brainstem pain regulatory system plays an important role in the pathophysiology of migraine by affecting trigeminal vascular nociceptive information transmission and central sensitization [10]. Current studies [11, 12] have found that the pain regulation system in the descending brainstem is mainly from the periaqueductal gray (PAG) -rostral ventromedial medulla (RVM) circuit to the medullary and spinal dorsal horn, and the pain regulation system can play a bidirectional role resulting in the suppression and facilitation of pain. Among them, PAG is the core part of the central endogenous analgesic system, which can inhibit the nociceptive afferents of the trigeminal neurovascular system to exert analgesic function.
Proton magnetic resonance spectroscopy (1H-MRS) is one of the methods for noninvasive quantitative analysis of metabolism, biochemical changes and compounds in human organs and tissues. Many metabolites such as N-acetyl aspartate (NAA), lactate (Lac), creatine (Cr) and glutamate (Glu) can be detected simultaneously [13]. In recent years, MRS has been used to study the changes of brain metabolites and neurotransmitters in migraine patients [14,15,16]. Therefore, in this study, MRS was used to observe the changes of metabolism and neurotransmitters of bilateral TNC and PAG through Glyceryl Trinitrate (GTN) -induced migraine rat model, and the relationship between the differences of metabolism and neurotransmitter changes during pain attack was compared, so as to reveal the correlation between the asymmetric regulation of PAG and bilateral TNC in migraine. Lastly, previous studies [17,18,19] explored the correlation between GTN-induced hyperalgesia and temporal changes in rats, and found that GTN concentration reached its maximum in the brain 2 hours after GTN injection, causing a significant increase in dural mast cell degranulation and an upregulation of mRNA and protein expression of the nociceptive marker c-fos in TNC; On this basis, this study selected 2 h after rat modeling to study the changes of metabolites.
Materials and methods
Animals
Thirty male Sprague–Dawley rats, weighing 140-170 g, purchased from Shanghai Xipuer-Bikai Laboratory Animal Co., Ltd. (License No. SCXK [hu] 2013–0016) were used in the study. The rats were housed in the Laboratory Animal Center of Zhejiang Chinese Medical University with a 12-h light–dark cycle (22.7 ± 0.2 °C and 49 ± 10% humidity) and (had) free access to water and food. The study was approved by the Laboratory Animal Center of Zhejiang Chinese Medical University. Besides, animal feeding management and animal experiment operation shall comply with《Zhejiang province Administration Rule of Laboratory Animal》.
Experimental design
Thirty rats were weighed and numbered, then the rats were randomly divided into 3 groups by the excel (n = 10/group): blank group, control group and model group. All of the three groups were gavaged with ultra-pure water (administration volume 1 ml/100 g) for 7 consecutive days, once a day in the morning. At the end of gavage on the seventh day, after 0.5 hours of adaptation in a transparent tempered glass box (30 cm × 30 cm × 40 cm in length, width and height), the blank group was intraperitoneally injected with ultra-pure water (administration volume: 0.2 ml/100 g), the control group was intraperitoneally injected with normal saline (administration volume: 0.2 ml/100 g), and the model group was intraperitoneally injected with GTN (administration dose: 10 mg/kg, administration volume: 0.2 ml / 100 g). Molding time: 9:30–10:00 a.m. After intraperitoneal injection, the rats were placed in the transparent tempered glass box, and the surrounding environment was kept quiet. All the animals were recorded for 2 hours of behavioral science. Within 2 h, the model animals showed burnout and reduced movement.
Samples collection
Two hours after modeling, the rats were completely anesthetized by intraperitoneal injection of 1% sodium pentobarbital (administration dose: 50 mg/kg, administration volume: 0.5 ml/100 g). The whole brain tissue removed from the rat was immediately stored in an ultra-low temperature refrigerator at − 80 °C. Then, the brain tissues of the PAG, LTNC and RTNC were respectively obtained according to the Stereoscopic Map of Rat Brain in the frozen microsection machine at − 20 °C and stored in the refrigerator at − 80 °C.
1H-MRS analysis
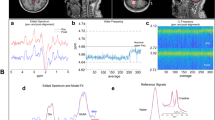
PAG/LTNC/RTNC samples were damaged in one rat in the blank group. Finally, 87 samples were included in the study. The MRS of all the samples was detected in College of Life Sciences, Zijingang Campus, Zhejiang University. Firstly, all the samples were weighed with an electronic balance and recorded. Secondly, the samples were individually ground in a grinding tube containing 600ul of 0.05% TSP-d4 heavy water and then transferred to a specific 5-mm diameter NMR tube. Lastly, all the samples were analyzed on a Bruker 600 MHz AVANCE III spectrometer equipped with a 5 mm-BBFO probe. After D2O locking field and automatic homogenization field, the pressure water peak map was obtained by NoesyPR1D sequence with the acquisition time of 1.7 s, the acquisition times of 128, the data points of 32 K, and the scanning time of 8.9 min. In the end, the original data was obtained by using Bruker Topspin 3.2.
1H-MRS data processing
All the free induction decays (FIDs) from 1H-MRS of the brain tissues were processed by MestRe Nove, a spectrum analysis and processing software, via fourier transformation, phase correction, and baseline correction with trimethylsilane propionate (TSP) as internal standard (chemical shift 0.00 PPM). The relevant metabolite concentrations were calculated by the formula CM = AM/ATSP*fP*CTSP*MWM/WM and denoted by “i”; The ratio of the relevant metabolite concentration to TSP and weight (i/TSP/weight) is considered as the absolute concentration and is denoted by “c”; The ratio of the relevant metabolite concentration to creatine phosphate (tCr) is considered as the relative concentration and is denoted by “r” (Fig. 1).

RTNC MRS image of the three groups sample graph, blank (14 mg), control9 (14 mg), model (18 mg)
Statistical analysis
Statistical analyses were performed with SPSS Statistics (version 25.0). GraphPad Prism(version 9.0) statistical graphics. The measurement data were expressed as mean ± standard deviation (−x ± s) if they were in accordance with normal distribution, otherwise, they were expressed as median (P25, P75). Enumeration data were expressed as rate or constituent ratio. The differences of brain metabolites in PAG, RTNG, LTNG of the three groups were analyzed by generalized estimating equation. For brain metabolites with interaction effects, it is necessary to fix the location and analyze the differences between groups in each location separately. In addition to that, for the data with normal distribution and homogeneity of variance, one-way analysis of variance was used for comparison between multiple groups (more than 2 groups), and LSD method was used for pairwise comparison between groups. For the data with were non-normal, Kruskal-Wallis H method was used for comparison between multiple groups, and all pairwise method was used for pairwise comparison between groups. The value of P < 0.05 was considered statistically significant.
Results
The interaction analysis of brain metabolites in PAG, LTNC and RTNC
We found that the group and site interaction effects of brain metabolites of rNAA (P = 0.036), rGlu (P = 0.001), rGln (P = 0.001), rTau (P = 0.023), rMl (P = 0.023) were statistically different, while the other brain metabolites (rLac, rAla, rAce, rGABA, rGly) group and site interaction effects were not statistically significant.
As shown in Fig. 2, there was no significant difference of rNAA in PAG and LTNC between groups (P > 0.05). However, the rNAA of control group (P = 0.021) and blank group (P = 0.023) were lower than those of model group in RTNC.

Comparison of rNAA, rGln, rGlu, rTau and rMl in PAG, LTNC and RTNC (n = 29); * denotes P < 0.05, ** denotes P < 0.01; The rNAA, rGln of RTNC in the control group were lower than the model group, while the rGlu, rTau of RTNC had no significant difference between groups
Furthermore, there was no significant difference of rGln in PAG and LNTC between groups (P > 0.05). However, the rGln of the control group (P = 0.028) was lower than that of the model group, and there was no significant difference between the blank group and the model group (p > 0.05) in RTNC.
Besides, there was no significant difference of rGlu in PAG,LTNC and RTNC between groups (P > 0.05). Likewise, there was no significant difference of rTau in PAG, LTNC and RTNC between groups (P > 0.05).
Lastly, in the Fig. 2, we can know that there was no significant difference of rMI in PAG between groups (P > 0.05). However, there was significant difference between LTNC rMl groups (P = 0.035) which suggested that there was no significant difference between control group and model group (P = 1.000), and no significant difference between blank group and model group (P = 0.050). There was significant difference between RTNC rMl groups (P = 0.039) which showed that there was no significant difference between control group and model group (P > 0.05), and significant difference between blank group and model group (P = 0.032).
Comparison of relative concentration of total brain metabolites in different parts
As shown in the Fig. 3, there were significant differences of rLac between groups (P = 0.000) which showed that the differences between blank group (P = 0.000), control group (P = 0.002) and model group were statistically significant. In addition to that, there were statistically significant differences of rLac between PAG, LTNC and RTNC (P = 0.000), suggesting the rLac difference between PAG and RTNC was statistically significant (P = 0.000), while the rLac difference between LTNC and RTNC was not statistically significant (P > 0.05).

Comparison of relative concentration of total brain metabolites in different parts. Compared with RTNC, * denotes P < 0.05, ** denotes P < 0.01; Compared with the model group, # indicates P < 0.05, and ## indicates P < 0.01. There were no significant differences in rLac, rAla, rAce, rGABA and rGly between LTNC and RTNC; There were significant differences in rLac, rAce, rGABA and rGly between PAG and RTNC which showed the rLac and rGly of PAG were lower than those of RTNC while the rAce and rGABA of PAG were higher than those of RTNC. The rLac, rAla and rGABA were statistically different among the groups (PAG, LTNC and RTNC), indicating the rAla and rGABA in the model group were lower than those in the control group and the blank group, when the rLac in the model group were higher than those in the control group and the blank group
There were significant differences of rAla between groups (P = 0.009) which showed that the differences between blank group (P = 0.041), control group (P = 0.006) and model group were statistically significant, on the contrary, there were no statistically significant differences of rAla between PAG, LTNC and RTNC (P > 0.05).
About the rAce, there were no significant differences between groups (P > 0.05). However, there were statistically significant differences between PAG, LTNC and RTNC (P = 0.000), suggesting the difference between PAG and RTNC was statistically significant (P = 0.000), while the difference between LTNC and RTNC was not statistically significant (P > 0.05).
Moreover, there were significant differences of rGABA between groups (P = 0.000) which showed that the differences between blank group (P = 0.000), control group (P = 0.001) and model group were statistically significant, likewise, there were statistically significant differences of rGABA between PAG, LTNC and RTNC (P = 0.000), suggesting the difference between PAG and RTNC was statistically significant (P = 0.000), while the difference between LTNC and RTNC was not statistically significant (P > 0.05).
Finally, about the rGly, there were no significant differences between groups (P > 0.05). However, there were statistically significant differences between PAG, LTNC and RTNC (P = 0.000), suggesting the difference between PAG and RTNC was statistically significant (P = 0.000), while the difference between LTNC and RTNC was not statistically significant (P > 0.05).
Comparison of brain metabolites in different parts (no interaction effect)
As shown in the Tables 1 and 2. In the PAG, the differences of rLac between groups were statistically significant (P = 0.026) which showed the model group was significantly higher than the blank group (P = 0.012) and the control group (P = 0.032).
In the LTNC, the differences of rLac between groups were statistically significant (P = 0.006) which showed the model group was significantly higher than the blank group (P = 0.002) and the control group (P = 0.037).
In the RTNC, the differences of rLac between groups were statistically significant (P = 0.000) which showed the model group was significantly higher than the blank group (P = 0.000) and the control group (P = 0.002).
In the RTNC, the differences of rNAA between groups were statistically significant (P = 0.030) which showed the model group was higher than that of the blank group (P = 0.023) and the control group (P = 0.021).
In the RTNC, the differences of rGABA between groups were statistically significant (P = 0.001) which showed the model group was lower than that of the blank group (P = 0.001) and the control group (P = 0.001).
In the RTNC, the differences of rGln between groups were statistically significant (P = 0.030) which showed the model group was higher than that of the control group (P = 0.028). However, there was no statistical difference between the model group and the blank group (P = 1.000).
Discussion
The current study was designed to explore the correlation of the asymmetric regulation between the periaqueductal gray (PAG) and the bilateral trigeminal nucleus caudalis (TNC) in migraine rats through the changes of metabolites and neurotransmitters of PAG and TNC in the pain regulatory pathway of acute migraine attack. In our study, we found that the metabolites and neurotransmitters of the PAG, the LTNC and the RTNC had interaction effects between groups and sites. The concentration of the rLac of the PAG, LTNC and RTNC increased in migraine rats, however, the rLac of the LTNC and the RTNC increased more than that of the PAG. Besides, the concentrations of metabolites and neurotransmitters were asymmetrical in the LTNC and the RTNC which showed that the rNAA and rGln increased in the RTNC, while rGABA decreased in the RTNC. Therefore, we can conclude that there is a correlation between the PAG, LTNC and RTNC in the regulation of pain during the acute attack of migraine, and the regulation of the LTNC and RTNC on pain is asymmetric.
Our study found that the rLac increased in PAG, RTNC, and LTNC in migraine rats. Lac is the product of anaerobic glycolysis. The increase of Lac in the brain is thought to be caused by mitochondrial dysfunction or insufficient oxygen content [20]. However, in recent years, it has been found that increased neuronal activity can lead to a rapid and transient increase of Lac in brain [21]. In addition, it has also been found that Lac increases in the occipital cortex and cerebellum during the interval of migraine attacks [22,23,24,25]. TNC is an important structure for nociceptive generation, so when pain stimuli are transmitted to TNC, the neuronal activity of TNC increases, which leads to the increase of Lac in TNC on both sides, suggesting that TNC neurons are activated bilaterally. PAG is the core part of the central endogenous analgesic system, in the early stage of migraine attack, the Lac of PAG increased less than that of TNC which suggests that the neuronal activity of PAG also increased less than that of TNC, indicating that PAG neurons respond to pain in the early stage, however, it needs further investigation whether this early response promotes pain or involved in pain suppression.
Our study also found that rNAA increased on RTNC in migraine rats, whereas rNAA did not show such a difference on LTNC. NAA is synthesized by aspartic acid and acetyl-CoA (CoA) under the catalysis of aspartic acid N-acetyltransferase [26], which is mainly concentrated in gray matter rich areas in the brain [27]. The synthesis of NAA depends on the integrity of neuronal mitochondria, and the fluctuation of concentration of NAA may occur simultaneously with the change of adenosine triphosphate (ATP) which suggests that it is closely related to metabolic energy [28, 29]. As a result, when neurons are stimulated, NAA will be released [30]. TNC is an important structure for nociceptive production, which contains a relatively rich ratio of neurons to glial cells, during pain stimulation, neurons release a large amount of stored NAA, which increases its concentration transiently. However, in our study, there is no similar phenomenon on LTNC and PAG which is an interesting phenomenon, According to Lac, we can know that bilateral TNC and PAG have activation changes in the early stage of migraine, but NAA, as a marker of neuronal activity, only increases in RTNC, therefore, it needs further study whether NAA is an important marker of neuronal activation related to pain generation or transmission. At the same time, this phenomenon also suggests that TNC on both sides have differences in conduction of pain sensation when exposed to pain stimuli.
In our study, we also found that rGABA decreased and Gln increased in RTNC of migraine rats. Gln is a metabolite of the excitatory neurotransmitter glutamate (Glu), specifically produced in brain astrocytes by glutamate and ammonia under the catalysis of glutamine synthetase. Then, glutamine is transported back to neurons, where it is converted to Glu by mitochondrial glutaminase [31]. This circulation is thought to prevent neuroexcitability caused by the production of too much Glu in the brain. GABA is a major inhibitory neurotransmitter in the central nervous system, which is directly synthesized by Glu through glutamate decarboxylase [32]. Meanwhile, the imbalance of Glu-GABA concentration has been considered as another hypothesis for the occurrence of migraine [33]. Zhe Ma et al. [18] found a significant increase of Glu/GABA+ in the thalamus of migraine rats, whereas Nastaren Abad et al. [19] did not find the similar increase in cortical regions of the migraine rat. The increased Gln in our study may be due to the rapid action of the excitatory neurotransmitter Glu early in migraine attack, causing pain to occur and producing excessive Gln. At the same time, when the pain stimulus is transmitted to the RTNC, the inhibitory neurotransmitter GABA decreases which reduces the pain threshold and improves the excitability of RTNC, and thus accelerates the conduction of pain stimulus signal.
In addition, our study found that the relative concentrations of brain metabolites rNAA, rGlu, rGln and rTau in the PAG and the bilateral TNC had interaction effects between groups and sites. However, the interaction effect could not be proved by current experiments and its significance needs to be further studied.
Our study identified asymmetric changes of metabolic substances and neurotransmitters in the left and right TNC during headache attack in a migraine rat model, which was first found in the study of migraine attack mechanism. These asymmetrical changes are consistent with our recent clinical findings which is that some migraine patients with patent foramen ovale (PFO) often have long-term unilateral headache, in recent years, many researches [34,35,36] have identified a relationship between PFO and migraine headache which also indirectly suggests the possibility of bilateral asymmetric regulation of migraine. In addition to that, Momoh-Ojewuyi et al. [37] found that a small percentage of people experienced long-term migraine with unilateral fixation, which is called side locked unilateral headache (SLUH). Besides, Shinoura N et al. [38] found that stress-related vascular responses were suppressed in the right cerebral hemisphere during migraine interictal periods, which indirectly suggested left-right cerebral functional asymmetry in migraine mechanisms. In addition, this functional asymmetry has been studied in other disorders, for example, antidepressants can increase activity in the left hemisphere [39, 40], Kong XZ et al. [41] have also shown that autism spectrum disorder (ASD) is associated with a subtle reduction in cortical thickness asymmetry that is widely distributed in cortical regions. Therefore, asymmetric regulation of brain function has been gradually discovered and studied in some diseases. Finally, our study points out that there are asymmetric changes of metabolites in left and right TNC during migraine attacks, these asymmetrical changes are consistent with Momoh-Ojewuyi et al’s found [37] which not only provides a new insight into the clinical application of unilateral nerve electrical stimulation in the treatment of migraine when drugs cannot provide sufficient relief, but also provides a new direction for future research on the pathogenesis of migraine.
In this study, the rat model of migraine was induced by GTN which mimic those migraine patients caused by meningeal inflammation and dural mast cell degranulation in clinic [18]. Although some migraine rats in the model group showed some complications such as burnout behavior at the end of the experiment, this model was preferred compared to other migraine rat models because of the other rat migraine models [42, 43] require invasive manipulation of the rat head, which can affect the MRI scan. Therefore, in order to obtain a more stable and undisturbed spectrum, we chose GTN for migraine induction.
The behavioral manifestations of migraine model induced by nitroglycerin were mostly as follows: allodynia [44] and hyperalgesia [45]. Allodynia test include heat sensitivity and Mechanical nociceptive thresholds, both are hurtful stimuli. GTN-induced hyperalgesia included the tail flick test and the formalin test, which is also the hurtful stimuli. These behavioral tests affect the purpose of the experiment. For another, the spontaneous behavior of headache induced by GTN was unstable. Although light sensitivity changes before and after the model, it does not have repeatability. Burrowing behavior, running wheel activity and light sensitivity cannot be used to establish successful behavioral criteria for GTN mode [46]. Recent studies have shown that reflex blink-associated EMG response [47] can be used as a successful evaluation criterion for 2 h modeling of migraine induced by GTN, but this test requires anesthesia rather than wakefulness, so it is not suitable for this study.
Conclusion
In this study, we found that both TNC and PAG were activated in the early stage of migraine, but there were differences in the pain response of the left and right TNC, and the right TNC played a key role in the generation and conduction of pain, which also suggested that the occurrence of migraine was asymmetric regulation of TNC on both sides. And the role of early activation of PAG in migraine regulation needs to be further studied.
In conclusion, we found the differences in the concentrations of metabolic substances between PAG and TNC in the left and right sides of migraine rats, revealing the asymmetry of TNC in the regulation of migraine, and PAG is also activated in the early stage of migraine, which provides theoretical help for further understanding the pathophysiology basis of migraine.
However, this study has some limitations: Firstly, this study only compared the changes of metabolites in brain tissue 2 h after modeling, so subsequent studies need to select multiple time points for measurement to conduct more in-depth research; Secondly, male rats were used in this study, and female rats need to be included in subsequent experiments which could make the experimental design more perfect. Thus, more researches are also necessary.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- PAG:
-
Periaqueductal gray
- SD:
-
Sprague–Dawley
- NAA:
-
N-acetylaspartate
- Gln:
-
Glutamine
- MI:
-
Myo-inositol
- GABA:
-
γ-aminobutyric acid
- Cho:
-
Choline
- 1H-MRS:
-
Proton magnetic resonance spectroscopy
- Ala:
-
Aminolevulinic acid
- Gly:
-
Glycine
- PFO:
-
Patent foramen ovale
- ASD:
-
Autism spectrum disorder
- TNC:
-
Trigeminal nucleus caudalis
- GTN:
-
Glyceryl Trinitrate
- Glu:
-
Glutamate
- Tau:
-
Taurine
- Lac:
-
Lactate
- Cr:
-
Creatine
- RVM:
-
Rostral ventromedial medulla
- Ace:
-
Angiotensin-Converting Enzyme
- ATP:
-
Adenosine triphosphate
- CGRP:
-
Calcitonin Gene-Related Peptide
References
GBD 2016 Disease and Injury Incidence and Prevalence Collaborators (2017) Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: a systematic analysis for the Global Burden of Disease Study 2016. Lancet 390(10100):1211–1259
Ashina M et al (2020) N Engl J Med 383(19):1866–1876
Ashina M, Katsarava Z, Do TP et al (2021) Migraine: epidemiology and systems of care. Lancet. 397(10283):1485–1495
Burstein R, Yamamura H, Malick A, Strassman AM (1998) Chemical stimulation of the intracranial dura induces enhanced responses to facial stimulation in brain stem trigeminal neurons. J Neurophysiol 79(2):964–982
Liu Y, Broman J, Edvinsson L (2004) Central projections of sensory innervation of the rat superior sagittal sinus. Neuroscience. 129(2):431–437
Wang X, Yu S, Dong Z, Jiang L (2011) The Fos expression in rat brain following electrical stimulation of dura mater surrounding the superior sagittal sinus changed with the pre-treatment of rizatriptan benzoate. Brain Res 1367:340–346
Pei P, Liu L, Zhao L, Cui Y, Qu Z, Wang L (2016) Effect of electroacupuncture pretreatment at GB20 on behaviour and the descending pain modulatory system in a rat model of migraine. Acupunct Med 34(2):127–135
Kuo F, Massoud TF (2022) Structural asymmetries in normal brain anatomy: A brief overview. Ann Anat 241:151894
Lin L, Ding R (2015) Effects of electroacupuncture on the metabolites of right thalamus and anterior cingulate cortex in patients with migraine: magnetic resonance spectroscopy study. J Tradit Chin Med. 56(14):1220–1223
Noseda R, Burstein R (2013) Migraine pathophysiology: anatomy of the trigeminovascular pathway and associated neurological symptoms, cortical spreading depression, sensitization, and modulation of pain. Pain. 154(Suppl 1):S44–S53
Hoffmann J, Baca SM, Akerman S (2019) Neurovascular mechanisms of migraine and cluster headache. J Cereb Blood Flow Metab 39(4):573–594
Solstrand Dahlberg L, Linnman CN, Lee D, Burstein R, Becerra L, Borsook D (2018) Responsivity of periaqueductal gray connectivity is related to headache frequency in episodic Migraine. Front Neurol 9:61
Egerton A (2021) The potential of 1H-MRS in CNS drug development. Psychopharmacology 238(5):1241–1254
Peek AL, Leaver AM, Foster S et al (2021) Increased GABA+ in people with Migraine, headache, and pain conditions- A potential marker of pain. J Pain 22(12):1631–1645
Wu X, Han S, Yang Y et al (2022) Decreased brain GABA levels in patients with Migraine without Aura: an exploratory proton magnetic resonance spectroscopy study. Neuroscience. 488:10–19
Younis S, Hougaard A, Christensen CE et al (2021) Interictal pontine metabolism in migraine without aura patients: A 3 tesla proton magnetic resonance spectroscopy study. Neuroimage Clin 32:102824
Tassorelli C, Greco R, Wang D, Sandrini M, Sandrini G, Nappi G (2003) Nitroglycerin induces hyperalgesia in rats--a time-course study. Eur J Pharmacol 464(2–3):159–162
Pedersen SH, Ramachandran R, Amrutkar DV, Petersen S, Olesen J, Jansen-Olesen I (2015) Mechanisms of glyceryl trinitrate provoked mast cell degranulation. Cephalalgia. 35(14):1287–1297
Ramachandran R, Bhatt DK, Ploug KB et al (2012) A naturalistic glyceryl trinitrate infusion migraine model in the rat. Cephalalgia. 32(1):73–84
Arngrim N, Schytz HW, Britze J et al (2016) Migraine induced by hypoxia: an MRI spectroscopy and angiography study. Brain. 139(Pt 3):723–737
Li B, Freeman RD (2015) Neurometabolic coupling between neural activity, glucose, and lactate in activated visual cortex. J Neurochem 135(4):742–754
Grimaldi D, Tonon C, Cevoli S et al (2010) Clinical and neuroimaging evidence of interictal cerebellar dysfunction in FHM2. Cephalalgia. 30(5):552–559
Watanabe H, Kuwabara T, Ohkubo M, Tsuji S, Yuasa T (1996) Elevation of cerebral lactate detected by localized 1H-magnetic resonance spectroscopy in migraine during the interictal period. Neurology. 47(4):1093–1095
Sándor PS, Dydak U, Schoenen J et al (2005) MR-spectroscopic imaging during visual stimulation in subgroups of migraine with aura. Cephalalgia. 25(7):507–518
Reyngoudt H, Achten E, Paemeleire K (2012) Magnetic resonance spectroscopy in migraine: what have we learned so far? Cephalalgia. 32(11):845–859
Patel TB, Clark JB (1979) Synthesis of N-acetyl-L-aspartate by rat brain mitochondria and its involvement in mitochondrial/cytosolic carbon transport. Biochem J 184(3):539–546
Tallan HH (1957) Studies on the distribution of N-acetyl-L-aspartic acid in brain. J Biol Chem 224(1):41–45
Signoretti S, Marmarou A, Tavazzi B, Lazzarino G, Beaumont A, Vagnozzi R (2001) N-Acetylaspartate reduction as a measure of injury severity and mitochondrial dysfunction following diffuse traumatic brain injury. J Neurotrauma 18(10):977–991
Dehghan A, Saatchian E, Sobhani M, Montazerabadi A (2020) Neurochemical metabolite alterations of the occipital lobe in migraine without aura by proton magnetic resonance spectroscopy. Neuroradiol J 33(5):410–415
Amaral IA, Hadera MG, Kotter M, Sonnewald U (2017) Oligodendrocytes Do not export NAA-derived aspartate in vitro. Neurochem Res 42(3):827–837
Andersen JV, Markussen KH, Jakobsen E et al (2021) Glutamate metabolism and recycling at the excitatory synapse in health and neurodegeneration. Neuropharmacology. 196:108719
Hao J, Jiang K, Zhang X, Wu M, Ma B (2021) “Glu/GABA-Gln” metabolic loop abnormalities in iminodipropionitrile (IDPN)-induced dyskinetic syndrome. Neurol Sci 42(11):4697–4706
Younis S, Hougaard A, Vestergaard MB, Larsson HBW, Ashina M (2017) Migraine and magnetic resonance spectroscopy: a systematic review. Curr Opin Neurol 30(3):246–262
Kumar P, Kijima Y, West BH, Tobis JM (2019) The connection between patent foramen Ovale and Migraine. Neuroimaging Clin N Am 29(2):261–270
Wang YL, Wang FZ, Zhang Y et al (2022) Association of migraine with patent foramen ovale closure: A systematic review and meta-analysis. Int J Cardiol Heart Vasc 39:100992 Published 2022 Mar 18
Qi Y, Zhang Y, Luo X et al (2021) Efficacy of patent foramen ovale closure for treating migraine: a prospective follow-up study. J Investig Med 69(1):7–12
Momoh-Ojewuyi A, Ayubi A, Abusamra R et al (2013) Side locked headaches. J Headache Pain 14(Suppl 1):P64
Shinoura N, Yamada R (2005) Decreased vasoreactivity to right cerebral hemisphere pressure in migraine without aura: a near-infrared spectroscopy study. Clin Neurophysiol 116(6):1280–1285
Rotenberg VS (2008) Functional brain asymmetry as a determinative factor in the treatment of depression: theoretical implications. Prog Neuro-Psychopharmacol Biol Psychiatry 32(8):1772–1777
Segarra AB, Prieto I, Martínez-Cañamero M, Ramírez-Sánchez M (2020) Is there a link between depression, neurochemical asymmetry and cardiovascular function? AIMS Neurosci 7(4):360–372 Published 2020 Sep 28
Kong XZ, Postema MC, Guadalupe T et al (2022) Mapping brain asymmetry in health and disease through the ENIGMA consortium. Hum Brain Mapp 43(1):167–181
Yao G, Huang Q, Wang M, Yang CL, Liu CF, Yu TM (2017) Behavioral study of a rat model of migraine induced by CGRP. Neurosci Lett 651:134–139
Pei P, Liu L, Zhao LP et al (2019) Electroacupuncture exerts an anti-migraine effect via modulation of the 5-HT7 receptor in the conscious rat. Acupunct Med 37(1):47–54
Bates EA, Nikai T, Brennan KC, Fu YH, Charles AC, Basbaum AI, Ptacek LJ, Ahn AH (2010) Sumatriptan alleviates nitroglycerin-induced mechanical and thermal allodynia in mice. Cephalalgia 30(2):170–178
Greco R, Mangione AS, Siani F, Blandini F, Vairetti M, Nappi G, Sandrini G, Buzzi MG, Tassorelli C (2014) Effects of CGRP receptor antagonism in nitroglycerin-induced hyperalgesia. Cephalalgia 34(8):594–604
Christensen SLT, Petersen S, Sorensen DB, Olesena J, Jansen-Olesen I (2016) Infusion of low dose glyceryl trinitrate has no consistent effect on burrowing behavior, running wheel activity and light sensitivity in female rats. J Pharmacol Toxicol Methods 80:43–50
Jones MG, Andreou AP, McMahon SB, Spanswick D (2016) Pharmacology of reflex blinks in the rat: a novel model for headache research. J Headache Pain 17(1):96
Acknowledgements
Not applicable.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Research was supported by the Zhejiang Provincial Natural Science Foundation of China Grant LY18H270009; The Health Commission of Zhejiang Province Grant 2022KY222; National Natural Science Foundation of China Grant 81302938; Medicine and Healthcare Scientific Research Foundation of Zhejiang Province of China Grant 2017KY118; and Zhejiang Provincial Inheritance Studio of Prominent Chinese Medicine Expert Prof. Qiu Changlin Grant GZS2021007).
Author information
Authors and Affiliations
Contributions
ZXZ conceived the study design and supervised the scientific work. WJY and LPZ performed the animal experiments. WJY, LPZ, JJY and JFL analyzed the data and prepared the tables and graphs. ZJC and WJY wrote the manuscript. MSX, ZJC and ZXZ revised the manuscript. All authors read, reviewed, edited, and approved the final manuscript. Zhijian Cao and Wenjing Yu contributed equally to this work.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The study was approved by the Laboratory Animal Center of Zhejiang Chinese Medical University.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Cao, Z., Yu, W., Zhang, L. et al. A study on the correlation of the asymmetric regulation between the periaqueductal gray and the bilateral trigeminal nucleus caudalis in migraine male rats. J Headache Pain 24, 27 (2023). https://doi.org/10.1186/s10194-023-01559-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s10194-023-01559-4