
Abstract
Background
Taxonomic uncertainties in the morphological species identification and taxonomic revisions in individual groups are known for all echinoderm classes. These uncertainties in morphological species identification and discrimination have spawned the application of molecular genetic identification techniques. However, as the fundamental step to allow and ensure future molecular species identification, valid and comprehensive reference library entries comprising morphological and molecular species information together with various metadata are essentially needed. In our study we compare morphological and molecular genetic species identification techniques for representatives of North Sea echinoderm classes, i.e. the Asteroidea, Ophiuroidea, Echinoidea and Holothuroidea.
Methods
Individuals were sampled during different surveys in different regions of the North Sea, identified to species level based on morphological diagnostic features, and were genetically analysed using a fragment of the mitochondrial cytochrome c oxidase subunit I (COI).
Results and Discussion
The morphological determination revealed 32 species including one taxon determined only to genus level. In contrast to this, the COI analysis supported 34 monophyletic clades with pronounced differences between the intra- and the inter-specific genetic variability (a barcoding gap of 4.93 %) with highest intra-specific variabilities found in the ophiuroid species Amphiura filiformis, A. chiajei and Ophiura sarsii. In 94 % of the investigated species, morphological identification and COI sequence clusters were congruent whereas for two asteroid species we found an underestimated diversity. For Astropecten irregularis, one of the most common starfish species of the North Sea, we found two distinct and possibly depth-related clades, probably sibling species, differing by 11.1–11.9 % sequence divergences (p-distances). For two starfish individuals, morphologically identified as Henricia sanguinolenta, the COI analysis revealed two monophyletic clades, of which one was classified as H. cf. oculata by comparison to published sequences.
Conclusions
This newly established sequence reference library for the North Sea Echinodermata allows and ensures future molecular species identification for various life-cycle stages including juveniles and meroplanktonic larvae and provides sequences for phylogeographic studies and the detection of sibling as well as cryptic species.
Similar content being viewed by others


Background
Echinoderms are a widespread marine invertebrate group, which can be found in a variety of different habitats ranging from intertidal zones to the depths of the world’s oceans [e.g. 1, 2]. In many regions, echinoderms form important physical and biological components in structuring marine benthic ecosystems. In the North Sea, they are one of the dominant groups characterising faunal communities [3] which dominate in abundance and biomass, represent major links in local food chains and thereby account for substantial transitions and fluxes of organic matter [4, 5]. The latter is particularly true for planktonic larval stages which seasonally occur in high and increasing abundances in the water column [6–8].
The echinoderm fauna of the North Sea has been well described based on their morphology with reports dating back to the late 19th and early 20th century [e.g. 9–11]. Mortensen [12] and Ursin [13] provided admirable synopses of the echinoderm fauna from the British Isles and the Central North Sea, respectively, and a number of extensive benthic monitoring surveys were conducted providing information on echinoderm species compositions in different parts of the North Sea [e.g. 14–20]. Long-term ecosystem surveys such as the German Small-scale Bottom Trawl Survey (GSBTS) [21] are the basis for studying long-term variability of echinoderm populations in the North Sea. Following studies revealed, for example, that r-selected opportunistic ophiuroids play a key role in the succession of benthic communities in the German Bight after extreme events such as cold winters [22, 23]. All these ecological ecosystem surveys, however, heavily rely on the accurate identification of species for a profound understanding of ecosystem functioning.
Taxonomic uncertainties in the morphological species identification of echinoderms and taxonomic revisions in individual groups are known for all classes [e.g. 24–27]. Such uncertainties can originate from unclear descriptions where—depending on the author—the same morphologically relevant structures may appear under different names. In this way, Clark [28], for example, traced back in detail the various names of the ‘oral tentacle scale’ (terminology proposed by her) in the brittle star’s family of Amphiuridae. Moreover, the morphological species identification of juvenile stages [e.g. 29] and pre-metamorphosed larval stages may prove far more difficult and identification literature is scarce [30]. Although it was reported that post-larvae of some brittle star species can be distinguished at a very small sizes [31], this still is a challenging task. Especially the phenomenon of adaptive plasticity during different developmental stages may cause taxonomic confusion in nominal species assignments. For example, in the echinoid’s and ophiuroid’s pluteus larva, the shape and length of the skeletal rods are, among others, relevant morphological identification features. However, various studies have revealed clear differences in the growth rates of the skeletal rods when reared under different food conditions [32] and references therein]. This may, therefore, also apply to varying environmental and food conditions in the field. Also, the identification of cryptic and sibling species is not possible without using advanced molecular techniques in addition to the traditional morphological species identification.
The uncertainties in morphological species identification and discrimination have spawned and advanced the application of molecular genetic identification techniques, for example DNA barcoding [33–38]. Molecular genetic approaches have led to far more detailed and/or contrasting results on cryptic and sibling species when compared to the morphological species identification alone. Spooner and Roy [39], for example, identified different lineages (‘hidden diversity’) in the ophiuroid Amphipholis squamata, originally thought to be a single species. This is also known for other taxonomic groups such as cnidarians and copepods from the North Sea. Here, recent studies detected sibling species by using molecular genetic discrimination techniques and by comparing the results with morphological species identifications [e.g. 40, 41]. Furthermore, morphologically distinct species showed high genetic similarity and thus give insights in speciation processes. For example, according to genetic similarity, Baric and Sturmbauer [42] suggested the endemic Mediterranean brittle star Ophiothrix quinquemaculata and the boreo-lusitanian O. fragilis to be ecotypes of the same species rather than actually different species. Also, early life-history stages, such as planktonic larvae, can be identified to species level based on their DNA sequence data. This approach can help to analyse planktonic species, difficult to be distinguished from one another, and to create long-term data sets of great ecological importance. Through applying such approaches, it was, for example, possible to identify Echinocardium cordatum larvae to be responsible for pronounced changes in the North Sea meroplankton due to increasing abundance and spatial distribution, which was viewed as a positive response to the increasing sea surface temperature in the North Sea [7, 8, 43]. Most of the comprehensive molecular genetic biodiversity studies on echinoderms were conducted off the coasts of southern and western Australia and New Zealand [44], in the coastal waters of Canada [45] and in the Arctic [46]. Ophiuroidea are studied in Icelandic waters [47]. In most of these biodiversity studies, mitochondrial DNA, mainly a fragment of the mitochondrial cytochrome c oxidase subunit I (COI), was analysed. This fragment is widely used for the discrimination of metazoan species [48, 49]. The published studies on echinoderms underline the applicability of this marker for successful (and economically important) echinoderm species discrimination or the discovery of cryptic species complexes [42, 50, 51].
The fundamental and essential step to allow and ensure future molecular species identification is the establishment of a valid reference library comprising data on both, morphological and molecular species information, together with various metadata (i.e. sampling location, identifier, etc.). Especially in times of developing and applying solely molecular-based biodiversity studies, using techniques like high-throughput DNA sequencing (i.e. metabarcoding [e.g. 52–54]), reference libraries connecting different species identification methods, are desperately needed. Once validated and established, they allow the identification of all different life-cycle stages, tissues or body parts.
To our knowledge, so far, no comprehensive molecular investigations have been conducted for the echinoderm fauna of the North Sea. To ensure the future molecular species identification for all different life-cycle stages, the prepending aim of the present study was to match morphology-based species assignments of adult North Sea echinoderms with the molecular species assignments using COI DNA sequences, and with this provide an accurate reference library as baseline for further studies.
Methods
Sampling and sample preparations
In the present study, a total of 317 adult echinoderm individuals were sampled at different stations in the North Sea (Fig. 1; Table 1). Sampling of individuals was conducted either using beam trawls or van Veen grabs from board the RV ‘Walther Herwig III’ during the International Bottom Trawl Survey (IBTS) and the German Small-scale Bottom Trawl Survey (GSBTS) in July and August 2007 (WH302), 2010 (WH335) and 2011 (WH345) as well as in January 2011 (WH340) and in March 2012 (WH352) (see [21] for further details). Additional van Veen grabs and beam trawl samples were taken from board of RV ‘Senckenberg’ in July and August 2010 and in November 2011. Specimens were also collected from Helgoland waters using van Veen grabs (RV ‘Uthörn’) and from the intertidal areas of the island of Helgoland in October 2011 as well as from the Gullmarsfjord in Sweden in August 2011. Subsequent to sampling, all individuals were—if possible—identified to species level using the identification guides by Mortensen [12], Paterson [55], Hayward and Ryland [56] and Southward and Campbell [57], and were then fixed either in absolute ethanol or were deep-frozen at −20 °C. In the home laboratory, digital images were taken from all investigated individuals and the deep-frozen samples were transferred into absolute ethanol. Depending on the taxonomic group, different body parts were dissected and used for the DNA extraction: parts of the outer legs (Ophiuroidea), tube feet (Asteroidea) and gonads and/or muscular tissue (Echinoidea, Holothuroidea). These tissue samples were stored in absolute ethanol at −20 °C. The individual specimens sampled for DNA extraction were stored in absolute ethanol as voucher material for potential re-identification. Voucher organisms, tissue and DNA extracts are stored at the German Center for Marine Biodiversity Research (DZMB), Senckenberg am Meer in Wilhelmshaven, Germany.

Locations of the 31 sampling stations in the North Sea (see Table 1 for detailed position data). The map was created with the statistical software R, version 3.2.3 [58] using bathymetrical data from http://topex.ucsd.edu [59]
Molecular genetic analyses
Genomic DNA was extracted from 317 echinoderm individuals, using the Qiagen DNeasy tissue kit with overnight lysis, following the manufacturer´s protocol. Amplification and sequencing of COI was partly conducted by the Canadian Center for DNA Barcoding (BIO, Guelph, Canada) and partly at the laboratories of the DZMB (amplification) and Macrogen (Amsterdam, Netherlands) (sequencing). At the DZMB, a COI fragment with the length of approximately 660 base pairs (bp) was amplified using the primer pair HCO2198 [60] and LCOech1aF1 [45]. PCRs started with denaturation at 95 °C (5 min) and were followed by 95 °C (30 s), 42 °C (60 s) and 72 °C (60 s) for 40 or in some cases 65 cycles. Final elongation was at 72 °C (7 min). The PCR products were checked on an agarose gel (1 %) with GelRed (0.1 %). Purification was conducted using the QIAquick PCR Purification Kit (Qiagen) or by incubating 10 µl of the PCR product in 0.5 µl Exonuclease I (20 U/µl) and 2 µl Alkalin Phosphatase (1 U/µl) for 15 min at 37 °C followed by 20 min at 75 °C. Sequencing was carried out by Macrogen (Amsterdam, Netherlands) using the primer pair HCO2198/LCOech1aF1 mentioned above.
Sequences were assembled, edited and checked for reading frames based on the mitochondrial genetic code for echinoderms using the software Geneious version 5.4.5 created by Biomatters [61]. Alignments were created using the software MUSCLE [62]. Pairwise genetic distances based on p-distances were calculated with the software MEGA (version 5.05 [63]) and gaps and/or missing data were treated as pairwise deletions. For the comparison to literature data, additionally Kimura-2-Parameter (K2P [64]) distances were calculated. The COI alignment comprised 317 sequences of a fragment length of 658 bp with 205 identical sites (31.2 %) and a pairwise identity of 77.2 %.
Neighbour Joining (NJ) analysis [65] based on p-distances with 1000 non-parametric bootstrap replicates [66] was performed using the software MEGA (version 5.05 [63]), again with gaps and/or missing data treated as pairwise deletions. Additionally, a maximum likelihood analysis was conducted using the GTRGAMMA model as recommended for data sets <50 taxa and with the generation of 1000 bootstrap replicates using the software RAxML-VI-HP [67]. P-distance matrices were also applied for non-metric multi-dimensional scaling (MDS) using the software Primer6 (version 6.1.6 [68]). For the three major groups of Ophiuroidea, Asteroidea and Echinoidea, MDS plots were performed with number of starts at 25 and minimum stress of 0.1.
Our data set was further analysed using the ‘BIN Discordance Report’ analysis tool available on Barcode of Life Data Systems (BOLD) without any filters [69]. One BIN represents a cluster of sequences differing by no more than 2 % [69]. Since it has been shown that BINs are highly congruent with existing species assignments [69], this analysis was used to (a) confirm North Sea echinoderm species clusters, (b) compare our data to published ones, (c) identify cryptic diversity, and (d) identify taxonomic inconsistencies. The taxonomically discordant BINs were reviewed for their actual cause and classified as follows: Identification Error (IE), Taxonomic problems (T), and Designation (D). Furthermore, each species was compared to available data in BOLD using the ‘Identification request’ tool (Species Level Barcode Records data base). These analyses were conducted on November 13th 2015.
Results
The 317 North Sea individuals of echinoderms identified by morphological and molecular genetic methods, comprised representatives of the four classes Asteroidea, Ophiuroidea, Echinoidea and Holothuroidea. The analysis based on morphological characters revealed 32 different taxa. It was possible to determine most specimens to species level, assigning 31 different species in total. However, one holothurian individual was identified to genus level only. In contrast to this, COI clustering as well as phylogenetic analyses (Neighbour Joining and Maximum Likelihood analyses) revealed 34 monophyletic clusters with two asteroid species, split into two species clusters each (Fig. 2).

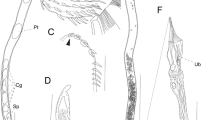
DNA sequence analyses: a Neighbour Joining analysis of the 658 bp COI fragment based on p-distances with 1000 non-parametric boostrap replicates. 317 individuals; b Maximum likelihood analysis of the 658 bp COI fragment based on the GTRGamma substitution model with 1000 bootstrap replicates. Numbers on branches represent bootstrap values ≥50 %. Drawings of echinoderms are taken and modified from Hayward and Ryland [56]. A Asteroidea, E Echinoidea, H Holothuroidea
In total, for 94.12 % of the species sampled and identified in this study, the morphological discrimination and species delimitation was concordant with the monophyletic clusters from the COI analyses. Taking into account the additional monophyletic clusters regarded as potentially different species clusters, inter-specific pairwise distances ranged from 8.37 to 34.64 % with the lowest value for the divergence for the two starfish species Solaster endeca and Crossaster papposus (Table 2; Fig. 3). Intra-specific variability ranged from 0.00 to 3.44 %, with highest values >2 % within the ophiuroids Amphiura filiformis (3.44 %), Ophiura sarsii (2.68 %) and Amphiura chiajei (2.43 %) (Table 2; Fig. 3). The difference between the intra- and the inter-specific variability revealed a minimum barcoding gap of 4.93 %.

Pairwise genetic p-distances of the North Sea Echinodermata (317 individuals; dark grey: intra-specific variability; light grey: inter-specific variability)
In the Neighbour Joining analysis, all 34 monophyletic lineages were supported by high bootstrap values (99 %) (Fig. 2a). On a higher taxonomic level, bootstrap support was found for clades of congeneric species like two putative Astropecten species (99 %), two Henricia species (99 %), and two Echinocardium species (98 %). Among the asteroids, S. endeca and C. papposus were supported by 99 %. A clade comprising the echinoids Strongylocentrotus droebachiensis, Psammechinus miliaris and Gracilechinus acutus was supported by 93 % with the latter two species showing a bootstrap support of 98 %. Another echinoid cluster was supported by 93 % comprising Brissopsis lyrifera, S. purpureus, E. cordatum and E. flavescens. All ophiuroid species clustered together with a bootstrap support of 94 % (Fig. 2a). The Maximum Likelihood Analysis revealed the same 34 monophyletic clusters as the Neighbour Joining analysis (Fig. 2b). Differences were detected only in a few lower bootstrap values, where the lowest value was ≤90 % recorded for O. ophiura (88 %) (Fig. 2b). In this analysis, there is a high divergence between Leptosynapta sp. and the other taxa. The grouping of the respective individuals to one species and the differences between the species is further demonstrated in the MDS plot (Fig. 4).

Multidimensional scaling (MDS) on the basis of the p-distance matrix for the Ophiuroidea, Asteroidea and Echinoidea from the present study
In the Asteroidea, we morphologically identified eleven species from four orders, nine families and eleven genera, i.e. Asterias rubens, Leptasterias muelleri, Stichastrella rosea, Astropecten irregularis, Luidia sarsii, Henricia sanguinolenta, Anseropoda placenta, Hippasterias phrygiana, Porania (Porania) pulvillus, Crossaster papposus, and Solaster endeca (Table 1). In contrast to these 11 morphological species assignments, the COI analyses revealed 13 different species clusters (Fig. 2). This was on account of each two clusters found for the species A. irregularis and for H. sanguinolenta, respectively. The results for A. irregularis revealed an intra-specific variability of 0.2–0.9 % in cluster 1 and 0–1.1 % in cluster 2. Distances between both clusters ranged from 11.1 to 11.9 %. Interestingly, the specimens from cluster 1 were sampled from depths between 87 and 111 m while individuals from cluster 2 were sampled from somewhat shallower depths (between 39 and 87 m) (see Table 1). At one sampling station (station 16 at 87 m depth; Fig. 1), each two specimens from both clusters occurred. The two specimens identified as H. sanguinolenta and originating from the same sampling station split into two clusters with a divergence of 13.07 %.
For all eight identified echinoid species, the morphological identifications and the molecular genetic discrimination, i.e. the species clusters based on COI divergences, were in accordance. The Echinoidea were represented by seven species from both the Irregularia and the Carinacea, providing three orders, seven families and six genera, i.e. Gracilechinus acutus, Psammechinus miliaris, Echinocyamus pusillus, Strongylocentrotus droebachiensis, Brissopsis lyrifera, Echinocardium cordatum, Echinocardium flavescens and Spatangus purpureus (Table 1).
From the Ophiuroidea, we identified eleven morphologically different species from two orders, five families and eight genera: Amphiura filiformis, Amphiura chiajei, Amphipholis squamata, Ophiothrix fragilis, Ophiura albida, Ophiura ophiura, Ophiura sarsii, Ophiopholis aculeata, Ophiactis balli, Ophiocten affinis, and Asteroynx loveni (Tabe 1). The latter species was found at the most northerly located station only (station 9; Fig. 1). Morphological and molecular discrimination was congruent in all identified species (Fig. 2).
The Holothuroidea were represented by two species from two orders and two families, which were both congruent in morphological and molecular genetic discrimination: Thyone fusus and Leptosynapta sp. identified only to genus level and represented by a single cluster.
The BIN analysis of our data set with 317 individuals revealed 316 records with BINs, thereby representing 35 BINs of which 18 were taxonomically concordant (147 records) and 17 which were taxonomically discordant (169 records) (Table 3). Highest rank of conflict for the taxonomically discordant BINs were mainly on species level (8 cases), followed by genus (5 cases), family (3 cases) and order level (1 case). For the ophiuroid O. sarsii, both one taxonomically concordant and one taxonomically discordant BIN was found. Except for two species (the ophiuroid A. squamata and the asteroid A. placenta), all other species from our data set clustered with those available in BOLD (partly published on higher taxonomic levels). Seven of the 17 taxonomically discordant BINs were renamed as taxonomically concordant due to designation problems (2 cases) and identification errors (5 cases). Because many species were found in the same BIN, the remaining 10 taxonomically discordant BINs were classified as taxonomic problems (T).
Particularly for the species split into two clusters, the application of the ‘Identification request’ tool allowed for their potential identification and designation. For both A. irregularis clusters we found high similarities (≤1 % divergence) to specimens unpublished in the BOLD data base [i.e. cluster 1 with one specimen sampled in Sweden; cluster 2 with specimens sampled in Sweden (1 specimen), Norway (1 specimen) and the Netherlands (2 specimens)]. Unfortunately, the sequences did not match with those from the comprehensive phylogeographic study on the genus Astropecten conducted by Zullinger and Lessios [50]—the only available published sequence data for this species in GenBank. The two individuals morphologically identified as H. sanguinolenta and originating from the same sampling station were referred to as Henricia sp. 1 and Henricia sp. 2. Henricia sp. 1 showed close similarity with Henricia cf. oculata from New Brunswick, St. Andrews, Canada (GenBank Accession numbers: HM400337, GU670162; 99.45 % pairwise identity), while Henricia sp. 2 did not reveal a close match (>98 %) with other Henricia species available in GenBank. Among the other identified asteroids in this study, we found close sequence matches with specimens from other regions (i.e. A. rubens 99.8 % similar to specimens from St. Andrews, New Brunswick, Canada and the United Kingdom (HM5420985, HM542098); C. papposus 99.7 % similar to specimens from Nunavut and New Brunswick, Canada (HM473811, HM543002, HM543003, HM542126-29), H. phrygiana 100 % similar to specimens from the Atlantic Ocean, Northwest of the United Kingdom (JQ896334, JQ896337-46, 48–50), S. endeca < 99.39 % similar to specimens from Nunavut and New Brunswick, Canada). Among the Echinoidea, the species were similar to those sampled from the same and to those from other regions (i.e. E. cordatum 99.39 % similar to those from Helgoland, Germany (NC_013881, FN562581), S. droebachiensis <99.39 % similar to those from Norway, Canada). Among the Ophiuroidea, high similarity was found for O. acuelata to specimens from Icelandic waters (>99.83 %, KJ620596–KJ620605). The North Sea specimens of other ophiuroid species, such as O. sarsii and the deep-sea species A. loveni showed large differences to those from other regions (i.e. O. sarsii 97.85 % similar to specimens from Nunavut, Canada (HM543041) and A. loveni <97.35 % similar to those from New Brunswick, Canada and Japan (HM542910-15, AB758757)). The holothuroid T. fusus showed a close sequence match (>98.92 %) with Thyonidium drummondii specimens from New Brunswick, Canada (HM400329, HM400330, HM400362, HM400363).
Discussion
Identification of North Sea echinoderms
Of the 94 echinoderm species described for the North Sea, the English Channel and the Irish Sea [57], we investigated more than 1/3 of the species. Among them, the Asteroidea showed the highest species number (13), followed by the Ophiuroidea (11), Echinoidea (8) and Holothuroidea (2). The same order in species richness by classes was observed for the eastern English Channel, Bristol Channel and Irish Sea [5], with the only difference, that in our study more ophiuroid and echinoid species were found.
Our study demonstrates concordance of morphological species identification with the results of the genetic analyses for about 94 % of the investigated North Sea echinoderm species. This result underlines the reliable species delimitation based on morphological diagnostic features presently applied for most of the species. However, the COI divergences effectively revealed deep lineage splits within various nominal species of starfish, such as A. irregularis, and also differentiated two morphologically very similar Henricia species, each treated and monitored as a single species so far. Both, the effectiveness of this approach in discriminating echinoderm species, and revealing cryptic lineages were already demonstrated by Ward and co-authors [44] for representatives of all five classes of echinoderms with 191 primarily Australian species, and by Corstorphine [45] for 131 mainly coastal Canadian species. The intra-specific divergences of the species in this study (p distances 0.00–3.44; Kimura-2-Parameter (K2P) distance for comparison: 0.00–3.55 %) are similar to those analysed by Ward and co-authors [44] (0.00–3.04 % K2P) and by Corstorphine [45] (<2 % K2P). Similar to the Australian findings [44], our results revealed the highest intra-specific variability in ophiuroid echinoderms. This variability within the North Sea brittle star species is similarly high as that revealed from the COI analysis from Icelandic waters (11 species, 66 specimens [47]).
In general, BIN analysis confirmed the morphologically and molecularly identified species and highlighted those with higher intra-specific variation or identification challenges. Here, it is important to note that one BIN is not equivalent to a species but represents a cluster of sequences differing by not more than 2 % [see 69]. The comparison of the present data set with published sequences and BIN analyses revealed 18 taxonomically concordant and 17 taxonomically discordant BINs. After review, 25 species were concordant and 10 discordant (Table 3). Reasons for classifying the discordant as concordant BINs were (1) designation problems and (2) presumably identification errors. The designation problems were caused by using the species names Astropecten irregularis cluster 1 and A. irregularis cluster 2 in order to distinguish the two species clusters. The presumable identification errors were assumed to occur, when only one specimen had another species name, i.e. in published data 60 specimens of A. rubens were classified as A. rubens and one as Leptasterias danica. The same was true for H. phrygiana, P. miliaris and O. ophiura. In the remaining ten taxonomically discordant BINs, more than one species name was found for more than one specimen in the published data. For example, for O. affinis, 16 individuals were identified as such, while two were identified as Ophiura robusta, one as Ophiactis balli and one as Ophiocten sericeum. Presumably, this can also be assigned to identification errors.
Hidden diversity
In the southern and shallower part of the North Sea, the starfish Asterias rubens is frequently found and yields both high biomasses and abundances [20, 70]. Further north in the central and northern North Sea, Astropecten irregularis is the dominating starfish species [3, 20]. Interestingly, A. irregularis species splits into two clades with a sequence divergence generally typical for inter-specific differences (11.1–11.9 %). This result may indicate the occurrence of two different species. The very species-rich genus Astropecten is supposed to be paraphyletic [50] and thereby comprises different species complexes and, most likely, cryptic species, which were already discovered when these authors had analysed 40 of the 150 species. For example, Astropecten specimens from Portuguese waters differed strongly from those sampled off the coasts of Sardinia, Greece and Madeira with uncorrected COI distances of 6.3–8.9 % (0.063–0.089 [50]). In this study we can also demonstrate two potentially different Astropecten lineages for the North Sea. Except for one station in the central North Sea with a depth of 87 m (station 16; Fig. 1), where both A. irregularis lineages overlapped, A. irregularis cluster 1 showed a more northerly distribution (in 100–111 m depth) and A. irregularis cluster 2 revealed a more southerly one (in 31–73 m depth). Interestingly, it is reported that A. irregularis has a very wide distributional range in the North Sea stretching from shallow to deep waters. For example, community clusters between the 100 and 200 m depth line are characterized by A. irregularis while at 50–100 m both Asterias rubens and Astropecten irregularis were, among others, the dominant species [3]. Our results might indicate a possible species splitting in relation to depth. Due to the great variation in the development of spines on the marginal plates, different varieties have been attributed to A. irregularis from the North Sea. Of the three forms defined as A. irregularis var. serratus (Müller and Troschel 1842), var. typicus and var. pentacanthus (Delle Chiaje 1827), Ursin [13] referred 62 % of the investigated species sampled in comprehensive surveys in the 1950s to var. typicus and 38 % to var. pentacanthus. In comparison with these data, A. irregularis cluster 1 with its deep and more northerly distribution would correspond to var. penthacanthus and A. irregularis cluster 2 with its occurrence at moderate depths in the more southern and eastern part of the North Sea would correspond to var. typicus. However, Ursin [13] reported the three forms rather to be modifications without systematic value and later, also Zulliger and Lessios [50] generally stated that in the genus Astropecten, the morphological diversity is very high and that characters are often continuous rather than discrete. In general, several subspecies (see references in [50]) and high genetic variability are described for A. irregularis distributed in European waters with the suggestion of three different species [50]. To elucidate the uncertainty of the depth-related splitting in the North Sea of A. irregularis specimens, detailed analyses of the morphological and ecological characters as well as of additional molecular makers are needed. Unfortunately, a combined analysis of the A. irregularis COI data from this study together with the published ones by Zulliger and Lessios [50] was not possible because the data sets did not result in a combined alignment without any gaps (even when no stop codons were found for both data sets). Since COI is a coding gene, the alignment and the identification of the reasons for this problem should be straightforward but still we cannot specify the reason for the alignment problem properly. The fact that A. irregularis sequences from this study show high similarities (<1 % divergence) to unpublished data in BOLD and that sequences from all other species analyzed in this study were comparable to published ones, demonstrates the amplification success of the COI metazoan barcoding region in this study. Maybe the fact that the two studies used very different primers/primer pairs resulted in the amplification of different COI target regions which are now not comparable? For example, Zulliger and Lessios [50] used different combinations of in total seven primers, both in the general and in the cycle-sequencing PCR. Comparing their A. irregularis sequences within the BOLD data base did not result in any close match.
Also, for the starfish Henricia, the COI indicated the existence of two different species. The difficulty in identifying the individuals morphologically to species level in the Henricia (Gray, 1840) complex has been noted by many authors [i.e. 12, 24, 70–73], who also differ in their descriptions of the species and of the genus. Fisher [73] suggested that the species within the Henricia genera hybridize, and that his species’ classification, therefore, had to be considered temporary. Madsen [74] divided the Henricia complex morphologically into two main groups: the H. pertusa group and the H. perforata group, based mainly on differences in the structure of the dorsal spines. Reproductive isolation of both Henricia groups was found by biochemical analyses [75]. Based on the work of Madsen [74], Southward and Campbell [57] listed H. perforata (O.F. Müller 1776), H. oculata (Pennant 1777), H. sanguinolenta (O.F. Müller 1776) and H. pertusa (O.F. Müller, 1776) in the ‘Synopses of the British Fauna’. High similarity with published sequences underlined that for at least one individual, we are possibly dealing with H. cf. oculata but for the other individual, the identification remains uncertain. Interestingly, for H. cf. oculata and H. sanguinolenta, Corstorphine [45] had similar morphological identification challenges/problems and tried to use sequence data (COI and 16S) in order to assign species names.
Apart from depth-related splitting within species, high intra-specific variability was found in the brittle stars. Eight of the eleven species are typical representatives for our study area, with four species regarded as ‘common’ species (i.e. A. filiformis, O. fragilis, O. albida and O. ophiura) and another four species regarded as ‘rare’ species in the German Bight (i.e. A. chiajei, O. aculeata, O. affinis and O. sarsii) [76]. The latter four are described to be found more often in northern regions of the North Sea [76]. Among these rare species, the high variability within O. sarsii—irrespective of geographic distribution—resulted in the classification of two different BINs: (1) one specimen in a taxonomically concordant BIN together with other 16 specimens from Canada and Norway identified as the same species, and (2) five specimens in a discordant BIN together with four other O. sarsii and two specimens identified as Ophiura robusta from Norway and Sweden. In general, species with high intra-specific variability resulting in more than one BIN, can give hints on cryptic diversity. Hereby, the fact that species from the same region reveal such high intra-specific variability, is a very interesting fact. For other ophiuroids, the high divergences may result from specimens originating from different sampling regions and thus from geographically large distances. For example, for the deep-sea dwelling A. loveni, the North Sea individuals showed high divergences to those from Canada, resulting in two different BINs. A phylogenetic pattern and possible indication for cryptic species was found for O. fragilis with a high level of haplotype diversity and COI divergences of up to 18.6 % between North Atlantic and Mediterranean/Galician coast populations [77]. Several very divergent mitochondrial and nuclear lineages were also found for the cosmopolitan brittle star A. squamata [78, 79]. In contrast to O. fragilis, divergences were high in co-existing individuals, thus sibling species were concluded to occur in the Mediterranean populations [79]. Hence, when thinking of their cosmopolitan distribution, many more cryptic species can be expected [79]. Distant A. squamata populations showed morphological similarities, while large genetic divergences were found, both, within and between populations when analysing mitochondrial DNA (i.e. 16S rDNA [78]). In contrast to this, O. fragilis has different morphological forms but does not markedly diverge on the COI basis [77].
Usage and applicability of the sequence data
Molecular approaches are useful when morphological identification is challenging or impossible, for example when diagnostic characters are subtle or missing in adults or are still undeveloped in early developmental stages. Here, this reference library can now find various applications in the future species identification for North Sea echinoderms. Especially the sequence data for the starfish genera Astropecten and Henricia allow for the future identification of these different clusters and can help and support further analyses on their morphology and ecology. For meroplanktic larvae and juveniles, the reference library allows for their species identification and for the evaluation of morphological diagnostic characters of the different developmental stages. This can then help to analyse plankton data, especially long-term data, and is of great ecological importance in the context of recruitment, succession patterns or monitoring. Another applicability of this reference library is the contribution to regional sequence data for species with a wide distribution range for phylogeographic analyses, as for example in A. squamata, S. droebachiensis, O. fragilis, A. rubens and H. phrygiana. For many of these species, haplotype diversity and population dynamics, both, on mitochondrial and nuclear markers, were studied to emphasize phylogeographic patterns. Finally, the analysis of molecular markers may help to detect cryptic diversity or to revise groups with taxonomic uncertainties and, therefore, represents a strong driving force for taxonomy in general.
Abbreviations
- COI:
-
mitochondrial cytochrome c oxidase subunit I
- D:
-
Designation
- GSBTS:
-
German Small-scale Bottom Trawl Survey
- IBTS:
-
International Bottom Trawl Survey
- IE:
-
identification error
- K2P:
-
Kimura-2-parameter distance
- MDS:
-
multi-dimensional scaling
- NJ:
-
neighbour joining
- T:
-
taxonomic problems
References
Hyman LH. The invertebrates: echinodermata. New York: McGraw-Hill Book Company Inc; 1955.
Uthicke S, Schaffelke B, Byrne M. A boom-bust phylum? Ecological and evolutionary consequences of density variation in echinoderms. Ecol Monogr. 2009;79:3–24. doi:10.1890/07-2136.1.
Callaway R, Alsvåg J, de Boois I, Cotter J, Ford A, Hinz H, Jennings S, Kröncke I, Lancaster J, Piet G, Prince P, Ehrich S. Diversity and community structure of epibenthic invertebrates and fish in the North Sea. ICES J Mar Sci. 2002;59:1199–214. doi:10.1006/jmsc.2002.1288.
Dahm C. Growth, production and ecological significance of Ophiura albida and O. ophiura (Echinodermata: Ophiuroidea) in the German Bight. Mar Biol. 1993;116:431–7. doi:10.1007/BF00350060.
Ellis JR, Rogers SI. The distribution, relative abundance and diversity of echinoderms in the eastern English Channel, Bristol Channel, and Irish Sea. J Mar Biol Assoc UK. 2000;80:127–38. doi:10.1017/S0025315499001642.
Lindley JA, Gamble JC, Hunt HG. A change in the zooplankton of the central North Sea (55° to 58° N): a possible consequence of changes in the benthos. Mar Ecol Prog Ser. 1995;119:299–303. doi:10.3354/meps119299.
Kirby RR, Beaugrand G, Lindley JA, Richardson AJ, Edwards M, Reid PC. Climate effects and benthic-pelagic coupling in the North Sea. Mar Ecol Prog Ser. 2007;330:31–8. doi:10.3354/meps330031.
Kirby RR, Beaugrand G, Lindley JA. Climate-induced effects on the meroplankton and the benthic-pelagic ecology of the North Sea. Limnol Oceanogr. 2008;53:1805–15. doi:10.4319/lo.2008.53.5.1805.
Möbius K, Bütschli O. Echinodermata. Jahresbericht der Commission zur wissenschaftlichen Untersuchung der deutschen Meere in Kiel für die Jahre 1872/73. Zoologische Ergebnisse der Nordseefahrt. 1875;143–151.
Meissner M, Collin A. Beiträge zur Fauna der südöstlichen und östlichen Nordsee Teil II. Echinodermen. Helgoland Wiss Meer. 1894;1:329–46.
Süßbach S, Breckner A. Die Seeigel, Seesterne und Schlangensterne der Nord- und Ostsee. Wiss Meeresunters. 1911;12:169–300.
Mortensen T. Handbook of the echinoderms of the British Isles. London: Oxford University Press; 1927.
Ursin E. A quantitative investigation of the echinoderm fauna of the central North Sea. Meddelelser fra Kommissionen for Danmarks Fiskeri- og Havundersøgelser: Ny Serie. 1960;2:1–204.
Ziegelmeier E. Macrobenthos investigations in the eastern part of the German Bight from 1950 to 1974. Rapport et Procès-verbaux des Réunion. J Cons Int Explor Mer. 1974;1978(172):432–44.
Basford D, Eleftheriou A, Raffaelli D. The infauna and epifauna of the northern North Sea. Neth J Sea Res. 1990;25:165–73. doi:10.1016/0077-7579(90)90017-B.
Duineveld GCA, Künitzer A, Niermann U, de Wilde PAWJ, Gray JS. The macrobenthos of the North Sea. Neth J Sea Res. 1991;28:53–65. doi:10.1016/0077-7579(91)90004-K.
Heip C, Basford D, Craeymeersch JA, Dewarumez J-M, Dörjes J, de Wilde PAWJ, Duineveld GCA, Eleftheriou A, Herman P, Niemann U, Kingston P, Künitzer A, Rachor E, Rumohr H, Soetaert K, Soltwedel T. Trends in biomass, density and diversity of North Sea macrofauna. ICES J Mar Sci. 1992;49:13–22. doi:10.1093/icesjms/49.1.13.
Künitzer A, Basford D, Craeymeersch JA, Dewarumez JM, Dörjes J, Duineveld GCA, Eleftheriou A, Heip C, Herman P, Kingston P, Niemann U, Rachor E, Rumohr H, de Wilde PAWJ. The benthic infauna of the North Sea: species distribution and assemblages. ICES J Mar Sci. 1992;49:127–43. doi:10.1093/icesjms/49.2.127.
Schroeder A. Community dynamics and development of softbottom macrozoobenthos in the German Bight (North Sea) 1969–2000. Ber Polar Meeresforsch. 2005;494:1–181.
Neumann H, Ehrich S, Kröncke I. Spatial variability of epifaunal communities in the North Sea in relation to sampling effort. Helgol Mar Res. 2008;62:215–25. doi:10.1007/s10152-008-0109-8.
Ehrich S, Adlerstein S, Brockmann U, Floeter J, Garthe S, Hinz H, Kröncke I, Neumann H, Reiss H, Sell AF, Stein M, Stelzenmüller V, Stransky C, Temming A, Wegner G, Zauke GP. 20 years of the German Small-Scale Bottom Trawl Survey (GSBTS): a review. Senck Marit. 2007;37:13–82. doi:10.1007/BF03043206.
Neumann H, Ehrich S, Kröncke I. The effects of cold winters and climate on the temporal variability of an epibenthic community in the German Bight. Clim Res. 2008;37:241–51. doi:10.3354/cr00769.
Neumann H, Reiss H, Rakers S, Ehrich S, Kröncke I. Temporal variability of southern North Sea epifauna communities after the cold winter 1995/1996. ICES J Mar Sci. 2009;66:2233–43. doi:10.1093/icesjms/fsp203.
Clark AM, Downey ME. Starfishes of the Atlantic. Lindon: Chapman and Hall; 1992.
Littlewood DTJ, Smith AB. A combined morphological and molecular phylogeny for sea urchins (Echinoidea: Echinodermata). Philos Trans R Soc B Biol Sci. 1995;347:213–34. doi:10.1098/rstb.1995.0023.
Pearse VB, Pearse JS, Hendler G, Byrne M. An accessible population of Ophiocanops off NE Sulawesi, Indonesia. In: Mooi R, Telford M, editors. Echinoderms. Rotterdam: Balkema; 1998. p. 413–8.
Lacey KMJ, McCormack GP, Keegan BF, Powell R. Phylogenetic relationships within the class Holothuroidea, inferred from 18S rRNA gene data. Mar Biol. 2005;147:1149–54. doi:10.1007/s00227-005-0009-2.
Clark AM. Notes on the family Amphiuridae (Ophiuroidea). Bull Br Mus Nat Hist Zool. 1970;19:1–81. doi:10.5962/bhl.part.24085.
Stöhr S. Who’s who among baby brittle stars (Echinodermata: Ophiuroidea): postmetamorphic development of some North Atlantic forms. Zool J Linn Soc. 2005;143:543–76. doi:10.1111/j.1096-3642.2005.00155.x.
Smith DL, Johnson KB. A guide to marine coastal plankton and marine invertebrate larvae. 2nd ed. Dubuque: Kendall/Hunt Publication Company; 1996.
Webb CM, Tyler PA. Post-larval development of the common north-west European brittle stars Ophiura ophiura, O. albida and Acrocnida brachiata (Echinodermata: Ophiuroidea). Mar Biol. 1985;89:281–92. doi:10.1007/BF00393662.
Podolsky RD, McAlister JS. Developmental plasticity in Macrophiothrix brittlestars: are morphologically convergent larvae also convergently plastic? Biol Bull. 2005;209:127–38. doi:10.2307/3593130.
Arndt A, Marquez C, Lambert P, Smith MJ. Molecular phylogeny of eastern Pacific sea cucumbers (Echinodermata: Holothuroidea) based on mitochondrial DNA sequence. Mol Phylogenet Evol. 1996;6:425–37. doi:10.1006/mpev.1996.0091.
Hart MW, Byrne M, Smith MJ. Molecular phylogenetic analysis of life-history evolution in asterinid starfish. Evolution. 1997;51:1848–61. doi:10.2307/2411007.
Cohen BL, Améziane N, Eleaume M, Richer de Forges B. Crinoid phylogeny: a preliminary analysis (Echinodermata: Crinoidea). Mar Biol. 2004;144:605–17. doi:10.1007/s00227-003-1212-7.
Hart MW, Podolsky RD. Mitochondrial DNA phylogeny and rates of larval evolution in Macrophiothrix brittlestars. Mol Phylogenet Evol. 2005;34:438–47. doi:10.1016/j.ympev.2004.09.011.
Byrne M, Rowe F, Uthicke S. Molecular taxonomy, phylogeny and evolution in the family Stichopodidae (Aspidochirotida: Holothuroidea) based on COI and 16S mitochondrial DNA. Mol Phylogenet Evol. 2010;56:1068–81. doi:10.1016/j.ympev.2010.04.013.
Okanishi M, Fujita T. Molecular phylogeny based on increased number of species and genes revealed more robust family-level systematics of the order Euryalida (Echinodermata: Ophiuroidea). Mol Phylogenet Evol. 2013;69:566–80. doi:10.1016/j.ympev.2013.07.021.
Sponer R, Roy MS. Phylogeographic analysis of the brooding brittle star Amphipholis squamata (Echinodermata) along the coast of New Zealand reveals high cryptic genetic variation and cryptic dispersal potential. Evolution. 2002;56:1954–67. doi:10.1111/j.0014-3820.2002.tb00121.x.
Laakmann S, Gerdts G, Erler R, Knebelsberger T, Martínez Arbizu P, Raupach MJ. Comparison of molecular species identification for North Sea calanoid copepods (Crustacea) using proteome fingerprints and DNA sequences. Mol Ecol Res. 2013;13:862–76. doi:10.1111/1755-0998.12139.
Laakmann S, Holst S. Emphasizing the diversity of North Sea hydromedusae by combined morphological and molecular methods. J Plankton Res. 2014;36:64–76. doi:10.1093/plankt/fbt078.
Baric S, Sturmbauer C. Ecological parallelism and cryptic species in the genus Ophiothrix derived from mitochondrial DNA sequences. Mol Phylogenet Evol. 1999;11:157–62. doi:10.1006/mpev.1998.0551.
Kirby RR, Lindley JA. Molecular analysis of Continuous Plankton Recorder samples, an examination of echinoderm larvae in the North Sea. J Mar Biol Assoc UK. 2005;85:451–9. doi:10.1017/S0025315405011392.
Ward RD, Holmes BH. O´Hara TD. DNA barcoding discriminates echinoderm species. Mol. Ecol Res. 2008;8:1202–11. doi:10.1111/j.1755-0998.2008.02332.x.
Corstorphine EA. DNA barcoding of echinoderms: species diversity and patterns of molecular evolution. Master Thesis, University of Guelph, Ontario, Canada; 2010.
Hardy SM, Carr CM, Hardman M, Steinke D, Corstorphine E, Mah C. Biodiversity and phylogeography of Arctic marine fauna: insights from molecular tools. Mar Biodiv. 2011;41:195–210. doi:10.1007/s12526-010-0056-x.
Khodami S, Martínez Arbizu P, Stöhr S, Laakmann S. Molecular species delimitation of Icelandic brittle stars (Ophiuroidea). Pol Polar Res. 2014;35:243–60. doi:10.2478/popore-2014-0011.
Hebert PDN, Ratnasingham S, deWaard JR. Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc Biol Sci. 2003;270(Suppl 1):96–9. doi:10.1098/rsbl.2003.0025.
Hebert PDN, Cywinska A, Ball SL, DeWaard JR. Biological identifications through DNA barcodes. Proc Biol Sci. 2003;270:313–21. doi:10.1098/rspb.2002.2218.
Zulliger DE, Lessios HA. Phylogenetic relationships in the genus Astropecten Gray (Paxillosida: Astropectinidae) on a global scale: molecular evidence for morphological convergence, species-complexes and possible cryptic speciation. Zootaxa. 2010;2504:1–19.
Uthicke S, Byrne M, Conand C. Genetic barcoding of commercial Bêche-de-mer species (Echinodermata: Holothuroidea). Mol Ecol Res. 2010;10:634–46. doi:10.1111/j.1755-0998.2009.02826.x.
Taberlet P, Coissac E, Pompanon F, Brochmann C, Willerslev E. Towards next-generation biodiversity assessment using DNA metabarcoding. Mol Ecol. 2012;21:2045–50. doi:10.1111/j.1365-294X.2012.05470.x.
Leray M, Knowlton N. DNA barcoding and metabarcoding of standardized samples reveal patterns of marine benthic diversity. PNAS. 2015;112:2076–81. doi:10.1073/pnas.1424997112.
Mohrbeck I, Raupach MJ, Martínez Arbizu P, Knebelsberger T, Laakmann S. High-throughput sequencing—The key to rapid biodiversity assessment of marine metazoa? PLoS One. 2015;10:e0140342. doi:10.1371/journal.pone.0140342.
Paterson GLJ. The deep-sea Ophiuroidea of the North Atlantic Ocean. Bull Br Mus Nat Hist. 1985;49:1–162.
Hayward PJ, Ryland JS. Marine fauna of the British Isles and North-West Europe. Oxford: Oxford University Press; 1990.
Southward EC, Campbell AC. Echinoderms. In: Crothers JH, Hayward PJ, editors. Synopses of the British fauna (New Series), No. 56. Shrewsbury: Field Studies Council; 2006. p. 272.
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2015. https://www.R-project.org/.
Smith WHF, Sandwell DT. Global seafloor topography from satellite altimetry and ship depth soundings. Science. 1997;277:1957–62. doi:10.1126/science.277.5334.1956.
Folmer OM, Black W, Hoen R, Lutz R, Vrijenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metozoan invertebrates. Mol Mar Biol Biotechnol. 1994;3:294–9.
Kearse M, Moir R, Wilson A, Stones-Havas S, Cheung M, Sturrock S, Buxton S, Cooper A, Markowith S, Duran C, Thierer T, Ashton B, Meintjes P, Drumond A. Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics. 2012;28:1647–9. doi:10.1093/bioinformatics/bts199.
Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32:1792–7. doi:10.1093/nar/gkh340.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28:2731–9. doi:10.1093/molbev/msr121.
Kimura M. A simple model for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol. 1980;16:111–20.
Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987;4:406–25.
Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39:783–91. doi:10.2307/2408678.
Stamatakis A. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006;22:2688–90. doi:10.1093/bioinformatics/btl446.
Clarke KR, Gorley RN. PRIMER v6: user manual/tutorial. Plymouth: PRIMER-E; 2006.
Ratnasingham S, Hebert PDN. A DNA-based registry for all animal species: the barcode index number (BIN) system. PLoS One. 2013;8:e66213. doi:10.1371/journal.pone.0066213.
Reiss H, Kröncke I. Seasonal variability of epibenthic communities in different areas of the southern North Sea. ICES J Mar Sci. 2004;61:882–905. doi:10.1016/j.icesjms.2004.06.020.
Bell FJ. Catalogue of the British echinoderms in the British Museum. London: British Museum; 1892.
Koehler R. Resultats scientifiques de la campagne du” Caudan” dans le golfe de Gascogne: Echinodermes, vol. 2. Amsterdam: Masson; 1896.
Fisher WK. Asteroidea of the North Pacific and adjacent waters. Part 1. Bull US Natl Mus. 1911;6:420. doi:10.5479/si.03629236.76.i.
Madsen FJ. The Henricia sanguinolenta complex (Echinodermata, Asteroidea) of the Norwegian Sea and adjacent waters. A re-evaluation, with notes on related species. Steenstrupia. 1987;13:201–68.
Ringvold H, Stien J. Biochemical differentiation of two groups within the species-complex Henricia Gray, 1840 (Echinodermata, Asteroidea) using starch-gel electrophoresis. Hydrobiologia. 2001;459:57–9. doi:10.1023/A:1012539821086.
Boos K, Franke H-D. Brittle stars (Echinodermata: Ophiuroidea) in the German Bight (North Sea)—species diversity during the past 130 years. J Mar Biol Assoc UK. 2006;86:1187–97. doi:10.1017/S0025315406014184.
Muths D, Jollivet D, Gentil F, Davoult D. Large-scale genetic patchiness among NE Atlantic populations of the brittle star Ophiothrix fragilis. Aquatic Biol. 2009;5:117–32. doi:10.3354/ab00138.
Sponer R, Deheyn D, Roy MS. Large genetic distances within a population of Amphipholis squamata (Echinodermata; Ophiuroidea) do not support colour varieties as sibling species. Mar Ecol Prog Ser. 2001;219:169–75. doi:10.3354/meps219169.
Boissin E, Feral JP, Chenuil A. Defining reproductively isolated units in a cryptic and syntopic species complex using mitochondrial and nuclear markers: the brooding brittle star, Amphipholis squamata (Ophiuroidea). Mol Ecol. 2008;17:1732–44. doi:10.1111/j.1365-294X.2007.03652.x.
Authors’ contributions
SL, TK and MJR designed the general approach of this study. All authors contributed to the sampling of the study material. Morphological identification was carried out by HN, KB and experts during GSBTS and IBTS. Molecular genetic and data analyses were performed by SL and the manuscript was written by SL, HN and KB. All authors read and approved the final manuscript.
Acknowledgements
We thank the captain and the crew of the RV ‘Walther Herwig III’, RV ´Uthörn´ and the RV ‘Senckenberg’ for their sampling support. Many thanks to the GSBTS/IBTS, especially Anne Sell and Matthias Kloppmann (Thuenen Institute of Sea Fisheries). Corinna Girschik, Rebekka Schüller and Valeska Borges assisted in the laboratory work.
Competing interests
The authors declare that they have no competing interests.
Availability of data and material
The datasets generated during the current study are available in the public data set “North Sea Echinodermata” (Dataset ID: DS-NSECH; dx.doi.org/10.5883/DS-NSECH) on the Barcode of Life Data Systems (BOLD; www.boldsystems.org). In addition, all barcode sequences were deposited on GenBank (Accession numbers KX458801 to KX459117; BankIt: 1931462).
Funding
This work was funded by the Federal Ministry of Education and Research (Grant No. 03F0499A) and the Land Niedersachsen. The present provider of funding had no role in the study design, data collection and analysis and the decision to publish or in the preparation of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Laakmann, S., Boos, K., Knebelsberger, T. et al. Species identification of echinoderms from the North Sea by combining morphology and molecular data. Helgol Mar Res 70, 18 (2017). https://doi.org/10.1186/s10152-016-0468-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s10152-016-0468-5